
Abstract
Over the last century, north-eastern North America has seen the gradual abandonment of much agricultural land that had become unsuitable for modern practices. This shift in land-use has released large areas that are now dominated by early-successional forests with limited mid- and late-successional species regeneration. Enrichment underplanting, with the partial preservation of the existing vegetation to serve as a shelterwood, has been proposed to restore the production of valuable timber. The present study reports mid-term (8–12 year) results from an enrichment planting experiment located at two sites in the St-Lawrence River Valley, eastern Canada. The objective was to identify the required frequency of low-intensity thinnings necessary to secure an optimal density of promising stems of black cherry (Prunus serotina Ehrh.) and red oak (Quercus rubra L.). Partial thinning treatments were applied during the 3rd and 7th growing seasons and compared to a single thinning applied at the 7th year. Each thinning treatment increased available light to 20–30 % in the understory (measured the following year at 2 m from ground level). The height of red oaks released twice was almost double that of red oaks thinned only once. The effect on black cherry was less dramatic and significant only at one of the two sites. A higher intensity or an earlier application of thinning regimes may be required for black cherry. The early thinning allowed a larger number of planted trees to react positively to the second thinning. The practice of double thinning over a period of 12 years allowed for the production of a significant number of competitively successful trees for the landowner.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Enrichment planting aims to restore a cohort of trees not only to replace aging overstorey trees (McCreary and Tecklin 2005), but also to improve forest composition, ecosystem functions and increase biodiversity (McComb et al. 1993; Hansen et al. 1991; Johnson et al. 2009), which can all contribute to increasing the value of otherwise degraded or undervalued forests, and thus increase the interest for their conservation in the landscape. An enrichment planting strategy using high-value native timber species may also be part of small-scale carbon offsetting, poverty alleviation, and conservation projects and opportunities (Paquette et al. 2009). We see enrichment planting as particularly promising in the specific context of post-agricultural forests.
Over the last century, the northeastern United States and eastern Canada have seen the gradual abandonment of land that has become unsuitable for modern agricultural practices (Bouchard and Domon 1997; Smith et al. 2004). This shift in land-use has released large areas that are now dominated by early-successional forests. Continued anthropogenic disturbance and isolation within an agricultural matrix may compromise the natural regeneration of these early-successional forests (D’Orangeville et al. 2008). These lands were originally forested, converted, and now represent an opportunity for restoration.
Enrichment underplanting, with the partial preservation of the existing vegetation to serve as a shelterwood, has been proposed (Martinez-Garza and Howe 2003; Gardiner et al. 2004; McCament and McCarthy 2005) to accelerate the return of tree species, as well as to restore valuable timber production in poor quality forests, such as those growing following the abandonment of agriculture. This silvicultural system aims to produce a shelterwood, an environment that is favourable to tree regeneration. Underplanting is an alternative to other practices such as planting following clear cut, which is less socially acceptable and which sometimes leads to failure (Schuler and Robison 2010).
The maintenance of a forest vegetation structure may also contribute to avoiding the successional setback and competition problems associated with more open stands and clearcuts (Carnevalea and Montagnini 2002; Lieffers and Stadt 1994; Truax et al. 2000; Balandier et al. 2006). The low value of the post-agricultural forests has often led to their conversion to single species conifer plantations, sometimes with negative environmental impacts (Borken et al. 2003; Makino et al. 2007; Aubin et al. 2008).
Among methods of artificial regeneration, enrichment planting is qualified as a more complex method. This is largely the consequence of unknown outcomes related to the competitive struggle between planted trees and competitors, in a relatively heterogeneous physical and biotic environment (Johnson et al. 2009). The shelterwood environment is a compromise between resource availability on the one hand, and protection from competition, herbivory, and climatic extremes on the other (Paquette et al. 2006b). Available light for tree regeneration establishment is increased through thinning (Drever and Lertzman 2003), but not so much as to promote competition (Carnevalea and Montagnini 2002; Lieffers and Titus 1989), or climatic stress (Agestam et al. 2003; Langvall and Lofvenius 2002; Man and Lieffers 1999). For enrichment planting applied in young successional forests, we proposed that a shelterwood can be thinned in several steps that are so gradual, that the regenerated stand developing beneath it eventually grows up and becomes dominant without major forest structure transformation. This procedure is similar to the irregular shelterwood described by Johnson et al. (2009). Thinning and clearing should be applied to release the planted trees from competition. Understanding the interplay between thinning frequency and enrichment underplanting regeneration response can result in a possible standardization of the method, to create successful conditions of survival and growth in post-agricultural forests. On the other hand, many treatments are perceived to be too labour intensive and costly (Groninger and Long 2008). Enrichment underplanting efforts may suffer from insufficient interventions where they are critical, mostly on mesic sites, where planted trees must compete with opportunistic species of these aggrading successional forests. Treatments have to show very high rates of success for these significant investments (Morrissey et al. 2010).
The present study reports mid-term (8–12 year) results from a long-term enrichment underplanting experiment of common but poorly understood young secondary succession low quality stands within private rural landscapes. A first paper based on this experimental design (Paquette et al. 2006a) had provided an analysis of the effect of a release treatment at year 3 on the growth of red oak (Quercus rubra L.) and black cherry (Prunus serotina Ehrh.) after 6 years of growth under shelterwood. This study uses the same experimental design to analyse the effects of two thinnings, at years 3 and 7, in comparison with a single thinning at year 7. There is a strong incentive for the latter option in order to reduce costs among regional and national forest management agencies. This paper reports on 12 years of growth for both tree species. Available light, tree height and released stem densities after 12 years are analyzed with respect to treatments.
Materials and methods
Study sites
Two sites located in the St-Lawrence River Valley region of eastern Canada were selected for this study. The region has a humid continental climate, with monthly means of 21 °C in July and −10 °C in January. Mean annual total precipitation is 1,030 mm (of which 18 % falls as snow), and is well distributed throughout the year (Environment Canada, climatological normals 1971–2000). Stands at both sites share similar till deposits and sandy loam soil textures (pH 5.2–5.8) with a potential terminal stage of sugar maple-bitternut hickory communities (Thibault 1985). Both sites originate from recent (ca 1962) agricultural abandonment (pasture), which followed extensive deforestation and high grading.
The St-Chrysostome site (45° 09′ N; 73° 45′ W) is dominated by grey birch (Betula populifolia Marsh.). Hophornbeam (Ostrya virginiana (Mill.) K. Koch) and white elm (Ulmus americana L.) are also present, while late successional species are almost absent. Tall shrubs (Crataegus spp. and Malus pumila Mill.) are also present and form dense thickets. This 2.5 ha site was deemed to be sufficiently open to be under-planted directly in the spring of 1998, without prior site preparation.
The Ste-Clotilde site (45° 08′ N; 73° 38′ W) is 3 ha and also mostly composed of grey birch, but less so than on the St-Chrysostome site because this species was the primary target of a preparation thinning. It is accompanied by hophornbeam, white ash (Fraxinus americana L.), hornbeam (Carpinus caroliniana Walt.) and white elm. Shrubs are less abundant. The Ste-Clotilde site was prepared during fall 1997 by thinning approximately 30 % of the stand’s basal area without creating major openings in the canopy, and primarily targeting early successional species. The residual basal area was 16 m2/ha, which was lower than that of the St-Chrysostome site without preparation (21 m2/ha), with comparable stem densities of over 6000 stems/ha and average stand height of 11 m.
Under-planting of black cherry and red oak seedlings
Red oak and black cherry were chosen to restore the potential for high value timber production. They are associated species of the climax forest of the region. They have respectively an intermediate (Sander 1990) and low tolerance (Marquis 1990) to shade. Although there is a large body of literature on the regeneration of red oak in North America, it deals mostly with poorly regenerated mature oak stands in the absence of forest fire. Very little data is available concerning black cherry in shelterwood environments.
Black cherry and red oak under-planting was carried out in spring 1998 (hereafter year 1) using one-year-old black cherry and red oak containerized seedlings (340 ml) over the entire area of the two sites. Mean height and diameter at the root collar for black cherry were 34 cm and 4 mm, respectively, and 27 cm and 6 mm for red oak. Black cherries were planted in greater numbers than red oaks at both sites (2:1 at St-Chrysostome and 6:1 at Ste-Clotilde) due to limited supplies of red oaks at the nursery. Trees were planted every 3 meters, on parallel planting lines also spaced at 3 meters. Species were distributed on alternate planting lines, according to their respective proportions. Inadequate micro-sites were avoided, due in particular to small depressions with drainage problems, producing an approximate final density of 900 trees per hectare.
Experimental design and treatments
This study began in 2000, 2 years after the trees had been planted. No other trials of this kind (hardwoods planted under a shelterwood of intolerant, pioneer species) were available at the time. This was a great opportunity for evaluative research, as the method was being proposed and applied by progressive foresters and producers. The research questions investigated in this study were the same as those of the foresters and policy makers involved in the project.
On each site we delineated two replication blocks, to account for possible spatial heterogeneity (Hulbert 1984). Inside these blocks, in early spring of 2000 (year 3), experimental rectangular plots were delineated so as to contain areas which were completely planted without interruption and with visually homogenous vegetation composition. They were scattered over the planted areas of each block, and buffers of at least 20 m were kept between them. Plots may contain underplanted red oak and black cherry seedlings, or one of the two: block area, plot number and seedling number per species are presented in Table 1. Plots were randomly assigned to either “single thinning (year 7)” or “two thinning (years 3 and 7)” treatment. The complete randomized block design for this study is formed by the four replication blocks (two on each site) and the two treatments. Red oak and black cherry were analysed separately. Herbivory by white-tailed deer still occurred, but was reduced by the application of a deer repellent (Deer-Away, IntAgra Inc., Minneapolis, MN, USA), twice annually (May and October), from year 3 to 6.
During the third year, on half of the plots, all trees and shrubs (1 cm < DBH < 10 cm) were cut within a 1-m radius around the planted trees, whereas the herbaceous vegetation and small woody stems (<1 cm) were cut flush with the ground within a smaller radius (50 cm), in proportion to their smaller size. Larger trees (>10 cm), within the same one-meter radius, were devitalized with glyphosate herbicide capsules (E-Z-Ject system, Waynesboro, MS, USA) and left standing. The remaining plots were left untouched.
At the end of the 7th year on all plots a thinning from above, also centered on the planted trees and removing approximately 30 % of the total cover (visually estimated), was carried out without creating large openings. The experimental design compares the effects of two post-plantation thinning, at years 3 and 7, to a single thinning at year 7.
Light availability
Available light was measured at 2 m from the ground for years 3, 4, 5, 6, 8, 10, and 12 on, respectively, 295, 101, 584, 589, 469, 574 and 592 trees. Light measurements were used as they integrate the competitive effects of all plants around the seedlings and are thus accurate descriptors of a seedling’s growing environment (Paquette et al. 2007a). They also respond to the thinning treatment, accounting for prior heterogeneity and possible variability in its application, including the presence of devitalized trees that were left standing.
From years 3 to 6, the instantaneous measurement of seasonal available light (% PPFD) was made according to the “Overcast” method of Parent and Messier (1996), using two quantum sensors on a complete overcast day. One sensor was used for in situ measurements below tree canopy while the second was placed in a nearby open field for reference. Light measurements were taken at each seedling location. Because required light measurement conditions (overcast sky) were difficult to obtain within a short period of time for such a large number of measurements, we limited the first measurements to only those plots which were to receive the release treatment in order to obtain pre-treatment values (year 3). At year 4, especially difficult for light measurements, we measured only a sample of trees. Measurements were taken for all trees at years 5 and 6.
During the following years (7, 8, 10 and 12), available light (% PPFD) was measured with an alternative method free of weather constraint we had previously developed that make use of the BF2 type Sunshine Sensor (Delta-T, Burwell, Cambridge, UK). The technique match both the well-established “Overcast” method and the accepted reference of hemispherical image analysis (Paquette et al. 2007a). The sensor comprises seven quantum sensors under a hemispherical dome with a pattern of clear and opaque areas (mask) such that at any given moment, in whatever direction the BF2 is oriented, at least one of the sensors will be in diffuse light (Wood et al. 2003). Only the diffuse signal was used and the ratio of available light was obtained in the same fashion as with the “Overcast” method, namely by placing another BF2 in a nearby open area and communicating by radio.
Survival, growth and height class density
Initial planted seedling density was estimated to ca 900/ha from the contractors’ report. At the beginning of the third season (year 2000) we measured tree plot density based on the surface of each plot measured by GPS and the number of underplanted trees present. These third year values by plot were used to estimate the fraction of living trees after 12 years. Mortality during the initial 2 years was excluded to compensate for factors which cannot be controlled for, such as occasional poor planting technique, poor quality planting stock, or predation. Among these third year living trees, damaged or diseased trees, as well as a small number of trees with obvious drainage problems at the microsite level (not identified at the time of plantation), were excluded from height growth analysis. The number of trees used for each year growth analysis is shown in Table 3. Height of planted trees was measured at the end of growing seasons of years 3 through 8, and of years 10 and 12.
From the area of experimental plots and the number of planted trees still alive at year 12 (see Table 1), the density per hectare of trees in each of four height classes (A < 2 m; 2 m ≤ B < 3 m; 3 m ≤ C < 4 m; D ≥ 4 m) was determined for each species.
Statistical analysis
The experimental design is composed of four replication blocks (two on each site), two treatments (single thinning and two thinning) and two species. Sites are thus considered random effects (Hooper et al. 2002) and eventual differences between sites, or among them, can be investigated using blocks effects. ANOVA with random effects was used to test the effect of the release treatments on available light and tree height for each species. Rank transformations were used for light data to meet assumptions of normality and homoscedasticity in parametric analyses (Iman and Conover 1979). Analyses were carried out on SAS software (SAS Institute Inc. 2009).
Results
The effects of the first thinning at year three are reported in details in Paquette et al. (2006a) which showed that the growth of underplanted trees responded mainly to the density of the upper canopy layer. The release treatment at year three was successful in significantly increasing available light for the next 3 years compared with the control (no early thinning). Planted trees, particularly red oak, responded well to this first thinning treatment. Also, deer herbivory was not increased by the thinning.
Light
The 30 % thinning of the canopy, done at the end of the 7th year of growth on the total area of both study sites, has produced an increase in available light measured at 2 m above ground (mean values ranging from 16 to 33 %) the year following thinning (year 8) (Table 2; Fig. 1). Overall, this thinning has produced more available light at the Ste-Clotilde site, with a mean increase of 14 percentage points for plots thinned twice (years 3 and 7) and of 20 percentage points in the case of the single thinning (year 7) (Fig. 1). The increase at the St-Chrysostome site was respectively of 10 and 12 percentage points of increased light for the double and the single thinning (Fig. 1). At this site, the lower light availability at 2 m can be explained by the presence of more abundant understory vegetation than at the Ste-Clotilde site.

Available light (% PPFD) measured at 2 m from ground level at St-Chrysostome (triangle) and Ste-Clotilde (square). Open symbols refer to the double thinning treatment (years 3 and 7) whereas closed symbols are used for the single thinning treatment (year 7). Data for year 7 at St-Chrysostome were not available; an estimation based on regression on the previous two measurements is presented using dotted lines
This increase in available light is rapidly followed by a gradual reduction at the two study sites (Fig. 1). In 4 years (from year 8 to 12), the mean light level (at 2 m height) reaches a comparable level to that measured before the thinning at year 7, and this for both treatments. Following the last thinning (year 7 cleaning), the difference between the double thinning and the single thinning was significant only at the year immediately following the treatment (Table 2).
Height growth
The mean height growth of red oak was increased by the double thinning, as the results show 3 and 5 years after the last thinning (years 10 and 12) (see treatment effect, Table 3; Fig. 2). The mean height of black cherry was less affected by the treatments. Only in the 3rd year after the last thinning (year 10) did black cherry have a mean height that was significantly higher in the double thinning plots (Table 3; Fig. 2). Five years after the last thinning (year 12), the difference between treatments for black cherry disappeared, whereas a Block × Treatment interaction was significant (Table 3). This indicates that the growth of black cherry was superior following the double thinning than to the single thinning in the blocks of only one of the study sites, the St-Chrysostome site (Table 3; Fig. 2). Unexpectedly, the site with the highest amount of available light at 2 m (Ste-Clotilde) produced the slowest growth for black cherry.

Height growth of red oak and black cherry underplanted at St-Chrysostome (triangle) and Ste-Clotilde (square).Open symbols refer to the double thinning treatment (years 3 and 7) whereas closed symbols are used for the single thinning treatment (year 7)
From year 8 to 12, and in plots with a single thinning, the mean annual height growth was 30 cm/year for red oak at both sites, and between 15 and 20 cm/year for black cherry (Ste-Clotilde and St-Chrysostome sites respectively) (Fig. 2). The double thinning at the St-Chrysostome site produced a mean annual height growth (from year 8 to 12) of 75 cm/year for red oak and of 38 cm/year for black cherry (Fig. 2). Mean annual height growth was respectively 65 and 15 cm/year for red oak and black cherry at the Ste-Clotilde site (Fig. 2). Thus, red oak obtained a mean height growth in the double thinning that was nearly double of that obtained in the single thinning and this for both sites. We observe a height decrease of red oak immediately following the thinning (Fig. 2) that is associated with the breakage of some crowns, caused by falling trees during the operation. However, the subsequent height growth remained strong.
Overall survival and height class density
With an initial density in the 3rd growing season of nearly 835 stems/ha on average for both tree species combined, on both sites, the survival at the St-Chrysostome site at the 12th growing season is 98 and 96 %, respectively, for black cherry and red oak that received the double thinning or the single thinning. The survival rate is lower for black cherry at the Ste-Clotilde site, with 87 and 89 % respectively for the double thinning and for the single thinning, in comparison with 93 and 97 % for red oak. Treatment type does not seem to have a major effect on the survival of planted trees after 12 years.
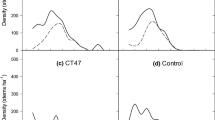
The densities shown in Fig. 3 were obtained following the assumption that red oak or black cherry trees would have been planted alone. Saplings of 3 m and more in height (classes C and D) are the most likely to represent future tree stems. At the St-Chrysostome site, the final density of stem size classes C and D of red oak or black cherry measured at the 12th year of growth was respectively 448 and 433 stems/ha for the double thinning and 129 and 223 stems/ha for the single thinning (Fig. 3). Density of stems in size classes C and D at the Ste-Clotilde site were 644 and 174 red oak and black cherry per ha respectively for the double thinning, and 106 and 134 stems/ha for the single thinning (Fig. 3). Comparatively, 61 % of the planted trees at the St-Chrysostome site have a height greater than 3 m in the double thinning and 25 % are taller than 3 m in the single thinning, against respectively 38 and 15 % of planted trees at the Ste-Clotilde site. For this site, we observe that black cherry trees of 4 m and more are particularly few (Fig. 3).

Underplanted tree densities of red oak and black cherry on each site at the end of the 12th year of growth by treatment (single or double thinning) and height class (A < 2 m; 2 m ≤ B < 3 m; 3 m ≤ C < 4 m; D ≥ 4 m)
Discussion
Canopy closure dynamics
During the first phase of this experiment (growing season 4–6) reported by Paquette et al. (2006a), we also observed a rapid canopy closure in the years following thinning. In this study, the measurements reveal that canopy closure was just as rapid following the second thinning done in the 7th year, in plots having been thinned only once as well as in those having been thing twice. The rate of light regime modification in hardwood forests was found to be more rapid in the first few years following selection cutting (Valverde and Silvertown 1997; Beaudet and Messier 2002). Following light to moderate thinning, a rapid height growth of the planted trees is necessary for them to attain a position that provides favourable light availability and protection from deer browsing. The rapid closure of the canopy and the objective of obtaining an optimal value of available light between 25 and 50 % [see review by Paquette et al. (2006b)] lead to the planning of more than one thinning treatment in order to maintain periods during which available light is optimal (Motsinger et al. 2010). Promoting regeneration development requires active management to reverse the trend of increasing stand density and limiting light in the understory (Dey et al. 2012).
Favour fast initial height growth
The use of only one late thinning (year 7) has produced trees that are limited in height (around 2 m high) after 12 years. This growth is below the minimum levels of 30 cm/year suggested by Johnson (1984) for ensuring the establishment of red oak in shelterwoods. At this height, a large number of planted trees are still likely to be browsed by deer, which reduces growth but also the probability of obtaining a straight bole. Survival rate has, however, remained very good.
The use of two thinning (years 3 and 7) has produced a very good height growth of red oak in spite of the moderate and sub-optimal (Paquette et al. 2006b) light levels reached following the second thinning at the St-Chrysostome site. Morrissey et al. (2010) have presented 5 year results were the larger gap sizes, ranging from 0.4 to 0.024 ha, improved red oak seedling height and diameter. However, they also underlined the greater numbers of larger competitors in larger gap sizes and the necessity of silvicultural cleaning treatments or crop tree release within group openings as crown closure begins. The method used in this study presents an alternative to creating large gaps, proceeding instead by repeated moderate and diffuse release treatments. A favourable release treatment effect was also shown by Povak et al. (2008), with their proposition to control tall understory saplings for successful red oak regeneration.
Back cherry reacted less strongly to the thinning than did red oak. According to Niinemets and Valladares (2006), Humbert et al. (2007), and Baker (1951) black cherry is slightly less tolerant to shade than red oak. We had proposed in 2006 that light availability induced by the first thinning was not sufficiently high for black cherry (Paquette et al. 2006a). The addition of a second thinning has produced growth results for black cherry that are clearly improved at one of the study sites (St-Chrysostome). Perhaps a more vigorous first thinning would have produced better growth for black cherry. But understory competition may have then become more important. We had, however, measured a favourable effect of understory density on the growth of black cherry, pointing to an increase in available light at the top of the seedlings (which would also explain the thicker understory), and possibly better protection from deer browsing (Paquette et al. 2006a). We add to that a possible benefit from lateral occupation by small shrubs and trees, as mentioned by Cogliastro et al. (2006) for white ash in a shrub community. The literature confirms that a partial cover is required for the optimal establishment of black cherry, but that full sunlight is necessary afterwards to ensure continued growth, which can be vigorous (Marquis 1990; Verheyen et al. 2007). Seedlings and saplings of P. serotina, introduced in Europe, have been shown to need a considerable amount of light, i.e., 58–80 and 21–47 % of full light, respectively (Godefroid et al. 2005). We had noted earlier the lack of response to thinning of black cherry and noted that “regeneration of this species would be successful under a more extensive and earlier opening of the canopy, thus taking advantage of reserves accumulated during early establishment under a shelterwood”. (Paquette et al. 2007b). Indeed, regeneration of this species is said to be favoured under more intensive disturbances (Marquis 1990).
The absence of an effect of the thinning on the growth of black cherry at one of the two sites (Ste-Clotilde), the one with the highest available light, as well as the lowest survival rate, could be the result of a microtopography of small mounds and depressions, with a greater amount of micro-sites with slower drainage. The greater number of black cherries planted at this site, in comparison with red oaks, may have over-represented them in locations with poor drainage conditions at this site. Black cherry shows rapid loss in productivity with increasingly wetter conditions (Marquis 1990), also observed in Belgium were the species is restricted to the driest areas (Godefroid et al. 2005).
Producing a significant number of free growing trees
For the silviculture of European oaks (Q. robur L. and Q. petraea Matt.), the final objective of 70 stems/ha in stands or in plantations has been proposed (Lemaire 2010). This target aims at rapidly producing large trees seems to be appropriate for the relatively small areas of private properties in Eastern Canada and in the north-eastern United States that could undergo enrichment planting. The double thinning used in this study allows for the possibility of concentrating the application of future silvicultural treatments (pruning, clearing and selection) on the stems that were more than 3 m in height (classes C and D) by the 12th growing season. The quantity of trees offers some insurance to the owner for the potential negative impact of eventual disturbances, and allows for the pre-selection of high quality future trees.
This study adds valuable information to research on enrichment planting done in different biomes, northern (Man and Lieffers 1999), temperate (Dey and Parker 1997; Gordon et al. 1995), continental (Maas-Hebner et al. 2005) and tropical (Montagnini et al. 1997). More specifically, this artificial hardwood regeneration experiment, done in post-agricultural forests, has contributed to better understand how species interact in different physical environments and are able to maintain dominance in changing combinations of competing vegetation (Dey et al. 2009).
For young and often species-poor stands that have developed following agricultural abandonment, a successful enrichment underplanting allows the forest structure to be conserved, the diversification of these stands by the establishment of mid-successional species, and the increase of their production potential. These results may increase the interest for the conservation of such stands, and ensure the maintenance of their ecological benefits to the landscape.
Conclusion
A first early thinning allowed a larger number of planted trees to achieve good growth and to be able to react adequately to the second thinning. A more vigorous thinning could be tested for black cherry. The practice of double thinning over a period of 12 years has allowed for the production of a significant number of competitively successful trees for the landowner. At their current size, these trees are less prone to deer browsing, can receive more light and generally have a single straight bole. This success has also allowed for the enrichment of the dominant species of the stand. We believe that a further cleaning intervention, as defined by Helms (1998), applied at the sapling stage and designed to free the favoured trees from those that overtop them, should be made at around year 15.
The present trend to thin late after enrichment underplanting remains strong among forest owners and managers, because of the costs of planting and the absence of revenues associated with the thinning of a young stand of intolerant hardwoods. The results presented here confirm the advantage of thinning early (year 3). A single late thinning (year 7) was not sufficient to compensate for the gains provided by the early thinning. The selection of sites with rich soils that are well-drained would maximize the yield on investments. In such prime conditions, where the closing of the canopy and the development of competitors can be rapid, thinning would be even more necessary. Artificial hardwood regeneration success can be accomplished by underplanting high quality seedlings, controlling competing vegetation and herbivores (see review in this issue by Dey et al. 2012). We believe these investments would still compare favourably with those required with the afforestation of recently abandoned agricultural fields with hardwoods.
References
Agestam E, Ekö PM, Nilsson U, Welander NT (2003) The effects of shelterwood density and site preparation on natural regeneration of Fagus sylvatica in southern Sweden. For Ecol Manag 176:61–73
Aubin I, Messier C, Bouchard A (2008) Can plantations develop understory biological and physical attributes of naturally regenerated forests? Biol Conserv 141:2461–2476
Baker FS (1951) A revised tolerance table. J For 47:179–181
Balandier P, Collet C, Miller JH, Reynolds PE, Zedaker SM (2006) Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 79(1):3–27
Beaudet M, Messier C (2002) Variation in canopy openness and light transmission following selection cutting in northern hardwood stands: an assessment based on hemispherical photographs. Agric For Meteorol 110:217–228
Borken W, Xu YJ, Beese F (2003) Conversion of hardwood forests to spruce and pine plantations strongly reduced soil methane sink in Germany. Glob Change Biol 9:956–966
Bouchard A, Domon G (1997) The transformation of the natural landscapes of the Haut-Saint-Laurent (Québec) and its implication on future resources management. Landsc Urban Plan 37:99–107
Carnevalea NJ, Montagnini F (2002) Facilitating regeneration of secondary forests with the use of mixed and pure plantations of indigenous tree species. For Ecol Manag 163(1–3):217–227
Cogliastro A, Benjamin K, Bouchard A (2006) Effects of full and partial clearing, with and without herbicide, on weed cover, light availability, and establishment success of white ash in shrub communities of abandoned pastureland in southwestern Quebec, Canada. New For 32:197–210
Dey DC, Parker WC (1997) Morphological indicators of stock quality and field performance of red oak (Quercus rubra L.) seedlings underplanted in a central Ontario shelterwood. New For 14:145–156
Dey DC, Gardiner ES, Schweitzer CJ, Kabrick JM, Jacobs DF (2012) A synthesis of underplanting oaks to sustain future oak stocking in temperate deciduous forests. New For 43(5/6)
Dey DC, Spetich MA, Weigel DR, Johnson PS, Graney DL, Kabrick JM (2009) A suggested approach for design of oak (Quercus L.) regeneration research considering regional differences. New For 37:123–135
D’Orangeville L, Bouchard A, Cogliastro A (2008) Post-agricultural forests: landscape patterns add to stand-scale factors in causing insufficient hardwood regeneration. For Ecol Manag 255:1637–1646
Drever CR, Lertzman KP (2003) Effects of a wide gradient of retained tree structure on understory light in coastal Douglas-fir forests. Can J For Res 33:137–146
Gardiner ES, Stanturf JA, Schweitzer CJ (2004) An afforestation system for restoring bottomland hardwood forests: biomass accumulation of Nuttall oak seedlings interplanted beneath eastern cottonwood. Restor Ecol 12:525–532
Godefroid S, Phartyal SS, Weyembergh G, Koedam N (2005) Ecological factors controlling the abundance of non-native invasive black cherry (Prunus serotina) in deciduous forest understory in Belgium. For Ecol Manag 210:91–105
Gordon AM, Simpson JA, Williams PA (1995) Six-year response of red oak seedlings planted under a shelterwood in central Ontario. Can J For Res 25:603–613
Groninger J, Long M (2008) Oak ecosystem management considerations for central hardwoods stands arising from silvicultural clearcutting. North J Appl For 25:173–179
Hansen AJ, Spies TA, Swanson FJ, Ohmann JL (1991) Conserving biodiversity in managed forests. Bioscience 41:382–392
Helms JA (1998) Dictionary of forestry. Society of American Foresters/CABI Publishing, Bethesda
Hooper E, Condit R, Legendre P (2002) Responses of 20 native tree species to reforestation strategies for abandoned farmland in Panama. Ecol Appl 12:1626–1641
Hulbert SH (1984) Pseudoreplication and the design of ecological field experiments. Ecol Monogr 54:187–211
Humbert L, Gagnon D, Kneeshaw D, Messier C (2007) A shade tolerance index for common understory species of northeastern North America. Ecol Ind 7(1):195–207
Iman RL, Conover WJ (1979) The use of the rank transform in regression. Technometrics 21(4):499–509
Johnson PS (1984) Responses of planted northern red oak to three overstory treatments. Can J For Res 14:536–542
Johnson PS, Shifley SR, Rogers R (2009) Ecology and silviculture of oaks, 2nd edn. CABI North American Office, Cambridge
Langvall O, Lofvenius MO (2002) Effect of shelterwood density on nocturnal near-ground temperature, frost injury risk and budburst date of Norway spruce. For Ecol Manag 168:149–161
Lemaire J (2010) Le chêne autrement: produire du chêne de qualité en moins de 100 ans en futaie régulière. Institut pour le développement forestier, Paris, 176 p
Lieffers VJ, Stadt KJ (1994) Growth of understorey Picea glauca, Calamagrostis canadensis, and Epilobium angustifolium in relation to overstory light. Can J For Res 24:1193–1198
Lieffers VJ, Titus SJ (1989) The effects of stem density and nutrient status on size inequality and resource allocation in lodgepole pine and white spruce seedlings. Can J Bot 67:2900–2903
Maas-Hebner KG, Emmingham WH, Larson DJ, Chan SS (2005) Establishment and growth of native hardwood and conifer seedlings underplanted in thinned Douglas-fir stands. For Ecol Manag 208:331–345
Makino S, Goto H, Hasegawa M, Okabe K, Tanaka H, Inoue T, Okochi I (2007) Degradation of longicorn beetle (Coleoptera, Cerambycidae, Disteniidae) fauna caused by conversion from broad-leaved to man-made conifer stands of Cryptomeria japonica (Taxodiaceae) in central Japan. Ecol Res 22:372–381. doi:10.1007/s11284-007-0359-y
Man R, Lieffers V (1999) Effects of shelterwood and site preparation on microclimate and establishment of white spruce seedlings in a boreal mixedwood forest. For Chron 75(5):837–844
Marquis DA (1990) Black cherry (Prunus serotina Ehrh.). In: Burns RM, Honkala BH (eds) Silvics of North America: Vol 2 Hardwoods. USDA Forest Service, Washington, pp 594–604
Martinez-Garza C, Howe HF (2003) Restoring tropical diversity: beating the time tax on species loss. J Appl Ecol 40:423–429
McCament CL, McCarthy BC (2005) Two-year response of American chestnut (Castanea dentata) seedlings to shelterwood harvesting and fire in a mixed-oak forest ecosystem. Can J For Res 35:740–749
McComb W, Spies T, Emmingham W (1993) Douglas-fir forests: managing for timber and wildlife habitats. J For 91:31–42
McCreary DD, Tecklin J (2005) Restoring native California oaks on grazed rangelands. USDA Gen Tech Rep, RMRS-P-35, pp 109–112
Montagnini F, Eibl B, Grance L, Maiocco D, Nozzi D (1997) Enrichment planting in overexploited subtropical forests of the Paranaense region of Misiones, Argentina. For Ecol Manag 99:237–246
Morrissey RC, Jacobs DF, Davis AS, Rathfon RA (2010) Survival and competitiveness of Quercus rubra regeneration associated with planting stocktype and harvest opening intensity. New For 40:273–287. doi:10.1007/s11056-010-9199-7
Motsinger J, Kabrick J, Dey D, Henderson D, Zenner E (2010) Effect of midstory and understory removal on the establishment and development of natural and artificial pin oak advance reproduction in bottomland forests. New For 39(2):195–213. doi:10.1007/s11056-009-9164-5
Niinemets Ü, Valladares F (2006) Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol Monogr 76(4):521–547
Paquette A, Bouchard A, Cogliastro A (2006a) Successful under-planting of red oak and black cherry in early-successional deciduous shelterwoods of North America. Ann For Sci 63(8):823–831
Paquette A, Bouchard A, Cogliastro A (2006b) Survival and growth of under-planted trees; a meta-analysis across four biomes. Ecol Appl 16(4):1575–1589
Paquette A, Bouchard A, Cogliastro A (2007a) A less restrictive technique for the estimation of understory light under variable weather conditions. For Ecol Manag 242(2–3):800–804. doi:10.1016/j.foreco.2007.01.067
Paquette A, Bouchard A, Cogliastro A (2007b) Morphological plasticity in seedlings of three deciduous species under shelterwood under-planting management does not correspond to shade tolerance ranks. For Ecol Manag 241(1–3):278–287. doi:10.1016/j.foreco.2007.01.004
Paquette A, Hawryshyn J, Senikas AV, Potvin C (2009) Enrichment planting in secondary forests: a promising clean development mechanism to increase terrestrial carbon sinks. Ecol Soc 14(1):31 [online]. http://www.ecologyandsociety.org/vol14/iss31/art31/
Parent S, Messier C (1996) A simple and efficient method to estimate microsite light availability under a forest canopy. Can J For Res 26:151–154
Povak NA, Lorimer CG, Guries RP (2008) Altering successional trends in oak forests: 19 year experimental results of low- and moderate-intensity silvicultural treatments. Can J For Res 38:2880–2895
Sander IL (1990) Quercus rubra L. In: Silvics of North America: vol 2, Hardwoods. Agriculture Handbook 654. Forest Service United State Department of Agriculture, Washington, pp 727–733
SAS Institute Inc (2009) SAS/STAT® 9.2 user’s guide: mixed modelling (book excerpt). SAS Institute Inc, Cary
Schuler JL, Robison DJ (2010) Performance of northern red oak enrichment plantings in naturally regenerating Southern Appalachian hardwood stands. New For 40:119–130
Smith WB, Miles PD, Visage JS, Pugh SS (2004) Forest resources of the United States, 2002. USDA Gen. Tech. Report NC-241. St. Paul, p 146
Thibault M (1985) Les régions écologiques du Québec méridional, (1:1 250 000). Service de la cartographie, Ministère de l’Énergie et des Ressources du Québec, Québec
Truax B, Lambert F, Gagnon D (2000) Herbicide-free plantations of oaks and ashes along a gradient of open forested mesic environments. For Ecol Manag 137:155–169
Valverde T, Silvertown J (1997) Canopy closure rate and forest structure. Ecology 78(5):1555–1562
Verheyen K, Vanhellemont M, Stock T, Hermy M (2007) Predicting patterns of invasion by black cherry (Prunus serotina Ehrh.) in Flanders (Belgium) and its impact on the forest understorey community. Divers Distrib 13:487–497
Wood J, Muneer T, Kubie J (2003) Evaluation of a newphotodiode sensor for measuring global and diffuse irradiance, and sunshine duration. J Sol Energy T Asme 125:43–48
Acknowledgments
We wish to thank the owners of the sites for their invaluable support over the years. We also gratefully acknowledge the work of technical staff (Stéphane Daigle, Léa Bouttier, André-Jean Lalanne, Valentin Arnaud, Réné Dulude) and two anonymous referees for providing constructive reviews. This work was supported by the Programme de Mise en Valeur des Ressources du Milieu Forestier—Volet II, Montérégie. We would also like to dedicate this work to the memory of our friend and long-time colleague André Bouchard who died during the course of this experiment.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cogliastro, A., Paquette, A. Thinning effect on light regime and growth of underplanted red oak and black cherry in post-agricultural forests of south-eastern Canada. New Forests 43, 941–954 (2012). https://doi.org/10.1007/s11056-012-9329-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-012-9329-5