
Abstract
Plant functional traits have been shown to vary with environmental conditions. However, we lack empirical data on how plant functional traits of different plant life forms respond to environmental factors. We studied the influence of environmental conditions on the distribution of plant functional traits in a Quercus wutaishanica forest with the aim of exploring the patterns of functional traits across different life forms and determining the driving factors of functional trait variation at fine spatial scales. We collected data on environmental factors (soil nutrients and soil moisture, canopy variables, topography) of 70 20 m × 20 m plots. Leaves were harvested from 26 species (4 tree species, 7 shrub species and 15 herbaceous species), and community-weighted mean (CWM) trait values for leaf area (LA), leaf mass per unit area (LMA), leaf carbon content (LCC), leaf nitrogen content (LNC), leaf phosphorus content (LPC), and leaf potassium content (LKC) were calculated. We also measured the height (H) of plants. The importance of biotic and abiotic factors in controlling plant functional traits was quantitatively assessed using redundancy analysis (RCA). Regression was used to determine relationships between CWM trait values and environment variables. We found that plant functional traits varied with life forms of plants. LA, LMA and LCC of trees were greater than those of the understory, whereas LNC, LPC and LKC of herbaceous were greater than those of trees. Responses of plant functional traits to environmental factors varied with different life forms. The combined effect of canopy, topography and soil factors had a greater impact on plant functional traits in understory layer than tree layer. General linear models showed that openness is the main factor affecting various functional traits of undergrowth plants, and the relationship between the element content in leaves of herbaceous plants and environmental factors is greater than that of shrub plants. The strong correlation of plant functional traits and environmental factors at fine spatial scales indicates that Q. wutaishanica forests have high spatial variability. Considering the variation of traits in different life form of plants and their interactions with biotic factors, it provides further insights into ecological mechanisms of shaping plant communities and driving plant community dynamics.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Plant functional traits considered as reflecting adaptations to variation in the abiotic and biotic environment (de Bello et al. 2010), can predict the response of important ecosystems to environmental change, and have a significant impact on the ecosystem processes (Cornelissen et al. 2003). In forest ecosystems, variations in plant functional traits along with abiotic and biotic factors determine the function of forest ecosystems (i.e., productivity or biomass) (Prado-Junior et al. 2016). Life forms of plants are very important for determining the gradients of plant functional traits, such as leaf size (Barboni et al. 2004). Different plant life forms have different plant functional traits, thus maximizing the utilization of resources and reducing adverse environmental impacts (Barboni et al. 2004; McIntyre et al. 1999). As a means to maximize the utilization of resources in specific environment, plants in forest communities present obvious vertical stratification. The tree layer is the dominant feature of forests, which controls the appearance of the forest community, affecting the micro-environment of forest and the distribution of understory vegetation (e.g., shrubs and herbs) (Lhotka and Loewenstein 2008; Parker and Dey 2008; Rodriguez-Calcerrada et al. 2008). Understory vegetation can restrict forest tree regeneration (Kobayashi et al. 2004), affect soil temperature and soil processes (e.g., the decomposition of organic matter and nitrogen mineralization) (Matsushima and Chang 2007), and influence forest soil properties and soil microbial biomass (Zhao et al. 2013, 2011). A deep understanding of the potential ecological mechanisms of the relationship between plant functional traits across forest strata (i.e., overstorey and understory) is crucial to understanding the importance of vertical stratification on ecosystem function in natural forests. Recent work has been paid to the relationships between functional traits of overstorey plant traits and environmental factors at large spatial scales (Kraft et al. 2008; Kunstler et al. 2016). The relationships between understory plant traits and environments have received less attention, especially the neglect of herbaceous species, which may have a great impact on forest dynamics (George and Bazzaz 2003).
Plant functional traits lie at the core of plant-environment interactions. Plants can improve the adaptability of population by regulating the utilization of individual resources (Kraft et al. 2008; Violle et al. 2007), thus affecting the function of ecosystem (Dı́az and Cabido 2001). For instance, plants preferentially assign new photosynthetic products to their aboveground parts in order to adapt to light environment (Wilsey 1996). Species with high leaf mass per unit area (LMA) appear in habitats with limited water and/or nutrients (Poorter et al. 2009). Some studies have shown that plant functional traits are related to physical and chemical properties of the soil (Hinsinger 2001; Ding et al. 2011; Laughlin et al. 2015; Coelho et al. 2018). Plant functional traits can also be strongly influenced by water (Chaves et al. 2003; Poorter et al. 2009) and light (Onoda et al. 2008; Valladares et al. 2000). Topography controls the spatial redistribution of light, heat, water and soil nutrients through geomorphic processes and morphological changes, and can greatly affect plant functional traits in forests (Bennie et al. 2008; Zhu et al. 2016). In addition, canopy influences the microclimate under the forest and changes the growth conditions of vegetation, thus affecting the growth, regeneration and species composition of understory vegetation (Barbier et al. 2008; Ellsworth and Reich 1993). The distribution, size, and orientation of leaves determines the pattern of light availability within the canopy and has an important influence on the understory, due to canopy trees competing with the understory for resources, both above- (e.g., light) and belowground (e.g., water and nutrients) (Ellsworth and Reich 1993; Yu and Sun 2013). Therefore, plant functional traits are related to canopy structure. The relationship between plant functional traits and the environment provides a foundation for studying plant growth strategies and ecosystem functions, under existing changing environmental conditions (Koide et al. 2014; Meng and Wang 2007; Schweiger et al. 2017).
Quercus wutaishanica is a dominant tree species in warm temperate deciduous broad-leaved forest area in China (Chen et al. 1999). Taiyue Mountain is a typical habitat for Q. wutaishanica, which is of great significance for maintaining the stability of the ecosystem in Shanxi Province (Yu and Sun 2013). To consider the functional strategies, it is therefore essential to gain insights into the relationship between plant functional traits and environment across forest strata (i.e. overstorey and understory). We postulated that plant functional traits differ among plant life forms. In addition, understanding the relationship between plant functional traits and environmental factors in different plant life forms is of great significance for forest management under heterogeneous vegetation. To address this need, we explored the variations on plant functional traits across different plant life forms and its relationships with environmental factors at fine spatial scales in a temperate deciduous forest. Specially, the objectives of this study were to: (1) what are the characteristics of plant functional traits in Q. wutaishanica forest? How did the plant functional traits vary, and how did it change across different plant life forms? (2) How did soil properties and canopy variables change along the spatial heterogeneity? (3) How do soil properties, canopy variables, and topographic factors affect the variations of plant functional traits at fine spatial scales?
Materials and methods
Study site
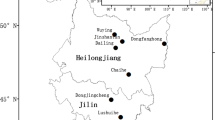
The study area was located in the Lingkong Mountain Nature Reserve (112°5′38″E, 36°39′24″N) of Taiyue Mountain, Shanxi Province, China. The Q. wutaishanica forest in the Reserve is typical of forests ranging from northeast to southwest China, with Q. wutaishanica occurring in most of its natural range (Yu and Sun 2013). Elevation ranges from 1450 to 1928 m. Mean annual temperature is 6.2 °C, and mean total annual precipitation is about 700 mm, with 60%—70% occurring in the summer (from July to September) (Cheng et al. 2014). The soil is identified as Cinnamon, which matches the alfisol type in the U.S. soil classification system (Lin et al. 2002; Yu and Sun 2013). The predominant tree species in the Reserve are Q. wutaishanica and Pinus tabuliformis, which are typical and frequently occurring temperate tree species of northern China. Typical shrub species include Lespedeza bicolor, Rosa xanthina, Corylus heterophylla, Spiraea trilobata. The most common herbaceous species are Echinochloa crusgalli, Melampyrum roseum, Carex duriuscula and Phlomis umbrosa.
Vegetation surveys
In August 2010, a permanent plot covering an area of 2.8 ha (200 m × 140 m) was established on a hillside in a secondary forest (Fig. 1). According to historical records, local forests have been frequently disturbed since the 1950s, including small-scale logging, which promoted the regeneration of P. tabuliformis and Larix principis-rupprechtii, and periodic fires. Since 1998, these forests have been protected as part of the National Natural Forest protection Project, but since then, no forest management measures have been taken for forests in this region. The existing forests are composed of Q. wutaishanica, Pinus tabuliformis and L. principis-rupprechtii with a certain proportion of pioneer species (Betula platyphylla).

Location and topography of the study site. DEM: Digital elevation model; The right on the right is a permanent plot, covering an area of 2.8 ha (200 m × 140 m) and divided into 70 quadrats of 20 m × 20 m
Filed surveys and measurements were carried out following the plot established. The plot was subdivided into 70 quadrats of 20 m × 20 m. Topographic attributes of each quadrats, including slope and aspect, were measured according to the method described in Liu et al. (2014). Position on slope was converted to numerical value of 1, 2, 3, and 4 for ridge top, upper-, mid-, and down-slope, respectively. All trees with a diameter at breast height (DBH; measured at 1.3 m above the soil) at least 1 cm and height were measured, mapped, and identified to species (Cheng et al. 2014). In August 2015, the DBH and height of trees (DBH > 1 cm) were re-measured. Additionally, five 5 m × 5 m shrub quadrats and five 1 m × 1 m herbaceous quadrats were set up within each quadrat of 20 m × 20 m. Within each quadrat, we recorded the height, occurrence, richness and coverage of each shrub and herbaceous plant.
Soil sampling and abiotic variables
In each quadrat, five independent soil samples with a depth of 0–20 cm were collected by using a circular soil auger with a diameter of 4.8 cm. Then, these samples were mixed within the same depth to analyze soil moisture, pH and soil nutrients. To measure soil moisture (M), 50 g of the soil was weighed and dried at 105℃ for 24 h, after removing stones, plant roots and litter. The remaining soil was air-dried and then sieved with a 1-mm mesh screen. 20 g of the 1 mm filtered soil was used to estimate pH with an acidity meter, and 50 g was filtered again with a 0.25 mm sieve to analyze soil organic carbon (SOC), soil total nitrogen (STN), soil total potassium (STK), and soil total phosphorus (STP). Samples were weighed and acidified by 25% HCl to remove inorganic carbon from the soil (Nieuwenhuize et al. 1994). SOC and STN were measured using an elemental analyzer (Thermo Fisher Flash 2000, the United States). STP was determined by digestion with perchloric acid and then measured by colorimetry (Sommers and Nelson 1972). STK was measured by digestion with hydrofluoric acid and perchloric acid (Pratt 1965).
Canopy variables measurements
In August 2015, samples were collected by using a canopy analyzer (WinCANOPY 2010a, Regent Instrument Inc., Quebec, Canada) for measuring canopy variables. Nine photos were taken randomly in each quadrat of 20 m × 20 m. We set the instrument at a height of 1.3 m. In order to avoid the influence of direct sunlight and scattering on canopy imaging, the photos were taken in cloudy weather between 8:00 to 10:00 am and 2:00 to 4:00 pm. The leaf area index (LAI), mean leaf angle (MLA), photosynthetic photon flux density above and below canopy (PPFD Over and PPFD Under), and openness were calculated by the canopy analyzer.
Plant functional traits
We sampled all individuals belonging to 97 species and 34 families. Based on results of the importance value (IV) index, 26 species within the 70 quadrats of 20 m × 20 m were chosen for measuring various functional traits to get community-weighted means of trait values (CWM), including 4 tree species, 7 shrub species and 15 herbaceous species (IV index ≥ 0.05) (Table S1). The species selected accounted for 79% (ranging from 68%—91%) individuals on average in a plot. The traits were calculated from field measurements of fully expanded fresh young and undamaged leaves of adult plants (Lebrija-Trejos et al. 2010). At least five individuals of comparable size per species were selected (tree ≥ 5 individuals; shrub ≥ 5 individuals; herbaceous ≥ 10 individuals), and five leaves per individual were sampled. Sampled tree leaves came from the outer layer of the crown, in the middle of the vertical length (Lebrija-Trejos et al. 2010). To prevent leaf wilting and the closing of compound leaves after collection, leaves were collected with their branches and immediately transported to the laboratory in plastic bags. The branches were put in water, and the leaves were detached individually to record their fresh mass and dimensions. Leaf traits were measured on entire leaves (without petioles). We measured leaf area (LA), leaf mass per unit area (LMA), leaf carbon content (LCC), leaf nitrogen content (LNC), leaf phosphorous content (LPC), and leaf potassium content (LKC) for each species in each plot. LA was calculated using a leaf area meter (LI-3000C Area Meter, Li-COR, Lincoln, USA). Leaves were oven dried for 48 h at 85 °C to obtain dry mass. After milling and sieving (through a 0.25 mm sieve), dry leaf samples were analyzed for leaf stoichiometric traits at the Key Laboratory for Silviculture and Conservation. LCC and LNC were measured by using an elemental analyzer (Thermo Fisher Flash 2000, USA). LPC and LKC were analyzed similarly to the soils.
Data analysis
The descriptive statistics (frequency distributions, means, and standard deviations) for each soil properties and canopy variables were calculated by using SPSS 18.0 software (SPSS InC., Chicago, IL, USA). The Kolmogorov–Smirnov test (K-S test) was used to determine if the frequency distributions departed significantly from normality. The spatial correlation of soil properties was analyzed by using GS + 10 software (Gamma Design Software, Plainwell, Michigan, USA). Three indices, nugget effect, range, and sill, were calculated to describe semi-variograms (Berry 2005). Then the soil properties data were fitted to spherical, exponential, and Gaussian models to obtain the best fit based on the lowest residual sum of squares (RSS). The nugget to sill ratio was divided into three classes: strong spatial correlation (< 25%), moderate spatial correlation (26–75%), and weak spatial correlation (> 75%) (Cambardella et al. 1994; Song et al. 2019; Negassa et al. 2019). All maps were produced using GIS software Arcmap 10.0 (Esri, Redlands, CA) according the residual value of ordinary kriging (Wang et al. 2017).
Community-weighted mean trait values (CWM) were calculated for each plant life form in each quadrat of 20 m × 20 m based on the relative contribution of the species to the community (Lavorel et al. 2008).
where Pi means the relative contribution of species i to the community for which we use the IV indices, and traiti is the trait value of species i.
ANOVAs were performed to evaluate differences among the different plant life forms for each plant functional traits. Bivariate correlation analysis was performed to investigate the relationship between plant functional traits at both the quadrat-level and species-level. These analyses were carried out in SPSS 18.0 software. Redundancy Analysis (RDA; (Rao 1964)) was performed to investigate the relationship between plant functional traits and environmental variables (soil properties, canopy variables, and topographic factors), as well as to identify the relative importance of the effect of each environmental factor on plant functional traits for each plant life form separately (Mcardle and Anderson 2001). The ordination analyses were performed using R software. Statistical significance was tested by the Monte Carlo permutation method based on 499 runs with randomized data (Wang et al. 2016). The multiple stepwise regressions were conducted with plant functional traits as the dependent variable and the environmental factors as independent variables for each plant life form separately. The absolute value of multivariate stepwise regression coefficients represented the impact of the environmental factors on plant functional traits (Bu et al. 2013).
Results
Comparison of plant functional traits for the different life forms
The CWM of plant functional traits for the 70 quadrats showed significant differences among the three plant life forms (Fig. 2). Plant height (H) and LCC were greatest for the tree layer, followed by the shrub layer, and the herbaceous layer, and there were significant differences among the three layers. The LMA and LA values for the tree layer were significantly higher than for the herbaceous layer and the shrub layer, but there were no significant differences between the herbaceous and shrub layers. The values of LNC, LPC for the herbaceous layer were the highest among the three groups, while the tree and shrub layer were not significantly different. The LKC values were higher significantly from the tree layer to the shrub layer to the herbaceous layer. The CWM of six traits (except plant height) tended to vary greatly among different quadrats, especially within the shrub layer.

Community-weighted mean trait values of different plant functional types in a Q. wutaishanica forest in Taiyue Mountain, Shanxi. H, height; LMA, leaf mass per unit area; LA, leaf area; LCC, leaf carbon content; LNC, leaf nitrogen content; LPC, leaf phosphorous content; LKC, leaf potassium content. a community-weighted means trait values of H; a community-weighted means trait values of LMA; b community-weighted means trait values of LA; c community-weighted means trait values of LCC; d community-weighted means trait values of LNC; e community-weighted means trait values of LPC; f community-weighted means trait values of plant height; g community-weighted means trait values of LKC. Boxes with different capital letters indicate significant differences among plant life forms at P < 0.01
At the quadrat-level, pairwise trait correlations were different across the three plant life forms (Table 1). In the tree layer, there were significantly negative correlations between LKC and LA, and between LKC and LCC (P < 0.01), but there were no significant relationships between other traits. In the shrub layer, significant positive correlations were detected among four leaf stoichiometric traits (LCC, LNC, LPC and LKC) (P < 0.01), while LPC and LA showed a significant positive correlation at the 0.05 level. In the herbaceous layer, LNC, LPC and LKC were significantly positively correlated (P < 0.01).
Compared to the quadrat-level, the correlations of plant functional traits at the species-level had higher values. Significant negative correlations were found between H and LPC, LPC and LKC (P < 0.01). On the contrary, significant positive correlations were found between H and LKC, LNC and LPC (P < 0.01) (Table 2). There were significant negative correlations between LA and LCC, and H and LNC (P < 0.05).
Spatial heterogeneity and correlation of soil properties in the Q. wutaishanica forest
As shown in Table 3, the mean value of soil properties was as follows: pH 6.48, SOC 2.62%, STN 2.46%, STP 0.42 mg/g, STK 0.15 mg/g in the surface soil. The CV of STN in the surface soil was particularly very high (176.21%). This suggests a high depends spatial heterogeneity of STN in the study region. Data of pH, SOC, STN, STK, STP were a normally distributed in surface soil layers, which met the requirement for geostatistical analysis.
Semi-variogram models and the best fit model parameters for soil properties are presented in Table S2 and Fig. S1. The scales for the spatial autocorrelations of the five soil properties were different. The nugget-to-sill ratios of the STN, STK, and pH were 0.31%, 8.33%, and 7.20% at surface soil, indicating a strong correlation. The other soil nutrients of SOC and STP showed moderate spatial correlation with nugget to sill ratios of 34.88% and 42.69% (Table S2). The coefficients of determination (R2) ranged from 0.426 to 0.999. The distribution maps of ordinary Kriging show that soil nutrients concentrations were obviously different at a small scale (Figs. S2).
Canopy variables characteristics in the Q. wutaishanica forest
The characteristics of canopy variables including openness, LAI, MLA, PPFD Over, PPFD Under are presented in Table 4. The LAI was in the range of 2.20 to 4.99 with CV of 21.17%. The MLA was in the range of 14.76 to 31.68°. PPFD Over and PPFD Under were 28.93 to 29.07 mol/m2∙day, and 2.55 to 13.95 mol/m2∙day, respectively, and the CV of PPFD Over was the lowest value, whereas the CV of PPFD Under was the highest value. The openness was in the range of 5.95 to 21.01, and its CV was 17.77%. MLA and PPFD Under were significantly correlated with openness (P < 0.01). The LAI was significantly negatively correlated with MLA and PPFD Over (P < 0.05), whereas MLA was significantly positively correlated with PPFD Over (P < 0.01) (Fig. S3).
Relationship between plant functional traits and environmental factors
The following 13 environmental variables were significantly related to plant functional traits across different plant life forms. The first two RDA axes explained 55.91%, 87.08%, 69.01% of the plant functional traits variation in tree layer, shrub layer, and herbaceous layer, respectively (Fig. 3). Canopy variables affecting the functional traits of tree layer plants were PPFD over and MLA (Fig. 3a). Plant functional traits showed profound correlations with PPFD under in the shrub layer and openness in the herbaceous layer (Fig. 3b, c). Plant functional traits of the different plant life forms were significantly correlated with abiotic and biotic factors as shown by stepwise multiple regression analysis (Table 5). In the tree layer, H was significantly negatively influenced by MLA, PPFD Over, STN, aspect and slope, and positively influenced by STK and soil moisture (P < 0.05). Soil moisture had a negative effect on LMA, while aspect had a positive effect on LMA. LA was significantly negatively influenced by stand openness, STK, pH and slope, and positively influenced by MLA and SOC (P < 0.05). LCC was mainly positively influenced by MLA, while LNC was negatively affected by SOC (Table 5). Compared with the tree layer, the functional traits of the understory layer were directly affected by the combine effect of environmental factors and the explanation of variation was obviously greater (Table 5). In the shrub layer, H was positively influenced by stand openness, LAI, and soil moisture, while H was negatively influenced by MLA, PPFD Under, STN, STK, and slope. PPFD Under, pH and slope had a negative effect on LA, while SOC ad soil moisture had a positive effect on LA. Stand openness had a positive effect on LCC, while LAI, MLA, STN, STK, pH had negative effect on LCC. LPC was negatively affected by stand openness, STK, pH, and aspect (P < 0.05) (Table 5). In the herbaceous layer, the environmental factors mainly influenced leaf stoichiometric traits (LCC, LNC, LPC and LKC). Stand openness, LAI, MLA, and slope had positive effects on LCC, while PPFD Under, SOC, STK, and soil moisture had negative effects on LCC. LNC was positively influenced by soil moisture and slope, while LNC was negatively influenced by stand openness, LAI, and aspect. LPC was positively influenced by soil moisture, while LPC was negatively influenced by stand openness, LAI, and SOC. LKC was positively influenced by STN and soil moisture, while LKC was negatively influenced by stand openness, LAI, and SOC (P < 0.05) (Table 5).

Standardized redundancy analysis of 7 functional traits by 11 environmental factors. RDA of community-weighted mean trait values (CWM) of the tree layer (a), the shrub layer (b), the herbaceous layer (c). SOC, soil organic carbon; STN, soil total nitrogen; STK, soil total potassium; STP, soil total phosphorus; pH, soil pH; M, soil moisture; LAI, leaf area index; MLA, mean leaf angle; OP, stand openness; PPFD Over and PPFD Under, photosynthetic photon flux density above and below canopy; TP, topographic position; H, height; LMA, leaf mass per unit area; LA, leaf area; LCC, leaf carbon content; LNC, leaf nitrogen content; LPC, leaf phosphorous content; LKC, leaf potassium content.Arrows indicate the direction and magnitude of variables. Blue arrows represent response variables; red arrows represent explanatory variables. Relationship between variables in RDA is expressed by a vector angle: if vector angle between each two variables is acute, they are positively correlated. Otherwise, if the angel is obtuse, the two variables are negatively correlated (right angle means correlation). The smaller the angle, the higher the degree of association between the two variables
Discussion
Plant functional traits and plant strategy in the three different plant life forms
Our main results show that in a Q. wutaishanica forest of China, at local scale, plant functional traits vary across the different plant life forms that conform the woodland structure, and these variations were related to the variations in environmental conditions. The stratification of plants in the community is beneficial for plants to fully utilize light, water and space resources. The difference of plant height leads to the imbalance of competition for light and space resources, and then leads to the difference of plant functional traits. Shrubs and herbaceous species were characterized by smaller leaf, but at the same time, lower LMA and LCC, and higher LNC than tree species. These combinations of traits indicate that most of the species that grow on the understory allocate more resources to produce leaf with more nutrient content and, probably more leaf production compared to overstory species.
Functional traits should not be viewed and analyzed in isolation, as many traits are often coordinated with each other. The trade-offs of plant functional traits determine the life history strategy of plants and the niche occupied by various species in the community, which further affects the stable coexistence of multiple species (Vandermeer, 1972; Silvertown, 2004). When two or more important plant traits have consistent correlation in different species, the change of these traits can be regarded as forming a strategic dimension (Wright et al. 2006). Across species, high LNC or LPC is generally associated with high nutritional quality to the consumers in food webs (Pérez-Harguindeguy et al. 2013). Leaf nitrogen content tends to be closely correlated with mass-based maximum photosynthetic rates and with SLA (specific leaf area, the inverse of LMA) (Pérez-Harguindeguy et al. 2013). Plants with low LMA (high SLA) have lower leaf thickness, a larger light capture area, and higher LNC, all of which contribute to a higher net photosynthetic rate and a higher relative growth rate (Niinemets 2010; Wright et al. 2001). In this study, LNC and LMA had a negative correlation in the shrub and herbaceous layers, but the tree layer showed a positive relationship, which might reflect the characteristics of Q. wutaishanica leaves. The leaves of Q. wutaishanica are leathery with a thick outer epidermis wall, thick cuticula, compact spongy parenchyma with few and small intercellular spaces, and transcurrent vascular bundles with a strong sclerenchymatous bundle sheath (Waldhoff and Furch 2002). The leathery leaf yields a high LMA value, and compact spongy parenchyma is closely related with maximum photosynthetic rate associated with LNC and LPC. Correlations among plant functional traits for the three life forms were different, indicating that the survival strategies of the life forms are different (Zhang et al. 2010). In addition, the morphological structure of different herbaceous plants is very different, which reflects their different environmental adaptation strategies, and to a certain extent affects their photosynthetic efficiency and nutrient distribution. In our study, Allium victorialis with higher LA meant that it had a strong ability to capture light resources, higher relative growth rate and shorter leaf life, while Allium victorialis with lower LNC meant that under lower light conditions the lower allocation of leaf N and decreased dark respiration rates, thus maintaining a lower photosynthetic light compensation point. Galium maximowiczii tended to have lower LA and higher LNC meant that it had low metabolic rates so as to maximize the rate of photosynthetic carbon assimilation per unit N allocated to a leaf. This would be helpful to better adapt to the dark environment under the forest. Meanwhile, some species in herbaceous layer had similar LA, but different LNC, or similar LNC but different LA. This may be because STN had the highest spatial heterogeneity of STN in this area. Some important physiological processes, including photosynthesis, may be limited due to lack of STN, which leads to different trade-offs in resource acquisition strategy under the same environment.
Effects of environmental factors on plant functional traits
The distribution characteristics of species with different life forms are related to various abiotic and biotic environmental factors (Dı́az et al. 1998). Differentiation of traits at local scale can be explained by a few key environmental variables, which act as filters on the available species assemblage (Grime 2006). Our results showed that there were differences in environmental factors affecting the distribution of traits for different life forms. In the tree layer, mean leaf angle was positively correlated with LCC, which was also reported by Niinemets (2010). This result was found because foliage inclination angle distribution and foliage spatial aggregation are the major determinants of light harvesting at the shoot scale. Soil is considered to be a dominant factor in determining the functional traits of plant communities, and is closely related to the variation in plant functional traits (Becknell and Powers 2014; Liu et al. 2012; Laughlin et al. 2015). Jager et al. (2015) found that soil nutrients could lead to multiple independent plant functional traits. At the local scale analyzed, we found that higher contents of SOC were associated with lower LNC in the tree life form. This result indicates that few substances were distributed to photosynthesis or growth, but more substances were distributed in the leaves of trees to endure stressful environments such as low N availability. Topography factors can affect various environmental factors such as water, temperature and soil nutrients, which are the main driving factor affecting plant functional traits (Loreau et al. 2001; Kichenin et al., 2013; Hu et al., 2014). The results showed that slope had a negative effect on plant height and leaf area. This observation indicates that trees tend to grow smaller leaves and limit their height to resist water and nutrient loss caused by erosion when the slope is steep. Additionally, aspect has negative impact on plant height and a positive impact on LMA, which would be due to increase the resistance to collapse caused by low water potential (Koch et al. 2004; Woodruff et al. 2004).
Canopy variables are not only an environmental factor in the forest, determining the redistribution of light and water, and shaping the local environment, but also a key factor affecting the functional trait of understory plants. The interception of water and light by the canopy not only influences species height and leaf traits directly (Ellsworth and Reich 1993; Qiu et al. 2008), but also affects plant functional traits indirectly, by impacting the forest microenvironment (Pelt and Franklin 2000). In our study, plant functional traits were mainly related to PPFD over and MLA in the tree layer (Fig. 3a). PPFD over and MLA had negative correlations with tree height, while MLA had a positive correlation with LA and LCC (Table 5). Canopy variables affecting functional traits of shrub layer and herbaceous layer were PPFD under and openness respectively (Fig. 3b, c). PPFD under had a negative correlation with height and LA in shrub layer (Table 5). Openness had a negative correlation with height, LNC, LPC and LKC, while had a positive correlation with LMA and LCC (Table 5). It can be seen that due to the different life forms of plants, the ways of obtaining light, water and other resources are also different, which leads to the different sensitivity of plant functional traits to certain canopy variable.
Conclusions
The plant function traits of different life forms showed high coefficient of variation, and the differences among different life forms were obvious, which was directly related to the complex structure and high biodiversity of forest community. Plant functional traits were significantly correlated with abiotic environmental factors, indicating that species composition was, at least partly, shaped by habitat filtering at the community scale. Key canopy variables, topographic and soil factors controlling plant functional traits changed across the different plant life forms. Each community-level plant functional trait was associated with 1–8 abiotic factors across different life forms. These results may contribute to forest management and increase our understanding of forest community assembly. The analysis demonstrates that it is important to consider the suites of traits at various hierarchical levels, and dynamic modifications in these traits in order to characterize species strategies across different plant life forms and understanding ecological processes in temperate forest ecosystem.
References
Barbier S, Gosselin F, Balandier P (2008) Influence of tree species on understory vegetation diversity and mechanisms involved-a critical review for temperate and boreal forests. For Ecol Manag 254:1–15. https://doi.org/10.1016/j.foreco.2007.09.038
Barboni D, Harrison SP, Bartlein PJ, Jalut G, New M, Prentice IC, Sanchez-Goni MF, Spessa A, Davis B, Stevenson AC (2004) Relationships between plant traits and climate in the Mediterranean region: a pollen data analysis. J Veg Sci 15:635–646. https://doi.org/10.1111/j.1654-1103.2004.tb02305.x
Becknell JM, Powers JS (2014) Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest. Can J Forest Res 44:604–613. https://doi.org/10.1139/cjfr-2013-0331
Bennie J, Huntley B, Wiltshire A, Hill MO, Baxter R (2008) Slope, aspect and climate: spatially explicit and implicit models of topographic microclimate in chalk grassland. Ecol Model 216:47–59. https://doi.org/10.1016/j.ecolmodel.2008.04.010
Berry JK (2005) Analyzing geo-spatial resource data: a hands-on case study in spatial analysis and data mining. University of Denver, Denver, CO
Bu WS, Zang RG, Ding Y, Zhang JY, Ruan YZ (2013) Relationships between plant functional traits at the community level and environmental factors during succession in a tropical lowland rainforest on Hainan Island, South China. Biodivers Sci 21:278–287 (in Chinese with English abstract)
Cambardella CA, Moorman TB, Novak JM, Parkin TB, Karlen DL, Turco RF, Konopka AE (1994) Field-scale variability of soil properties in Central Iowa soils. Soil Sci Soc Am J 58:1501–1511. https://doi.org/10.2136/sssaj1994.03615995005800050033x
Chaves MM, Maroco JP, Pereira JS (2003) Understanding plant responses to drought—from genes to the whole plant. Funct Plant Biol 30:239–264. https://doi.org/10.1071/FP02076
Chen J, Wu ZY, Raven PH (1999) Flora of China. Science Press, Beijing
Cheng XQ, Han HR, Kang FF, Song YL, Liu K (2014) Point pattern analysis of different life stages of Quercus liaotungensis in Lingkong Mountain, Shanxi Province, China. J Plant Interact 9:233–240. https://doi.org/10.1080/17429145.2013.818167
Coelho MS, Carlos PP, Pinto VD, Meireles A, Negreiros D, Morellato LPC, Fernandes W (2018) Connection between tree functional traits and environmental parameters in an archipelago of montane forests surrounded by rupestrian grasslands. Flora 238:51–59. https://doi.org/10.1016/j.flora.2017.04.003
Cornelissen JHC, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich DE, Pausas JG (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380. https://doi.org/10.1071/BT02124
de Bello F, Lavorel S, Díaz S, Harrington R, Cornelissen JH, Bardgett RD, da Silva PM (2010) Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers Conserv 19:2873–2893. https://doi.org/10.1007/s10531-010-9850-9
Dı́az S, Cabido M (2001) Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655. https://doi.org/10.1016/S0169-5347(01)02283-2
Dı́az S, Cabido M, Casanoves F (1998) Plant functional traits and environmental filters at a regional scale. J Veg Sci 9:113–122. https://doi.org/10.2307/3237229
Ding J, Wu Q, Yan H, Zhang SR (2011) Effects of topographic variations and soil characteristics on plant functional traits in a subtropical evergreen broad-leaved forest. Biodivers Sci 19:158–167 (in Chinese with English abstract)
Ellsworth DS, Reich PB (1993) Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia 96:169–178. https://doi.org/10.1007/BF00317729
George LO, Bazzaz FA (2003) The herbaceous layer as a filter determining spatial pattern in forest tree regeneration. In: Gilliam FS, Roberts MR (eds) The herbaceous layer in forests of eastern North America. Oxford University Press, New York, pp 265–282
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences. J Veg Sci 17:255–260. https://doi.org/10.1111/j.1654-1103.2006.tb02444.x
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237:173–195. https://doi.org/10.1023/A:1013351617532
Hu YS, Yao XY, Liu YH (2014) The functional traits of forests at different succession stages and their relationship to terrain factors in Changbai Mountains. Acta Ecol Sinica 34(20):5915–5924 (in Chinese with English abstract)
Jager MM, Richardson SJ, Bellingham PJ, Clearwater MJ, Laughlin DC (2015) Soil fertility induces coordinated responses of multiple independent functional traits. J Ecol 103:374–385. https://doi.org/10.1111/1365-2745.12366
Kichenin E, Wardle DA, Peltzer DA, Morse CW, Freschet GT (2013) Contrasting effects of plant inter- and intraspecific variation on community-level trait measures along an environmental gradient. Funct Ecol 27(5):1254–1261. https://doi.org/10.1111/1365-2435.12116
Kobayashi T, Shimano K, Muraoka H (2004) Effect of light availability on the carbon gain of beech (Fagus crenata) seedlings with reference to the density of dwarf bamboo (Sasa kurilensis) in an understory of Japan Sea type beech forest. Plant Spec Biol 19:33–46. https://doi.org/10.1111/j.1442-1984.2004.00099.x
Koide RT, Fernandez C, Malcolm G (2014) Determining place and process: functional traits of ectomycorrhizal fungi that affect both community structure and ecosystem function. New Phytol 201:433–439. https://doi.org/10.1111/nph.12538
Koch GW, Sillett SC, Jennings GM, Davis SD (2004) The limits to tree height. Nature 428:851–854. https://doi.org/10.1038/nature02417
Kraft NJ, Valencia R, Ackerly DD (2008) Functional traits and niche-based tree community assembly in an Amazonian forest. Science 322:580–582. https://doi.org/10.1126/science.1160662
Kunstler G, Falster D, Coomes DA, Hui F, Kooyman RM, Laughlin DC, Poorter L, Vanderwel M, Vieilledent G, Wright SJ, Aiba M, Baraloto C, Caspersen J, Cornelissen JHC, Gourlet-Fleury S, Hanewinkel M, Herault B, Kattge J, Kurokawa H, Onoda Y, Peñuelas J, Poorter H, Uriarte M, Richardson S, Ruiz-Benito P, Sun IF, Ståhl G, Swenson NG, Thompson J, Westerlund B, Wirth C, Zavala MA, Zeng H, Zimmerman JK, Zimmermann NE, Westoby M (2016) Plant functional traits have globally consistent effects on competition. Nature 529:204–207. https://doi.org/10.1038/nature16476
Laughlin DC, Richardson SJ, Wright EF, Bellingham PJ (2015) Environmental filtering and positive plant litter feedback simultaneously explain correlations between leaf traits and soil fertility. Ecosystems 18:1269–1280. https://doi.org/10.1007/s10021-015-9899-0
Lavorel S, Grigulis K, McIntyre S, Williams NS, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A (2008) Assessing functional diversity in the field–methodology matters! Funct Ecol 22:134–147. https://doi.org/10.1111/j.1365-2435.2007.01339.x
Lebrija-Trejos E, Pérez-García EA, Meave JA, Bongers F, Poorter L (2010) Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology 91:386–398. https://doi.org/10.1890/08-1449.1
Lhotka JM, Loewenstein EF (2008) Influence of canopy structure on the survival and growth of underplanted seedlings. New For 35:89–104. https://doi.org/10.1007/s11056-007-9063-6
Lin C, Ma QY, Han HR, Cao WQ, Wang ZZ, Wang ZQ, Zhang BX (2002) Phytosynthesis characteristic of Quercus liaotungensis in Taiyue mountain Region. Acta Ecol Sinica 22:1399–1406 (in Chinese with English abstract)
Liu J, Yunhong T, Slik JF (2014) Topography related habitat associations of tree species traits, composition and diversity in a Chinese tropical forest. For Ecol Manage 330:75–81. https://doi.org/10.1016/j.foreco.2014.06.045
Liu XJ, Swenson NG, Wright SJ, Zhang L, Song K, Du YJ, Zhang JL, Mi XC, Ren HB, Ma KP (2012) Covariation in plant functional traits and soil fertility within two species-rich forests. PLoS ONE 7:e34767. https://doi.org/10.1371/journal.pone.0034767
Loreau M, Naeem P (2001) Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294(5543):804–808. https://doi.org/10.1126/science.1064088
Matsushima M, Chang SX (2007) Effects of understory removal, N fertilization, and litter layer removal on soil N cycling in a 13-year-old white spruce plantation infested with Canada bluejoint grass. Plant Soil 292:243–258. https://doi.org/10.1007/s11104-007-9220-x
Mcardle BH, Anderson MJ (2001) Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology 82:290–297. https://doi.org/10.1890/0012-9658(2001)082[0290:FMMTCD]2.0.CO;2
McIntyre S, Lavorel S, Landsberg J, Forbes T (1999) Disturbance response in vegetation towards a global perspective on functional traits. J Veg Sci 10:621–630. https://doi.org/10.2307/3237077
Meng TT, Ni J, Wang GH (2007) Plant functional traits, environments and ecosystem functioning. J Plant Ecol 31:150–165 (in Chinese with English abstract)
Negassa W, Baum C, Schlichting A, Müller J, Leinweber P (2019) Small-scale spatial variability of soil chemical and biochemical properties in a rewetted degraded peatland. Front Environ Sci 7:1–16. https://doi.org/10.3389/fenvs.2019.00116
Nieuwenhuize J, Maas YE, Middelburg JJ (1994) Rapid analysis of organic carbon and nitrogen in particulate materials. Mar Chem 45:217–224. https://doi.org/10.1016/0304-4203(94)90005-1
Niinemets Ü (2010) A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol Res 25:693–714. https://doi.org/10.1007/s11284-010-0712-4
Onoda Y, Schieving F, Anten NP (2008) Effects of light and nutrient availability on leaf mechanical properties of Plantago major: a conceptual approach. Ann Bot-London 101:727–736. https://doi.org/10.1093/aob/mcn013
Parker WC, Dey DC (2008) Influence of overstory density on ecophysiology of red oak (Quercus rubra) and sugar maple (Acer saccharum) seedlings in central Ontario shelterwoods. Tree Physiol 28:797–804. https://doi.org/10.1093/treephys/28.5.797
Pelt RV, Franklin JF (2000) Influence of canopy structure on the understory environment in tall, old-growth, conifer forests. Can J Forest Res 30:1231–1245. https://doi.org/10.1139/x00-050
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte MS, Cornwell WK, Craine JM, Gurvich DE (2013) New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot 61:167–234. https://doi.org/10.1071/BT12225_CO
Poorter H, Niinemets Ü, Poorter L, Wright IJ, Villar R (2009) Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol 182:565–588. https://doi.org/10.1111/j.1469-8137.2009.02830.x
Prado-Junior JA, Schiavini I, Vale VS, Arantes CS, van der Sande MT, Lohbeck M, Poorter L (2016) Conservative species drive biomass productivity in tropical dry forests. J Ecol 104:817–827. https://doi.org/10.1111/1365-2745.12543
Pratt PF (1965) Digestion with hydrofluoric and perchloric acids for total. Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties (methods of soil anb) pp 1019–1021.
Qiu JL, Li YD, Chen DX (2008) The research progress and the significance of canopy structure in forest ecology. J Guangdong For Sci Technol 24(1):75–82 (in Chinese with English abstract)
Rao CR (1964) The use and interpretation of principal component analysis in applied research. Sankhyā Ind. J. Stat. Ser. A 26:329–358
Rodriguez-Calcerrada J, Mutke S, Alonso J, Gil L, Pardos JA, Aranda I (2008) Influence of overstory density on understory light, soil moisture, and survival of two underplanted oak species in a Mediterranean montane Scots pine forest. Forest Syst 17:31–38. https://doi.org/10.5424/srf/2008171-01021
Schweiger AK, Schütz M, Risch AC, Kneubühler M, Haller R, Schaepman ME (2017) How to predict plant functional types using imaging spectroscopy: Linking vegetation community traits, plant functional types and spectral response. Methods Ecol Evol 8:86–95. https://doi.org/10.1111/2041-210X.12642
Silvertown J (2004) Plant coexistence and the niche. Trends Ecol Evol 19:605–611. https://doi.org/10.1016/j.tree.2004.09.003
Sommers LE, Nelson DW (1972) Determination of total phosphorus in soils: a rapid perchloric acid digestion procedure. Soil Sci Soc Am J 36:902–904. https://doi.org/10.2136/sssaj1972.03615995003600060020x
Song DP, Li H, Liu SJ, Zou GY, Liu DS (2019) A geostatistic investigation of the comprehensive evaluation of fertility and spatial heterogeneity of forest soil nutrients in hilly and mountainous regions of southern China. Arab J Geosci 12:292. https://doi.org/10.1007/s12517-019-4480-3
Valladares F, Wright SJ, Lasso E, Kitajima K, Pearcy RW (2000) Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology 81:1925–1936. https://doi.org/10.1890/0012-9658(2000)081[1925:PPRTLO]2.0.CO;2
Vandermeer JH (1972) Niche theory. Annu Rev Ecol Syst 3:107–132. https://doi.org/10.1146/annurev.es.03.110172.000543
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007) Let the concept of trait be functional! Oikos 116:882–892. https://doi.org/10.1111/j.0030-1299.2007.15559.x
Waldhoff D, Furch B (2002) Leaf morphology and anatomy in eleven tree species from Central Amazonian floodplains (Brazil). Amazoniana XVII 17:79–94. https://doi.org/10.1080/00207217.2012.743081
Wang T, Kang F, Cheng X, Han H, Ji W (2016) Soil organic carbon and total nitrogen stocks under different land uses in a hilly ecological restoration area of North China. Soil Till Res 163:176–184. https://doi.org/10.1016/j.still.2016.05.015
Wang T, Kang FF, Cheng XQ, Han HR, Bai YC, Ma JY (2017) Spatial variability of organic carbon and total nitrogen in the soils of a subalpine forested catchment at Mt. Taiyue, China, Catena 155:41–52. https://doi.org/10.1016/j.catena.2017.03.004
Woodruff DR, Bond BJ, Meinzer FC (2004) Does turgor limit growth in tall trees? Plant Cell Environ 27:229–236. https://doi.org/10.1111/j.1365-3040.2003.01141.x
Wilsey BJ (1996) Urea additions and defoliation affect plant responses to elevated CO2 in a C3 grass from Yellowstone National Park. Oecologia 108:321–327. https://doi.org/10.1007/BF00334657
Wright IJ, Falster DS, Pickup M, Westoby M (2006) Cross-species patterns in the coordination between leaf and stem traits, and their implications for plant hydraulics. Physiol Plantarum 127:445–456. https://doi.org/10.1111/j.1399-3054.2006.00699.x
Wright IJ, Reich PB, Westoby M (2001) Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low- nutrient habitats. Funct Ecol 15:423–434. https://doi.org/10.1046/j.0269-8463.2001.00542.x
Yu M, Sun OJ (2013) Effects of forest patch type and site on herb-layer vegetation in a temperate forest ecosystem. For Ecol Manag 300:14–20. https://doi.org/10.1016/j.foreco.2012.12.039
Zhang HW, Ma JY, Sun W, Chen FH (2010) Altitudinal variation in functional traits of Picea schrenkiana vat. tianschanica and their relationship to soil factors in Tianshan Mountains. Northwest China Acta Ecol Sinica 30:5747–5758 (in Chinese with English abstract)
Zhao J, Wan SZ, Fu SL, Wang XL, Wang M, Liang C, Chen YQ, Zhu XL (2013) Effects of understory removal and nitrogen fertilization on soil microbial communities in Eucalyptus plantations. For Ecol Manag 310:80–86. https://doi.org/10.1016/j.foreco.2013.08.013
Zhao J, Wang X, Shao Y, Xu G, Fu S (2011) Effects of vegetation removal on soil properties and decomposer organisms. Soil Biol Biochem 43:954–960. https://doi.org/10.1016/j.soilbio.2011.01.010
Zhu YY, Wang XA, Xian W, Deng MJ (2016) Effect of slope aspect on the functional diversity of grass communities in the Loess Plateau. Acta Ecol Sinica 36:6823–6833 (in Chinese with English abstract)
Acknowledgements
The study was supported by the National Natural Science Foundation of China (No. 31700372), the National Key Research and Development Program of China (No. 2019YFA0607304), and National Natural Science Foundation of China (No. 31800392). We gratefully acknowledge the fieldwork support from the Taiyue Forestry Bureau. We also thank all those who provided helpful suggestions and comments on improving the quality of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Cheng, X., Ping, T., Li, Z. et al. Effects of environmental factors on plant functional traits across different plant life forms in a temperate forest ecosystem. New Forests 53, 125–142 (2022). https://doi.org/10.1007/s11056-021-09847-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-021-09847-0