
Abstract
Backgrounds
Epithelial mesenchymal transition (EMT) is a critical process involved in the invasion and metastasis of cancer, including lung cancer (LC). Transforming growth factor (TGF)-β is one of factors capable of inducing EMT. Polyinosinic-polycytidylic acid (polyI:C), a synthetic agonist for toll-like receptor (TLR) 3, can enhance immune responses and has been used as an adjuvant for cancer vaccines; however, it remains unclear whether it influences other process, such as EMT. In the present study, we examined the effects of polyI:C on TGF-β-treated A549 human LC cells.
Methods and results
By in vitro cell proliferation assay, polyI:C showed no effect on the growth of A549 cells treated with TGF-β1 at the concentration range up to 10 μg/ml; however, it markedly suppressed the motility in a cell scratch and a cell invasion assay. By Western blotting, polyI:C dramatically decreased TGF-β1-induced Ak strain transforming (Akt) phosphorylation and increased phosphatase and tensin homologue (PTEN) expression without affecting the Son of mothers against decapentaplegic (Smad) 3 phosphorylation or the expression level of E-cadherin, N-cadherin or Snail, indicating that polyI:C suppressed cell motility independently of the ‘cadherin switching’. The Akt inhibitor perifosine inhibited TGF-β1-induced cell invasion, and the PTEN-specific inhibitor VO-OHpic appeared to reverse the inhibitory effect of polyI:C.
Conclusion
PolyI:C has a novel function to suppress the motility of LC cells undergoing EMT by targeting the phosphatidylinositol 3-kinase/Akt pathway partly via PTEN and may prevent or reduce the metastasis of LC cells.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Lung cancer (LC) arises from respiratory epithelial cells and are divided into two groups; small cell lung cancer (SCLC) and non–small cell lung cancer (NSCLC) [1, 2]. LC is one of the most common cancer worldwide and the survival rate of LC patients is low among all cancer patients [1, 3]. LC cells metastasize to remote tissues, such as brain, bone, lymph nodes, and liver, and the presence of LC metastasis is associated with poor prognosis [4]. Significant progress has been made in both diagnosis and treatment of LC; however, the exact mechanisms of LC metastasis remain unclear. Additional studies aiming to better understand the mechanisms of LC metastasis and to develop new means to prevent or slow the metastasis are needed.
During cancer metastasis, cancer cells first dissociate from the primary tumor and invade the surrounding tissue and move into the circulation. Epithelial-mesenchymal transition (EMT) is a well-studied cellular process thought to play a critical role in cancer cell invasion and metastasis. EMT is characterized by the downregulation of epithelial genes, such as E-cadherin, and the upregulation of mesenchymal genes, such as neural (N)-cadherin and a zinc-finger transcriptional repressor snail [5]. One of the major molecules that regulate EMT is transforming growth factor-β (TGF-β) [6]. The cellular responses to TGF-β are mediated through two major pathways; the canonical pathway involving son of mothers against decapentaplegic (Smad) proteins, and the noncanonical pathways signaled through mitogen-activated protein kinase (MAPK), phosphatidylinositol 3′-kinase (PI3K) and protein kinase B (also known as Ak strain transforming; Akt). Previous studies showed that treatment of epithelial cells with TGF-β rapidly activated mammalian target of rapamycin (mTOR) via the PI3K/Akt pathway, which correlated with increases in cell size and protein synthesis. Inhibition of mTOR by rapamycin blocked the migration of TGF-β-treated epithelial cells [7, 8]. It was recently reported that farnesol contained in plant oils inhibited TGF-β-induced EMT of LC cells by targeting the Akt/mTOR pathway [9], reiterating the importance of this signaling pathway in EMT. There are three isoforms of TGF-β, TGF-β1, β2 and β3, among which TGF-β1 is the most studied [10].
Activation of toll-like receptors (TLRs) by their agonists is critical for the development of immune responses and TLR agonists have been used as adjuvants for cancer vaccines and therapies [11, 12]. Systemic administration of TLR agonists activated innate immune responses, upregulated antibody-dependent effector functions, and increased adaptive immune responses in preclinical studies [11]. A double stranded RNA, polyinosinic-polycytidylic acid (polyI:C), was synthesized as a ligand that activates human TLR3 [11]. A phase II study of polyI:C (Poly-ICLC, Hiltonol®), a more stable form of polyI:C stabilized by adding poly-L-lysine and carboxymethylcellulose [13], in patients with malignant embryonal rhabdomyosarcoma, was shown to have a dramatic antitumor effect. These anti-tumor effects are considered caused by stimulating the immune system [14].
In addition to the immunological effects, polyI:C appears to directly induce apoptosis in cancer cells via TLR3 [15, 16], and suppress their motility [16,17,18]. It is well known that a wide variety of cytokines and growth factors are present in tumor microenvironments and cancer cells are exposed to those factors, including TGF-β. During our research, we observed that the treatment of the A549 human lung adenocarcinoma cells with control double-stranded siRNA inhibited TGF-β1-induced EMT. Since A549 cells express TLR3 [19], we hypothesized that activation of TLR3 with polyI:C may interfere with TGF-β1-induced EMT. In this report, we tested our hypothesis by using A549 cells as a model and found that polyI:C suppresses the motility of TGF-β1-treated A549 cells by counteracting TGF-β1-induced Akt phosphorylation without affecting the phosphorylation of Smad3 and the expression of E-cadherin, N-cadherin or Snail. Thus, polyI:C exert its anti-tumor effect by suppressing the cell motility enhanced by TGF-β, and polyI:C may be useful to prevent or reduce the metastasis of lung cancer cells.
Materials and methods
Cell culture
A549 [20] and NCI-H1993 NSCLC cells were obtained from the American Type Culture Collection (Manassas, VA) and grown in RPMI‐1640 tissue culture medium (Sigma-Aldrich, St. Louis, MO) supplemented with 10% heat-inactivated (HI)-fetal bovine serum (FBS, Gibco, Grand Island, NY), 100 U/ml penicillin and 100 μg/ml streptomycin (Sigma-Aldrich) at 37 °C in a humidified atmosphere containing 5% CO2. One million cells in the medium were seeded into 60-mm tissue culture dishes (BD Falcon, Cat. No. 353002). After incubation at 37 °C for 24 h, medium was replaced by fresh RPMI 1640 containing 0.5% HI-FBS with or without 10 μg/ml polyI:C (PolyI:C HMW, InvivoGen, San Diego, CA) and/or 10 ng/ml recombinant human TGF-β1 (Peprotech, Rocky Hill, NJ), and cells were incubated for additional 24 h. In some experiments, 5 mM perifosine (Cell Signaling Technology, Danvers, MA) or 10 μM VO-OHpic (Santa Cruz Biotechnology, Santa Cruz, CA) was used to inhibit Akt or phosphatase and tensin homologue (PTEN), respectively.
Western blotting
After cultured under different conditions, cells were directly lysed in 2% SDS lysis buffer (2% SDS, 62.5 mM Tris–HCl, 10% glycerol and 50 mM DTT). Cell lysates were incubated immediately at 95 °C for 5 min, sonicated briefly, spun, and the supernatants were stored at − 80 °C until use. Ten µg of cell lysates were loaded into wells of 4–12% polyacrylamide gels (Thermo Fisher Scientific, Waltham, MA, USA) and proteins were separated by sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and then transferred onto PVDF membranes by a wet transfer system using 1 × transfer buffer (3 g Tris, 14.4 g glycine and 200 ml ethanol in 800 ml dH2O). After overnight exposure to an appropriate primary antibody at 4 °C, the membranes were rinsed and incubated with either anti-rabbit or anti-mouse secondary IgG conjugated with horseradish peroxidase (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Target proteins on the membranes were visualized using ImmunoStar LD reagent (Wako Pure Chemicals, Tokyo, Japan). The intensity of detected bands was analyzed by the ImageJ image analysis software developed by the NIH [21]. Primary antibodies against E-cadherin (#3195), N-cadherin (#4061), Snail (#3879), p-Akt (#9271), Akt (#9272), p-PI3K (#4228), PI3K (#4292), p-Smad3 (#9520), Smad3 (#9523), TLR3 (#6961), p-Erk (#4370), Erk (#4695), p-P38 (#9211), P38 (#9212), p-JNK (#9251), JNK (#9252), PTEN (#9552), p-GSK-3α/β(#9331), GSK-3α/β(#5676), p-β-Catenin (#9561), β-Catenin (#9562), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (#5174), were purchased from Cell Signaling Technology (Danvers, MA).
RNA extraction and quantitative real-time PCR (RT-qPCR)
Total RNA was extracted from cultured cells using High Pure RNA Isolation Kit (Roche Applied Science, Mannheim, Germany) and cDNA was reverse transcribed (RT) using a kit from Thermo Fisher Scientific. The expression of each gene was determined by quantitative polymerase chain reaction (qPCR) on a StepOne™ Real-time PCR system with Taqman PCR master mix (Applied Biosystems, Foster City, CA, USA). Primers for PTEN (Hs02621230_s1) and GAPDH (Hs02758991_g1) were from Thermo Fisher Scientific. The expression level of each gene was adjusted by examining the expression level of the GAPDH gene used as an internal control, and relative mRNA expression was calculated using the ∆∆CT method.
Cell proliferation assay
The effect of polyI:C on cell proliferation weas determined using cell counting kit-8 (CCK-8, Dojindo Laboratories, Kumamoto, Japan). Briefly, A549 cells (1.5 × 105 cells in 100 ml medium) were plated in 96- well microplates. After incubation at 37 °C in a humidified 5% CO2 incubator for 24 h, various concentrations of polyI:C (0, 5, 10, 20 μg/ml) were added and the plates were incubated for additional 24 and 48 h with or without 10 ng/ml TGF-β1. A549 cells were also incubated with 10 μg/ml polyI:C without TGF- β1. The cell number was quantitated by adding 10 μl CCK-8 solution to each well of 96-well tissue culture plates and incubating the plates for 1 h at 37 °C. Absorbance at 450 nm was then measured on a microplate reader (SH-1000Lab, Corona Electric Co., Ibaraki, Japan).
Cell scratch wound healing assay
Cell scratch wound healing assay was performed according to the method previously described [22]. Briefly, 2 × 106 A549 cells were plated in 60-mm plates and cultured until they became 90% confluent. Wounds were made by gently scratching cell monolayers with sterile yellow pipette tips and plates were incubated for 48 h in the presence of 10 μg/ml polyI:C alone or in combination with 10 ng/ml TGF-β1 in RPMI 1640 supplemented with 0.5% HI-FBS. Wounded area at each time point is presented as the percentage of the original wounded area at time 0.
Matrigel cell invasion assay
Cell invasion assay was performed as previously described [23]. Cells were harvested by trypsin- ethylenediamine tetraacetic acid (EDTA) (Gibco) at 90% confluency and resuspended in RPMI with 0.5% HI-FBS. Two hundred-fifty thousand cells (in 1 ml medium) were added to rehydrated inserts of BioCoat™ Matrigel® Invasion Chamber (Corning, catalog number: 354578), and the inserts were placed in 24-well plates containing RPMI 1640 containing 10% HI-FBS. After a 22-h incubation, cells that remained above the insert membrane were wiped off using cotton tips. Cells attaching the bottom of the filters were fixed in 100% methanol for 2 min and stained with crystal violet. Invaded cells in four high-power fields (HPFs) were counted by phase contrast microscopy (CKX41; Olympus, Tokyo, Japan).
Statistical analysis
Statistical significance was analyzed by Student t test using the Prism software (GraphPad Software, San Diego, CA, USA). A value of p < 0.05 was considered statistically significant.
Results
PolyI:C suppresses the migration of TGF-β1-treated A549 cell
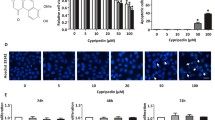
We first examined the effect of polyI:C on the proliferation of the human A549 lung cancer cells. A549 cells were cultured for 24 or 48 h in the absence of TGF-β1 or polyI:C or in the presence of each stimulant alone or in combination, and the proliferation rates were evaluated by the CCK-8 assay. Addition of polyI:C at the concentration range up to 10 μg/ml had no significant effect in the absence or presence of TGF-β1, but it caused a slight but significant decrease at its highest concentration of 20 μg/ml only in the presence of TGF-β1 (7.9% inhibition at 24 h and 10% inhibition at 48 h) (Fig. 1a). Ten μg/ml polyI:C showed modest growth inhibition (approximately 10%) at 72 h, but no acute cytotoxicity (Fig. 1b). We used 10 μg/ml of polyI:C for the rest of our study.

The effects of polyI:C on the proliferation and shape change of A549 cells treated with TGF-β1. a A549 cells (1.5 × 104) were cultured with 10 ng/ml TGF-β1 plus different doses of polyI:C for 24 or 48 h and the number of cells in each well was quantitated using CCK-8. Data is presented as mean ± SEM. **p < 0.001, n = 12. b A549 cells (1.5 × 104) were cultured with 10 μg/ml polyI:C for up to 96 h and the number of cells in each well was quantitated using CCK-8. Data is presented as mean ± SEM. *p < 0.05, n = 12. c A549 cells (2.5 × 105) were incubated in 60-mm plastic dishes as indicated for 24 h and changes in cell morphology were examined under microscopy (× 200). TGF-β1; 10 ng/ml, polyI:C; 10 μg/ml
We next examined its effects on the morphological change. As previously reported [7], untreated A549 cells showed a cobblestone shape (Fig. 1c, left upper panel). TGF-β1 induced morphological changes characterized by elongated spindle shape and the space between cell clusters became less (Fig. 1c, left lower panel). PolyI:C alone did not induce any significant morphological changes (Fig. 1c, right upper panel); however, it partly inhibited TGF-β1-induced morphological changes and elongated cell shape was less obvious (Fig. 1c, right lower panel), suggesting that polyI:C may inhibit TGF-β1-induced cell motility and invasiveness of A549 cells. Almost identical results were obtained by using NCI-H1993 cells (Supplementary Fig. 1a).
To test the effects of polyI:C on the migration of A549 cells, we used a cell scratch wound healing assay. As shown in Fig. 2a, b, TGF-β1 significantly accelerated the restoration of the wounds. Although polyI:C did not alter the restoration of the wound-width by itself, it significantly suppressed TGF-β1-induced restoration of wound-width at 48 h (Fig. 2a, b). We next used a matrigel cell invasion assay. TGF-β1 significantly promoted the invasion of A549 cells. PolyI:C did not affect the invasion of A549 cells by itself (in the absence of TGF-β1), but it inhibited the invasion of A549 cells in the presence of TGF-β1 (Fig. 2c). The effect of polyI:C was dose-dependent (Fig. 2d). These results indicated that polyI:C can inhibit the motility and invasiveness of A549 cells activated by TGF-β1.

The effects of polyI:C on TGF-β1-induced migration and invasion of A549 cells. a A549 cells (2 × 106) were incubate at 37 °C for overnight in RPMI-1640 containing 10% HI-FBS. Wounds were made by gently scratching cell monolayers with sterile yellow pipette tips, and monolayers were cultured for 48 h in RPMI 1640 supplemented with 0.5% HI-FBS containing 10 μg/ml polyI:C alone, 10 ng/ml TGF-β1 alone or both. Wounded areas were observed under microscopy and photos were taken. × 100. b Wounded areas were quantitated and represented as the percentage of wounded area at time 0. Data is presented as mean ± SEM of four measures of each wounded area. The summary of ten independent experiments. ***p < 0.001. c A549 cells (2.5 × 105) were suspended in 0.5 ml RPMI-1640 containing 0.5% HI-FBS and 10 μg/ml polyI:C, 10 ng/ml TGF-β1 or both and placed in the inner wells of BioCoat™ Matrigel® Invasion Chamber and incubated at 37 °C for 22 h. Membranes were fixed, stained and the number of invaded cells was counted under phase contrast microscopy. Data is presented as mean ± SEM. The summary of two independent experiments. *p < 0.05, **p < 0.01. n = 8. d Different doses of polyI:C and 10 ng/ml TGF-β1 were used. Data is presented as mean ± SEM. The summary of two independent experiments. *p < 0.05, ***p < 0.001
PolyI:C suppresses Akt phosphorylation in TGF-β1-treated A549 cells
To study the mechanisms by which polyI:C suppresses cell motility and invasiveness induced by TGF-β1, we examined the level of TLR3 present in A549 cells cultured in different conditions. Similar levels of TLR3 were detected by Western blotting after culture with either TGF-β1, polyI:C alone or TGF-β1 plus polyI:C (Fig. 3a).

The effects of polyI:C on the expression of TLR3 and EMT markers and on the phosphorylation of Smad3 and MAPKs. a One million A549 cells were incubated in 60-mm plastic dishes for 24 h at 37 °C in RPMI 1640 with 10% HI-FBS. After the incubation, cells were incubated for additional 24 h in fresh medium containing 0.5% HI-FBS with 10 ng/ml TGF-β1, 10 μg/ml polyI:C or both. The level of each protein and the level of phosphorylation were evaluated by Western blotting. b The level of phosphorylation in each cell lysate was evaluated by Western blotting, followed by analyses using densitometry (lower panels). Data is present as mean ± SEM. A summary of three independent experiments. **p < 0.01, ***p < 0.001. c A549 cells (1 × 106) were suspended in RPMI-1640 containing 10% HI-FBS. After a 24-h incubation at 37ºC, medium was replaced by fresh medium containing 0.5% HI-FBS and cells were incubated in the presence of 10 ng/ml TGF-β1 with or without 5 mM perifosine at 37 °C for 24 h. The level of phosphorylated Akt at Ser 473 was evaluated by Western blotting. d Two hundred and fifty thousand A549 cells were suspended in 0.5 ml RPMI 1640 containing 0.5% HI-FBS and incubated for 24 h with 10 ng/ml TGF-β1 in the presence or absence of 5 mM perifosine. Results are representative of two independent experiments. Data is present as mean ± SEM per HPF. ***p < 0.001. n = 8
As noted above, TGF-β1 induces cell motility in cancer cells, and it also induces epithelial mesenchymal transformation (EMT) by up-regulating Snail via a Smad-dependent manner, and repressing E-cadherin which is thought to suppress metastasis during tumor progression [24]. EMT is also accompanied by an increase in N-cadherin expression [7]. In agreement with the previous observation, TGF-β1 treatment induced the phosphorylation of Smad3, increased the protein level of N-cadherin and Snail, and reduced the protein level of E-cadherin in A549 cells. PolyI:C showed little effects on the expression of these molecules (Fig. 3a). In addition, polyI:C did not alter the level of p-ERK, p-P38 or p-JNK in TGF-β1-treated cells (Fig. 3a). Thus, treatment with polyI:C appeared to inhibit the motility and invasiveness of A549 cells undergoing EMT in response to TGF-β1 by a Smad-independent mechanism.
It was previously shown that the activation of the PI3K/Akt pathway plays a critical role in increased migratory ability of murine mammalian epithelial cells treated with TGF-β1 and undergoing EMT [7]. Since polyI:C inhibited TGF-β1-induced increased migration without altering the protein level of E-cadherin, N-cadherin or Snail, we hypothesized that polyI:C may target the PI3K/Akt pathway. As shown in Fig. 3b, TGF-β1 increased the phosphorylation of PI3K and Akt. PolyI:C did not reduce the TGF-β1-induced PI3K phosphorylation level, but markedly reduced the Akt phosphorylation level. Inhibition of Akt phosphorylation by the Akt inhibitor perifosine [25] (Fig. 3c) suppressed the migration of TGF-β1-treated A549 cells (Fig. 3d), indicating a critical role of Akt in the migration of TGF-β1-treated cells. Reduced TGF-β1-induced Akt phosphorylation by polyI:C was also detected in NCI-H1993 cells (Supplementary Fig. 1b).
PolyI:C suppresses Akt phosphorylation in TGF-β1-treated A549 cells partly by upregulating PTEN expression
PTEN is shown to downregulate the PI3K/Akt pathway by dephosphorylating PIP3; thus, it opposes PI3K activity and subsequently downregulates Akt [26]. To examine a potential contribution of PTEN to the polyI:C-mediated inhibition of Akt phosphorylation and cell migration in TGF-β1-treated A549 cells, we first evaluated the level of PTEN. As shown in Fig. 4a, b, polyI:C modestly but significantly increased the level of PTEN protein and mRNA in TGF-β1-treated A549 cells by Western blotting and RT-qPCR, respectively. TGF-β1-induced Akt phosphorylation was dose-dependently reduced by polyI:C, whereas the level of PTEN protein increased (Fig. 4c).

The effects of polyI:C on the PTEN/Akt signaling pathway. a A549 cells (1 × 106) were incubated in 60-mm plastic dishes at 37 °C for 24 h. Medium was replaced by fresh medium containing 0.5% HI-FBS and 10 ng/ml TGF-β1, 10 μg/ml polyI:C or both for 24 h. The level of PTEN in each cell lysate was evaluated by Western blotting, followed by densitometry (right panel). Data is present as mean ± SEM. The summary of three independent experiments. **p < 0.01. b Expression of PTEN mRNA was analyzed by quantitative RT-PCR. The summary of two independent experiments. Data is present as mean ± SEM. **p < 0.01. c A549 cells (1 × 106) were suspended in RPMI-1640 medium containing 10% HI-FBS. After 24 h incubation at 37 °C, medium was replaced by fresh medium containing 0.5% HI-FBS and 10 ng/ml TGF-β1 with different doses of polyI:C. The level of each protein in each cell lysate and the level of phosphorylation were evaluated by Western blotting, followed by densitometry (lower panels). Data is present as mean ± SEM. The summary of four independent experiments. *p < 0.05, **p < 0.01, ***p < 0.0001
To further examine the role of PTEN in TGF-β1-induced cell migration and invasion and their suppression by polyI:C, we used a PTEN-specific inhibitor, VO-OHpic [27, 28]. Among several PTEN inhibitors, VO-OHpic was found to be most potent and specific [28]. As shown in Fig. 5a, TGF-β1-induced Akt phosphorylation (lane 1) was reduced by polyI:C (lane 2). When VO-OHpic was used with TGF-β1 and polyI:C, it tended to reverse polyI:C-mediated inhibition of Akt phosphorylation although the difference was not statistically significant (lane 3). By a cell invasion assay, VO-OHpic again showed a tendency to reverse the inhibitory effect by polyI:C on the invasion of TGF-β1-treated cells (Fig. 5b). These results suggested a role for PTEN in polyI:C-mediated inhibition of Akt phosphorylation and invasion in TGF-β1-treated A549 cells.

The effects of VO-OHpic on the inhibition of Akt phosphorylation and invasion of TGF-β1-treated cells by polyI:C. a A549 cells (1 × 106) were suspended in RPMI-1640 medium containing 10% HI-FBS. After 24 h incubation at 37 °C, the medium was replaced by fresh medium containing 0.5% HI-FBS and incubated at 37 °C for 24 h as indicated. TGF-β1; 10 ng/ml, polyI:C; 10 μg/ml, VO-OHpic; 10 μM. The level of Akt and the phosphorylation at Ser 473 were evaluated by Western blotting. Data is present as mean ± SEM. *p < 0.05. Results are representative of three independent experiments. b A549 cells (2.5 × 105) were pre-treated with 10 μM VO-OHpic for 1 h at 4 °C and then seeded in Matrigel® Invasion Chamber. After a 22-h incubation with 10 ng/ml TGF-β1 with or without 10 μg/ml polyI:C, cells that migrated through the membranes of the inserts were counted under phase contrast microscopy. Data is presented as mean ± SEM per HPF. *p < 0.05, ***p < 0.001. Results are representative of three independent experiments
Discussion
Studies using polyI:C and its analogs used as a cancer adjuvant have produced encouraging data in both human cancer patients and murine cancer models [13, 29]. In addition to its function in immune responses, polyI:C was demonstrated to inhibit the growth and migration of TLR3-expressing tumor cells in vitro [16]. However, tumor cells are exposed to an array of cancer-promoting factors in cancer microenvironments; thus, it remains unclear whether polyI:C has the same effects on cancer cells in the presence of cytokines, such as TGF-β1 that induces EMT. Here, we demonstrated our novel results that polyI:C treatment significantly suppressed the migration and invasion of TGF-β1-treated A549 cells. This effect by polyI:C was via inhibition of Akt phosphorylation. We identified PTEN as a molecule partly involved in the process. Thus, polyI:C can inhibit cancer progression by not only upregulating immune responses but also inhibiting the motility and subsequent invasion and metastasis.
EMT leads to an increased ability in cell motility and invasion, and it occurs during cancer progression. TGF-β is an inducer of EMT and it acts through Smad proteins-dependent and Smad proteins-independent pathways. It was previously reported that TGF-β increases cell size and migration by rapidly activating mammalian target of rapamycin (mTOR) via the PI3K/Akt pathway [30]. In the present study, polyI:C did not alter either downregulation of E-cadherin or upregulation of N-cadherin and Snail levels in TGF-β1-treated A549 cells, which are regulated by the Smad3 pathway. By contrast, polyI:C inhibited TGF-β1-induced Akt phosphorylation and cell migration. These results suggest that the PI3K/Akt pathway is the specific target of polyI:C.
To determine a potential mechanism of polyI:C-mediated inhibition of Akt phosphorylation in TGF-β1-treated cells, we examined the role for several signaling molecules, such as MAPKs and PTEN, and identified PTEN as a candidate molecule. PTEN opposes PI3K activity, leading to the inactivation of Akt [31], and PTEN could inhibit cell migration [32]. The elevated level of PTEN in A549 cells treated with TGF-β1 plus polyI:C appears to occur at a transcriptional level because we detected increased PTEN mRNA expression. However, exact mechanism that leads to the increased PTEN level remains unclear.
GSK-3 is a serine/threonine protein kinase active in resting cells and inactivated by phosphorylation [33]. GSK-3 is downstream of several major signaling pathways, including the PI3K/Akt pathway, and the major kinase to phosphorylate β-catenin, leading to its degradation. A previous study showed that GSK-3β, an isoform of GSK-3, is a critical signaling molecule for TGF-β1-induced EMT in the ARPE-19 retinal epithelial pigment cells [34]. Therefore, GSK-3β is another candidate molecule regulating polyI:C-mediated inhibition of Akt phosphorylation and migration induced by TGF-β1. We examined the phosphorylation of GSK-3β in A549 cells and did not find any effect by polyI:C (date not shown), indicating that the Akt/GSK-3β/β-catenin pathway does not play a role (Supplementary Fig. 2a).
PolyI:C induces type I interferon (IFN) production by TLR3-expressing cells, such as fibroblasts [35], and IFNs produced by A549 cells in response to polyI:C may have affected Akt phosphorylation and motility. Therefore, we examined a potential role of autocrine production of type I IFNs in our model. Treatment with polyI:C inhibited the TGF-β1-induced Akt phosphorylation but had no effect on the phosphorylation of Stat1. This suggested that polyI:C did not cause IFNs production by A549 cells to induce Stat1 phosphorylation; thus, it is unlikely that the induction of IFN production by polyI:C was responsible for the inhibition of TGF-β1-induced Akt phosphorylation in A549 cells (Supplementary Fig. 2b, c).
We demonstrated a novel function of polyI:C that contributes to the inhibition of cancer progression. However, there are some limitations. We used A549 cells for all experiments and some of the results were validated by using NCI-H1993 cells. These cells were morphologically very similar. We tested additional lung carcinoma cells, such as the human HCC4006 lung adenocarcinoma and the mouse Lewis lung carcinoma cells. These cells showed the morphology of cells that have gone through EMT without any additional treatment and there was no additional induction of Akt phosphorylation after TGF-β1 treatment (data not shown), suggesting that the polyI:C effects found in this study may be limited to cells at a certain stage of cancer cell development. We also considered to determine the role of polyI:C in vivo using a mouse model; however, since polyI:C affects the growth of cancer cells via activation of the immune response, it is difficult to evaluate its direct effects on cancer cells in vivo.
Conclusion
PolyI:C and its analogues are being clinically tested as a cancer vaccine adjuvant for anti-cancer treatment. A phase II study in patients with malignant embryonal rhabdomyosarcoma showed a dramatic antitumor effect [14]. In clinical studies of subjects with melanoma or advanced renal cell cancer [13], the administration of Poly-ICLC could be well-tolerated, but no clinical benefit was observed. The results of our study suggest that polyI:C can significantly reduce the metastatic potential of lung cancer cells undergoing EMT; thus, polyI:C and its analogue may be listed as an option to be used in combination with existing other anti-cancer therapies.
References
Dela Cruz CS, Tanoue LT, Matthay RA (2011) Lung cancer: epidemiology, etiology, and prevention. Clin Chest Med 32:605–644
Lemjabbar-Alaoui H, Hassan OU, Yang YW, Buchanan P (2015) Lung cancer: biology and treatment options. Biochim Biophys Acta 1856:189–210
Siegel RL, Miller KD, Fuchs HE, Jemal A (2021) Cancer statistics, 2021. CA Cancer J Clin 71:7–33
Xie S, Wu Z, Qi Y, Wu B, Zhu X (2021) The metastasizing mechanisms of lung cancer: recent advances and therapeutic challenges. Biomed Pharmacother 138:111450
Menju T, Date H (2021) Lung cancer and epithelial-mesenchymal transition. Gen Thorac Cardiovasc Surg 69:781–789
Roberts AB, Thompson NL, Heine U, Flanders C, Sporn MB (1988) Transforming growth factor-beta: possible roles in carcinogenesis. Br J Cancer 57:594–600
Lamouille S, Derynck R (2007) Cell size and invasion in TGF-beta-induced epithelial to mesenchymal transition is regulated by activation of the mTOR pathway. J Cell Biol 178:437–451
Hamidi A, Song J, Thakur N et al (2017) TGF-β promotes PI3K-AKT signaling and prostate cancer cell migration through the TRAF6-mediated ubiquitylation of p85α. Sci Signal 10:eaa14186
Lee JH, Chinnathambi A, Alharbi SA, Shair OHM, Sethi G, Ahn KS (2019) Farnesol abrogates epithelial to mesenchymal transition process through regulating Akt/mTOR pathway. Pharmacol Res 150:104504
Moses HL, Roberts AB, Derynck R (2016) The discovery and early days of TGF-β: a historical perspective. Cold Spring Harb Perspect Biol 88:a021865
Forte G, Rega A, Morello S et al (2012) Polyinosinic-polycytidylic acid limits tumor outgrowth in a mouse model of metastatic lung cancer. J Immunol 188:5357–5364
Urban-Wojciuk Z, Khan MM, Oyler BL et al (2019) The role of TLRs in anti-cancer immunity and tumor rejection. Front Immunol 10:2388
Gnjatic S, Sawhney NB, Bhardwaj N (2010) Toll-like receptor agonists: are they good adjuvants. Cancer J 16:382–391
Salazar AM, Erlich RB, Mark A, Bhardwaj N, Herberman RB (2014) Therapeutic in situ autovaccination against solid cancers with intratumoral poly-ICLC: case report, hypothesis, and clinical trial. Cancer Immunol Res 2:720–724
Jiang Q, Wei H, Tian Z (2008) Poly I:C enhances cycloheximide-induced apoptosis of tumor cells through TLR3 pathway. BMC Cancer 8:12
Lau WH, Zhu XG, Ho SWT, Chang SC, Ding JL (2017) Combinatorial treatment with polyI:C and anti-IL6 enhances apoptosis and suppresses metastasis of lung cancer cells. Oncotarget 8:32884–32904
Guo Z, Chen L, Zhu Y et al (2012) Double-stranded RNA-induced TLR3 activation inhibits angiogenesis and triggers apoptosis of human hepatocellular carcinoma cells. Oncol Rep 27:396–402
Ma B, Herzog EL, Moore M et al (2016) RIG-like helicase regulation of chitinase 3-like 1 axis and pulmonary metastasis. Sci Rep 6:26299
Tissari J, Sirén J, Meri S, Julkunen I, Matikainen S (2005) IFN-alpha enhances TLR3-mediated antiviral cytokine expression in human endothelial and epithelial cells by up-regulating TLR3 expression. J Immunol 174:4289–4294
Lieber M, Smith B, Szakal A, Nelson-Rees W, Todaro G (1976) A continuous tumor-cell line from a human lung carcinoma with properties of type II alveolar epithelial cells. Int J Cancer 17:62–70
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9:671–675
Liang CC, Park AY, Guan JL (2007) In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nat Protoc 2:329–333
Repesh LA (1989) A new in vitro assay for quantitating tumor cell invasion. Invasion Metastasis 9:192–208
Wang Y, Shi J, Chai K, Ying X, Zhou BP (2013) The role of snail in EMT and tumorigenesis. Curr Cancer Drug Targets 13:963–972
Kondapaka SB, Singh SS, Dasmahapatra GP, Sausville EA, Roy KK (2003) Perifosine, a novel alkylphospholipid, inhibits protein kinase B activation. Mol Cancer Ther 2:1093–1103
Govender D, Chetty R (2012) Gene of the month: PTEN. J Clin Pathol 65:601–603
Rosivatz E, Matthews JG, McDonald NQ et al (2006) A small molecule inhibitor for phosphatase and tensin homologue deleted on chromosome 10 (PTEN). ACS Chem Biol 1:780–790
Zhu X, Shao ZH, Li C et al (2014) TAT-protein blockade during ischemia/reperfusion reveals critical role for p85 PI3K-PTEN interaction in cardiomyocyte injury. PLoS One 9:e95622
Varthaman A, Moreau HD, Maurin M, Benaroch P (2016) TLR3-induced maturation of murine dendritic cells regulates CTL responses by modulating PD-L1 trafficking. PLoS One 11:e0167057
Xu J, Lamouille S, Derynck R (2009) TGF-beta-induced epithelial to mesenchymal transition. Cell Res 19:156–172
Georgescu MM (2010) PTEN tumor suppressor network in PI3K-Akt pathway control. Genes Cancer 1:1170–1177
Leslie NR, Yang X, Downes CP, Weijer CJ (2005) The regulation of cell migration by PTEN. Biochem Soc Trans 33:1507–1508
Yucel G, Oro AE (2011) Cell migration: GSK3β steers the cytoskeleton’s tip. Cell 144:319–321
Huang L, Zhang C, Su L, Song Z (2017) GSK3β attenuates TGF-β1 induced epithelial-mesenchymal transition and metabolic alterations in ARPE-19 cells. Biochem Biophys Res Commun 486:744–751
Matsumoto M, Kikkawa S, Kohase M, Miyake K, Seya T (2002) Establishment of a monoclonal antibody against human toll-like receptor 3 that blocks double-stranded RNA-mediated signaling. Biochem Biophys Res Commun 293:1364–1369
Acknowledgements
We are grateful to the members of the Department of Pathology and Experimental Medicine, Okayama University, for their useful discussion.
Funding
This study was funded by the Department of Pathology and Experimental Medicine, Okayama University.
Author information
Authors and Affiliations
Contributions
Conceptualization: TY; Formal analysis and investigation: TY, GT; Writing—TY, TY; Writing—review and editing: TY, TO, MF, AM; Funding acquisition: MF, TO, AM.
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Yamaguchi, T., Yoshimura, T., Ohara, T. et al. PolyI:C suppresses TGF-β1-induced Akt phosphorylation and reduces the motility of A549 lung carcinoma cells. Mol Biol Rep 48, 6313–6321 (2021). https://doi.org/10.1007/s11033-021-06625-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-021-06625-1