
Abstract
In Brazil, most Eucalyptus plantations are located in regions experiencing periods of water shortage where fertilizers are intensively used to achieve high productivity. Fertilization can affect water use. However, the effects of fertilization on tree growth patterns during extreme droughts periods remain unknown. A throughfall exclusion experiment was set up in São Paulo State-Brazil to study the effects of potassium (K) and sodium (Na) fertilization and their interaction with water supply in the growth of Eucalyptus grandis trees over an abnormal season of 6 months of extreme drought in comparison with that in normal seasons, as well as the differences in responsiveness to intra-annual meteorological variability. Arranged in a split-plot design, the factors were water supply (37% throughfall exclusion vs. no throughfall exclusion) and fertilization regime (K, Na, and control). Basal area growth was monitored by band dendrometers measurements at 14-day intervals over 2 years. Meteorological and soil water content data were also collected. K and Na fertilization increased the tree basal area by four and three-fold, respectively, during normal seasons. During a severe drought season, these positive effects were suppressed. However, K- and Na-fertilized trees achieved a similar cumulative basal area increment to that of the control trees. The 37% throughfall exclusion significantly decreased tree growth in all treatments only in the severe drought period, and these effects were stronger in K-fertilized trees. K-fertilized trees were highly responsive to intra-annual meteorological variability. Our results suggest that extreme drought has similar effects on E. grandis tree growth regardless of the K/Na fertilization regime.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Forests are subject to climate change effects, such as long drought periods, changes in rainfall pattern frequency and amount, and temperature increase (IPCC 2013). Drought periods affect the growth of trees in natural forests, which can lead to tree death (Williamson et al. 2000; Cox et al. 2004; Allen et al. 2010), and some studies have shown that the same effects could occur in Eucalyptus planted forests (Cabral et al. 2011).
In Brazil, most Eucalyptus forest plantations are located in Southeast regions subject to long dry periods of 3–5 months during which the monthly rainfall is less than 50 mm (Gonçalves et al. 2013), such as São Paulo State, which covers 17% (0.95 million ha) of the total area of Eucalyptus plantations in Brazil (5.7 million ha) (IBÁ 2017). São Paulo and other states of the southeast have historically been characterized by drought events with serious effects on water availability (Coelho et al. 2016b). Since 2000, the frequency of the dry periods (monthly rainfall below 50 mm) has increased from that in 1965–2000, and their duration has increased in the last decade (Nobre et al. 2016). In fact, the long dry period that occurred in southeastern Brazil from the end of 2013 until the end of 2014 is considered to be the most severe drought since 1961 (Coelho et al. 2016a, b) or even since 1930 (Dearden 2015), and led to the worst water crisis in São Paulo since 1961 (Nobre et al. 2016).
Eucalyptus grandis W. Hill ex Maiden is the most planted and one of the most productive species in Brazil (Campinhos 1999; Gonçalves et al. 2013). The fertilization in the soil fertility improvement is known to be essential for the successful establishment of eucalyptus plantations (Gonçalves and de Barros 1999; Zeng et al. 2013). Depending on the characteristics of the plantation site, nitrogen, phosphorus, and potassium fertilization are needed to achieve high productivity rates in E. grandis plantations (Gonçalves et al. 2004). Recently, sodium has been considered a potential fertilizer in K-deficient soils because E. grandis tree growth showed a positive response to NaCl applications in a partial replacement of KCl (Almeida et al. 2010).
Fertilization can affect the water balance of the trees (Gonçalves and Benedetti 2000; Battie-Laclau et al. 2014; Ward et al. 2015). In Eucalyptus globulus, nitrogen supply can increase the water stress and mortality of trees at water-limited sites (White et al. 2009). In Pinus taeda, fertilizers increase resiliency to minor water stress in the short term (Ward et al. 2015) and a reduction in water availability does not affect the growth of fertilized trees (Samuelson et al. 2014). However, the long-term effects of fertilization with respect to drought sensitivity remain a concern (Ward et al. 2015), and long-term studies on the effects of fertilizers in interaction with rainfall variability are needed to better understand how climate change affects forest productivity (Beier et al. 2012; Samuelson et al. 2014).
Previous studies have revealed that K and Na fertilization improves water use efficiency and increases wood production in E. grandis trees two to three-fold (de Almeida 2009; Almeida et al. 2010; Battie-Laclau et al. 2016). However, K and Na fertilization can also increase stomatal conductance, accompanied by a reduction in soil water storage (Battie-Laclau et al. 2014). This finding, along with a high leaf area index (LAI) and time series patterns of a soil-to-leaf water potential gradient, suggested that K and Na increase the risk of hydraulic dysfunction following extreme water deficiency (Battie-Laclau et al. 2014). Therefore, there is concern that, in an extreme and prolonged drought, K and Na could aggravate the effects of water stress and cause tree mortality (Battie-Laclau et al. 2014). In this context, studies during severe droughts are necessary to understand the actual effects of K and Na fertilization in the development of E. grandis trees.
Studies conducted on the early growth stages of Eucalyptus trees improved the understanding of the effects of K and Na on drought adaptation mechanisms at the physiological level (Battie-Laclau et al. 2014, 2016). However, an understanding of the growth patterns of E. grandis trees over the complete rotation period is also important to determine the effect of fertilization and water availability, because growth is dependent on short-term changes in environmental conditions, such as temperature and rainfall (Deslauriers et al. 2007). In this regard, high-frequency dendrometer band measurements are used to assess secondary growth changes over long periods and investigate their relationships with climate (Knott 2004; Vitas 2011; Bošeľa et al. 2013; Campoe et al. 2016). Previous studies using dendrometer bands have demonstrated a strong effect of intra-annual meteorological variability on basal area growth (Sette et al. 2010, 2012; Campoe et al. 2016). However, the growth patterns of E. grandis trees fertilized with K and Na under throughfall exclusion in response to intra-annual meteorological variability are still unknown.
Given the widespread practice of incorporating high amounts of K during the fertilization of Eucalyptus plantations in water-deficit regions (Gonçalves and de Barros 1999) and the potential use of Na as a substitute for K (Almeida et al. 2010), an experiment of fertilization levels and throughfall exclusion was initiated in 2010 in Itatinga, Brazil, to evaluate the interactions of K and Na supply with water availability. Here, we monitored changes in tree secondary growth at fortnightly intervals from 3 to 5 years after planting, over rainy and dry seasons, including the period of drought in 2014. Our objectives were: (1) to evaluate the basal area growth patterns of E. grandis trees fertilized with K and Na in interaction with 37% throughfall exclusion over the abnormal dry season of 2014, characterized by a severe drought (44% less rainfall than historical average, and monthly rainfall < 50 mm), compared with those during normal seasonal periods (seasons with rainfall volume within the range of the historical average). (2) To evaluate their differences in responsiveness to intra-annual meteorological variability.
We hypothesize that, over normal rainy and dry seasonal periods, K and Na fertilization would promote high growth rates, increasing the basal area growth up to three to four-fold. Over an abnormal dry season, the positive effects of K and Na fertilization on basal area increment would be suppressed and a lower cumulative basal area than that in K and Na deficient trees would be observed. Based on the literature that reported declines of tree growth under water availability reductions (Battie-Laclau et al. 2014; Samuelson et al. 2014), we expected that 37% throughfall exclusion would decrease tree growth independent of fertilization regime and climatic conditions. Finally, we hypothesized that K and Na fertilization would significantly increase the responsiveness of trees to intra-annual meteorological variability.
Materials and methods
Study area
The experiment was conducted at the Itatinga Experimental Station of the University of São Paulo in Brazil (23°02′S, 48°38′W). The mean annual rainfall and temperature are 1360 mm and 19.9 °C, respectively. The experiment was set up on an elevated terrain with 3% slope at an altitude of 850 m. The soils are deep ferralsols with 14–23% clay content (for more details about the soil characteristics, see Battie-Laclau et al. 2014 and Laclau et al. 2010).
Experimental design
A split plot design was initiated on June 20, 2010, with a highly productive E. grandis clone used in commercial plantations by the Suzano Company (Brazil). At the time of planting, all plants were fertilized with 75 kg ha−1 of P2O5, 80 kg ha−1 of N (NH4(SO4)2), and 20 kg ha−1 of FTE BR-12 (micronutrient source) applied in holes next to the plants. Furthermore, 2000 kg ha−1 of dolomitic limestone was broadcast on the soil surface. These nutrients were not limiting factors for tree growth in our study site (Laclau et al. 2009).
Six treatments (2 water supply regimes × 3 fertilization regimes) were installed in three blocks. The whole-plot factor was the water supply regime, namely, 37% throughfall exclusion in the plot (− W) and no throughfall exclusion in the plot (+ W). The split-plot factor (nested within each plot) was the fertilization regime, namely, potassium supply (K) with 335 kg ha−1 (or 4.5 kmol ha−1) of KCl, sodium supply (Na) with 263 kg ha−1 (or 4.5 kmol ha−1) of NaCl, and control (C) without K or Na supply. KCl and NaCl were broadcast on the soil surface in a single dose 3 months after planting.
The area of each subplot was 864 m2 with 144 trees and a spacing of 2 m × 3 m. Throughfall exclusion devices on − W plots were installed in September 2010 and retained thereafter. Transparent plastic panels were used, mounted on wooden frames at a height varying between 0.5 and 1.6 m (Battie-Laclau et al. 2016) and covering 37% of the area in − W plots.
Tree growth monitoring
Secondary growth was monitored via periodic measurements of circumference at breast height (CBH) increment using dendrometer bands installed in three trees in each subplot (nine trees per treatment), located in the third row. Measurements were recorded from October 2013 (40 months after planting) to October 2015 (65 months after planting) at 14-day intervals in the early hours of the morning. The first measurements were recorded 1 month after band dendrometer installation to avoid underestimations of CBH owing to initial slack in tree boles (Fuller et al. 1988; Keeland and Sharitz 1993). In addition, CBH was measured directly with a tape measure at the beginning of each season in order to verify and correct possible underestimations of band dendrometers.
Meteorological and soil water content data
Rainfall, mean air temperature (Tmean), minimum air temperature (Tmin), maximum air temperature (Tmax), and vapor pressure deficit (VPD) data were collected at 30-min intervals from an automatic weather station in the Itatinga Experimental Station located 50 m away from the experiment. Raw meteorological data were converted to a daily timescale.
Soil water content (SWC) in m3 m−3, was measured weekly in the subplots of the first block. In each subplot, three soil moisture sensors (Trase Soilmoisture, Santa Barbara, CA, USA) were installed at 0.15 m, 0.5 m, 1 m, 1.5 m, 3 m, 4.5 m, and 6 m depths, 1 month before the plantation was established. TDR sensors were calibrated via gravimetry of soil water content and density measurements.
Daily meteorological and weekly soil moisture data were organized into fortnightly periods to match the dendrometer readings. Moreover, daily rainfall data were summarized to obtain cumulative rainfall for 6 month-interval periods: rainy seasons (October to March) and dry seasons (April to September) in the first and second year of assessment.
Rainfall deficiency was calculated based on differences between historical monthly average rainfall data available from 2006 to 2013 and monthly rainfall data from October 2013 to October 2015, registered at the weather station in the Itatinga Experimental Station.
Data analysis
For all sampled trees from each treatment, the basal area increment (cm2 tree−1) was calculated from bole CBH increment data generated by the band dendrometer readings.
Analysis of variance (ANOVA) with a split-plot model and Tukey’s test were used to test the effects of water supply regime, fertilization regime, and the interaction of these factors (fixed effects) on cumulative basal area increment within each rainy and dry season along the 2 years of assessment. Blocks and water regime × block were considered random effects.
Stepwise multiple linear regression analysis for each treatment (387 data points) was used to identify the meteorological variables with the greatest effect on tree basal area increments and establish models to predict basal area growth rates as a function of meteorological variables. Additional tests were performed including preceding meteorological data in 1, 2 and 3 days at a given growth interval, in order to prove the absence of any lag in growth response.
In addition, Pearson correlation was used to assess the relationships between meteorological variables, soil water content (SWC), and tree basal area increment in each treatment.
Results
Meteorological seasonality and rainfall anomalies
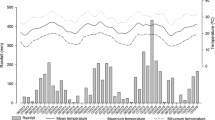
Over the 2 years, marked seasonality was observed in cumulative rainfall, Tmean, Tmin, and Tmax, which were lower during the dry seasons (April to September) than during the rainy seasons (October to March) (Fig. 1b). In contrast, the variability of VPD between the dry and rainy seasons was less marked, although the lowest values of VPD occurred at the end of the rainy seasons and the beginning of the dry seasons (March to June) (Fig. 1a).

Daily variations of vapor pressure deficit (VPD) (a) and daily variations of rainfall, mean air temperature (Tmean), minimum air temperature (Tmin) and maximum air temperature (Tmax) (b) from September 2013 until October 2015, at Itatinga Experimental Station, Itatinga, São Paulo, Brazil. The shaded areas represent the dry season (April–September)
Historical data from 2006/2007 to 2012/2013 shown an average annual rainfall of 1690 mm in Itatinga-SP. Normally, in rainy seasons (October to March) cumulative rainfall is 1146 mm, while in dry seasons (April to September) it is 544 mm.
However, during the first year of study (October 2013 to September 2014), cumulative rainfall was 946 mm and a considerable monthly rainfall deficiency was observed ranging from 61 to 186 mm during November, December 2013, and January, February, April, June, and July 2014 (Fig. 2b). Cumulative rainfall in the rainy season of 2013/2014 (October 2013 to March 2014) was 621 mm, while in the dry season of 2014 (April to September 2014) it was 325 mm, in which the driest months were recorded in June, July and August 2014 (Fig. 2a).

Monthly rainfall time series (a) and monthly rainfall anomaly time series (b) from October 2013 to October 2015 for the Itatinga Experimental Station. Monthly rainfall series were compared to historical average of 2006–2012/2013 and anomalies were relative to the 2006–2012/2013 monthly mean. The shaded areas represent the dry season (April–September)
In contrast, during the second year (October 2014 to September 2015), cumulative rainfall was 1686 mm and monthly rainfall deficiency occurred only in October 2014 and January, April, and June 2015. The remaining months showed a rainfall surplus (Fig. 2b). Cumulative rainfall in the rainy season of 2014/2015 (October 2014 to March 2015) was 1106 mm, whereas, in the dry season of 2015 (April to September 2015), it was 580 mm.
In the rainy season of 2013/2014 and dry season of 2014, there was 44% less rainfall than the 2014/2015 rainy season and dry season of 2015, respectively, as well as 46% and 40% less rainfall than historical average.
Soil water content (SWC)
The lowest soil water content (SWC) values occurred from June to October 2014, independent of treatment owing to the extreme rainfall deficiency in these months. The SWC in superficial layers (0.15–1.5 m) was highly responsive to rainfall (Fig. 3a). However, in deep layers (3–6 m) SWC was less responsive to rainfall events and varied less, except in the first months of 2015 when the SWC increased considerably owing to high rainfall (Fig. 3b).

Series of soil water content (SWC) in 0.15–1.5 m soil layer (a) and in 3–6 m soil layer (b). Since February 18, 2015, there was no data due to TDR sensor malfunction. The shaded areas represent the dry season (April–September)
In both superficial and deep soil layers, SWC was maximum in the C + W subplots and minimum in the K − W subplots. + W plots always had higher SWC than did the − W plots within the same fertilization regime. Throughfall exclusion reduced SWC by 12%–27% in the superficial soil layers, and the largest reductions were observed in K plots (22%–27%) (Fig. 4a). However, in soil deep layers, the reduction of SWC was less than 16%, and the largest reductions occurred also in K plots (15%–16%), whereas the lowest reduction occurred in Na and C plots (0%–5%), except in the 2015 rainy season (Fig. 4b). Reductions of SWC were larger in rainy seasons and proportional to high amounts of rainfall over these periods.

Mean soil water content (SWC) in 0.15–1.5 m soil layer (a) and in 3–6 m soil layer (b) as a response to treatments of fertilization and water regime. Means sorted by season: rainy season 2013/2014 (October 2013 to March 2014), dry season 2014 (April 2014 to September 2014), and rainy season 2014/2015 (October 2014 to March 2015). Since February 18, 2015, there was no data due to TDR sensor malfunction
The mean comparisons showed that throughfall exclusion affected SWC reduction more strongly than the K and Na fertilization regime in superficial soil layers. In contrast, in deep soil layers, the effects of K and Na fertilization were stronger than those 37% throughfall exclusion (Fig. 4a, b). Thus, these patterns reflected the high water consumption of the K- and Na-fertilized trees relative to that of the control trees in the deep soil layers.
Time series patterns of SWC revealed effective SWC reduction by the throughfall exclusion device installed in − W plots.
Tree basal area growth
During the first year, the cumulative basal area followed the same increment pattern in all treatments. However, in the second year, after October 2014, cumulative basal area in K + W, K − W, and Na − W treatments increased considerably, whereas cumulative basal area in the Na + W, C + W, and C − W maintained similar increment patterns to those in the previous year (Fig. 5a).

Time series of cumulative basal area (a) and basal areal fortnightly increment (b) in cm−2 tree−1. Blue bars indicate daily rainfall (mm). Vertical bars indicate standard errors between trees (n = 9). From October 2013 to September 2015, white area represents the rainy seasons and the shaded area represents dry seasons. Extreme drought period corresponds to 2014 dry season
Fortnightly basal area increments did not show clear patterns of variation between treatments, i.e. K- and Na-fertilized trees did not always show the highest basal increments, and, trees under 37% throughfall exclusion did not always show the lowest increments (Fig. 5b). During rainy periods, K-fertilized trees showed significantly higher basal area increments than Na- fertilized trees (p < 0.0001), and the increments of both were significantly higher than those in the control trees (p < 0.0001). Increments were often significantly higher for + W trees than that for − W trees (p < 0.0001). However, after February 2015, − W trees showed higher or equal basal increments to those of + W trees (Fig. 5b).
In periods with lowest monthly rainfall values, which occurred in February, April, June, July, August, and October 2014 (Fig. 2a), trees did not grow (except those in the C + W treatments) and the increments were negative (between − 0.79 and 0 cm2 tree−1). However, after any rainy events, trees under K + W, K − W, Na + W, and Na− W treatments achieved high growth rates. C + W trees grew at low rates (approximately 0.3 cm2 tree−1) during the same periods (Fig. 5b). In contrast, during the 2015 dry season, patterns in basal area fortnightly increments were similar in all treatments and they were only decreased in July and August (Fig. 5b).
Variations in basal area fortnightly increments were higher in K and Na treatments, varying from − 0.84 to 1.96 cm2 tree−1, − 0.54 to 2.00 cm2 tree−1, − 0.33 to 1.77 cm2 tree−1, and − 0.33 to 1.59 cm2 tree−1, for K + W, K − W, Na + W, and Na − W, respectively. The range of variations for C+ W and C − W was from − 0.24 to 1.06 cm2 tree−1 and from − 0.18 to 0.87 cm2 tree−1, respectively.
Analysis of cumulative basal area in rainy and dry seasons revealed different tree growth responses (Table 1 and Fig. 6). During the 2013/2014 rainy season, despite considerable water deficiency (Fig. 2b), the positive effects of K and Na supply on tree growth were maintained (cumulative basal area increment increased by 51% and 33% from that of control) even in − W plots. In addition, in this period, there were no significant differences between K and Na supply with respect to basal area growth. However, during the 2014 dry season, with extreme water deficiency, the basal area increment was not significantly influenced by the fertilization regime. Over this particularly dry period, cumulative basal area increment significantly reduced (by 46%) in − W plots owing to the additional effect of 37% throughfall exclusion. In contrast, on good rainfall condition of 2014/2015 rainy season and the 2015 dry season, basal area growth was significantly influenced by the fertilization regime but not by the water supply regime. K and Na supply increased basal area by 298% and 107%, respectively, over the 2014/2015 rainy season with respect to control, whereas during the 2015 dry season, it increased by 393% and 163%, respectively.

Cumulative basal area increment during the rainy season of 2013/2014 (October 2013 to March 2014), the dry season of 2014 (April 2014 to September 2014), the rainy season of 2014/2015 (October 2014 to March 2015), and the dry season of 2015 (April 2015 to September 2015). Vertical bars indicate standard deviations
A significant interaction between the water regime and fertilization regime occurred only during the 2015 dry season, which reflected a negative effect of throughfall exclusion on tree growth for K-fertilized trees and a positive effect for Na-fertilized trees and control trees (Table 1 and Fig. 6).
Influence of intra-annual meteorological variability in tree basal area growth
Basal area fortnightly increments were significantly influenced by rainfall, VPD, and SWC in all treatments, whereas the relationship with Tmin was significant only in K-fertilized and Na-fertilized trees. Similarly, the relationship with Tmax was only significant in K-fertilized trees (Fig. 7). There was no correlation with Tmean.

Pearson correlation coefficients between basal area fortnightly increments vs. rainfall, mean air temperature (Tmean), minimum air temperature (Tmin), maximum air temperature (Tmax), vapor pressure deficit (VPD), soil water content (SWC) at 0.15–1.5 m depth (SWC 0.15–1.5) and at 3–6 m depth (SWC 3–6). nsNot significant at 0.05 level, *significant at 0.05 level, **significant at 0.01 level
Responsiveness of basal area fortnightly increment to meteorological variables was stronger in trees with K supply (K + W and K − W) than in trees with Na supply (Na + W and Na − W) and control (C + W and C − W). Moreover, the relationships were always stronger in the − W than in the + W regime (Fig. 7).
Meteorological data can be useful to predict the basal area growth of K-fertilized trees (R2adj = 0.51 p < 0.001 in K + W, and R2adj = 0.54 p < 0.001 in K − W), whereas the growth of Na-fertilized trees cannot be efficiently predicted using only the meteorological data (R2adj = 0.16 p < 0.001 in Na + W, and R2adj = 0.35 p < 0.001 in Na − W) (Table 2). Control trees were unresponsive to intra-annual meteorological variability during the study period.
Additional tests of regression models including 1, 2 and 3 days lag in growth response to meteorological variability are showed in Table 3. The greater the lag, the lower the goodness-of-fit of the growth equations. This indicates the absence of any discernible lag in growth response to intra-annual meteorological variability.
Discussion
Interactive effects of fertilization and water supply regimes in dry and rainy seasons
The long dry period that occurred in southeastern Brazil, from the end of 2013 until the end of 2014 is considered to be the most severe drought since 1961 (Coelho et al. 2016a, b), generating the worst water deficiency in São Paulo to date (Nobre et al. 2016). Particularly in Itatinga, over this period, cumulative rainfall was lower by 44% than that historical average, with the driest months of the 2014 dry season (monthly rainfall < 50 mm) occurring at this time (Fig. 2a). Thus, the 2014 dry season, with just 325 mm of cumulative rainfall over 6 months, can be regarded as an extreme drought period. In contrast, from October 2014 to September 2015, high rainfall was observed even during the dry season, exceeding the historical average in some months.
The higher basal area increments promoted by K and Na supply were maintained only under traditional conditions of rainy and dry seasons (i.e., normal amounts and frequency of rainfall). In this period, the higher increments of K- and Na-fertilized trees can be principally explained by an increase in water use efficiency for stemwood production elicited by K and Na in non-limiting water conditions (Battie-Laclau et al. 2016).
However, during the extreme drought period, the positive effects of K and Na fertilization on tree growth were suppressed. In fact, in periods with more than 15 days without rain, K- and Na-fertilized trees stopped their basal area growth, whereas control trees maintained very low growth rates (Fig. 5b).
Negative stem fortnightly basal area increments occurred in K- and Na-fertilized trees, during driest periods, probably reflecting variations in xylem diameter due to lower sap flow in trees over dry periods (Battie-Laclau 2013) because xylem water potential (proportional to sap flow) is closely related to xylem diameter variations (Sevanto et al. 2008).
However, under any rainfall event, K- and Na-fertilized trees recovered quickly their growth, reaching a cumulative basal area similar to that of the control trees during the extreme drought period.
Subsequently, the high rainfall amounts during 2014/2015 rainy season and 2015 dry season created favorable conditions for the rapid growth of K-fertilized trees (Fig. 5b). As consequence, the cumulative basal area was considerably higher in K + W and K − W, which further increased the difference between treatments (Fig. 5a). In addition, in the wettest periods, throughfall exclusion was favorable for high growth rates in the Na − W treatment, achieving an equal basal area size to that in Na + W trees. The higher SWC in superficial layers in Na + W plots could have affected respiration and nutrient translocation by roots (Sette et al. 2010), decreasing tree growth rates. Similarly, excess water could have negatively affected the trees in C + W and C − W treatments, because in these treatments, the water demand by trees was even lower.
Nonetheless, the higher cumulative basal area increment of trees with K and Na supply from the third to the fifth year after planting confirmed the positive effect of K and Na (higher in K) as reported in the same experiment during the first half of the rotation (Battie-Laclau et al. 2014, 2016; Castro et al. 2017). Thus, the positive effect of these nutrients on tree growth is maintained throughout the 5 years. Similarly, the intermediate effects of Na supply on the tree growth allow corroborating the previous results in younger E. grandis trees (Almeida et al. 2010; Battie-Laclau et al. 2014; Castro et al. 2017). The positive effects of other nutrients, such as nitrogen and phosphorus, in Eucalyptus plantations have been demonstrated (Stoneman et al. 1997; Laclau et al. 2009; White et al. 2009). As such, considering that all treatments in our experiment included nitrogen and phosphorus supply, the large differences on growth between K-fertilized and K-deficient trees indicated a strong additive effect of K supply, demonstrating the importance of this nutrient on the basic fertilization on E. grandis plantations in Itatinga region, Brazil.
On the other hand, although 37% throughfall exclusion significantly reduced soil water content in − W plots (Fig. 3), the effects on tree growth were not significant, especially on Na-fertilized and control trees, reaching a similar basal area after 2 years of assessments. However, the differences between K + W and K − W were larger but not significant (Fig. 5a) probably due to the proportion of soil water content reduction via throughfall exclusion device, which was higher in K − W plots (22–27%) compared to Na − W and C − W plots (12–23% and 20–23% respectively) (Fig. 4). Na-fertilized and control trees were more able to maintain their growth rates during dry periods as a result of lower water consumption than K-fertilized trees, which made it possible to store water in deep soil layers during rainy periods (Battie-Laclau et al. 2014). By contrast, the consequences of 37% throughfall exclusion were more intense on K-fertilized trees, decreasing basal area increments by 18%, because the high water consumption of these trees was necessary to maintain high growth rates promoted by the K supply (Battie-Laclau et al. 2016). However, the negative effects of 37% throughfall exclusion on tree growth in all treatments were significant only over the extreme drought period (Fig. 6, Table 1).
These interaction results are in agreement with observations over the first 3 years for this experimental plantation regarding stemwood biomass accumulation, aboveground net primary production, and gross primary production (Battie-Laclau et al. 2014, 2016; Christina et al. 2015).
Thus, the negative effect of 37% throughfall exclusion on tree growth was lower than that of K or Na deficiency. In addition, over normal seasonal periods, the 37% throughfall exclusion may not be enough to cause strong water stress in the trees, independent of the treatments. The absence of throughfall exclusion effects over no drought periods had also occurred in Pinus taeda L. plantation in the southeastern United States, where surplus rainfall occurred in 2013, reaching 1413 mm cumulative rainfall (Samuelson et al. 2014), and even, throughfall exclusion were beneficial to increase standing volume by 4% (Ward et al. 2015). Consistent with our results, throughfall exclusion in Pinus taeda was effective only over drought seasons, decreasing stemwood production by 13% (Samuelson et al. 2014).
The results of SWC on − W plots compared to + W plots, show that a 37% throughfall exclusion device get reduce SWC in a proportion from 12 to 27% at 0.15–1.5 m deep (Figs. 3, 4). Similarly, other experiments with 30% throughfall exclusion showed a reduction in SWC about 25% at 0–1.2 m deep (Ward et al. 2015). At greater depth, the reduction in SWC is lower (Fig. 3). We suppose that the reduction of SWC via throughfall exclusion device is not greater because part of the rainfall is intercepted by the tree crown and drained to the ground through the stem. However, the soil moisture sensors allowed to know the real water availability reduction at different depths of the soil.
Tree growth responsiveness to intra-annual meteorological variability
The differences in responsiveness of tree growth to the intra-annual meteorological variability among treatments (Fig. 7) were expressed in their substantial differences in patterns of basal area growth (Fig. 5b). Rainfall was the most important meteorological variable affecting tree growth in all treatments. The strong sensitivity of K-fertilized trees to intra-annual rainfall variability was reflected as growth cessation during the driest periods and high growth rates upon the occurrence of any rainfall event (Fig. 7). As consequence, after 6 months of extreme drought period, K-fertilized trees had grown at similar growth rates as in Na- and control fertilized trees. In agreement with our study, correlations between daily patterns of radial growth and daily rainfall variability were in the same order of magnitude in Eucalyptus globulus Labill. trees (r = 0.61) and Eucalyptus nitens (H. Deane & Maiden) Maiden trees (r = 0.51) (Downes et al. 1999). Despite the low temporal resolution of band dendrometers compared to the high temporal resolution of electronic dendrometers, band dendrometers were efficient enough to measure the responsiveness of the trees to climate.
K-fertilized trees were responsive to the minimum (positive relationship) and maximum (negative relationship) temperature, and Na-supplied trees were only affected by minimum temperature. In contrast, control trees were not responsive to the temperature. Minimum and maximum temperatures were also related to bole basal area growth in K-fertilized trees in older E. grandis trees of adjacent experiments nearby our study area (Sette et al. 2010; Campoe et al. 2016) and it has been also observed in E. globulus on a daily time scale (Downes et al. 1999). Temperature is considered as an important meteorological variable involved in biochemical processes, respiration, and stomatal conductance (Lin 2012). In addition, maximum temperature negatively influences stem growth by increasing respiration, reducing stomatal conductance to mitigate a higher transpiration demand, and reducing photosynthesis (Schippers et al. 2015).
Generally, bigger Eucalyptus trees are more responsive to meteorological variables than smaller trees in the same stands as a result of their higher light interception rates and light use efficiency (Binkley et al. 2010; Campoe et al. 2013, 2016). In our study, K + W trees were considerably bigger than K − W trees (Fig. 5a). However, K + W trees were less responsive to intra-annual meteorological variability compared to K − W trees (R2adj = 0.51 and 0.54 for K + W and K − W, respectively). In addition, tree size between Na + W and Na − W or between C + W and C − W treatments (Fig. 5a) were similar. However, the growth of trees in − W plots was considerably more responsive to intra-annual meteorological variability (R2adj = 0.35 and 0.19 for Na − W and C − W, respectively) compared to trees growing in + W plots (R2adj = 0.16 and 0.07 for Na + W and C + W, respectively) (Table 2). In the southeastern regions of Brazil with risks of prolonged droughts, this finding may suggest that the growth response of E. grandis trees to environmental conditions increases considerably during low water availability periods, regardless of the tree size and growth rate.
Effects of K and Na fertilization over extreme drought periods
Until the first 3 years of E. grandis tree plantation in the same experiment, K and Na addition improved stomatal regulation and enhanced osmotic adjustment over the study period accompanied by a reduction of soil water storage (Battie-Laclau et al. 2014). Moreover, high LAI and tendency of soil-to-leaf water potential gradient suggested that K and Na increase the risk of hydraulic dysfunction following extreme water deficiency (Battie-Laclau et al. 2014). Therefore, there is a concern if, under extreme water deficiency, the addition of K and Na would considerably increase the risk of tree death by an embolism (Battie-Laclau et al. 2014; Christina et al. 2015) or in lesser severity, decrease the tree growth. In our study, in older trees (3–5 years-old), we found that over an extreme drought period (2014 dry season), K and Na fertilization did not lead to mortality, and the cumulative basal area of K- and Na-fertilized trees over this period was similar to that of the control trees. The high responsiveness of K- and Na-fertilized trees to intra-annual rainfall variability showed that these trees can effectively respond by growing rapidly after rainy events and by completely stopping their cambial activity during long the driest periods. Some factors, reported in previous studies during the first 3 years after planting, can contribute to these growth patterns, such as the high stomatal conductance, optimal stomatal regulation, and high water-use efficiency for stem biomass production, performed by K and Na (Battie-Laclau et al. 2014, 2016). This factors may have accelerated the growth recovery as well as cambial inactivity. However, another important factor could be the alteration in fine roots density and its distribution in the soil profile in K- and Na-fertilized trees. Recently, in the same experiment, a simulation of K and throughfall effects using a MAESPA model found that fines roots of K + W and K − W trees can explore greater depths for water and reach the water table located at more than 16 and 18 meters deep in the soil, respectively (Christina et al. 2018). In fact, field studies proved that fine roots of fertilized E. grandis can seek water deeper than 10 m (Laclau et al. 2013; Germon et al. 2017). Moreover, combined effects of the fertilization and water deficit suggest a modification of fine roots configuration to increase the soil water access (Ewers et al. 1999; Samuelson et al. 2014).
Our study shows that an extreme drought season can affect the growth of trees independent of K or Na fertilization. Contrary to our hypothesis, over an extreme drought period as occurred in 2014 dry season, the effects of K and Na fertilization were less severe than expected because trees without K or Na application had similar cumulative basal area increment. On the other hand, as expected, during normal rainy and dry seasons, K and Na fertilization can increase the basal area growth by three to four-fold.
Simulation of future climate conditions suggests an increase in air temperature as well as changes in the amount and distribution of rainfall in South America (IPCC 2013; Chou et al. 2014). Eucalyptus plantations in Brazil are increasingly planted into areas with water deficiency and a higher risk of prolonged droughts (Gonçalves et al. 2013), where intensive fertilizer use is adopted to increase productivity (Gonçalves and de Barros 1999). In these contexts, understanding how fertilization is related to changes in growth responses to intra-annual meteorological variability and extreme drought conditions is important for the development of more appropriate management practices in these regions.
Conclusion
Our study found that K and Na fertilization increased the basal area growth by three to four-fold, and two to three-fold, respectively, over normal dry and rainy seasonal periods. As we expected, positive effects of K and Na fertilization were suppressed over a particularly dry season, characterized by extreme drought. However, contrary to our hypothesis, after these 6 months of the extreme drought conditions, K- and Na-fertilized trees reached a similar cumulative basal area increment to that of the control trees.
A 37% throughfall exclusion may not be enough to cause strong water stress in E. grandis trees and negatively affect the growth of trees over normal seasonal periods in southeast Brazil. The throughfall exclusion significantly decreased tree growth only under extreme drought. Because of stronger water reduction in − W plots and their high water consumption, K-fertilized trees were more affected than Na-fertilized trees and control trees, whose growth was not decreased.
Differences in growth patterns between treatments were related to responsiveness to intra-annual meteorological variability. Basal area growth of K-fertilized trees showed a strong relationship with rainfall, VPD, and minimum air temperature. However, the response of Na-fertilized and control trees to meteorological variables was relatively weak. Excluding the 37% throughfall considerably amplified the growth response of Na-fertilized and control trees to seasonal changes.
In areas with very deep tropical soils and annual rainfall < 1500 mm in southeast Brazil, our results suggest that prolonged droughts may affect the E. grandis tree growth independent of the K/Na fertilization regime. K and Na fertilized trees, in our experiment, could cease growth during the driest days and quickly recover their growth rates in response to small rainfall events. However, studies at a physiological level during extreme drought periods are necessary to better understand the effects of K and Na fertilization under future drought scenarios, particularly in a climate change context.
Supplementary material
Full data of the study: Basal area increment, rainfall, temperature (mean, minimum and maximum), vapor pressure deficit, and soil water content (0.15–1.5 m deep, and 3–6 m deep), collected in fortnightly periods, for each tree sampled by treatment of fertilization and water regime. http://dx.doi.org/10.17632/98fk3rvkrm.1.
References
Allen CD, Macalady AK, Chenchouni H et al (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Almeida JCR, Laclau JP, Gonçalves JLM et al (2010) A positive growth response to NaCl applications in Eucalyptus plantations established on K-deficient soils. For Ecol Manag 259:1786–1795. https://doi.org/10.1016/j.foreco.2009.08.032
Battie-Laclau PRF (2013) Efeitos da adubação potássica sobre a adaptação à seca do Eucalyptus grandis. Universidade de São Paulo
Battie-Laclau P, Laclau JP, Domec JC et al (2014) Effects of potassium and sodium supply on drought-adaptive mechanisms in Eucalyptus grandis plantations. New Phytol 203:401–413. https://doi.org/10.1111/nph.12810
Battie-Laclau P, Delgado-Rojas JS, Christina M et al (2016) Potassium fertilization increases water-use efficiency for stem biomass production without affecting intrinsic water-use efficiency in Eucalyptus grandis plantations. For Ecol Manag 364:77–89. https://doi.org/10.1016/j.foreco.2016.01.004
Beier C, Beierkuhnlein C, Wohlgemuth T et al (2012) Precipitation manipulation experiments-challenges and recommendations for the future. Ecol Lett 15:899–911. https://doi.org/10.1111/j.1461-0248.2012.01793.x
Binkley D, Stape JL, Bauerle WL, Ryan MG (2010) Explaining growth of individual trees: light interception and efficiency of light use by Eucalyptus at four sites in Brazil. For Ecol Manag 259:1704–1713. https://doi.org/10.1016/j.foreco.2009.05.037
Bošeľa M, Pajtík J, Konôpka B et al (2013) Modelling effects of weather condition on seasonal dynamics of the stem circumference increment in a mixed stand of Norway spruce and European beech. For J 59:180–188. https://doi.org/10.2478/v10114-011-0025-8
Cabral OMR, Gash JHC, Rocha HR et al (2011) Fluxes of CO2 above a plantation of Eucalyptus in southeast Brazil. Agric For Meteorol 151:49–59. https://doi.org/10.1016/j.agrformet.2010.09.003
Campinhos E Jr (1999) Sustainable plantations of high-yield shape Eucalyptus trees for production of fiber: the Aracruz case. New For 17:129–143. https://doi.org/10.1023/A:1006562225915
Campoe OC, Stape JL, Nouvellon Y et al (2013) Stem production, light absorption and light use efficiency between dominant and non-dominant trees of Eucalyptus grandis across a productivity gradient in Brazil. For Ecol Manag 288:14–20. https://doi.org/10.1016/j.foreco.2012.07.035
Campoe OC, Munhoz JSB, Alvares CA et al (2016) Meteorological seasonality affecting individual tree growth in forest plantations in Brazil. For Ecol Manag 380:149–160. https://doi.org/10.1016/j.foreco.2016.08.048
Castro VR, Surdi PG, Tomazello Filho M et al (2017) Efeito da disponibilidade hídrica e da aplicação de potássio e sódio no crescimento em diâmetro do tronco de árvores de Eucalyptus grandis. Sci For 45:89–99. https://doi.org/10.18671/scifor.v45n113.08
Chou SC, Lyra A, Mourão C et al (2014) Assessment of climate change over South America under RCP 4.5 and 8.5 downscaling scenarios. Am J Clim Chang 03:512–527. https://doi.org/10.4236/ajcc.2014.35043
Christina M, Le Maire G, Battie-Laclau P et al (2015) Measured and modeled interactive effects of potassium deficiency and water deficit on gross primary productivity and light-use efficiency in Eucalyptus grandis plantations. Glob Chang Biol 21:2022–2039. https://doi.org/10.1111/gcb.12817
Christina M, le Maire G, Nouvellon Y et al (2018) Simulating the effects of different potassium and water supply regimes on soil water content and water table depth over a rotation of a tropical Eucalyptus grandis plantation. For Ecol Manag 418:4–14. https://doi.org/10.1016/j.foreco.2017.12.048
Coelho CAS, Cardoso DHF, Firpo MAF (2016a) Precipitation diagnostics of an exceptionally dry event in São Paulo, Brazil. Theor Appl Climatol 125:769–784. https://doi.org/10.1007/s00704-015-1540-9
Coelho CAS, de Oliveira CP, Ambrizzi T et al (2016b) The 2014 southeast Brazil austral summer drought: regional scale mechanisms and teleconnections. Clim Dyn 46:3737–3752. https://doi.org/10.1007/s00382-015-2800-1
Cox PM, Betts RA, Collins M et al (2004) Amazonian forest dieback under climate-carbon cycle projections for the 21st century. Theor Appl Climatol 78:137–156. https://doi.org/10.1007/s00704-004-0049-4
de Almeida JCR (2009) Nutrição, crescimento, eficiência de uso de água e de nutrientes em povoamentos de Eucalyptus grandis fertilizados com potássio e sódio. Universidade de São Paulo
Dearden L (2015) Brazil hit by worst drought since 1930 as taps in Sao Paulo and Rio de Janeiro run dry|The Independent. In: Brazil hit by worst drought since 1930 as taps Sao Paulo Rio Janeiro run dry. http://www.independent.co.uk/news/world/americas/brazil-hit-by-worst-drought-since-1930-as-taps-in-sao-paulo-and-rio-de-janeiro-run-dry-10000234.html. Accessed 16 Feb 2017
Deslauriers A, Rossi S, Anfodillo T (2007) Dendrometer and intra-annual tree growth: what kind of information can be inferred? Dendrochronologia 25:113–124. https://doi.org/10.1016/j.dendro.2007.05.003
Downes G, Beadle C, Worledge D (1999) Daily stem growth patterns in irrigated Eucalyptus globulus and E. nitens in relation to climate. Trees Struct Funct 14:102–111. https://doi.org/10.1007/s004680050214
Ewers BE, Oren R, Albaugh TJ, Dougherty PM (1999) Carry-over effects of water and nutrient supply on water use of Pinus taeda. Ecol Appl 9:513–525. https://doi.org/10.2307/2641141
Fuller LG, Cattelino PJ, Reed DD (1988) Correction equations for dendrometer band measurements of five hardwood species. North J Appl For 5:111–113
Germon A, Guerrini IA, Bordron B et al (2017) Consequences of mixing Acacia mangium and Eucalyptus grandis trees on soil exploration by fine-roots down to a depth of 17 m. Plant Soil. https://doi.org/10.1007/s11104-017-3428-1
Gonçalves JLM, Benedetti V (2000) Nutrição e fertilização florestal. IPEF
Gonçalves JLM, de Barros NF (1999) Improvement of site productivity for short-rotation plantations in Brazil. Bosque 20:89–106
Gonçalves JLM, Stape JL, Laclau J-P et al (2004) Silvicultural effects on the productivity and wood quality of eucalypt plantations. For Ecol Manag 193:45–61. https://doi.org/10.1016/j.foreco.2004.01.022
Gonçalves JLM, Alvares CA, Higa AR et al (2013) Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalypt plantations. For Ecol Manag 301:6–27. https://doi.org/10.1016/j.foreco.2012.12.030
IBÁ (2017) Relatório IBÁ 2017, São Paulo
IPCC (2013) Climate change 2014 : synthesis report. Contribution of Working Groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change. Geneva, Switzerland
Keeland BD, Sharitz RR (1993) Accuracy of tree growth measurements using dendrometer bands. Can J For Res 23:2454–2457. https://doi.org/10.1139/x93-304
Knott R (2004) Seasonal dynamics of the diameter increment of fir (Abies alba mill.) and beech (Fagus sylvatica L.) in a mixed stand. J For Sci 50:149–160
Laclau JP, Almeida JCR, Gonalves JLM et al (2009) Influence of nitrogen and potassium fertilization on leaf lifespan and allocation of above-ground growth in Eucalyptus plantations. Tree Physiol 29:111–124. https://doi.org/10.1093/treephys/tpn010
Laclau JP, Ranger J, Gonçalves JLM et al (2010) Biogeochemical cycles of nutrients in tropical Eucalyptus plantations. Main features shown by intensive monitoring in Congo and Brazil. For Ecol Manag 259:1771–1785. https://doi.org/10.1016/j.foreco.2009.06.010
Laclau J-P, da Silva EA, Rodrigues Lambais G et al (2013) Dynamics of soil exploration by fine roots down to a depth of 10 m throughout the entire rotation in Eucalyptus grandis plantations. Front Plant Sci 4:1–12. https://doi.org/10.3389/fpls.2013.00243
Lin Y-S (2012) How will Eucalyptus tree species respond to global climate change? A comparison of temperature responses of photosynthesis. University of Western Sydney, Sydney
Nobre CA, Marengo JA, Seluchi ME et al (2016) Some characteristics and impacts of the drought and water crisis in southeastern Brazil during 2014 and 2015 some characteristics and impacts of the drought and water crisis in southeastern Brazil during 2014 and 2015. J Water Resour Prot 8:252–262. https://doi.org/10.4236/jwarp.2016.82022
Samuelson LJ, Pell CJ, Stokes TA et al (2014) Two-year throughfall and fertilization effects on leaf physiology and growth of loblolly pine in the Georgia Piedmont. For Ecol Manag 330:29–37. https://doi.org/10.1016/j.foreco.2014.06.030
Schippers P, Sterck F, Vlam M, Zuidema PA (2015) Tree growth variation in the tropical forest: understanding effects of temperature, rainfall and CO 2. Glob Chang Biol 21:2749–2761. https://doi.org/10.1111/gcb.12877
Sette CR Jr, Tomazello Filho M, dos Santos Dias CT, Laclau JP (2010) Crescimento em diâmetro do tronco das árvores de Eucalyptus grandis W. Hill. ex. Maiden e relação com as variáveis climáticas e fertilização mineral. Rev Árvore 34:979–990. https://doi.org/10.1590/S0100-67622010000600003
Sette CR Jr, Tomazello Filho M, Lousada JL, Laclau JP (2012) Seasonal increment in trunk diameter of Eucalyptus grandis tress applying dendrometer bands. Cienc Florest 22:763–775
Sevanto S, Nikinmaa E, Riikonen A et al (2008) Linking xylem diameter variations with sap flow measurements. Plant Soil 305:77–90. https://doi.org/10.1007/s11104-008-9566-8
Stoneman GL, Crombie DS, Whitford K et al (1997) Growth and water relations of Eucalyptus marginata (jarrah) stands in response to thinning and fertilization. Tree Physiol 17:267–274
Vitas A (2011) Seasonal growth variations of pine, spruce, and birch recorded by band dendrometers in ne Lithuania. Balt For 17:30–35
Ward EJ, Domec JC, Laviner MA et al (2015) Fertilization intensifies drought stress: water use and stomatal conductance of Pinus taeda in a midrotation fertilization and throughfall reduction experiment. For Ecol Manag 355:72–82. https://doi.org/10.1016/j.foreco.2015.04.009
White DA, Crombie DS, Kinal J et al (2009) Managing productivity and drought risk in Eucalyptus globulus plantations in south-western Australia. For Ecol Manag 259:33–44. https://doi.org/10.1016/j.foreco.2009.09.039
Williamson GB, Laurance WF, Oliveira AA et al (2000) Amazonian tree mortality during the 1997 El Nino drought. Conserv Biol 14:1538–1542. https://doi.org/10.1046/j.1523-1739.2000.99298.x
Zeng S, Jacobs DF, Sloan JL et al (2013) Split fertilizer application affects growth, biomass allocation, and fertilizer uptake efficiency of hybrid Eucalyptus. New For 44:703–718. https://doi.org/10.1007/s11056-013-9371-y
Acknowledgements
We gratefully acknowledge Rildo M. Moreira and the staff of Itatinga Research Station (ESALQ/USP) as well as Eder Araujo da Silva (http://www.floragroapoio.com.br), for their technical support. We also thank Jean-Paul Laclau and Juan Sinforiano Delgado-Rojas for sharing the soil water content data and meteorological data of the experimental plantation.
Funding
This research was funded partially by Agropolis Fondation under the reference ID1203-003 through the «Investissements d’avenir» programme (LabexAgro/ANR-10-LABX-0001-01), National Council for Scientific and Technological Development (CNPq) (Process Numbers 170461/2017-3, 444793/2014-3 and 014/2012), Fundação de Amparo à Pesquisa do Estado de São Paulo (Fapesp) (2013/25642-5) and Fondo Nacional de Desarrollo Científico, Tecnológico y de Innovación Tecnológica (FONDECYT-CONCYTEC, Perú) (Award Number 239-2018).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chambi-Legoas, R., Chaix, G. & Tomazello-Filho, M. Effects of potassium/sodium fertilization and throughfall exclusion on growth patterns of Eucalyptus grandis W. Hill ex Maiden during extreme drought periods. New Forests 51, 21–40 (2020). https://doi.org/10.1007/s11056-019-09716-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-019-09716-x