
Abstract
The projected northwards shift in the distribution suggests an expansion of European beech (Fagus sylvatica L.) into the Baltics and an increase of its commercial potential there. Furthermore, recent studies have shown good long-term survival of underplanted, yet non-rereleased beech under the presumedly limiting climate of the eastern part of Latvia, thus supporting the advance of the thermophilic species. Hence, the estimation of the responses of the underplanted beech to shelterwood removal is a logical step in the assessment of the specie’s growth potential. To analyse the effect of conventionally managed mature coniferous shelterwood thinning on the growth of underplanted European beech, six thinning treatments (100, 75, and 50% of canopy, second canopy storey, second canopy storey with understorey removed, and the control) were tested in the 33-year-old northeasternmost experimental plantation in Europe. Growth responses were evaluated five years after the treatment, when beech was 38 years old. The survival and growth responses to shelterwood thinning and local characteristics at the tree and stand level were estimated using linear models. The post-treatment survival/mortality was independent, yet the dimensions of beech, particularly stem diameter, were significantly affected by shelterwood thinning intensity (positively). Regarding tree height, the response was somewhat weaker compared to that of stem diameter, while being disproportional to thinning intensity, with moderate thinning having the strongest positive effect. The estimated positive responsiveness of beech to shelterwood thinning suggested increasing growth potential of the species and expansion in the Baltics. Though the responses of beech were moderate probably due to the age and harsher climate.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The distribution of European beech (Fagus sylvatica L.), which is a common tree species in Central and Western Europe (Durrant et al. 2016), is projected to shift northwards as a consequence of the anticipated climatic changes (Fortu 2023; Martinez del Castillo et al. 2022; Hickler et al. 2012; Allen et al. 2010; Kramer et al. 2010). Accordingly, the abundance of the species is projected to increase northwards of its current range, particularly in the lowland southern Sweden and the Baltics, where species can become economically and ecologically important (Fortu 2023; Jansone et al. 2023; Hickler et al. 2012). Furthermore, the disproportional contraction of the trailing part of the species range emphasizes the importance of the eastern Baltic region for the future beech forests (Martinez del Castillo et al. 2022; Kramer et al. 2010).
The extension of vegetation season is the principal driver of the northward expansion of beech (Fortu 2023; Hickler et al. 2012). Still, the species is susceptible to cold spells and, particularly, late frosts, especially at a young age (Weigel et al. 2018; Petritan et al. 2011; Tompson and Fennessy 2011; Bolte et al. 2007). Late frosts are considered the major threats to the commercial potential of beech under cool climates of the leading parts of the distribution (Petritan et al. 2011; Bolte et al. 2007). Accordingly, shelterwood is widely applied to protect young beech from frost and winter desiccation (Podrázský et al. 2019).
The means (intensity and design) of shelterwood removal and its effects on the sustainability of the stand and the advance growth (understorey/underplanted saplings), in particular, vary regionally and locally (Reventlow et al. 2023, 2018; Weidig and Wagner 2021; Diaconu et al. 2015; Boncina et al. 2007; Löf et al. 2005). Furthermore, the underlying effects of shelterwood removal are complex as changes in light, climate and microclimate interact (Čater et al. 2014; Övergaard et al. 2009; Lichtenthaler et al. 2007; Agestam et al. 2003). Shelter removal affects the stand moisture regime, to which beech is sensitive, and which might affect competitiveness (Leuschner 2020; Roibu et al. 2022; Kramer et al. 2010). Hence, a locally/regionally optimized sheltering removal strategy is critical for the regeneration, survival, browsing protection, and stem quality of the regenerating trees (Löf, 2000; Podrázský et al. 2019).
Regional differences in the effect of shelterwood are explicit for the survival of beech advance growth, with the positive effect increasing towards a cooler climate (Podrázský et al. 2019; Caquet et al. 2010; Kunstler et al. 2005). The dimensions of the advance growth trees generally show a positive response to decreasing shelterwood density (Weidig and Wagner 2021; Reventlow et al. 2018; Diaconu et al. 2015; Löf et al. 2007). However, excessively released trees can form large and thick branches that often develop into forks (Fortu 2023), thus resulting in slower height growth and poorer stem quality (Reventlow et al. 2018). Also, in the case of extreme shelter removal under cooler climates, height growth can even be decreased due to frost, winter desiccation, drought, or changes in crown shape (Reventlow et al. 2023; Agestam et al. 2003).
In the Baltics, Latvia currently appears as the Northeastern limit of beech growing in forest stands in Europe (Puriņa et al. 2016a; b; Jansons et al. 2015). Still, the preconditions for wider advance of the species are already demonstrated by growth, successful self-regenerations, and acclimation of beech in stands in the coastal western part of Latvia (Matisons et al. 2018, 2017; Puriņa et al. 2016a, b; Jansons et al. 2015) and Lithuania (Linkevičius and Junevičiūtė, 2021). In a young (12 years) plantation in the central part of Latvia, good survival and increment have been observed also for unsheltered saplings (Puriņa et al. 2016a). However, there is insufficient information about the responsiveness of beech to management (including sheltering) under such presumedly marginal conditions. Furthermore, in an experimental plantation in the eastern part of Latvia, where the climate is presumedly extreme, underplanted beech has shown good (~ 80%) long-term (33 years) survival under mixed coniferous canopy despite several cold spells (Puriņa et al. 2016b), thus indicating the efficiency of shelterwood. Hence in this site, which currently is the northeasternmost plantation of beech in Europe, the responses of beech to shelter removal appear indicative of growth potential, as well as of the optimal thinning intensity under extreme climate.
The study aimed to assess the growth response of underplanted European beech at the tree and stand levels concerning different shelterwood thinning intensities at the northeasternmost plantation in Europe. We hypothesized that more intense thinning facilitated diameter growth, but had a lesser effect on height growth of underplanted beech. We also hypothesized that the responses would be moderate, as the harsher climate would have burdened acclimation of beech to the rapid changes in light conditions.
Material and methods
Experimental site
The studied plantation was located in lowland conditions in the hemiboreal forest zone, in the central part of Latvia (56° 42′ N, 25° 56′ E). The regional climate in the area is moist continental; the mean monthly temperature ranges from − 4.5 °C in February being the coldest to 17.6 °C in July the warmest month (LEGMC 2020). During the last three decades, the six coldest six-hour mean temperature events were recorded in January 1987 (− 31.1 °C), 2003 (− 31.7 °C) and 2006 (− 28.7 °C), and in February 1985 (− 31.2 °C), 2007 (− 29.6 °C) and 2012 (− 28.9 °C). The late-spring frosts are frequently observed at the start of the growing season in May and up to the first days of June (LEGMC 2020). The mean annual sum of precipitation is 698.1 mm.
The study site was an unfenced sheltered experimental plantation of beech established in 1983, which to the authors’ knowledge is the northeasternmost forest plantation of the species in Europe (Purina et al. 2016b). The stand was growing on moderately dry, freely draining mesotrophic loamy soil with an acidic reaction (comparable to a site index of 28–30 m for conifers). The topography of the site was generally flat with a relative elevation of < 6 m. The plantlets of beech raised from the seed material from beech stands in the western part of Latvia (2–3 years old) were planted along the trailing lines (3 m width skidding, maintenance corridors; distance between them ranged from 3.5 to 8.5 m) with the spacing of 0.75–0.95 m. The shelterwood was a mature (ca. 90 years old) naturally regenerated and conventionally managed (two thinnings from the below with occasional deadwood/firewood collection) mixed coniferous stand dominated by mature Scots pine (Pinus sylvestris L.) or Norway spruce (Picea abies L. H. Karst.) with some silver birch (Betula pendula Roth.) admixture. The stand had a scarce second canopy storey (trees of 50–75% of the mean dominant canopy height) prevailed by spruce with a slight birch and oak (Quercus robur L.) admixture. The understory was scarce with hazel (Corylus avellana L.) being the most common. More details on the shelterwood and underplanted birch are given in Supplementary material (Table S1). Prior to shelterwood thinning, the planted beeches have had high survival (~ 4000 plantlets alive, ~ 80% survival over 33 years), their height and diameter at breast height were variable ranging 0.3–13.0 m and 2–149 mm, respectively (Puriņa et al., 2016B).
Thinning treatments and measurements
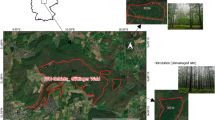
In the autumn/winter of 2016, the stand was divided into 45 regular 25 × 25 m sample plots (Fig. 1) and a complete shelterwood inventory was conducted (measuring all canopy trees and estimating understory density). Tree height (H) was measured by the Vertex clinometer (Haglöf Sweden AB) with a precision of 0.10 m and stem diameter at 1.3 m height (DBH) was measured with a calliper with a precision of one mm precision. Within each plot, beech saplings (advance growth) exceeding 30 cm in height have been mapped; 4094 trees in total. For each beech, height, and for those above 1.6 m, also DBH were measured. The condition of trees (living or dead), as well as damages (cut or damaged due to thinning performance, browsing damages) for each beech were recorded (Puriņa et al., 2016b).

Schematic depiction of thinning treatment design of the underplanted European beech in the northeasternmost plantation in Europe in the eastern part of Latvia. For each plot (total of 45) shelterwood species and treatment intensity are shown
In December 2016, the shelterwood thinning was performed by manual harvesting to minimize damage to the remaining trees and the beech advanced growth. The experimental design included six different levels of thinning treatments with diverse intensities (100, 75, and 50% of the upper canopy, second canopy storey, and second canopy storey with understorey removed) each carried under Scots pine and Norway spruce canopy (Table 1). Hence, for each treatment intensity and canopy species, two to seven plots were established (Table 1). The thinning was systematic (single stem), and the intensity was defined by the basal area removed (which correlated with the number of canopy trees). Second canopy storey removal and understory removal corresponded to < 20% of basal areas of shelterwood. The repeated measurements were done five years after the thinning treatment in autumn of 2021.
Statistical analysis
For the description of the mortality of beech in the plots of different thinning intensities during the five years, the total mortality rate r% was calculated as:
where N0 is the number of living beeches at the beginning of the observation; Nt is the number of living beeches at the end of the observation. For the description of shelterwood before and after the thinning, stand density, mean diameter and height increment, basal area, and standing volume (and their changes) were calculated. Stemwood volume was estimated based on local exponential species-specific equations (Liepa 1996).
To assess the effects of shelterwood treatments on the performance of stand, as well as on individual tree growth, according to the regional differences in management approaches, the effects of the treatments were estimated using linear models (analysis of covariance) at the tree and plot levels, respectively. At the plot (stand) level analysis, multiple linear (fixed) regression was used for each response variable. The models in general form were as follows:
where yi is the mean plot-level characteristics of beech (mortality, mean changes in DBH and H, basal area and stemwood volume five years after thinning) of the i-th plot; Ti is the thinning treatment (factorial, six levels); Si is the dominant species of shelterwood prior thinning (factorial, two levels); DENSi is the density of beech prior to thinning (numeric covariate); DBHi and Hi is the mean stem diameter at 1.3 m height and mean height of beech before thinning, respectively (numeric covariates). Generalization according to the binomial distribution was applied for survival data.
At the tree level, linear mixed-effects models with a plot as the random effect (intercept) were used to evaluate the effects of treatment on the characteristics of individual trees, while accounting for the spatial dependencies. The models fit for each response variable in general form were as follows:
where yi is the mean individual characteristics (stem diameter at 1.3 m height, tree height, and their absolute and relative increments five year after thinning) of the j-th beech in the i-th plot; Ti is the thinning treatment (factorial, six levels); Si is the dominant species of shelterwood prior thinning (factorial, two levels); DENSi is the density of beech prior to thinning (numeric covariate); dbhij and hij is the diameter at 1.3 m height and height of beech before thinning, respectively (numeric covariates); (pi) is the random effect of plot (factorial, 45 levels). The models were fit using a restricted maximum likelihood approach. The Wald’s type II χ2 test was used to estimate the significance of the fixed effects. The levels of treatment intensity were compared using the Tukey’s test. The data analysis was performed in R v. 4.2.2 (R Core Team 2022) using the package “lme4” (Bates et al. 2015).
Results
Before the shelterwood thinning in the winter of 2016, beech averaged 3.94 ± 0.09 m (mean ± standard error) in height and 2.38 ± 0.69 cm in DBH, which varied moderately among the sample plots (coefficient of variation was 0.23 and 0.17, respectively). The density of beech between the plots ranged from 336 to 2320 with the mean of 1446 ± 68 trees ha−1 (Supplementary material, Table S1). Overall during the five years after the thinning, mean H and DBH have increased by 1.55 ± 0.07 m and 2.42 ± 0.07 cm, respectively. The density of beech advance growth (underplanted saplings) in the second survey decreased to 1264 ± 370 trees ha−1, while showing a similar range as before (334–1888 trees ha−1). Five years after the thinning, 87.4% of beeches (3578 trees) had successfully survived, indicating overall low mortality. Among the 12.6% mortality of beech, 1.9% of trees died explicitly due to thinning (cut or broken). Although 14.3% of beeches had some browsing damage, it alone comprised < 1%, while likely contributing to 4.2% of the total mortality.
The tree-level analysis showed that the dimensions (size) of individual beeches and their increment in 2022 were significantly affected by treatment intensity, yet the effects were considerably stronger on DBH than H (Table 2). There, however, were no differences in responses of beech regarding the dominant shelterwood species, which were both conifers. The tree size covariates were strictly significant for all responses, yet the initial (before thinning) beech density lacked a significant effect. The marginal R2 values of the statistical models were high for the dimension and their increment, while the relative increments were considerably less affected by the treatments and pre-treatment conditions of trees. The effect of the plot was generally low, as the random variance comprised up to 10% (ICC ≤ 0.10), indicating spatially consistent responses.
Thinning intensity (treatments) had a generally positive, yet disproportional effect on DBH and, particularly, H of the underplanted beech (Fig. 2A). The lowest and highest DBH, which differed significantly, were estimated in the case of unthinned (control) and heavily (100% of canopy) thinned shelterwood, respectively. The differences between the weaker contrasts (understorey and/or second canopy storey, 50%, and 75% of shelterwood removed) were non-significant. The lowest H was estimated for the unthinned, slightly (second canopy storey and understorey removed) and heavily (100% of canopy) thinned shelterwood, which mutually lacked significant differences. The highest H and its increment were estimated under the intermediate thinning intensities (understorey and second canopy storey or 50% and 75% of shelterwood removed), which were similar. Still, the relative H increment lacked significant differences among the thinning treatments due to higher variability, although it followed a pattern similar to H.

The estimated marginal mean values of the underplanted beech size, survival, and increment according to shelterwood thinning treatments at the tree (A) and stand (B) level five years after the treatment in the easternmost forest plantation in Europe. The whiskers denote the estimated 95% confidence intervals of the marginal means. Similar letters above the bars denote a lack of significant differences according to Tukey’s test (p-value < 0.05). DBH—stem diameter at the 1.3 m height; H—tree height; G—basal area of stand; M—standing stock
The effect of shelterwood thinning at the stand level was generally weaker (Table 2) being significant (p-value < 0.05) for the increment of DBH, H, and standing volume of the underplanted beech. The thinning treatment showed a near significant effect (p-value ~ 0.08) on survival, while the changes in the stand basal area of beech were not affected. Regarding the increment of DBH and hence the basal area of beech, the dominant shelterwood species showed a significant effect. The tree size covariate (DBH before the shelterwood treatment) had a significant effect on the increment of H, basal area, and standing volume of the underplanted beech.
The highest mortality of beech (16.5%) occurred in the plots where 100% canopy was removed, however, in the other plots, it was still quite similar (ca.11.66%; Fig. 2B), though the significance of the differences was near marginal (p-value ~ 0.06). Surprisingly, the second highest mean DBH, though with the highest variability, was estimated for the control treatment, where all shelterwood was maintained. Still, the highest mean DBH was estimated when 75% of shelterwood was removed. The lowest DBH was estimated for the low intensity shelterwood thinning treatments, particularly if only the second canopy storey was removed; however, the differences were significant only between the strongest contrasts. The DBH increment under pine and spruce shelter was 2.67 ± 0.19 and 2.16 ± 0.09 cm 5y−1, respectively.
Contrasting DBH, the moderate shelterwood thinning (second canopy storey and 50% of the canopy removed) was estimated with the highest H increment, while the lowest H increment occurred in the unthinned plots, as well as in the case when second canopy storey and understorey was removed (Fig. 2B). The basal area increment followed the intensity of thinning, yet the differences among the treatments were not significant. Still, the basal area of beech was significantly higher (Table 2) under pine than spruce (2.22 ± 0.19 and 1.86 ± 0.17 m2 ha−1 5y−1, respectively). Moderate shelterwood thinning, when 50% of shelterwood was removed, had a positive effect on the standing volume of beech, which was considerably and significantly higher compared to the control treatment (Fig. 2B).
Discussion
Shelterwood thinning generally had a positive effect on the growth of 33-year-old underplanted beech supporting the ability of the species to utilize canopy openings (Weidig and Wagner 2021; Annighöfer 2018; Collet et al. 2011; Caquet et al. 2010). This agrees with previous studies, which have highlighted the importance of light gradients on growth patterns of beech under different shelterwood removal intensities (e.g., Weidig and Wagner 2021; Caquet et al. 2010; Collet et al. 2001). Beech growth is sensitive to environmental impacts and changes (Diaconu et al. 2015; Dittmar et al. 2003), hence growth responses can be explicit even in short-term surveys (Bontemps et al. 2010; Collet et al. 2001). The increase in DBH after the release, though, was more pronounced compared to H, similarly as observed within the current distribution range (Weidig and Wagner 2021; Collet and Chenost 2006). The sampling plot, had a low effect on the responses of beech, as suggested by the variance of random effect (Table 2) indicating homogeneous growing conditions. Alternatively, this might also indicate minimal effects of micro-site conditions despite the harsher climate (Cavin and Jump 2017). Still, the relative changes in beech dimensions were highly stochastic as indicated by low R2 values, while the dimension and increment were rather deterministic, implying dependence on the management (Bontemps et al. 2010; Dittmar et al. 2003; Collet et al. 2001).
The estimated overall mortality (including the thinning-related) over the post-treatment period (12.6% over five years, which corresponds to 2.7% annually) was slightly higher than observed in the core part of the distribution (~ 1.7% annually; Caquet et al. 2010; Kunstler et al. 2005), probably due to harsher climate (Weigel et al. 2018; Tompson and Fennessy 2011; Bolte et al. 2007). Nevertheless, the survival of beech did not differ by treatment, as observed previously (Caquet et al. 2010; Kunstler et al. 2005), indicating the robustness of underplanted beech regarding changes in illumination and microclimate (Wang et al. 2022; Čater et al. 2014; Lichtenthaler et al. 2007; Aussenac 2000). This contrasts the responsiveness to canopy opening of Norway spruce, which can be considered the main shade-tolerant competitor of beech in the northern part of the range (Parish and Antos 2005; Örlander and Karlsson 2000), supporting preconditions for the spread of beech in the understorey of coniferous stands. Although the studied plantation was unfenced, the beech mortality due to animal browsing was negligible, likely as the trees have long been present on the site (Puriņa et al. 2016b). Still, shelterwood thinning had increased browsing damage, similar to previous studies (Löf et al. 2005).
The pre-release size (H and, particularly, DBH) was a strong predictor of post-treatment dimensions of individual beeches (Table 2), thus indicating the effects of the initial condition and long-term beech seedling dominance (Caquet et al. 2010). However, at the stand level, the pre-release size of beech had a weaker effect on post treatment stand characteristics (Table 2), supporting robustness and resilience of the underplanted advance growth. The pre-release stand density had a significant positive effect on stand basal area and volume increment (Table 2), as previously observed by Weidig and Wagner (2021), which might be explained by the low initial density of beech saplings (ca. 1800 saplings ha−1; Puriņa et al. 2016b). Under such a low density, beech intra-specific competition was low, favouring horizontal crown development and diameter growth, hence volume growth of the stand (Collet and Chenost 2006; Leder and Weihs 2000). The increased mean DBH and basal area increment under Scots pine compared to Norway spruce canopy (Table 2) might be related to light transmission (Felton et al. 2020), which favours diameter growth of the shade tolerant species (Hara et al. 1991).
The explicit positive effect of shelterwood thinning, particularly that of high intensity (Fig. 2A) on individual trees complied with growth release in response to light availability (Weidig and Wagner 2021; Annighöfer 2018; Collet et al. 2011; Caquet et al. 2010; Collet and Chenost 2006). The additional increment of DBH of individual beech caused by shelterwood thinning over the 5 years (~ 12 mm; Fig. 2A), however, was lower compared to that in Germany (26.9 mm over 6 years; Weidig and Wagner 2021) likely due to harsher conditions. Though the stand/plot DBH responded to thinning disproportionally (Fig. 2B), suggesting the effect of light saturation (Linnert 2009; Petritan et al. 2009), which apparently reached an optimum level at 75% shelterwood removed, while excessive light caused inhibitory effects (Kunstler et al. 2005; Ammer 2003). The high plot DBH in the case of control treatment, however, can be an artefact of site design, as the control plots bordered with the plots of 50, 75 and 100% shelterwood thinning intensity, which likely provided additional light sufficient to stimulate growth (Kunstler et al. 2005; Ammer 2003).
The estimated additional annual H increment of beech under thinned shelterwood of 4–12 cm year−1 (compared to control; Fig. 2A, B) was comparable to that in Germany (15–20 cm year−1; Weidig and Wagner 2021), indicating quite efficient growth release. Although light availability and height growth of underplanted beech are correlated (Stiers et al. 2019; Petritan et al. 2007; Collet et al. 2002), the responses of H to thinning intensity were disproportional both at the tree and stand level (Fig. 2A, B) implying independence of density. Nevertheless, the disproportional responses can be explained by the interaction between the effects of light saturation (Linnert 2009; Petritan et al. 2009) and microclimate under harsher climate (Petritan et al. 2011; Klumpp et al. 2010). The lowest H increment was observed in plots where the second canopy and understorey were removed (Fig. 2), likely as the protection from frosts was absent (Petritan et al. 2011), yet light has not reached an optimal level (Ammer 2003). Accordingly, removing the second canopy storey maintaining the understorey likely minimized the effect of frosts aiding height growth. The highest H was estimated when 50% of shelterwood was removed, which likely matched optimal light conditions for primary growth (Kunstler et al. 2005; Ammer 2003), with radial increment being moderate (Collet and Chenost 2006; Leder and Weihs 2000). The reduction of H exceeding 50% of canopy removal (Fig. 2) might be associated with a saturation threshold, which corresponds to approximately 25% of the irradiation in an open field (Petritan et al. 2009; Collet and Chenost 2006; Stancioiu and O’Hara 2006). Exceeding the threshold, inhibitory effects of light on height growth can occur (Linnert 2009; Petritan et al. 2009; Robakowski and Antczak 2008; Aranda et al. 2001), as well as the growth tradeoffs might shift to radial growth (Collet and Chenost 2006; Leder and Weihs 2000).
The disproportional responses beech to shelterwood thinning intensity (Fig. 2A, B) might also be related to the dynamics of adjustments to changes in light conditions (Weidig and Wagner 2021; Lüttge et al. 2005; Valladares et al. 2002). For the understorey beech, adaptation of the photosynthetic apparatus to increased light might take up to a couple of years (Ammer 2003; Valladares et al. 2002; Aussenac 2000; Tognetti et al. 1998), implying an inability to immediately get advantage of ameliorated conditions (Annighöfer 2018; Martens and Preißler 2010; Collet and Chenost 2006). Furthermore, radial increment usually responds faster than height increment, which can show up to a three-year lag (Annighöfer 2018; Martens and Preißler 2010; Collet and Chenost 2006), as well as the duration of the environmental effects differ between them (Weidig and Wagner 2021; Martens and Preißler 2010). Furthermore, these relationships can be modulated by climatic conditions (Collet and Chenost 2006). Unfortunately, the study data disabled such analysis as the increment was not measured annually, nor they could be reliably reconstructed in the field (Weidig and Wagner 2021). Hence further monitoring of growth dynamics is necessary for the estimated mid-term responses to the treatment and sustainability of the stand.
The positive responsiveness of beech to shelterwood thinning (Fig. 2, Table 2), despite the reduction of protection against frosts and winter desiccation (Podrázský et al. 2019), supports the increasing growth potential of the species in the northern part of its projected distribution (Fortu 2023; Martinez del Castillo et al. 2022). Although the dataset was limited, the observed response of H suggests that the thinning of 50% of sheltering canopy trees in terms of basal area optimal for releasing underplanted beech. Regarding DBH, thinning of 75% of shelterwood caused the strongest increase of the basal area of underplanted beech, still, such an intense release might facilitate branching and reduce stem quality. Considering high survival before and after thinning and positive responses to moderate shelterwood thinning, underplanting of beech also appears as a promising tool for introducing deciduous admixture in coniferous stands thus facilitating their transformation according to the continuous cover approach. Still, as a single plantation was analysed, the obtained results encourage the extension of the research aiming to assess of responses of beech to management across a wider gradient of site conditions and compositions, as well as the timing of thinning.
Conclusions
Underplanted beech under marginal conditions in the northeasternmost outpost plantation in Europe responded positively to shelterwood thinning showing growth release despite the presumedly harsh climate. Such a positive response indicated ameliorating growing conditions and hence growing potential of the species in the Baltics. As hypothesized, thinning had a somewhat stronger effect on diameter than height growth. However, the estimated responses of beech growth proxies suggested the presence of thinning intensity when the basal area of canopy trees was reduced by half, yet undergrowth retained. Still, the growth release in the absolute numbers was moderate, likely due to relatively harsh climate and mesotrophic site. Considering high long-term survival and positive responsiveness to shelterwood thinning, underplating of beech appears as a promising tool for facilitating the transformation of coniferous stands according to the continuous cover approach. Based on the estimated responses, an extension of the research aiming to assessment of responses of beech to management across the wider ecological gradients in Latvia and the Baltics is encouraged.
References
Agestam E, Ekö PM, Nilsson U, Welander NT (2003) The effects of shelterwood density and site preparation on natural regeneration of Fagus sylvatica in southern Sweden. For Ecol Manag 176(1/3):61–73. https://doi.org/10.1016/S0378-1127(02)00277-3
Allen CD, Macalady AK, Chenchouni H et al (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259(4):660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Ammer C (2003) Growth and biomass partitioning of Fagus sylvatica L. and Quercus robur L. seedlings in response to shading and small changes in the R/FR-ratio of radiation. Ann for Sci 60(2):163–171. https://doi.org/10.1051/forest:2003009
Annighöfer P (2018) Stress relief through gap creation? Growth response of a shade tolerant species (Fagus sylvatica L.) to a changed light environment. For Ecol Manag 415:139–147. https://doi.org/10.1016/j.foreco.2018.02.027
Aranda I, Bergasa LF, Gil L, Pardos J (2001) Effects of relative irradiance on the leaf structure of Fagus sylvatica L. seedlings planted in the understory of a Pinus sylvestris L. stand after thinning. Ann for Sci 58:673–680. https://doi.org/10.1051/forest:2001154
Aussenac G (2000) Interactions between forest stands and microclimate: ecophysiological aspects and consequences for silviculture. Ann for Sci 57:287–301
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.48550/arXiv.1406.5823
Bolte A, Czajkowski T, Kompa T (2007) The north-eastern distribution range of European beech—a review. Forestry 80:413–429. https://doi.org/10.1093/forestry/cpm028
Boncina A, Kadunc A, Robic D (2007) Effects of selective thinning on growth and development of beech (Fagus sylvatica L.) forest stands in south-eastern Slovenia. Ann for Sci 64(1):47–57. https://doi.org/10.1051/forest:20060874
Bontemps JD, Hervé JC, Dhôte JF (2010) Dominant radial and height growth reveal comparable historical variations for common beech in north-eastern France. For Ecol Manag 259(8):1455–1463. https://doi.org/10.1016/j.foreco.2010.01.019
Caquet B, Montpied P, Dreyer E, Epron D, Collet C (2010) Response to canopy opening does not act as a filter to Fagus sylvatica and Acer sp. advance regeneration in a mixed temperate forest. Ann for Sci 67:105. https://doi.org/10.1051/forest/2009086
Čater M, Diaci J, Roženbergar D (2014) Gap size and position influence variable response of Fagus sylvatica L. and Abies alba Mill. For Ecol Manag 325:128–135. https://doi.org/10.1016/j.foreco.2014.04.001
Cavin L, Jump AS (2017) Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge. Glob Change Biol 23(1):362–379. https://doi.org/10.1111/gcb.13366
Collet C, Chenost C (2006) Using competition and light estimates to predict diameter and height growth of naturally regenerated beech seedlings growing under changing canopy conditions. Forestry 79(5):489–502. https://doi.org/10.1093/forestry/cpl033
Collet C, Fournier M, Ningre F, Hounzandji AP-I, Constant T (2011) Growth and posture control strategies in Fagus sylvatica and Acer pseudoplatanus saplings in response to canopy disturbance. Ann Bot 107:1345–1353. https://doi.org/10.1093/aob/mcr058
Collet C, Lanter O, Pardos M (2001) Effects of canopy opening on height and diameter growth in naturally regenerated beech seedlings. Ann for Sci 58(2):127–134. https://doi.org/10.1051/forest:2001112
Collet C, Lanter O, Pardos M (2002) Effects of canopy opening on the morphology and anatomy of naturally regenerated beech seedlings. Trees 16(4–5):291–298. https://doi.org/10.1007/s00468-001-0159-x
Diaconu D, Kahle HP, Spiecker H (2015) Tree-and stand-level thinning effects on growth of European beech (Fagus sylvatica L.) on a northeast-and a southwest-facing slope in southwest Germany. Forests 6(9):3256–3277. https://doi.org/10.3390/f6093256
Dittmar C, Zech W, Elling W (2003) Growth variations of common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe—A dendroecological study. For Ecol Manag 173:63–78. https://doi.org/10.1016/S0378-1127(01)00816-7
Durrant, TH, De Rigo D, Caudullo G (2016) Fagus sylvatica in Europe: distribution, habitat, usage and threats. European atlas of forest tree species. Luxembourg, Europe: European Commission. https://www.org/mtv/FISE-Comm/v01/e012b90. Accessed 30 September 2023
Felton A, Petersson L, Nilsson O et al (2020) The tree species matters: biodiversity and ecosystem service implications of replacing Scots pine production stands with Norway spruce. Ambio 49(5):1035–1049. https://doi.org/10.1007/s13280-019-01259-x
Fortu DA (2023) A comparison of thinning programmes in European beech (Fagus sylvatica L.) based on permanent plots in Southern Sweden. Master’s thesis, Swedish University of Agricultural Sciences
Tompson D, Fennessy J (2011) Beech (Fagus sylvatica) in Irish Forestry. In: Frýdl J, Novotný P, Fennessy J, von Wühlisch G (ed) Genetic Resources of Beech in Europe—Current State, Vol. 350, VTI, Braunschweig, pp 167–170
Hara T, Kimura M, Kikuzawa K (1991) Growth patterns of tree height and stem diameter in populations of Abies veitchii, A. mariesii and Betula ermanii. J Ecol 79:1085–1098. https://doi.org/10.2307/2261100
Hickler T, Vohland K, Feehan J, Miller PA, Smith B, Costa L, Giesecke T, Fronzek S, Carter TR, Cramer W, Kuhn I, Sykes MT (2012) Projecting the future distribution of European potential natural vegetation zones with a generalized, tree species-based dynamic vegetation model. Global Ecol Biogeogr 21:60–63. https://doi.org/10.1111/j.1466-8238.2010.00613.x
Jansone D, Matisons R, Kārše V, Bāders E, Kaupe D, Jansons Ā (2023) Structural heterogeneity of European beech (Fagus sylvatica L.) stands at its northernmost limits. Sustainability 15(20):14681. https://doi.org/10.3390/su152014681
Jansons Ā, Matisons R, Puriņa L, Neimane U, Jansons J (2015) Relationships between climatic variables and tree-ring width of European beech and European larch growing outside of their natural distribution area. Silva Fenn 49(1):1255. https://doi.org/10.14214/sf.1255
Klumpp R, Steiner H, Hochbichler E (2010) Current state of the European beech (Fagus sylvatica L.) gene-pool in Austria. In: Frýdl J, Novotný P, Fennessy J, von Wühlisch G (ed) Genetic Resources of Beech in Europe—Current State, vol 350. VTI, Braunschweig, pp 38–45
Kramer K, Degen B, Buschbom J, Hickler T, Thuiller W, Sykes M, de Winter W (2010) Modelling exploration of the future of European beech (Fagus sylvatica L.) under climate change—range, abundance, genetic diversity and adaptive response. For Ecol Manag 259:2213–2222. https://doi.org/10.1016/j.foreco.2009.12.023
Kunstler G, Curt T, Bouchaud M, Lepart J (2005) Growth, mortality, and morphological response of European beech and downy oak along a light gradient in sub-Mediterranean forest. Can J for Res 35:1657–1668. https://doi.org/10.1139/x05-097
Latvian Environment, Geology and Meteorology Centre (LEGMC) (2020) Climate in Latvia. https://klimats.meteo.lv/klimats/latvijas_klimats. Accessed 29 Oct 2023
Leder B, Weihs U (2000) Wachstum und qualitative Entwicklung eines 8 Jahre alten Buchen-Verbandsversuches unter Kiefer im Niederrheinischen Tiefland. Forst Und Holz 55(6):172–176
Leuschner C (2020) Drought response of European beech (Fagus sylvatica L.)-A review. Perspect Plant Ecol 47:125576. https://doi.org/10.1016/j.ppees.2020.125576
Lichtenthaler HK, Ač A, Marek MV, Kalina J, Urban O (2007) Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol Biochem 45:577–588. https://doi.org/10.1016/j.plaphy.2007.04.006
Liepa I (1996) Tree Growth Study. LUA, Jelgava (Latvia)
Linkevičius E, Junevičiūtė G (2021) Effect of competition and climatic conditions on the growth of beech in the mixed Pine Beech Stand: Lithuanian Case Study. Environ Sci Proc 3:49. https://doi.org/10.3390/IECF2020-07961
Linnert M (2009) Wachstum und Qualität junger Buchen in einem unterschiedlich aufgelichteten Fichtenaltbestand. Dissertation, Universität Göttingen
Löf M (2000) Influence of patch scarification and insect herbivory on growth and survival in Fagus sylvatica L., Picea abies L. Karst. and Quercus robur L. seedlings following a Norway spruce forest. For Ecol Manag 134:111–123. https://doi.org/10.1016/S0378-1127(99)00250-9
Löf M, Karlsson M, Sonesson K, Welander TN, Collet C (2007) Growth and mortality in underplanted tree seedlings in response to variations in canopy closure of Norway spruce stands. Forestry 80(4):371–383. https://doi.org/10.1093/forestry/cpm022
Löf M, Paulsson R, Rydberg D, Welander NT (2005) The influence of different overstory removal on planted spruce and several broadleaved tree species: survival, growth and pine weevil damage during three years. Ann for Sci 62:237–244. https://doi.org/10.1051/forest:2005015
Lüttge U, Kluge M, Bauer G (2005) Botanik, 5th edn. Wiley-VCH, Weinheim
Martens S, Preißler C (2010) Reaktion von Verjüngungen unter Fichtenschirm auf plötzliche Freistellung. AFZ-Der Wald 65(10):14–18
Martinez del Castillo E, Zang CS, Buras A et al (2022) Climate-change-driven growth decline of European beech forests. Commun Biol 5(1):163. https://doi.org/10.1038/s42003-022-03107-3
Matisons R, Puriņa L, Adamovičs A, Robalte L, Jansons Ā (2017) European beech in its northeasternmost stands in Europe: Varying climate-growth relationships among generations and diameter classes. Dendrohronologia 45:123–131. https://doi.org/10.1016/j.dendro.2017.08.004
Matisons R, Šņepsts G, Puriņa L, Donis J, Jansons Ā (2018) Dominant height growth of European beech at the northeasternmost stands in Europe. Silva Fenn 52(1):7818. https://doi.org/10.14214/sf.7818
Örlander G, Karlsson C (2000) Influence of shelterwood density on survival and height increment of Picea abies advance growth. Scand J for Res 10:20–29. https://doi.org/10.1080/02827580050160439
Övergaard R, Agestam E, Ekö PM, Johansson U (2009) A method for natural regeneration of beech (Fagus sylvatica L.) practiced in southern Sweden. Stud for Suecica 218:30
Parish R, Antos JA (2005) Advanced regeneration and seedling establishment in small cutblocks in high-elevation spruce-fir forest at Sicamous Creek, southern British Columbia. Can J for Res 35:1877–1888. https://doi.org/10.1139/x05-108
Petritan AM, von Lüpke B, Petritan IC (2007) Effects of shade on growth and mortality of maple (Acer pseudoplatanus), ash (Fraxinus excelsior) and beech (Fagus sylvatica) saplings. Forestry 80(4):397–441. https://doi.org/10.1093/forestry/cpm030
Petritan AM, von Lüpke B, Petritan IC (2009) Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.) maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur J for Res 128(1):61–74. https://doi.org/10.1007/s10342-008-0239-1
Petritan IC, Lüpke BV, Petritan AM (2011) Influence of shelterwood and ground vegetation on late spring frost damages of planted beech (Fagus sylvatica) and Douglas-Fir (Pseudotsuga menziesii) saplings. Balt for 17(2):227–234
Podrázský V, Baláš M, Linda R, Křivohlavý O (2019) State of beech pole stands established at the clear-cut and in the underplanting. J for Sci 65:256–262. https://doi.org/10.17221/59/2019-JFS
Purina L, Adamovics A, Katrevics J, Katrevica Z, Dzerina B (2016) Growth of Fagus sylvatica in young mixed stand: case study in central Latvia. Res Rur Dev 2:21–27
Puriņa L, Matisons R, Jansons Ā, Šēnhofa S (2016) Survival of European beech in the central part of Latvia 33 years since the plantation. Silva Fenn 50(4):1. https://doi.org/10.14214/sf.1656
R Core Team. (2022) R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/. Accessed 15 Oct 2023.
Reventlow DOJ, Nord-Larsen T, Sørensen H, Skovsgaard JP (2023) Effects of Norway spruce shelter density and soil scarification method on mortality and height growth of underplanted European beech, European silver fir and Douglas fir. Forestry 96(4):481–497. https://doi.org/10.1093/forestry/cpac051
Reventlow DOJ, Nord-Larsen T, Skovsgaard JP (2018) Pre-commercial thinning in naturally regenerated stands of European beech (Fagus sylvatica L.): effects of thinning pattern, stand density and pruning on tree growth and stem quality. Forestry 92:120–132. https://doi.org/10.1093/forestry/cpy039
Robakowski P, Antczak P (2008) Ability of silver fir and European beech saplings to acclimate photochemical processes to the light environment under different canopies of trees. Pol J Ecol 56(1):3–16
Roibu CC, Palaghianu C, Nagavciuc V et al (2022) The response of beech (Fagus sylvatica L.) populations to climate in the easternmost sites of its European distribution. Plants 11(23):3310. https://doi.org/10.3390/plants11233310
Stancioiu PT, O’Hara KL (2006) Regeneration growth in different light environments of mixed species, multiaged, mountainous forests of Romania. Eur J for Res 125:151–162. https://doi.org/10.1007/s10342-005-0069-3
Stiers M, Willim K, Seidel D, Ammer C, Kabal M, Stillhard J, Annighöfer P (2019) Analyzing spatial distribution patterns of European beech (Fagus sylvatica L.) regeneration in dependence of canopy openings. Forests 10:637. https://doi.org/10.3390/f10080637
Tognetti R, Minotta G, Pinzauti S, Michelozzi M, Borghetti M (1998) Acclimation to changing light conditions of long-term shadegrown beech (Fagus sylvatica) seedlings of different geographic origins. Trees 12:326–333. https://doi.org/10.1007/PL00009719
Valladares F, Chico JM, Aranda I, Balaguer L, Dizengremel P, Marinque E, Dreyer E (2002) The greater seedling high-light tolerance of Quercus robur over Fagus sylvatica is linked to greater physiological plasticity. Trees 16:395–403. https://doi.org/10.1007/s00468-002-0184-4
Wang H, Lin S, Dai J, Ge Q (2022) Modeling the effect of adaptation to future climate change on spring phenological trend of European beech (Fagus sylvatica L.). Sci of Total Environ 846:157540. https://doi.org/10.1016/j.scitotenv.2022.157540
Weidig J, Wagner S (2021) Growth response of advanced planted European beech (Fagus sylvatica L.) after storm-caused loss of shelterwood. Eur J for Res 140:931–946. https://doi.org/10.1007/s10342-021-01376-x
Weigel R, Muffler L, Klisz M, Kreyling J, van der Maaten-Theunissen M, Wilmking M, van der Maaten E (2018) Winter matters: Sensitivity to winter climate and cold events increases towards the cold distribution margin of European beech (Fagus sylvatica L.). J Biogeogr 45(12):2779–2790. https://doi.org/10.1111/jbi.13444
Funding
This study was supported by JSC Latvia’s State Forests project “Support for Forest Seed production”.
Author information
Authors and Affiliations
Contributions
Conceptualization, I.M., A.J., methodology, A.J., I.J., software, I.M., R.M., validation, A.J., formal analysis, I.M., R.M., investigation, I.J., A.A., resources, A.J., data curation, I.M., A.J., I.J., writing—original draft preparation, I.M., writing—review and editing, I.M., A.J., R.M., visualization, I.M., R.M., supervision, A.J., project administration, A.J., funding acquisition, A.J. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Matisone, I., Jaunslaviete, I., Adamovičs, A. et al. Response of underplanted European beech to shelterwood thinning at the northeasternmost forpost plantation in Europe. New Forests 55, 1267–1281 (2024). https://doi.org/10.1007/s11056-024-10033-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-024-10033-1