
Abstract
Eucalyptus are trees sensitive to interference imposed by weeds mainly during the early growth phase. The repetitive use of herbicides with the same mechanism of action has led to the selection of resistant weeds, and Digitaria insularis stands out for its resistance to glyphosate. Due to its occurrence in eucalyptus plantations, knowledge regarding D. insularis interference in eucalyptus early growth is needed to develop management strategies. This study aimed to confirm D. insularis glyphosate-resistence; and also to evaluate the effect of densities and distances of glyphosate-resistant D. insularis, seeded and in regrowth, on the early growth of Eucalyptus urophylla × E. grandis (Clone AEC-144). Prior to eucalyptus’ experiments, D. insularis glyphosate-resistance was confirmed through a dose–response curve experiment. After that, two experiments [one for weed’s first growth (seeded) and other for second growth (regrowth)] were carried out in an open area, in 700 L pots, for 105 days after planting eucalyptus seedlings. The treatments consisted of increasing densities of D. insularis (0, 1, 2 and 3 plants m−2) planted at 10, 20 or 30 cm from the eucalyptus seedlings. Eucalyptus height, stem diameter, leaf area and dry matter were evaluated. The eucalyptus variables most sensitive to interference were leaf and stem dry matter, with reductions of 45% and 88.4%, respectively for weeds seeded and regrowth. Weed second growth (regrowth) interfered more aggressively in eucalyptus growth. The coexistence with D. insularis, regardless of density and distance, negatively interfered early tree growth.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Eucalyptus are the most important tree crop for the Brazilian plantation forest sector comprising a total planted area of 9.39 million hectares in 2021, in which 75.8% is made up of eucalyptus plantations (Ibá 2022). In the same year, Brazilian eucalyptus forests had an average productivity of 38.9 m3 ha−1 year−1, making them the most productive eucalyptus forests in the world (Ibá 2022), where the species (hybrid) mainly planted is Eucalyptus urophylla × E. grandis (Gonçalves et al. 2013). The high productivity of this crop in Brazil is due to the development of breeding programs, as well as crop management strategies to optimise productivity, including weed management (Stape et al. 2004; Pereira et al. 2012; Gonçalves et al. 2013).
Planted forests can be negatively affected by interference from several ecological factors, which directly influence their development (Pitelli and Marchi 1991) and, consequently, their productivity (Garau et al. 2009; Cruz et al. 2010; Tiburcio et al. 2023). During the initial phase of eucalyptus growth, which is the period comprising the first year after planting the crop, the plants are more vulnerable to interference caused by abiotic (such as water and nutritional deficiencies) and biotic factors (Florentine and Fox 2003; Schaller et al. 2003; Coll et al. 2004; Garau et al. 2008; Bacha et al. 2018).
Among the biotic factors, the competition imposed by neighbouring vegetation stands out, since the lack of control of the on-site vegetation during the first six months after planting eucalyptus can cause losses of up to 38% in wood volume after 7 years of cultivation (Little and Staden 2005; Hunt et al. 2006a, b; Little et al. 2007; Qin et al. 2018; Tiburcio et al. 2023). In addition to competing with the crop for water and nutrients, weeds can also release allelochemicals into the environment, negatively affecting eucalyptus growth (Watt et al. 2003; Graat et al. 2018). Thus, studies to this effect have been developed around the world in recent decades (Sands and Nambiar 1984; Caldwell et al. 1995; Adams et al. 2003; Coll et al. 2004; Garau et al. 2008; Colmanetti et al. 2017; Tiburcio et al. 2023). Despite this, information about competition of eucalyptus with herbicide-resistant weeds is scarce (Braga et al. 2018), a notable gap in information, especially due to the increase in cases of herbicide-resistant biotypes in Brazil and worldwide (Heap 2023).
Weed herbicide-resistance is defined as the ability of a plant to survive and reproduce after exposure to a dose of an herbicide normally lethal to wild individuals of this species (Powles and Yu 2010). Glyphosate is one of the most important herbicides in the world, being used to control annual or perennial weeds in several production systems (Blackshaw and Harker 2002; Hébert et al. 2019; Clapp 2021). The wide genetic variability of weeds allows their survival in different environmental conditions (Christoffoleti and López-Ovejero 2003); additionally, the selection pressure of resistant biotypes, offered by the repetitive use of the same mechanism of action, has considerably increased the reports of herbicide-resistant biotypes around the world (Heap and Duke 2018; Netto et al. 2021; Heap 2023).
The native grass species Digitaria insularis (L.) Fedde (sourgrass), whose resistance to glyphosate has already been proven in Brazil (Carvalho et al. 2011; Heap 2023), is now considered one of the main weeds of crops tolerant to this herbicide (Cerdeira et al. 2011; Cesarin et al. 2019; Netto et al. 2021). This species was considered marginal in areas planted to eucalyptus, but recently it has been recorded in eucalyptus plantations (Barroso et al. 2021), a concern for Brazilian forest growers, given its rapid dispersion and the increase in reports of glyphosate-resistant biotypes (Heap 2023).
In addition to the species of neighbouring vegetation that can co-occur with planted trees, other factors also directly affect the degree of interference of the neighbouring plant community in the crop (Bleasdale 1960; Pitelli 1985), such as: density and distance from eucalyptus plants (Dinardo et al. 2003; Graat et al. 2015; Costa et al. 2021); the period of coexistence between the neighbouring vegetation and planted trees (Toledo et al. 2000; Adams et al. 2003; Tarouco et al. 2009); and also eucalyptus clone resistance to competition (Pereira et al. 2013; Resende et al. 2016; Colmanetti et al. 2017; Pavan et al. 2021). From this perspective, the amount of neighbouring vegetation plants per unit area, or density, is highlighted, since even at low density (3 plants m−2) some species can lead to reductions of up to 48% in eucalyptus dry matter, such as Urochloa decumbens (Stapf) R.D.Webster (signal grass) (Bacha et al. 2016). Thus, as a result of the increase in cases of glyphosate-resistant D. insularis in eucalyptus plantations in Brazil, and also because there are no studies on the interference of this species on planted eucalyptus establishment, studies are needed to determine whether this could be a concern for forest growers. Furthermore, glyphosate-resistance can result in inadequate chemical-control of this species, causing regrowth (or second growth) of plants after necrosis of leaves affected by the herbicide, which can also influence the degree of interference in the crop (Bacha et al. 2016).
Such information is necessary to support effective weed management strategies in Eucalyptus plantations, such as, for example, the timing of intervention or management and the width of the control strip to be adopted (Silva et al. 2012; Tiburcio et al. 2023). Thus, the present study aimed to: (i) confirm D. insularis glyphosate-resistence; (ii) evaluate the effect of densities and distances of glyphosate-resistant D. insularis, seeded and in regrowth, on the early growth of Eucalyptus urograndis.
Materials and methods
Experiments 1 and 2—Digitaria insularis first growth (from seeds) and second growth (regrowth from first experiment’s plants)
Experimental area, plant material and treatments
Two experiments [one for the weed’s first growth (from seeds, i.e. weed seeded hereafter) and other for the second growth (after cutting, i.e. weed regrowth hereafter)] were conducted in an open experimental area at Sao Paulo State University (UNESP), Jaboticabal-SP, Brazil. The altitude of the site is 590 m and the geographic coordinates are: latitude 21°15′17″ S and longitude 48°19′20″ W. The climate of the region, according to the Köppen (1948) classification, is Cwa, subtropical, dry in winter, with summer rains, with an annual average temperature of 23.3 °C and precipitation of 1243.92 mm between 2018 and 2022. The first experiment was conducted between September and December 2019, and the second experiment was conducted between April and July 2020.
In both experiments, 700 L pots were used (1 × 1 × 0.7 m), with a hollow bottom, filled with soil from a Dystrophic Red Latosol (physical–chemical analysis of the substrate in Table 1). For the first experiment, one week before eucalyptus planting, an amount equivalent to 3 t ha−1 of dolomitic limestone per pot was added. A planting fertilization was carried out with the 27–00–10 formula (N–P–K) at the equivalent dose of 300 kg ha−1.
For both experiments, regional representative commercial seedlings [E. urophylla × E. grandis (Clone AEC-144)], purchased from Agriflora® (approximately 100 days old), were transplanted in the center of the pots. Each pot, with one eucalyptus plant, was considered an experimental plot. For experiment 1, eucalyptus seedlings had an average height of 35 cm and a stem diameter of 2.96 mm. In experiment 2, the average height was 32 cm and stem diameter of 2.85 mm.
For both experiments, a randomized block design was used, with treatments arranged in a 3 × 3 + 1 factorial scheme (double factorial plus one additional treatment), with 5 replications. The treatments consisted of the densities of 1, 2 and 3 D. insularis plants m−2, planted at 10, 20 or 30 cm away from the eucalyptus seedlings, in addition to a weed-free control, totaling 10 treatments. For each experiment, a total of 50 pots were used (five for each treatment). The distribution of plants in the pots and the treatments’ layout are presented in the supplementary material (Figures S1 and S2).
In the first experiment, 15 days after planting the eucalyptus seedlings (DAPL), the D. insularis (Digitaria insularis) seedlings were transplanted to the pots, acquired from previous sowing in styrofoam trays with 128 cells filled with horticultural substrate. The seeds were collected at the UNESP Experimental Farm (Jaboticabal-SP, Brazil), where the case of glyphosate-resistant D. insularis has already been proven (Carvalho et al. 2011). At the time of planting, seedlings were chosen according to their size uniformity (two fully expanded leaves and 5–7 cm height), and positioned in the pots according to the proposed treatments.
For the second experiment, the regrowth of weeds previously planted on Experiment 1 was used, at the same pots, densities and distances aforementioned. Thus, the day before eucalyptus planting, all D. insularis plants were cut at 5 cm from the ground, so that plants were allowed to regrowth uniformly. At 75 DAPL, a cover fertilization was carried out, applying the same formulation and dose previously described.
Both experiments were conducted without water restriction, for 105 DAPL. In order to avoid the interference of any other weed species in the experiments, weekly manual weeding was carried out in all experimental plots.
Assessed variables
For both experiments, fortnightly, eucalyptus height (from the base of the plant to its apex, with the aid of a 100 cm ruler) and stem diameter (measured at 5 cm from the ground, using a digital caliper) assessments were carried out up to 105 DAPL. In the second experiment, at 75 DAPL, the relative total chlorophyll content was evaluated on the third fully expanded leaf, using a chlorophyll meter (Falker®, mod. ClorofiLog CFL 1030).
At the end of both experimental period (105 DAPL each), the eucalyptus plants were cut at their base and the leaves detached for leaf area measuring (LiCor®, mod. LI-3000A). Then, leaves and stems were dried in an oven with forced air circulation (~ 60 °C) for 7 days, for subsequent determination of dry matter mass, after weighing on a precision electronic scale.
Experiment 3—dose–response curve for Digitaria insularis glyphosate-resistance confirmation
In order to confirm D. insularis glyphosate-resistance, a third experiment was conducted.
When planting D. insularis seedlings in Experiment 1, the plants were also transplanted into pots with a capacity of 0.5 L, filled with a mixture of soil, river sand and horticultural substrate (1:1:1 v/v/v) and cultivated in open area, without water restriction.
When the plants reached an average of 15–20 cm in height and the beginning of tillering [stage 21 of the Bleiholder et al. (1991)], they were subjected to an application of glyphosate (Roundup® Transorb R). Treatments consisted of doses equivalent to 0, 210, 420, 840, 1680, 3360 and 6720 g a.e. ha−1 of glyphosate, corresponding to 0%, 12.5%, 25%, 50%, 100%, 200% and 400% of the recommended dose for D. insularis control in eucalyptus crop. A completely randomized experimental design was used, with seven treatments, in four replications.
The herbicide application was carried out with a backpack sprayer at constant pressure (CO2), equipped with a bar with four XR11002 tips and regulated for a tank volume of 200 L ha−1. At the time of application, which was carried out in a spray room, the air temperature was 25 °C and the relative humidity was 76%.
Visual control evaluations were carried out according to the scale proposed by the Asociación Latinoamericana de Malezas (ALAM 1974), considering a score of 0 for absence of injury, up to 100% for plant death. These evaluations were performed at 7, 14, 21 and 28 days after glyphosate application (DAA). At 28 DAA, the remaining green shoots of the plants were cut close to the ground and placed to dry in an oven with forced air circulation at 60 °C, for seven days, for subsequent determination of the dry matter mass after weighing on a precision scale.
Statistical analysis
For both eucalyptus competition experiments, the data obtained were submitted to analysis of variance (ANOVA) (Fisher 1925) by the F test and the averages compared by the Tukey test at 5% probability, using the AgroEstat® software.
For the dose response study, the control scores data were graphically expressed, while dry matter results were submitted to non-linear log-logistic regression to determine the necessary glyphosate dose for a 10%, 50% and 90% reduction in dry matter (Eq. 1).
where Y is the dry matter reduction expressed in percentage of reduction compared to the untreated control; a represents the maximum asymptote, xc the necessary value for 50% of the dry mass reduction per biotype, and k a regression constant.
Furthermore, to identify the effect of all treatments (from the first and second weed growth) on the eucalyptus plants, a principal component analysis (PCA) was performed. The process was carried out by reducing the multivariate data matrix to an interpretable two-dimensional biplot that explains the greater proportion of variation in the data obtained in both experiments. All graphics were made with Origin® software.
Results
Experiment 1—Digitaria insularis first growth (from seeds)
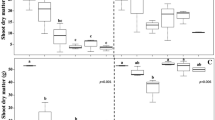
For all variables (except for leaf area), the factorial differed from the control (additional factor) (Table 2). The competition with D. insularis plants, regardless of density and distance, reduced, on average, 14.4% eucalyptus height, 45% leaf dry matter (LDM) and 41% total dry matter (TDM) (Table 2).
Regarding eucalyptus height, there was no effect of D. insularis density. Plants at 10 and 20 cm from the eucalyptus caused an 11.5% reduction compared to those grown at 30 cm, considering the mean densities (Table 2). Compared to control, these treatments caused reductions of 17.9% to eucalyptus’ height (Table 2).
Next, emphazis will be given to the key outcomes regarding the interaction between the factors (densities × distances) for eucalyptus’ stem diameter, leaf area, leaf dry matter, stem dry matter and total dry matter (Table 3).
Considering plants grown 10 cm away from eucalyptus, the densities of 2 and 3 plants m−2 reduced 15% eucalyptus stem diameter compared to 1 plant m−2 (Table 3A). The highest density was the only treatment that showed no difference in relation to the evaluated distances (Table 3A).
With respect to leaf area—LA (Table 3B), for plants grown at 10 cm, the densities of 2 and 3 plants m−2 caused an average reduction of 32%, when compared to 1 plant m−2. Considering the distances within the densities, there was a difference between the treatments only for 3 plants m−2, in which the cultivation of eucalyptus 10 cm from the D. insularis caused a 23% reduction in the LA, compared to the average of the greatest distances (Table 3B).
Considering eucalyptus leaf dry matter (LDM) (Table 3C), the highest density was different from the others only when the plants were 10 cm away, causing an average reduction of 34.8%. For this density, the closest distances between the weed and eucalyptus (10 and 20 cm) reduced the LDM by 34.9%, compared to those located at 30 cm (Table 3C). This response pattern was different for the densities of 1 and 2 plants m−2, in which the closest proximity between the plants (10 cm) was the one that least affected the eucalyptus LDM (Table 3C). Compared to weed-free control, D. insularis grown 20 cm away from eucalyptus caused reductions of 55.9% in this variable (means of all densities – Table 3C).
For the stem dry matter (SDM) (Table 3D), considering the distance of 10 cm, the interference in eucalyptus growth was progressive, being directly proportional to the increase in density. Thus, the densities of 2 and 3 plants m−2 were responsible for reductions of 30% and 53%, respectively, compared to 1 plant m−2 (Table 3D). Considering the higher density, the distances of 10 and 20 cm between plants caused greater reductions for eucalyptus in relation to cultivation at 30 cm. The other densities showed different responses regarding distances (Table 3D).
Eucalyptus total dry matter (TDM) indicated the same response pattern observed for SDM, considering both densities and distances. For 10 cm, there was a progressive increase in the interference of eucalyptus growth according to the increase in density (Table 3E). On that occasion, 3 plants m−2 reduced eucalyptus TDM by 46.4%, compared to 1 plant m−2. For the treatment with 3 plants m−2, the cultivation at 30 cm obtained values 42% higher than 10 and 20 cm (Table 3E).
For eucalyptus height growth during the experimental period, evaluations prior to 90 DAPL were not significant comparing treatments (p > 0.05—data not shown); from that date onwards, weed densities and distances interfered with crop growth (Fig. 1a and c). At 105 DAPL, 3 D. insularis plants m−2 reduced, on average, eucalyptus height by 12.9% compared to control (Fig. 1a). The plants positioned at 10 and 20 cm caused an average reduction of 17.9% in relation to the control, differing from the 30 cm treatment which caused 7.22% reduction (Fig. 1c).

Effect of different densities of glyphosate-resistant Digitaria insularis first growth (seeded) on height (A) and stem diameter (B) of E. urophylla × E. grandis (Clone AEC-144) plants during 105 days after planting. Graphs C and D indicate the different distances considering the mean densities, for height and stem diameter of eucalyptus plants, respectively. For the densities (A and B), the indicated values are composed by the average distances from the weed. Different letters indicate a significant difference by Tukey’s test at 5% probability
The stem diameter, on the other hand, proved to be more sensitive than height, regarding the competition imposed by D. insularis, since at 75 DAPL the control reached the same values observed for treatments with 2 and 3 plants m−2 (mean distances—Fig. 1b) and for 20 cm (density averages—Fig. 1d) at 105 DAPL. At the end of the experiment, the reductions compared to the control were 20.2% and 22.7%, respectively for the densities of 2 and 3 m−2 and 20 cm (Fig. 1b and d).
Experiment 2—Digitaria insularis second growth (regrowth from first experiment’s plants)
For the second experiment, the factorial differed from the control (additional factor) for all evaluated variables (Table 4). The interference of D. insularis plants on eucalyptus growth was greater in regrowth (experiment 2), compared to the weed seeded (Experiment 1—Table 2). The coexistence with D. insularis, regardless of density and distance, reduced, on average, 44.5% eucalyptus height, 65% LA and 88.4% SDM (Table 4).
For the eucalyptus height and stem diameter, 2 D. insularis m−2 caused mean reductions of 12 and 19%, respectively, compared with the density of 1 plant m−2, regardless of distance (Table 4). The weeds positioned 10 cm from the eucalyptus caused the greatest interference in the stem diameter, compared to those positioned at 30 cm (Table 4). Considering the total chlorophyll content, all treatments obtained values 16% lower than control, regardless of densities and distances (Table 4).
With respect to LA of eucalyptus (Table 5A), the densities of 2 and 3 plants m−2 caused reductions of 57%, compared to 1 plant m−2, when grown at 20 cm apart. At the lowest density, only the 10 cm treatment caused a reduction in the LA, while for 2 and 3 plants m−2, the 30 cm distance obtained higher values compared to the other treatments (Table 5A).
For LDM, SDM and TDM (Table 5B–D, respectively), the response pattern was the same as that observed for LA, in that the two highest densities caused greater interference in eucalyptus growth compared to 1 m−2, for 20 cm. Regarding the distances for the 1 plant m−2, it was observed that the closer the weed was to the eucalyptus, the lower the values obtained for LDM and TDM, with average reductions of 64.7% compared to 20 cm (Table 5B and D). As for the density of 3 D. insularis m−2, the cultivation at 30 cm provided values 86% and 73% higher than the other distances, respectively for LDM and TDM (Table 5B and D). As for the SDM, considering 2 plants m−2, the coexistence between the plants at 30 cm resulted in a stem diameter growth 174% greater than the other distances (Table 5C). Despite this, compared to control, the average reduction caused by coexistence with 2 plants m−2 was 90% (mean of the distances) (Table 5C).
Although the first and second experiments were not compared by ANOVA’s test, the results indicate that Digitaria insularis regrowth was considerably more aggressive to eucalyptus growth compared to seeded plants, since before 60 DAPL, the control had already reached the same values as the treatments with higher densities and shorter distances at 105 DAPL (Fig. 2).

Effect of different densities of glyphosate-resistant Digitaria insularis second growth (regrowth) on height (A) and stem diameter (B) of E. urophylla × E. grandis (Clone AEC-144) plants during 105 days after planting. Graphs D and D indicate the different distances considering the mean densities, for height and stem diameter of the eucalyptus, respectively. For the densities (A and B), the indicated values are composed by the average distances from the weed. Different letters indicate a significant difference by Tukey’s test at 5% probability
For eucalyptus height, at 60 DAPL the treatments already differed from the control (p < 0.05—data not shown), considering the mean densities and distances (Fig. 2a and c). At the end of the experiment, the densities of 2 and 3 plants m−2 caused reductions of 46.7% in eucalyptus height (average distances—Fig. 2a). The distances tested did not differ from each other, with reductions of 44.4%, on average, compared to control (Fig. 2c).
Regarding stem diameter, as early as 45 DAPL the treatments already differed from the control (p < 0.05—data not shown), regardless of density or distance (Fig. 2b and d). At the end of the experimental period, the highest densities reduced the eucalyptus stem diameter by 57.9% (mean distances—Fig. 2b), while plants at 10 cm caused a reduction of 61.5% in this variable, considering the mean from tested densities (Fig. 2d).
Principal component analysis (PCA) accounted for a total of 98.63% of the original information, corresponding to 95.4% for PC1 and 3.23% for PC2 (Fig. 3). This result is in agreement with the criteria established by Sneath and Sokal (1973), in which the main components must explain at least 70% of the total variance of the presented data.

Principal Component Analysis (PCA) of growth variables of Eucalyptus urophylla × E. grandis (Clone AEC-144) after 105 days of competition with glyphosate-resistant Digitaria insularis planted at different densities and distances, seeded or in regrowth. DM = dry matter. S or R = weeds seeded or in regrowth. 10, 20 or 30 = distance between eucalyptus plants and weeds. 1, 2 or 3 = D. insularis density (plants m−2)
The PCA resulted in the formation of four treatment groups. The first group consisted of control plants, conducted in the experiment in regrowth (red dot); the second group was formed by treatments with D. insularis regrowth, regardless of density and distance, inversely correlated to PC1 (purple dots); the third group consisted of eucalyptus cultivated without weeds, in experiment 1 (green dot); and the fourth group was formed by treatments with D. insularis plants seeded, regardless of density or distance (blue dots) (Fig. 3). All evaluated variables showed direct correlation with PC1.
Experiment 3—dose–response curve for Digitaria insularis glyphosate-resistance confirmation
Glyphosate was effective in controlling (at least 90% of control) the D. insularis genotype used in the experiments only with doses of 3360 and 6720 g a.e. ha−1 (double and four times the recommended dose—Figs. 4a and S2C). The recommended dose and the others (x/2, x/4 and x/8) obtained control scores lower than 70% at the end of the evaluation period, not being effective for control of D. insularis (Fig. 4a), thus demonstrating a possible herbicide-resistance.

Dose–response curve for Digitaria insularis control with glyphosate (A) and for the weed dry mass at 28 days after herbicide application (B)
To obtain a 50% reduction in dry mass (LD50) of this D. insularis genotype, a dose of 1045 g a.e. ha−1 of glyphosate was needed (Fig. 4b), which possibly reveals a low resistance factor of this plant material. For reductions of 10% (LD10) and 90% (LD90) in weed dry matter, glyphosate doses of 196 and 3360 g a.e. ha−1 were required, respectively (Fig. 4b).
Discussion
The reduction of some planted eucalyptus size variables evaluated in the present study, such as plant’s height, stem diameter and dry matter (Tables 2, 3, 4, 5 and Figs. 1, 2, 3), is a direct result of inter-specific competition between D. insularis and E. urograndis in the period following planting and corroborate results found by several authors (Toledo et al. 2001; Hunt et al. 2006a, b; Garau et al. 2009; Bacha et al. 2016; Colmanetti et al. 2019). Eucalyptus plants tend to be highly sensitive to competition imposed by weeds in the first year after planting the crop, a period known as initial growth (Adams et al. 2003; Florentine and Fox 2003; Garau et al. 2009). Such sensitivity in this period should be highlighted especially due to the fact that most commercial eucalyptus plantations in Brazil have short rotation cycles (approximately 7 years), which can considerably impact productivity (Gonçalves et al. 2013; Tiburcio et al. 2023). Despite the short term evaluation carried out in this study, others with longer periods of competition indicated response trends similar to those observed here. Thus, the lower difference between treatments with and without competition found in experiment 1, for height and stem diameter (Fig. 1), may be due to this factor. Possibly the difference between these treatments would be increasingly greater as the period of coexistence was prolonged, similarly to those reported in studies longer than one year (Toledo et al. 2000; Adams et al. 2003; Garau et al. 2009; Tarouco et al. 2009). Thus, the lack of control of competing plants can cause eucalyptus early growth losses, as shown here (Tables 2, 3, 4, 5 and Figs. 1, 2, 3).
The distribution of weeds in the area is also a relevant factor to be considered in eucalyptus plantations. The results obtained for the D. insularis first growth showed that the leaf area did not differ from the control (additional factor) in realtion to the three distances tested, regardless of the densities (Table 2). However, in regrowth condition, the weed coexistence 10 cm away from eucalyptus resulted in reductions of 79.5%, whereas at 30 cm, the reductions were of 53% compared to control, regardless of the densities (Table 4). These data show that in the first weed growth (i.e., from seeds) distances of up to 30 cm from the planted eucalyptus equally interfere with crop growth, whereas in regrowth, the interference was more aggressive the closer the weed was to the eucalyptus plants. Corroborating these results for weeds’ first growth, Graat et al. (2015) observed that only one plant of U. decumbens or Urochloa ruziziensis [(R.Germ. & C.M.Evrard) Crins (Congo grass)] grown at increasing distances of up to 40 cm from E. urograndis reduced crop growth. However, no significant differences regarding the tested distances were observed by the authors.
In addition to the distance between weeds and the crop plants, other biotic and abiotic factors can also affect the degree of competition from the weed community in eucalyptus establishment (Bleasdale 1960; Pitelli 1985), such as: region’s edaphoclimatic conditions, species or clone studied, and also weed species and density. In this context, Colmanetti et al. (2019) observed that a density of 33 plants m−2 of Urochloa brizantha [(A.Rich.) R.D.Webster (palisade grass)] caused reductions of approximately 42% in dry matter and 47% in eucalyptus leaf area. In the present study, three D. insularis m−2 were necessary for the eucalyptus dry matter to be reduced by 46% (Table 2). This difference between the previously cited studies is possibly due to the different competitive capacities intrinsic to the weed species, which can cause physiological changes in the eucalyptus, resulting in a decrease in plant growth. Such results are in agreement with those of Santos et al. (2015), who observed that the coexistence of eucalyptus with grasses resulted in decreases in stomatal conductance, transpiration rate and water use efficiency of E. urograndis. In fact, the increase in the number of specimens competing for the environmental resources (according to the increase in weed density) may result in a low supply of essential resources for eucalyptus growth (Adams et al. 2003; Bacha et al. 2016; Costa et al. 2021), causing deficiencies that result in changes of some parameters related to photosynthesis (Huang et al. 2008), like low intensity or quality of incident light (Red/Far red ratio) (Sharkey and Raschke 1981; Ma and Upadhyaya 2017) and water deficit (Grossnickle and Folk 1993; Lamhamedi et al. 1998; Garau et al. 2008). On the other hand, Galvan-Cisneros et al. (2023) emphasized that old eucalyptus plantations can cause increased environmental filtering (by influencing shading, soil water availability and soil nutrients), which also influences the species composition of neighboring vegetation, given the different competitive abilities among different functional vegetation types (woody, grasses and herbaceous species), evidencing that such ecological interaction has a mutual influence on competing species.
Comparing the planting conditions of both experiments, considering the means of all densities and distances, it was observed that for D. insularis seeded (Experiment 1), the reductions of TDM and LDM were 45% and 41%, respectively; whereas for regrowth (Experiment 2), the values reached 85.7% and 83.5% of reduction for the same variables (Tables 2 and 4, respectively). Such results demonstrated that the reduction rates of both characteristics were greater in the regrowth condition, corroborating those of Bacha et al. (2016). Those authors elucidated that U. decumbens competed for nutrients more effectively in regrowth, possibly because the weed roots were already fully developed when E. urograndis was planted, which may have conferred a competitive advantage to the weed, especially regarding nutrient absorption. Nitrogen is a key nutrient for chlorophyll formation, and its availability in soil may directly affect leaf chlorophyll concentration (Donahue et al. 1990), as well as tree growth under competition (Adams et al. 2003). This statement can be illustrated by the highest values of total chlorophyll concentration obtained by control plants not subjected to coexistence (Table 4). As highlighted by several studies (Porra et al. 1989; Chappelle and Kim 1992; Ripullone et al. 2003), the amount of chlorophyll is directly related to the photosynthetic rate and consequently, plant growth, supporting the results observed in this study (Figs. 2 and 3). For E. globulus, Adams et al. (2003) demonstrated that nitrogen fertilization lessen the negative effect from coexistence with grasses, suggesting that the availability and the competition for nitrogen were limiting tree growth.
The four groups formed on PCA analysis showed considerable distinction among themselves, in which the regrowth experiment indicated the most severe impacts on growth of planted eucalyptus, regardless of density and distance. This response is due to the fact that the D. insularis plants already had their root system fully established when the eucalypt was planted, which possibly guaranteed them a greater competitive advantage. In contrast to the regrowth group, the weeds’ first growth also formed an isolated group, but directly related to PC1 and also to the evaluated variables (Fig. 3). Thus, regardless of density and distance, the weed in regrowth clearly caused greater interference in eucalyptus growth, corroborating the results obtained by Bacha et al. (2016), who evaluated the competition between E. urograndis (clone GG-100) and another common brazilian grass weed species (U. decumbens) in a short-term response.
With respect to D. insularis resistance to glyphosate, several authors reported that when an allele of resistance to herbicides confers an adaptation-cost to the plant, these biotypes tend to present less competitive aggressiveness. As a result, the limitation in herbicide-binding to the site of action, caused by a structural modification in the target enzyme, can compromise the efficiency of the enzymatic function, resulting in the cost of resistance (or “fitness”) (Powles and Yu 2010; Vila-Aiub et al. 2011, 2015). However, as this type of comparison between resistant and susceptible biotypes was not carried out in this study, such information cannot be conclusive based on what was exposed here. Despite this, the possibility is suggested that biotypes of D. insularis susceptible to glyphosate present an even greater competitive capacity than those observed in this work (Yanniccari et al. 2016; Braga et al. 2018; Barroso et al 2022). However, other studies should be carried out in this sens in order to confirm such hypothesis.
In the present study, the eucalyptus variable most affected by the weed’s first growth interference was the SDM (Table 2); the D. insularis planted at a density of 3 plants m−2 reduced the eucalyptus SDM by 45%, compared to the control, regardless of the distance (Table 2). Graat et al. (2015), studying increasing distances between signalgrass (U. decumbens) and eucalyptus, also reported that the characteristic that most affected by competition was SDM, with reductions of approximately 51%. Similarly, several studies with other weed species [Panicum maximum Jacq. (Guinea grass) (Dinardo et al. 2003), Commelina benghalensis L. (Bengal dayflower) (Costa et al. 2021) and U. brizantha (Colmanetti et al. 2017)] have elucidated that the stem dry mass is the variable most sensitive to interference imposed by weeds. On the other hand, the eucalyptus height was the characteristic that showed less sensitivity to the effects of competition imposed by D. insularis in regrowth (Table 4). These results corroborate those obtained in several studies, with different eucalyptus clones and grasses weed species (Toledo et al. 2001; Cruz et al. 2010; Braga et al. 2018; Tiburcio et al. 2023). Thus, the information that eucalyptus height is the least sensitive characteristic to interference imposed by weeds (and SDM is the most sentsitive) is essential for eucalyptus producers, since the non-destructive evaluations commonly performed throughout the crop cycle in commercial areas (e.g., height of eucalyptus plants) may demonstrate interference values lower than those observed for real productivity (in wood mass). Taken togheter, these results indicate that adequate control of the weed community must be paramount, since the real loss in biomass will possibly be greater than those indicated by the height assessment (Tiburcio et al. 2023).
Conclusions
The coexistence with Digitaria insularis plants, regardless of density and distance, had a negative effect on the Eucalyptus urophylla × E. grandis (Clone AEC-144) early growth.
After cutting, the second growth (regrowth condition) of D. insularis interfered more aggressively in the early growth of eucalyptus plants, compared to the weed’s first growth (seeded), possibly due to the root system already established at the moment of eucalyptus planting.
Due to D. insularis competitive aggressiveness as well as glyphosate resistance, it is important that forest growers consider its management with pre-emergence herbicides. In case of not-suficient pre-emergence control (due to high infestation, and/or also due to a possible herbicide’s short residual effect, considering eucalyptus as a perennial crop), the use of post-emergence herbicides with different mechanisms of action must be carried out (e.g., graminicides utilization). As a result, these management strategies will may avoid weed regrowth, as well as ensure that these plants do not spread seeds, avoiding the increasing areas with glyphosate-resistant D. insularis.
References
Adams P, Beadle C, Mendham N, Smethurst P (2003) The impact of timing and duration of grass control on the growth of a young Eucalyptus globulus Labill. plantation. New for 26:147–165. https://doi.org/10.1023/A:1024490707175
ALAM – Asociación Latinoamericana De Malezas (1974) Recomendaciones sobre unificación de los sistemas de evaluación en ensayos de control de malezas. Asociación Latinoamericana De Malezas 1:35–38
Bacha AL, Martins PFRB, Alves PLCA, Paula RC (2018) Trinexapac-ethyl causes stimulatory effect on eucalyptus initial growth under nutritional deficiency. Can J for Res 48(1):94–100. https://doi.org/10.1139/cjfr-2017-0245
Bacha AL, Pereira FCM, Pires RN, Nepomuceno MP, Alves PLCA (2016) Interference of seeding and regrowth of signal grass weed (Urochloa decumbens) during the initial development of Eucalyptus urograndis (E. grandis × E. urophylla). Aust J Crop Sci. 10(3):322–330. https://doi.org/10.21475/ajcs.2016.10.03.p6995
Barroso AAM, Michelon TB, Alves PLCA, Han H, Yu Q, Powles SB, Vila-Aiub MM (2022) Challenging glyphosate resistance EPSPS P106S and TIPS mutations with soybean competition and glyphosate: implications for management. Pest Manag Sci 78(11):4764–4773. https://doi.org/10.1002/ps.7096
Barroso GM, da Silva RS, Mucida DP, Borges CE, Ferreira SR, dos Santos JCB, Lins HA, Mendonça V, Silva DV, dos Santos JB (2021) Spatio-temporal distribution of Digitaria insularis: risk analysis of areas with potential for selection of glyphosate-resistant biotypes in eucalyptus crops in Brazil. Sustainability 13:10405. https://doi.org/10.3390/su131810405
Blackshaw RE, Harker KN (2002) Selective weed control with glyphosate in glyphosate-resistant spring wheat (Triticum aestivum). Weed Technol 16:885–892. https://doi.org/10.1614/0890-037X(2002)016[0885:SWCWGI]2.0.CO;2
Bleasdale JKA (1960) Studies on plant competition. In: Harper JL (ed) The biology of weeds. Oxford, pp 133–142
Bleiholder H, Kirfel H, Langelüddecke P, Stauss R (1991) Codificação unificada dos estádios fenológicos de culturas e ervas daninhas. Pesq Agropec Bras 26(9):1423–1429
Braga AF, Barroso AAM, Amaral CL, Nepomuceno MP, Alves PLCA (2018) Population interference of glyphosate resistant and susceptible ryegrass on eucalyptus initial development. Planta Daninha 36:e018170148. https://doi.org/10.1590/S0100-83582018360100086
Caldwell J, Sucoff E, Dixon K (1995) Grass interference limits resource availability and reduces growth of juvenile red pine in the field. New for 10:1–15. https://doi.org/10.1007/BF00034173
Carvalho LB, Cruz-Hipolito H, González-Torralva F, Alves PLCA, Christoffoleti PJ, Prado R (2011) Detection of sourgrass (Digitaria insularis) biotypes resistant to glyphosate in Brazil. Weed Sci 59(2):171–176. https://doi.org/10.1614/WS-D-10-00113.1
Cerdeira AL, Gazziero DLP, Duke SO, Matallo MB (2011) Agricultural impacts of glyphosate-resistant soybean cultivation in South America. J Agric Food Chem 59(11):5799–5807. https://doi.org/10.1021/jf102652y
Cesarin AE, Martins PFRB, Barroso AAM, Carrega WC, Alves PLCA, Bianco S (2019) Dry matter and macronutrient accumulation in Digitaria insularis biotypes susceptible and resistant to glyphosate. J Agric Sci 11(4):151–159. https://doi.org/10.5539/jas.v11n4p151
Chappelle EW, Kim MS (1992) Ratio analysis of reflectance spectra (RARS): an algorithm for the remote estimation of the concentrations of chlorophyll a, chlorophyll b, and carotenoids in soybean leaves. Remote Sens Environ 39(3):239–247. https://doi.org/10.1016/0034-4257(92)90089-3
Christoffoleti PJ, López-Ovejero R (2003) Main aspects on weed herbicide resistance to glyphosate. Planta Daninha 21(3):507–515. https://doi.org/10.1590/S0100-83582003000300020
Clapp J (2021) Explaining growing glyphosate use: the political economy of herbicide-dependent agriculture. Glob Environ Chang 67:102239. https://doi.org/10.1016/j.gloenvcha.2021.102239
Coll L, Balandier P, Picon-Cochard C (2004) Morphological and physiological responses of beech (Fagus sylvatica) seedlings to grass-induced belowground competition. Tree Physiol 24(1):45–54. https://doi.org/10.1093/treephys/24.1.45
Colmanetti MAA, Bacha AL, Alves PLCA, Paula RC (2019) Effect of increasing densities of Urochloa brizantha cv. Marandu on Eucalyptus urograndis initial development in silvopastoral system. J for Res 30:537–543. https://doi.org/10.1007/s11676-018-0635-4
Colmanetti MAA, Bacha AL, Braga AF, Alves PLCA, Paula RC (2017) Interference of Urochloa decumbens and Panicum maximum in the initial growth of six clones of Eucalyptus urograndis. Aust J Crop Sci 11(10):1261–1267. https://doi.org/10.21475/ajcs.17.11.10.pne514
Costa AGF, Bacha AL, Pires RN, Pavani MCMD, Alves PLCA (2021) Interference of Commelina benghalensis in the initial growth of Eucalyptus grandis in winter and summer. Ciência Florestal 31(2):590–606. https://doi.org/10.5902/1980509825556
Cruz MB, Alves PLCA, Karam D, Ferraudo AS (2010) Capim-colonião e seus efeitos sobre o crescimento inicial de clones de Eucalyptus × urograndis. Ciência Florestal 20(3):391–401. https://doi.org/10.5902/198050982054
Dinardo W, Toledo REB, Alves PLCA, Pitelli RA (2003) Efeito da densidade de plantas de Panicum maximum. Jacq. Sobre o crescimento inicial de Eucalyptus grandis W. Hill ex Maiden. Scientia Forestalis 64:59–68
Donahue RA, Berg VS, Vogelmann TC (1990) Assessment of the potential of the blue light gradient in soybean pulvini as a leaf orientation signal. Physiol Plant 79(4):593–598. https://doi.org/10.1111/j.1399-3054.1990.tb00030.x
Fisher RA (1925) Statistical methods for research workers. Oliver & Boyd, Edinburgh
Florentine S, Fox J (2003) Competition between Eucalyptus victrix seedlings and grass species. Ecol Res 18:25–39. https://doi.org/10.1046/j.1440-1703.2003.00531.x
Galvan-Cisneros CM, Gastauer M, Massante C, Villa PM, Meira-Neto JAA (2023) Simultaneous competition and environmental filtering in woody communities of the understory of Eucalyptus plantations in the Cerrado. Perspect Plant Ecol Evol Syst 59:12573. https://doi.org/10.1016/j.ppees.2023.125731
Garau AM, Ghersa CM, Lemcoff JH, Barañao JJ (2009) Weeds in Eucalyptus globulus subsp. maidenii (F. Muell) establishment: effects of competition on sapling growth and survivorship. New for 37:251–264. https://doi.org/10.1007/s11056-008-9121-8
Garau AM, Lemcoff JH, Ghersa CM, Beadle CL (2008) Water stress tolerance in Eucalyptus globulus Labill. subsp. maidenii (F. Muell.) saplings induced by water restrictions imposed by weeds. Forest Ecol Manag 255:2811–2819. https://doi.org/10.1016/j.foreco.2008.01.054
Gonçalves JLM, Alvares CA, Higa AR, Silva LD, Alfenas AC, Stahl J, Ferraz SFB, Lima WP, Brancalion PHS, Hubner A, Bouillet JPD, Laclau JP, Nouvellon Y, Epron D (2013) Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalyptus plantations. For Ecol Manag 301:6–27. https://doi.org/10.1016/j.foreco.2012.12.030
Graat Y, Bacha AL, Nepomuceno MP, Alves PLCA (2018) Initial development of eucalyptus according to different desiccation periods of signalgrass. Planta Daninha 2018(v36):e018145168. https://doi.org/10.1590/S0100-83582018360100020
Graat Y, Rosa JO, Nepomuceno MP, Carvalho LB, Alves PLCA (2015) Grass weeds interfering with eucalyptus: effects of the distance of coexistence on the initial plant growth. Planta Daninha 33:203–211. https://doi.org/10.1590/0100-83582015000200005
Grossnickle S, Folk R (1993) Stock quality assessment: forecasting survival and performance on a reforestation niche. Tree Plant Notes 44(3):113–121
Heap I (2023) The international survey of herbicide resistant weeds. www.weedscience.com. Accessed 5 March 2023
Heap I, Duke SO (2018) Overview of glyphosate-resistant weeds worldwide. Pest Manag Sci 74:1040–1049. https://doi.org/10.1002/ps.4760
Hébert MP, Fugère V, Gonzalez A (2019) The overlooked impact of rising glyphosate use on phosphorus loading in agricultural watersheds. Front Ecol Environ 17(1):48–56. https://doi.org/10.1002/fee.1985
Huang Z, Xu Z, Blumfield TJ, Bubb K (2008) Variations in relative stomatal and biochemical limitations to photosynthesis in a young blackbutt (Eucalyptus pilularis) plantation subjected to different weed control regimes. Tree Physiol 28:997–1005. https://doi.org/10.1093/treephys/28.7.997
Hunt MA, Battaglia M, Davidson NJ, Unwin GL (2006a) Competition between plantation Eucalyptus nitens and Acacia dealbata weeds in northeastern Tasmania. Forest Ecol Manag 233:260–274. https://doi.org/10.1016/j.foreco.2006.05.017
Hunt MA, Battaglia M, Davidson NJ, Unwin GL (2006b) Competition between plantation Eucalyptus nitens and Acacia dealbata weeds in northeastern Tasmania. For Ecol Manag 233:260–274. https://doi.org/10.1016/j.foreco.2006.05.017
Ibá (2022) Indústria brasileira de árvores. Relatório Ibá. https://iba.org/datafiles/publicacoes/relatorios/relatorio-anual-iba2022-compactado.pdf. Accessed 5 March 2023
Köppen W (1948) Climatologia: con un estudio de los climas de la tierra. Fondo de Cultura Econômica, México
Lamhamedi M, Bernier P, Hebert C, Jobidon R (1998) Physiological and growth responses of three sizes of containerized Picea mariana seedlings outplanted with and without vegetation control. For Ecol Manag 110:13–23
Little KM, Rolando CA, Morris CD (2007) An integrated analysis of 33 Eucalyptus trials linking the onset of competition-induced tree growth suppression with management, physiographic and climatic factors. Ann for Sci 64:585–591. https://doi.org/10.1051/forest:2007036
Little KM, Staden JV (2005) Effects of vegetation control on Eucalyptus grandis × E. camaldulensis volume and economics. S Afr J Bot 71:418–425
Ma L, Upadhyaya MK (2017) Effects of red/far-red light ratio on common lamb’s-quarters, redroot pigweed, and tomato plants. Can J Plant Sci 97(3):494–500. https://doi.org/10.1139/cjps-2016-0089
Netto AG, Cordeiro EM, Nicolai M, Carvalho SJ, Ovejero RFL, Brunharo CA, Zucchi MI, Christoffoleti PJ (2021) Population genomics of Digitaria insularis from soybean areas in Brazil. Pest Manag Sci 77:5375–5381. https://doi.org/10.1002/ps.6577
Pavan BE, Amaral RG, Pupin S, Costa RML, Dias DC, Scarpinati EA, Paula RC (2021) Competitive ability among Eucalyptus spp. commercial clones in Mato Grosso do Sul state. Forest Ecol Manag 494:119297. https://doi.org/10.1016/j.foreco.2021.119297
Pereira FCM, Alves PLCA, Martins JVF (2013) Interference of grasses on the growth of eucalyptus clones. J Agric Sci 5:173–180. https://doi.org/10.5539/jas.v5n11p173
Pereira FCM, Yamauti MS, Alves PLCA (2012) Interaction between weed management and covering fertilization in the initial growth of Eucalyptus grandis x E. urophylla. Revista Árvore 36:941–950. https://doi.org/10.1590/S0100-67622012000500016
Pitelli RA (1985) Interferência das plantas daninhas em culturas agrícolas. Informe Agropecuário 11(129):16–27
Pitelli RA, Marchi SR (1991) Interferência das plantas invasoras nas áreas de reflorestamento. III Seminário técnico sobre plantas daninhas e o uso de herbicidas em reflorestamento, Belo Horizonte, pp 1–11
Porra RJ, Thompson WA, Kriedemann PE (1989) Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim Biophys Acta Bioenerg 975(3):384–394. https://doi.org/10.1016/S0005-2728(89)80347-0
Powles SB, Yu Q (2010) Evolution in action: plants resistant to herbicides. Annu Rev Plant Biol 61:317–347. https://doi.org/10.1146/annurev-arplant-042809-112119
Qin F, Liu S, Yu S (2018) Effects of allelopathy and competition for water and nutrients on survival and growth of tree species in Eucalyptus urophylla plantations. For Ecol Manag 424:387–395. https://doi.org/10.1016/j.foreco.2018.05.017
Resende RT, Marcatti GE, Pinto DS, Takahashi EK, Cruz CD, Resende MDV (2016) Intra-genotypic competition of Eucalyptus clones generated by environmental heterogeneity can optimize productivity in forest stands. For Ecol Manag 15:50–58. https://doi.org/10.1016/j.foreco.2016.08.041
Ripullone F, Grassi G, Lauteri M, Borghetti M (2003) Photosynthesis–nitrogen relationships: interpretation of different patterns between Pseudotsuga menziesii and Populus× euroamericana in a mini-stand experiment. Tree Physiol 23(2):137–144. https://doi.org/10.1093/treephys/23.2.137
Sands R, Nambiar EKS (1984) Water relations of Pinus radiata in competition with weeds. Can J Forest Res 14:233–237. https://doi.org/10.1139/x84-045
Santos MV, Ferreira EA, Tuffi Santos LD, da Fonseca DM, Aspiazu I, Silva DV, Porto JMP, Braga RR (2015) Physiological aspects of acacia and eucalyptus in competition with Brachiaria. Aust J Crop Sci 9(3):210–214
Schaller M, Schroth G, Beer J, Jimenez F (2003) Root interactions between young Eucalyptus deglupta trees and competitive grass species in contour strips. For Ecol Manage 179:429–440. https://doi.org/10.1016/S0378-1127(02)00534-0
Sharkey TD, Raschke K (1981) Effect of light quality on stomatal opening in leaves of Xanthium strumarium L. Plant Physiol 68(5):1170–1174. https://doi.org/10.1104/pp.68.5.1170
Silva JRV, Alves PLCA, Toledo REB (2012) Weed control strip influences the initial growth of Eucalyptus grandis. Acta Sci-Agron 34(1):29–35. https://doi.org/10.4025/actasciagron.v34i1.12252
Sneath PH, Sokal RR (1973) Numerical taxonomy: the principles and practice of numerical classification. Freeman, San Francisco, WH
Stape JL, Binkley D, Ryan MG, Gomes AND (2004) Water use, water limitation, and water use efficiency in a Eucalyptus plantation. Bosque 25(2):35–41. https://doi.org/10.4067/S0717-92002004000200004
Tarouco CP, Agostinetto D, Panozzo LE, Santos LS, Vignolo GK, Ramos LOO (2009) Weed interference periods on in the initial growth of eucalyptus. Pesq Agrop Brasileira 44(9):1131–1137. https://doi.org/10.1590/S0100-204X2009000900010
Tiburcio RAS, Bacha AL, Alves PLCA, Salgado TP (2023) Long-term growth response to weed-control strips in Eucalyptus urograndis plantations in Brazil. Aust for 86(1):22–30. https://doi.org/10.1080/00049158.2023.2167155
Toledo REB, Victória Filho R, Alves PLCA, Pitelli RA, Cadini MTD (2000) Effects of Brachiaria decumbens strip control on eucalyptus initial plants development. Planta Daninha 18(3):383–393. https://doi.org/10.1590/S0100-83582000000300001
Toledo REB, Victória Filho R, Alves PLCA, Pitelli RA, Cadini MTD (2001) Efeito da densidade de plantas de Brachiaria decumbens Stapf. sobre o crescimento inicial de mudas de Eucalyptus grandis W. Hill ex Maiden. Scientia Forestalis 60:109–117
Vila-Aiub MM, Gundel PE, Preston C (2015) Experimental methods for estimation of plant fitness costs associated with herbicide-resistance genes. Weed Sci 63:203–216. https://doi.org/10.1614/WS-D-14-00062.1
Vila-Aiub MM, Neve P, Roux F (2011) A unified approach to the estimation and interpretation of resistance costs in plants. Heredity 107(5):386–394. https://doi.org/10.1038/hdy.2011.29
Watt MS, Whitehead D, Mason EG, Richardson B, Kimberley MO (2003) The influence of weed competition for light and water on growth and dry matter partitioning of young Pinus radiata, at a dryland site. Forest Ecol Manag 183:363–376. https://doi.org/10.1016/S0378-1127(03)00139-7
Yanniccari M, Vila-Aiub M, Istilart C, Acciaresi H, Castro AM (2016) Glyphosate resistance in perennial ryegrass (Lolium perenne L.) is associated with a fitness penalty. Weed Sci 64(1):71–79. https://doi.org/10.1614/WS-D-15-00065.1
Acknowledgements
The authors are grateful for the technical support provided by Weed Sciences Laboratory (LAPDA) and also to the financial support provided by São Paulo Research Foundation (FAPESP) (grant #20/06162-6) to the first author.
Funding
São Paulo Research Foundation (FAPESP) (grant #20/06162-6).
Fundação de Amparo à Pesquisa do Estado de São Paulo, 20/06162-6
Author information
Authors and Affiliations
Contributions
Study conception and design: T.B.G., A.L.B. and P.L.C.A.A.; Methodology implementation, experiment execution and data collection: T.B.G. and A.L.B.; Data analysis/interpretation: T.B.G., A.L.B. and P.L.C.A.A.; Manuscript writing/revision: T.B.G., A.L.B., H.L.M., B.D.N. and P.L.C.A.A.
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that there is no conflict of interest regarding the publication of this manuscript.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Guidugli, T.B., Bacha, A.L., Martins, H.L. et al. Glyphosate-resistant Digitaria insularis effect on eucalyptus initial growth. New Forests 55, 1209–1229 (2024). https://doi.org/10.1007/s11056-024-10028-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-024-10028-y