
Abstract
Many second growth forests have poor regeneration and in some spatial and economic context, such as post-agricultural forests, it may be beneficial to increase tree diversity. We planted seedlings of yellow birch (Betula alleghaniensis) and bur oak (Quercus macrocarpa) in strip openings of 3, 6 and 9 m width to control light. Individual mesh sleeves were used to reduce browsing pressure but they also reduced available light for seedlings: a large mesh and a small mesh sleeves were tested in each opening. Available light for seedlings ranged from 5 to 46% of full sunlight. First year growth was almost non-responsive to all treatments for both species, but birch showed strong responses during second year. Birches in the large mesh sleeve and the 9 m opening had the largest height and diameter increments, while those in the small mesh sleeve and the 3 m opening had the smallest increments. Small mesh sleeves increased the height over stem diameter ratio for both species. Treatments affected several other morphometric variables of birch but none of the oak. More time is needed to assert if bur oak can be a candidate species for enrichment planting. We recommend strip openings of 9 m width and large mesh sleeves, for protection against deer browsing, while controlling neighbouring competition, to promote artificial regeneration of hardwood species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Recolonization of abandoned land by forests is a phenomenon quite frequent and can be seen in a wide variety of climates (Bouchard and Domon 1997; Buisson et al. 2006; Bowen et al. 2007). Many of these lands were previously used for human activities, such as pastures or crops. In these cases, natural regeneration can sometimes lead, through succession, to healthy ecosystems and tree communities (Benjamin et al. 2005; Bowen et al. 2007). However, succession often will be heavily altered in these post-agricultural forests: recolonization is limited to wind disperse species and succession is stopped to its first stage. Many of these forests are surrounded by agricultural fields thus, distance from mature stand reduces the propagation of new seeds (Butaye et al. 2001; D’Orangeville et al. 2008) and disturbances from past human activities impede recruitment of new species (Cramer et al. 2008; Brown and Boutin 2009). This pattern can be seen in many post-agricultural forests across the world (Butaye et al. 2001; Cramer et al. 2008; D’Orangeville et al. 2008; Brown and Boutin 2009). Regeneration will often depend on the level of human disturbance. Lower levels, such as pasture or ancestral farming which have little impact on the soil, will often follow a natural trajectory without further human intervention (Benjamin et al. 2005; Cramer et al. 2008). Nevertheless, higher level disturbance, such as intensive agriculture on large area will almost every time develop to poor communities with low diversity (Cramer et al. 2008). Low biodiversity makes these forests vulnerable to disturbances, such as diseases or strong meteorological events (Dey et al. 2012; Sánchez-Pinillos et al. 2016). Moreover, the habitat may be inadequate for many animal species and little ecosystem services are provided compared to a more mature forest (Hansen et al. 1991; Bowen et al. 2007). Without human intervention, some forests could take more than an 100 years before seeing some recruitment of late successional species (Oosting 1942).
Many forests in North America and Europe are second growth forests, and a good proportion of them are on abandoned agricultural lands. Moreover, these forests are often close to urban areas (Ramankutty and Foley 1999; Verheyen et al. 1999; Drummond and Loveland 2010). The abundance and the proximity to our living space of this type of forests raise the importance of understanding the modalities of seedlings’ early growth for an efficient management, if we are to keep forested areas and their ecological services.
Therefore, some post-agricultural forests must be managed to increase their resilience and keep and improve the existing ecosystem services. A direct economic value could be added by managing with the aim of producing timber over time, which would stimulate interventions (Benjamin et al. 2008). This is particularly true when targeting forests nearest urban center and wood transformation industries, due to a lower abundance of forests in these areas, thus a greater potential for increasing local biodiversity and reduced transportation costs. Such an added value could help preserve forest and reduce their potential return to agriculture. The choice of which forest should be managed must be weighed carefully, as some of them represent a transitional stage and offer a somewhat rare habitat for many species (Bowen et al. 2007).
There are two factors known to be major constraints everywhere on the success of artificial and natural regeneration of mid to late successional species: light and competition (van Kuijk et al. 2014; Stanturf et al. 2014). Light levels must be high enough to ensure a vigorous growth of seedlings while restraining competitive, fast growing, species. Control of the overstory alone cannot achieve this goal, as there may be strong competition in the understory as well (Balandier et al. 2006). Therefore, interventions are required to control that competition (Dubois et al. 2000).
In recent years, another factor stalling both artificial and natural regeneration has reached an alarming threshold: deer browsing. This affects mostly North America and Europe, where deer will eat seedlings, to the point where some species will never reach the canopy and others will take many more years than they usually do (Kuiters and Slim 2002). Selective browsing might favour unwanted species and counter measure adds a constraint: protective sleeves are required around seedlings but they hinder light reception. Even though there is a light reduction, most type of protection induce an increase in height growth (Ward et al. 2000; Ponder 2003). However, such effects are reduced when there is over 55% of the canopy (Walters 1993; DuPlissis et al. 2000). Few studies were available about the use and effects of shelters under varying canopy and even fewer about the use of mesh sleeves under canopy openings.
Enrichment underplanting in strip openings is a way of increasing light levels for planted trees. By using this method, it is possible to increase forest biodiversity while maintaining part of the environment untouched, thus keeping the ecosystem services provided and reducing successional setback probabilities (Stanturf et al. 2014). Remaining trees will provide several benefits, such as shelter against wind, heavy snow and strong sun, or reduce soil temperature drop during winter (Langvall and Löfvenius 2002; Dey et al. 2012). Deer protections have been used in the past in open areas and a few times in mature forests in combination with underplanting (Dubois et al. 2000; Pinna et al. 2012). While there are many papers detailing the effects of protective sleeves on growth of seedlings, most of them are either about tube shelters and/or in an open area. Tube shelters give little to no benefits to growth under forested cover, while open areas have different light environments (Walters 1993; DuPlissis et al. 2000). Low light, induced by canopy is a condition reducing seedlings’ growth. Browsing pressure makes protections a necessity for many areas in the northern hemisphere. However, tube shelters, a common type of protection, are often mentioned to increase establishment and growth of seedlings under high levels of light (Taylor et al. 2006; Andrews et al. 2010; Mariotti et al. 2015). Mesh sleeves are another type of protection, less restrictive to light, which have some beneficial effects on growth (Walters 1993). Tube shelters being too restrictive for use in forests, is there a forest cover density (i.e. light intensity) under which mesh shelters could be useful to enhanced seedlings’ early growth?
The aim of this study was to compare seedlings establishment and early growth of two hardwood species under different light environments, in a post-agricultural forest, using (1) different width of strip openings and (2) different mesh size of protective sleeves (instead of tube shelters) against browsing. The two species, yellow birch (Betula alleghaniensis Britt.) and bur oak (Quercus macrocarpa Michx.), are mid-tolerant species. Yellow birch is often seen in a gap dynamic in mature forests, whereas bur oak is sometimes seen as pioneer but also in mature forests (Burns and Honkala 1990). We hypothesize that an intermediate light level (under 50%, but above 25%PAR), provided by one or few of our combinations will have the best success: low enough to slow competitors’ growth but high enough to ensure seedlings’ early development.
Methods
Study area
Study site was a post-agricultural forest located near Montreal (QC, Canada, 45°40′34″ north, 73°19′19″ west), in a sugar maple-hickory (Acer sacharrum–Carya cordiformis) zone (Grandtner 1966). Mean annual temperature is 6.6 °C and precipitation average 983 mm per year of which 186 fall as snow, with an average of 457 mm during growing season from may to September (Environment Canada 2010). The soil is loamy sand to sand with an average pH of 4.9. Fertility is low, but the water table is high, at less than 1 m. Deer often visit the grounds (7–12/km2), leading to a high browsing pressure (Huot and Lebel 2012). The stand is composed of red maple (Acer rubrum L.) and grey birch (Betula populifolia Marbo.), with few trembling aspen (Populus tremuloides Michx.) and black cherry (Prunus serotina Ehrh.), with a mean age of 26 years old. Seedlings and saplings were 92 and 89% red maple, respectively, emphasizing the need for a greater diversity. The most abundant herbaceous species are bristly dewberry (Rubus hispidus L.), blackberry (Rubus alleghaniensis Porter), Canada mayflower (Maienthemum canadense Desf.) and broadleaves meadowsweet (Spiraea latifolia Borkh.).
Experimental design
In each of four blocks, one strip of 3, 6 and 9 m width, separated by 6 m of untouched forest, were randomly positioned with a north/south orientation, during autumn of 2013, few months before the plantation in the following spring of 2014. Distance between blocks ranged from 6 to 300 m. In each strip, three sections were assigned a different protection sleeve. Two commercially available protections were tested: VexarMD (MasterNet LTD Mississauga, ON, Canada) (called hereafter “large mesh”) and ClimaNetMD (Nortene technologies, Lomme, France) (called hereafter “small mesh”). The third type was built using chicken wire and aimed to be a control maximising light transmission and lessening physical stress while also protecting the stem. All vegetation was cut down in a 50 cm radius around seedlings in August, using machetes, for the first two growing season. We made sure no branches were shading seedlings. Competition was mostly sprouts from stumps in the strips and shrubs that took advantage of increased light. It reached around 80 cm high the first year and over 1.5 m the second. Any given section had four groups of two trees, one per species, which were yellow birch and bur oak. Four trees for each combination of treatments were planted in a line at the center of the opening with a spacing of 2 m. Bur oaks were bare root seedlings of 2 years while yellow birches were 1 year seedlings from containers. Mean initial height and standard deviation were 56 ± 10 cm and 52 ± 15 cm for yellow birch and bur oak respectively.
Yellow birch is a species common in mature forests of eastern North America. It has a fast growth when young, which slows later, as it follows a gap dynamic in forests of Acer saccharum Marsh. and Fagus grandifolia Ehrh. The species has been in decline for many years now (Ward and Stephens 1997). Bur oak has a slower growth, but usually a longer lifespan and can tolerate a good range of hydrologic conditions (Abrams 1996). His fruits are a food source for many animal species. Both species are indigenous in the study area and are of good economic yield.
Protectors
Mesh sleeves are tube like shelters of 30 cm in diameter, with walls made of plastic mesh, similar to a fence. The large mesh sleeves were made of green plastic, with a square mesh of 2.5 cm per side, while the small mesh sleeves were made of black plastic with a square mesh of 2 mm per side. The control was a metal square mesh of 2.5 cm per side. The large mesh and the control were 140 cm high, while the small mesh was 80 cm, but was raised several times to stay ahead of the top leader until reaching a height of 140 cm. Browsing by deer (Odocoileus virginianus) occurs mostly under 150 cm, but depends on the density of deer population (Dubois et al. 2000; Paquette et al. 2006a; Oswalt et al. 2006).
Sampling
We measured photosynthetic active radiation (PAR), from 400 to 700 nm, at 0.05, 0.75, 1.25 and 2 m, year one and year two, using sunshine sensor (BF2 and BF5 Sunshine Sensor, Delta-T, Cambridge, UK). For each forest measure, another one was taken simultaneously in an open field nearby. The field was an open space without any obstacle for at least 100 m in any direction. It allowed the creation of a ratio of PAR (%PAR), which is a good estimator of total light received during the growing season (Parent and Messier 1996; Paquette et al. 2007). This measure gives the average %PAR received during the whole season, but does not give information about daily variations, such as cloud cover or sun flecks. It does take into account an opening, if there is one. Diffuse light is increased in an opening, even thought the sun is not shining directly in the sample point.
The light attenuation inside each protector and inside the control was measured in an open space with quantum sensor (LI-190, LI-COR, Lincoln, Nebraska, USA), using the overcast method (Parent and Messier 1996). We then applied these light attenuations to the values measured at 2 m to calculate the %PAR received by seedlings.
Seedlings diameter and height increments were measured on each individual by subtracting year 1 from year 2, while additional measurements were taken on a subsample (2 per treatments and per species, on 3 blocks, n = 108): shoot growth, chlorophyll concentration (atLEAF+, FT Green LLC, Wilmington, USA), leaf area, length of secondary branches, and crown length. Using these variables, several ratios were made to monitor seedling’s morphology: height/stem diameter (H/D ratio), annual shoot growth/mean length of secondary branches (extension growth index), height/sum of secondary branches, leaf area/sum of secondary branches (leaf display), crown length/stem diameter and crown length/stem length (living crown). Leaf area was calculated with leaf length, using a regression of leaf blade length with leaf area (Delta-T area meter mk2, Delta T Devices LTD, Cambridge, UK). The regression was built using 36 and 42 leaves, for yellow birch and bur oak respectively, from extra seedlings planted between the openings: seedlings were 3 m away from an opening, on both sides. We made sure to have a wide range of leaf size to include any size that could be found on the seedlings in the openings.
Environmental variables (light, stand characterization and air temperature) were sampled using six transects made at regular intervals, perpendicular to the openings. Light measurements taken in transects were only used for comparison between openings and untouched forest for both year one and year two. They were taken in the forest, 15 m from each side of a block, in the openings and between the openings, all at a height of 1.25 m. Stand vegetation was characterized using the point-centered quarter method (Mitchell 2007) (n = 60, tree DBH130: θ > 5 cm, seedling DBH130: 1 cm < θ < 5 cm). Herbaceous vegetation was described using the species with cover superior to 50% on 1 m2 square (n = 60). Air temperature was measured every half hour for 82 days, at a height of 80 cm, in the middle of the openings from June to August of the second year, using data loggers (DS1922L, iButton, Maxim integrated, CA, USA) protected from direct sunlight by a green plastic plate, 20 cm above. We made sure the sensors were always under shaded conditions.
Analysis
We used a three-way ANOVA to compare growth and light level across treatments, using protections and opening width as fixed factors and blocks as random factor, and Tukey HSD for multiple means comparison. Chi square tests were used for survival. Some data were transformed to reach normality. Linear and logarithm regressions were made using growth variables and light. Most of the analyses were made with JMP v10.0.0 (SAS Institute, Cary, USA) but for the regressions (R v3.0.2, R Development Core Team, Vienna, AUS).
Results
Light distribution under treatments
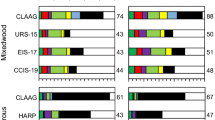
All openings increased available light compared to the untouched forest, from 5 to 10 times more light (Table 1). There was a decrease in %PAR from year one to year two, faster in the smaller openings, reducing by 50% in the 3 m width, 39% in the 6 m width and 22% in the 9 m width (see measurements at 2 m high, Table 1). Light available for a seedling is further reduced by the use of protective sleeve. Small mesh allows 34% of the light to get through while large mesh and control allow 74% (Fig. 1). Vegetation competition reduced light reaching the ground from 2 m high by 42 up to 75% PAR, from the 3 m openings to the 9 m openings (Table 1).

Mean % of PAR during growing season inside sleeves (integrated for the whole sleeve), for each width of opening. Year 1: opening effect p = 0.0015; sleeve effect p < 0.0001 Year 2: opening effect p < 0.0001; sleeve effect p = 0.0001. Effects were the same for both years. Values followed by the same letter are not significantly different (alpha = 0.05). Small mesh (S), large mesh (L), control (C)
Survival
Small mesh sleeves provided yellow birch with a better survival (94%) than the large mesh (79%) and the control (75%) (Fig. 2). There were no interactions between the opening width and the sleeves. Survival of bur oak seedlings was slightly lower with the large mesh (92%) compared to the other (97%) and the control (100%). Openings width had no effect on survival, for both species (Fig. 2).

Survival rate of yellow birch and bur oak after two growth seasons under different openings’ width and protective sleeves. Yellow birch: openings effect p = 0.57, sleeve effect p = 0.0138, interaction p = 0.1682. Bur oak: openings effect p = 0.4627, sleeve effect p = 0.0457, interaction p = 0.8862. Values followed by the same letter are not significantly different (alpha = 0.05). Small mesh (light grey), large mesh (dark grey), control (black)
Growth
Data from the first year are not shown, as there was much uncertainty during the first growth season, for both species and all treatments due to stress of transplantation. For each growth variable analyzed, no interaction has been detected for yellow birch or bur oak between protective sleeves and openings, for the second year. During that same year, yellow birch shoot and diameter growth increased with the size of opening, without affecting the H/D ratio (Table 2). Diameter growth seemed more sensitive to openings than shoot growth, as differences were significant for all size, while only between the 9 m and the two others for shoot growth. Small mesh sleeve reduced birch’s growth for the same two variables in all openings compared to control (Table 2). It also reduced final height and shoot growth, although it increased the H/D ratio. Treatments had no effect on shoot growth and final height of bur oak, but larger openings had a greater diameter increment, for all protections, while small mesh reduced diameter increment and increased H/D ratio, for all openings (Table 2).
Seedlings morphology and chlorophyll concentration
Treatments had no significant direct effects on bur oak morphology. Opening width and large mesh sleeve increased chlorophyll concentration for yellow birch. Small mesh sleeves increased crown length/stem diameter ratio for birch, but reduced the total length of secondary branches. While treatments had no significant effects on the other ratios, some were quite close (0.05 < p < 0.1): leaf area, leaf display and number of secondary branches had a tendency to be decreased by small mesh sleeves while height/total length of secondary branches was increased by that same protection (Table 3).
Taking only light measurements into account allowed several regressions to be seen, some that were otherwise not detected: yellow birch increased the length of its secondary branches (R2 = 0.22), total leaf area (R2 = 0.156), leaf display (R2 = 0.129) and diameter (R2 = 0.318), while bur oak increased its extension growth index (R2 = 0.105), total leaf area (R2 = 0.147) and diameter (R2 = 0.188) (Table 4).
Temperature
Wider openings meant higher air temperature, close to 5 °C more than in the control forest in the afternoon. Temperature differences between openings were minimal before 10:30 in the morning and after 18:00 and reached their highest at 14:00 (Fig. 3).

Mean air temperature for 24 h at 80 cm height, by opening, data from June to August 2015
Discussion
Light levels
Paquette et al. (2006b) in a meta-analysis identified the interval between 25 and 50%PAR to be best for survival and growth of trees under residual cover. The high end of this interval has shown better growth results (Beaudet and Messier 1998; Gasser et al. 2010; Kern et al. 2012). With an optimal growth around 36–46%PAR inside sleeves, our results support this conclusion for post-agricultural forests when using protective mesh sleeves, in a temperate climate. Many experiments on different protector types, if not almost all of them, were made in open areas, either fields or large forest gaps. Therefore, light intensity was never low, the only reduction being the protector. These experiments showed some success for mesh sleeves, increasing their height growth (Ward and Stephens 1995; Ward et al. 2000; Devine and Harrington 2008). Some studies on natural regeneration did have low light intensity (Beaudet and Messier 1998; Forrester et al. 2014), but ours might be one of the first to have a wide range of %PAR using protections in a forested environment.
Over time, openings will start closing and change the light environment (Cogliastro et al. 2006). Canopy closure is faster when comprised of pioneer species than in a more mature forest (Valverde and Silvertown 1997) and should be considered when planning enrichment under-planting. Our results show that small openings may allow enough %PAR for the first year only with protective sleeves, as canopy closes. While small openings are not a long-term option, there is a setback to wider strip opening: we have demonstrated that wider strips promote vegetation competition in young post-agricultural forest (Table 1). Pioneer species allow a greater light input through the canopy (Canham 1988), therefore competition should thrive compared with a more mature forest (Beaudet and Messier 1998; Cogliastro and Paquette 2012). Hence, it may require particular care, specifically around slow growing tree species like bur oak. Bur oak reached its photosynthetic maximum in the 6 and 9 m opening while yellow birch was close to it only in the 9 m opening.
Survival
Light intensity had some influence on survival and growth pattern of both species. What was less expected is that sleeves were more influential than openings. Proportionally, light reduction was greater by the small mesh sleeve in a given opening than between the 9 and 3 m openings, which would explain observing effects of sleeves but not openings, for survival and several growth variables.
The higher survival for yellow birch seedlings in the small mesh sleeve is surprising, as it greatly reduced light and was not observed with bur oak, even though both species are mid tolerant to shade (Niinemets and Valladares 2006). However, it has been noticed during fieldwork (personal observation) that almost no vegetation was able to grow inside the small mesh sleeve, while the other protection allowed a somewhat stronger competition. It appears that the %PAR, under the combined shades casted by the seedling and the small mesh, were low enough to inhibit all growth: 24% of available PAR reaches the bottom quarter of the sleeve without a seedling inside, while this number increases to 67% for the large mesh. A difference in survival rate was expected between the openings, as our range of %PAR was wide, yet none was seen. Competition was stronger with wider opening and since its control was done late in the growing season, light disparities may have been reduced to a level were no difference in survival could be seen. Maybe, considering that species are of intermediate shade tolerance, our lowest level (11%PAR the first year and 5% the second) was still high enough to allow survival of most individuals.
Seedlings growth
Decreasing forest cover to sustain seedlings regeneration is a technique often used, but usually restrained to natural regeneration and uncommon in combination with planting of deciduous tree species. Therefore, few studies on the growth of underplanted North American hardwood trees are available for comparison. Nevertheless, we compared our results to assisted regeneration studies and noticed that the height growth of yellow birch was greater for the same amount of %PAR, sometimes two to three times yet with an increased H/D ratio (Lorenzetti et al. 2008; Kern et al. 2012; Yang et al. 2012). Amongst studies, deer protections are often deficient or lacking and browsing reduces growth and drains resource storage over time (Kern et al. 2012). Similar growths have been observed in natural regeneration with the same light conditions (Beaudet and Messier 1998; Ricard et al. 2003), although they selected older and taller seedlings that were above competitors and had very few data for high range of %PAR. Protective sleeves may have enhanced height growth, since we observed the same growth as assisted regeneration, without choosing the best seedlings, by funnelling the trees towards the top of the sleeve. Microclimate in tree shelter is known to increase height gain, but its effects are reduced by ventilating tubes and almost non-existent when using mesh sleeves (Bergez and Dupraz 2000; Devine and Harrington 2008). Another factor of importance affecting growth patterns are mechanical stimuli. Coutand et al. (2008) reported that the absence of mechanical stimuli increases height growth, at the cost of diameter growth, and that wind rustling through the leaves could be a sufficient stimulus. Therefore, our protective sleeves may have allowed enough wind to create mechanical stimulation, but for the small mesh sleeve, which has a higher H/D ratio. The large diameter of our protections allowed enough room for some trunk movements.
Growth should vary over a wide range of %PAR (Beaudet and Messier 1998; Ricard et al. 2003; Paquette et al. 2006b; Lof et al. 2007; Gasser et al. 2010) but we did not see such changes, even though %PAR had a wide range in our experiment (5–46%PAR). The proliferation of competition in wider openings might have reduced light disparities, hence a range of %PAR smaller than what was measured (our light measures were taken above vegetation). However, weed can provide protection against harsh condition such as snow and ice storm, a role that may have been taken, in our case, by the protective sleeves, by providing a physical protection (Close et al. 2009; Puértolas et al. 2010). Protective mesh sleeves may also affect morphology, by modifying the H/D ratio. In comparison with the present work, most studies observed a lower H/D ratio for every level of light (Lorenzetti et al. 2008; Kern et al. 2012), but not all (Beaudet and Messier 1998). Most of them also had no protective sleeves: the %PAR increases with height inside a protective sleeve, therefore compelling seedlings toward a faster height growth, detrimental to the diameter. Several studies reported such results for tube shelters (DuPlissis et al. 2000; Ward et al. 2000; Ponder 2003). Every oak could stand on his own without their protection, while over 75% of the yellow birches could, for each treatment, and only a small curve was visible for almost a quarter of the yellow birches (data not shown). Yellow birches that completely toppled without their protection represent less than 2% of total number of birches. Competition may have had a similar effect, as it reduces %PAR close to the ground, but not as much at the top, adding to the variation of H/D ratio between protective sleeves. Competition also has another effect, by reducing the red/far red ratio (R/FR), which then stimulates height growth (Ballaré et al. 1990; Schmitt and Wulff 1993). The advantage of mesh sleeves in this case is that they trap less FR inside the sleeve (Laliberté et al. 2008), while it is a condition known to tube shelters (Bergez and Dupraz 2009).
Results for bur oak’s H/D ratio were slightly different from yellow birch because seedlings grew slower and were smaller, thus affected only by the bottom portion of the sleeve, where disparities in light reduction were stronger. The large mesh intercepted more light at its bottom than the control, but less than the small mesh and is thus positioned between it and the small mesh in terms of H/D ratio. Moreover, yellow birch has a maximum photosynthesis around 600 µmol m−2 s−1 while it is close to 400 µmol m−2 s−1 for bur oak, meaning it could take advantage of increased light when bur oak could not (Logan 1970; Hamerlynck and Knapp 1994; Beaudet et al. 2000).
Diameter growth was more sensitive to treatments, for both species. In the larger opening, larger diameters could be explained by a less constraining light, which allowed seedlings to increase allocation to diameter and roots instead of crown. Moreover, increased light is linked with an increase in air temperature and soil temperature, which may stimulate underground activities such as bacterial communities and mycorrhizas, favouring root growth (Cheng et al. 2005; Song and Cheng 2010).
While the H/D ratio of yellow birch was high, several variables showed that growth was sustained, and without too much etiolation. The small mesh sleeves increased the crown/stem diameter ratio for yellow birches, meaning that with the other protection, which allows more light, more resources are allocated towards diameter and less toward the crown, hence the lower H/D ratio. Leaf display correlated to light clearly indicates that with higher %PAR, height growth is sustained by a stronger photosynthesis potential. Moreover, photosynthesis per leaf area also increased, as chlorophyll content showed, hence the larger opening increased overall growth and not only biomass allocation. An increase in aerial biomass is usually sustained by a more developed root system (Wayne and Bazzaz 1993).
Canopy is closing relatively fast and thus the transmitted %PAR will follow. However, seedlings will eventually grow outside of their protection and leave the limited light environment it represents. Also, light requirement tends to increase with tree age (Messier and Nikinmaa 2000). Hence, if their height growth is fast enough, it is plausible to think that the first intervention to open the canopy will be sufficient for the 9 m opening, because it will maintain a relatively stable %PAR at the crown of trees, during their growth.
Treatments seemed to have little to no effects on the morphometric measurements and survival rate of bur oak. However, it is a species known to build a strong root system and 2 years may not have been enough to see such development (Bragg et al. 1993; Rebbeck et al. 2011). Also, shoot dieback is often observed on bur oak, which consequently helps to increase root to shoot ratio, thus survivability (Johnson et al. 2009). This would explain the slow growth and the high survival rate seen in this study. Moreover, it may have suffered more from light competition than yellow birch, which has a faster height growth and will get outside the influence of neighbouring competition.
Nonetheless, a few growth parameters of bur oak reacted to light, as observed before (Laliberté et al. 2008). Therefore, its growth could be enhanced with the correct light environment, although focus could be put elsewhere, as many oak species have been shown to react more strongly to other variable than strictly light, such as water content and microsite (Collins and Battaglia 2008).
Conclusions
Seedlings establishment can be facilitated by the use of protective mesh sleeves under canopy strip openings. However, several conditions must be met to obtain successful growth.
-
1.
Mesh sleeves must be permissive to light: we saw only increase in growth with higher %PAR, with nothing to suggest that high %PAR could be detrimental to seedlings’ growth.
-
2.
Openings of 9 m are a good compromise between increased seedlings’ growth and stronger competition. Wider openings might increase light induced stress, such as leaf transpiration, and competition’s growth, thus, loosing the advantage of increased light.
-
3.
For slow growing species, like bur oak, a strict control of competition is necessary to ensure that seedlings are not overtopped by neighbouring vegetation.
-
4.
The choice of planted species should be carefully thought. Species should have at least some tolerance to shade, as canopy closure will reduce incident light, as well as a fast height growth, to reduce the frequency of competition control. For species with slower growth, a second thinning may be required after few years.
-
5.
Small mesh sleeves may be adapted for restoration of underplanted yellow birch, as they increased survival however, the slower growth in these protector is not an option for wood production and long term survival needs to be studied.
References
Abrams MD (1996) Distribution, historical development and ecophysiological attributes of oak species in the eastern United States. Ann For Sci 53:487–512
Andrews DM, Barton CD, Czapka SJ, Kolka RK, Sweeney BW (2010) Influence of tree shelters on seedling success in an afforested riparian zone. New For 39(2):157–167
Balandier P, Collet C, Miller JH, Reynolds PE, Zedaker SM (2006) Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 79:3–27
Ballaré CL, Scopel AL, Sánchez RA (1990) Far-red radiation reflected from adjacent leaves: an early signal of competition in plant canopies. Science 247(4940):329–332
Beaudet M, Messier C (1998) Growth and morphological responses of yellow birch, sugar maple, and beech seedlings growing under a natural light gradient. Can J For Res 28:1007–1015
Beaudet M, Messier C, Hilbert DW, Lo E, Wang ZM, Lechowicz MJ (2000) Leaf-and plant-level carbon gain in yellow birch, sugar maple, and beech seedlings from contrasting forest light environments. Can J For Res 30(3):390–404
Benjamin K, Domon G, Bouchard A (2005) Vegetation composition and succession of abandoned farmland: effects of ecological, historical and spatial factors. Landsc Ecol 20(6):627–647
Benjamin K, Bouchard A, Domon G (2008) Managing abandoned farmland: the need to link biological and sociological aspects. Environ Manage 42(4):603–619
Bergez JE, Dupraz C (2000) Effect of ventilation on growth of Prunus avium seedlings grown in treeshelters. Agric For Meteorol 104(3):199–214
Bergez JE, Dupraz C (2009) Radiation and thermal microclimate in tree shelter. Agric For Meteorol 149(1):179–186
Bouchard A, Domon G (1997) The transformation of the natural landscapes of the Haut-Saint-Laurent (Québec) and their implications on future resource management. Landsc Urban Plann 37:99–107
Bowen ME, McAlpine CA, House APN, Smith GC (2007) Regrowth forests on abandoned agricultural land: a review of their habitat values for recovering forest fauna. Biol Conserv 140:23–296
Bragg WK, Knapp AK, Briggs JM (1993) Comparative water relations of seedlings and adult Quercus species during gallery forest expansion in tallgrass prairie. For Ecol Manage 56:29–41
Brown CD, Boutin C (2009) Linking past land use, recent disturbance, and dispersal mechanism to forest composition. Biol Conserv 142:1647–1656
Buisson E, Dutoit T, Torre F, Römermann C, Poschlod P (2006) The implications of seed rain and seed bank patterns for plant succession at the edges of abandoned fields in Mediterranean landscapes. Agr Ecosyst Environ 115(1–4):6–14
Burns RM, Honkala BH (1990) Silvics of North America: 2. Hardwoods, agriculture handbook 654, vol 2. US Department of Agriculture, Forest Service, Washington
Butaye J, Jacquemyn H, Hermy M (2001) Differential colonization causing non-random forest plant community structure in a fragmented agricultural landscape. Ecography 24:369–380
Canham CD (1988) An index for understory light levels in and around Canopy Gaps. Ecology 69:1634–1638
Cheng S, Widden P, Messier C (2005) Light and tree size influence belowground development in yellow birch and sugar maple. Plant Soil 270:321–330
Close DC, Ruthrof KX, Turner S, Rokich DP, Dixon KW (2009) Ecophysiology of species with distinct leaf morphologies: effects of plastic and shadecloth tree guards. Restor Ecol 17:33–41
Cogliastro A, Paquette A (2012) Thinning effect on light regime and growth of underplanted red oak and black cherry in post-agricultural forests of south-eastern Canada. New For 43:941–954
Cogliastro A, Benjamin K, Bouchard A (2006) Effects of full and partial clearing, with and without herbicide, on weed cover, light availability, and establishment success of white ash in shrub communities of abandoned pastureland in southwestern Quebec, Canada. New For 32(2):197–210
Collins B, Battaglia LL (2008) Oak regeneration in southeastern bottomland hardwood forest. For Ecol Manage 255:3026–3034
Coutand C, Dupraz C, Jaouen G, Ploquin S, Adam B (2008) Mechanical stimuli regulate the allocation of biomass in trees: demonstration with young Prunus avium trees. Ann Bot 101(9):1421–1432
Cramer V, Hobbs R, Standish R (2008) What’s new about old fields? Land abandonment and ecosystem assembly. Trends Ecol Evol 23:104–112
D’Orangeville L, Bouchard A, Cogliastro A (2008) Post-agricultural forests: landscape patterns add to stand-scale factors in causing insufficient hardwood regeneration. For Ecol Manage 255:1637–1646
Devine WD, Harrington CA (2008) Influence of four tree shelter types on microclimate and seedling performance of Oregon white oak and western redcedar. Res. Pap. PNW-RP-576. Portland, OR: US Department of Agriculture, Forest Service, Pacific Northwest Research Station
Dey DC, Gardiner ES, Schweitzer CJ, Kabrick JM, Jacobs DF (2012) Underplanting to sustain future stocking of oak (Quercus) in temperate deciduous forests. New For 43:955–978
Drummond MA, Loveland TR (2010) Land-use pressure and a transition to forest-cover loss in the eastern United States. Bioscience 60:286–298
Dubois MR, Chappelka AH, Robbins E, Somers G, Baker K (2000) Tree shelters and weed control: effects on protection, survival and growth of cherrybark oak seedlings planted on a cutover site. New For 20:105–118
DuPlissis J, Yin X, Baughman MJ (2000) Effects of site preparation, seedling quality, and tree shelters on planted northern red oaks. Staff Paper Series No. 141. College of Natural Resources and Minnesota Agricultural Experiment Station University of Minnesota, St Paul
Environnement Canada (2010) Données des stations pour le calcul des normales climatiques au Canada de 1981–2010. Government of Canada. http://climat.meteo.gc.ca/climate_normals/results_1981_2010_f.html. Accessed April 2016
Forrester JA, Lorimer CG, Dyer JH, Gowe ST, Mladenoff DJ (2014) Response of tree regeneration to experimental gap creation and deer herbivory in north temperate forests. For Ecol Manage 329:137–147
Gasser D, Messier C, Beaudet M, Lechowicz MJ (2010) Sugar maple and yellow birch regeneration in response to canopy opening, liming and vegetation control in a temperate deciduous forest of Quebec. For Ecol Manage 259:2006–2014
Grandtner MM (1966) La végétation forestière du Québec méridional. Presses de l’Université Laval, Québec, p 216
Hamerlynck E, Knapp AK (1994) Stomatal responses to variable sunlight in bur oak (Quercus macrocarpa Michx.) leaves with different photosynthetic capacities. Int J Plant Sci 155:583–587
Hansen AJ, Spies TA, Swanson FJ, Ohmann JL (1991) Conserving biodiversity in managed forests. Bioscience 41:382–392
Huot M, Lebel F (2012) Plan de gestion du cerf de Virginie au Québec 2010–2017. Québec, Ministère des Ressources naturelles et de la Faune
Johnson PS, Shifley SR, Rogers R (2009) Ecology and silviculture of oaks -, 2nd edn. CABI North American Office, Cambridge
Kern CC, Reich PB, Montgomery RA, Strong TF (2012) Do deer and shrubs override canopy gap size effects on growth and survival of yellow birch, northern red oak, eastern white pine, and eastern hemlock seedlings? For Ecol Manage 267:134–143
Kuiters AT, Slim PA (2002) Regeneration of mixed deciduous forest in a Dutch forest-heathland, following a reduction of ungulate densities. Biol Conserv 105:65–74
Laliberté E, Bouchard A, Cogliastro A (2008) Optimizing hardwood reforestation in old fields: the effects of treeshelters and environmental factors on tree seedling growth and physiology. Restor Ecol 16:270–280
Langvall O, Löfvenius MO (2002) Effect of shelterwood density on nocturnal near-ground temperature, frost injury risk and budburst date of Norway spruce. For Ecol Manage 168:149–161
Lof M, Karlsson M, Sonesson K, Welander TN, Collet C (2007) Growth and mortality in underplanted tree seedlings in response to variations in canopy closure of Norway spruce stands. Forestry 80(4):371–383
Logan KT (1970). Adaptations of photosynthetic apparatus of sun- and shade- grown yellow birch (Betula alleghaniensis Britt.). Can J Bot 48(9):1681–1688
Lorenzetti F, Delagrange S, Bouffard D, Nolet P (2008) Establishment, survivorship, and growth of yellow birch seedlings after site preparation treatments in large gaps. For Ecol Manage 254:350–361
Mariotti B, Maltoni A, Jacobs DF, Tani A (2015) Tree shelters affect shoot and root system growth and structure in Quercus robur during regeneration establishment. Eur J Forest Res 134(4):641–652
Messier C, Nikinmaa E (2000) Effects of light availability and sapling size on the growth, biomass allocation, and crown morphology of understory sugar maple, yellow birch, and beech. Écoscience 7:345–356
Mitchell K (2007) Quantitative analysis by the point-centered quarter method. New York: Hobart and William Smith Colleges. arXiv:1010.3303v2 [q-bio.QM]
Niinemets Ü, Valladares F (2006) Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecol Monogr 76(4):521–547
Oosting HJ (1942) An ecological analysis of the plant communities of piedmont, North Carolina. Am Midl Nat 28:1–126
Oswalt CM, Clatterbuck WK, Houston AE (2006) Impacts of deer herbivory and visual grading on the early performance of high-quality oak planting stock in Tennessee, USA. For Ecol Manage 229(1–3):128–135
Paquette A, Bouchard A, Cogliastro A (2006a) Successful under-planting of red oak and black cherry in early-successional deciduous shelterwoods of North America. Annals Forest Sci 63(8):823–831
Paquette A, Bouchard A, Cogliastro A (2006b) Survival and growth of under-planted trees: a meta-analysis across four biomes. Ecol Appl 16:1575–1589
Paquette A, Bouchard A, Cogliastro A (2007) A less restrictive technique for the estimation of understory light under variable weather conditions. For Ecol Manage 242:800–804
Parent S, Messier C (1996) A simple and efficient method to estimate microsite light availability under a forest canopy. Can J For Res 26:151–154
Pinna S, Malenfant A, Côté M (2012) Vigor and growth responses of sugar maple and yellow birch seedlings according to different competing vegetation types and fabric shelter use. North J Appl For 29:133–140
Ponder F Jr (2003) Ten-year results of tree shelters on survival and growth of planted hardwoods. North J Appl For 20(3):104–108
Puértolas J, Oliet JA, Jacobs DF, Benito LF, Peñuelas JL (2010) Is light the key factor for success of tube shelters in forest restoration plantings under Mediterranean climates? For Ecol Manage 260:610–617
Ramankutty N, Foley JA (1999) Estimating historical changes in land cover: North American croplands from 1850 to 1992. Glob Ecol Biogeogr 8:381–396
Rebbeck J, Gottschalk K, Scherzer A (2011) Do chestnut, northern red, and white oak germinant seedlings respond similarly to light treatments? Growth and biomass. Can J For Res 41:2219–2230
Ricard JP, Messier C, Delagrange S, Beaudet M (2003) Do understory sapling respond to both light and below-ground competition?: a field experiment in a north-eastern American hardwood forest and a literature review. Ann For Sci 60:749–756
Sánchez-Pinillos M, Coll L, De Cáceres M, Ameztegui A (2016) Assessing the persistence capacity of communities facing natural disturbances on the basis of species response traits. Ecol Ind 66:76–85
Schmitt J, Wulff RD (1993) Light spectral quality, phytochrome and plant competition. Trend Ecol Evol 8(2):47–51
Song HT, Cheng S (2010) Various growth strategies of yellow birch seedlings in multiple-abiotic factors changing environments. Plant Soil Environ 56:235–243
Stanturf JA, Palik BJ, Dumroese RK (2014) Contemporary forest restoration: a review emphasizing function. For Ecol Manage 331:292–323
Taylor TS, Loewenstein EF, Chappelka AH (2006) Effect of animal browse protection and fertilizer application on the establishment of planted Nuttall oak seedlings. New For 32(2):133–143
Valverde T, Silvertown J (1997) Canopy closure rate and forest structure. Ecology 78:1555–1562
van Kuijk M, Anten NPR, Oomen RJ, Schieving F (2014) Stimulating seedling growth in early stages of secondary forest succession: a modeling approach to guide tree liberation. Front Plant Sci 5:345
Verheyen K, Bossuyt B, Hermy M, Tack G (1999) The land use history (1278–1990) of a mixed hardwood forest in western Belgium and its relationship with chemical soil characteristics. J Biogeogr 26(5):1115–1128
Walters RS (1993) Protecting red oak seedlings with tree shelters in northwestern Pennsylvania. Pennsylvania. Res. Pap. NE-679. Radnor, PA: US Department of Agriculture, Forest Service, Northeastern Forest Experiment Station
Ward JS, Stephens GR (1995) Protection of tree seedlings from deer browsing. In: Gottschalk KW, Fosbroke SLC (Eds.) 10th Central Hardwood Forest Conference. USDA For. Serv. Gen. Tech. Rep. NE-197, pp 507–514
Ward JS, Stephens GR (1997) Protecting red oak seedlings with tree shelters in northwestern Pennsylvania. Res. Pap. NE-679. Radnor, PA: US Department of Agriculture, Forest Service, Northeastern Forest Experiment Station
Ward JS, Gent MP, Stephens GR (2000) Effects of planting stock quality and browse protection-type on height growth of northern red oak and eastern white pine. For Ecol Manage 127(1):205–216
Wayne PM, Bazzaz FA (1993) Birch seedling responses to daily time courses of light in experimental forest gaps and shadehouses. Ecology 74:1500–1515
Yang G, Chen H, Cheng S, Yu H (2012) Interactive effects of canopy gap, liming and understory control on aboveground growth of yellow birch and sugar maple seedlings. Ekoloji 21:1–8
Acknowledgements
We thank the Ministère des Forêts, de la Faune et des Parcs (MFFP) of Québec and the Institut de Recherche en Biologie Végétale for financial support. We also thank Hafid Amrane and Paul Mazime Otye Moto from the local bureau of the MFFP for help on choosing the land for the experiment. Thanks also go to the many people that helped for the field work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Magnoux, A., Cogliastro, A. & Paquette, A. Growth of planted seedlings inside protective sleeves under strip openings of different widths in a post-agricultural forest. New Forests 49, 279–296 (2018). https://doi.org/10.1007/s11056-017-9619-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-017-9619-z