
Abstract
Rice (Oryza sativa L.) is a major food for more than half of the world’s population and serves as a model monocot plant. DEP1 (dense and erect panicle 1), corresponding to an erect-panicle architecture, may bring the third breakthrough in rice breeding after the introduction of the semi-dwarf trait and the application of heterosis in hybrid rice. The aim of this study was to determine the effects of DEP1 on yield components and vascular bundle-related traits under indica and japonica genetic backgrounds. We analyzed a series of recombinant inbred lines, which were derived from a cross between the japonica variety SN265 with an erect panicle and the indica variety R99 with a curved panicle. The results showed that effects of the DEP1/dep1 allele were much stronger than the effects of the indica/japonica genetic background on vascular bundle-related traits and yield components. As the frequency of indica alleles increased, the grain yield per plant and the harvest index significantly increased in lines with the dep1 allele. Among the indica-type lines, those with the dep1 allele showed superior values for almost all of the vascular bundle-related traits and yield components. Our results provide new insights into the use of a high-yield-related gene derived from japonica in the indica genetic background.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rice is a major food for more than half of the world’s population, and it serves as a model plant for monocots. With continuing population growth and the decreasing availability of arable land, increased grain yield per unit area is one of the most important goals of rice production and scientific research.
In rice breeding, two major breakthroughs have been made in the last century. The first breakthrough, known as the ‘Green Revolution,’ was the breeding of semi-dwarf lines that greatly enhanced rice yields in the 1960s (Suh and Heu 1978). The second breakthrough was the application of heterosis in hybrid rice, which further increased rice yields in the 1970s (Virmani et al. 1982; Yuan 1998b). To achieve the third breakthrough, national and international rice breeding programs have focused on improving crop productivity by selecting for superior grain yield components and ideal plant architecture (Huang et al. 2009; Khush 1999; Peng et al. 2008; Yuan 1998a). Recently, several genes controlling the yield components of rice have been identified. These genes include GW2, qSW5, and GS5, which contribute to grain width (Li et al. 2011; Shomura et al. 2008; Song et al. 2007), GS3 that regulates grain length (Fan et al. 2006), and FZP and LAX1, which control the number of branches per panicle (Komatsu et al. 2003; Oikawa and Kyozuka 2009). Among these genes, DEP1 has been extensively researched and widely used in rice breeding (Huang et al. 2009; Wang et al. 2009; Zhou et al. 2009).
DEP1 (EP/qPE9-1) was successfully cloned and characterized by three research groups independently (Huang et al. 2009; Wang et al. 2009; Zhou et al. 2009). The dominate allele dep1 at the DEP1 locus corresponds to a gain-of-function mutation causing truncation of a phosphatidylethanolamine-binding protein-like domain protein. This mutation was shown to be responsible for the erect-panicle architecture and increased grain yield (Huang et al. 2009). A recent study demonstrated that DEP1 encodes a conventional plant-specific Gγ subunit protein domain that interacts strongly with RGB1 (a rice Gβ subunit protein). Plants with the dep1 allele showed nitrogen-insensitive vegetative growth alongside increased nitrogen uptake and assimilation, which resulted in improved harvest index and grain yield (Sun et al. 2014). These effects of DEP1 make it pivotal in rice breeding. However, there has been little consensus about the effects of DEP1 under different genetic backgrounds. The dominant allele at the DEP1 locus in the japonica variety Shao313 caused increased grain yield per plant, resulting from an increase in the number of grains per panicle (Huang et al. 2009). In the indica variety R6574, the erect-type allele at the DEP1 locus caused an increase in the number of grains per panicle, but the grain yield per plant was decreased because of lower 1000-grain weight and fewer panicles (Zhou et al. 2009). Similar results were observed in Nongken57, a japonica variety (Yi et al. 2011). These results and observations indicated that the effects of DEP1 may differ under different genetic backgrounds.
In rice, the vascular bundle system provides mechanical support for the plant body (Zhang et al. 2002). Panicle architecture is closely related to the number of vascular bundles. Compared with curved-panicle varieties, erect-panicle varieties have more large and small vascular bundles (Huang et al. 2009). The number of vascular bundles differs between indica and japonica; this is one of the traits that can be used to differentiate between indica and japonica varieties (Zhang et al. 2002). Consequently, the number of vascular bundles is strongly affected by the indica/japonica genetic background and by the erect-/curved-panicle architecture. However, few studies have analyzed the effects of interactions between subspecies and panicle architecture on vascular bundle-related traits.
In this study, we used a series of recombinant inbred lines (RILs) derived from the cross between the japonica variety SN265 with an erect panicle and the indica variety R99 with a curved panicle to elucidate the effects of DEP1 on yield components and vascular bundle-related traits under indica and japonica genetic backgrounds.
Materials and methods
Plant materials
We used 200 RILs derived from the cross between SN265 and R99. SN265 is widely cultivated in northern China. In this variety, a 12-bp nucleotide sequence replaces a 637-bp region in the middle of exon 5 at the DEP1 locus. This results in erect panicles and an increased number of grains per plant and consequently increased grain yield (Huang et al. 2009). R99 is an indica restore line with a curved panicle. Alleles with sequences matching the Nipponbare reference genome sequence are represented by DEP1, while high-yield alleles responsible for erect panicles are represented by dep1. SN265 harbors the dep1 allele, and R99 harbors the DEP1 allele. The amino acid sequence alignment of the DEP1 locus in SN265 and R99 is shown in Figure S1.
Field experiments
All of the RILs were grown in a rice paddy field at the experimental farm of Shenyang Agricultural University, Shenyang, China (41.8°N; 123.4°E), during the summer of 2014. Seeds were sown in a seedling nursery on April 24, 2014, with one seedling transplanted per hill on 23 May. At least 120 plants of each RIL were transplanted at 30 cm × 15 cm spacing. The RILs were arranged in a randomized block design with three replications, and each replication included at least 40 plants. Fertilizer was applied as basal dressing at an application rate of 75 kg ha−1 N, 150 kg ha−1 P, and 75 kg ha−1 K. Three weeks after transplanting, DNA was extracted from leaves of eight plants per line to determine the frequency of indica-type markers.
Evaluation of vascular bundles
We evaluated the vascular bundles after the flowering stage. Main stems with spikes were sampled from eight plants per RIL. A transverse section (approx. 1 mm thick) was cut 1 cm below the neck node and observed under a dissecting microscope. We counted the number of large vascular bundles (LVB) and small vascular bundles (SVB) in the inner parenchyma of the stem and primary branches. The ratio of the number of LVB to the number of primary branches (hereafter, the V/R ratio) was calculated for each panicle. Using the same method, we determined the V/R ratio at the upper, middle, and lower sections of the panicle. We also evaluated the vascular bundles at the second-to-top node. Images of transverse sections of the panicle neck and the second-to-top node were screened after counting the number of vascular bundles.
DNA extraction and subspecies-specific genotyping
Genomic DNA was isolated from fresh-frozen leaves using the CTAB method (Doyle 1991). In total, 90 sets of subspecies-specific insertion–deletion (INDEL) and subspecies-specific intron length polymorphism (SSILP) markers in indica and japonica rice cultivars were used to analyze the proportion of indica alleles in the genome of each RIL. The clone names and physical distances of 34 INDEL and 56 SSILP markers were obtained from the marker-based physical maps produced by the International Rice Genome Sequencing Project, based on previous studies (Sun et al. 2012; Lu et al. 2009; Zhao et al. 2009a). The two marker types were integrated into one genetic map based on physical distances using MapChart software (Fig. S2). All of the INDEL and SSILP markers were used to genotype the 200 RILs. The PCRs were performed as described elsewhere (Shen et al. 2004; Wang et al. 2006; Zhao et al. 2009b; Sun et al. 2012) using the primers listed in Table S1. The indica allele frequency was calculated as the ratio of the number of indica-type markers to the total number of markers. The DEP1 locus was genotyped as described previously (Xu et al. 2014).
Measurements of yield components
At the mature stage (35 days after flowering), the aboveground portions of eight plants per RIL were harvested from each plot. After counting the number of panicles and measuring plant height, the panicles were hand-threshed and placed in water to separate filled grains, which sank in water, from unfilled grains. Seed sterility was determined as the ratio of non-fertilized grains to total grains. To determine dry weight, the filled and unfilled grains were oven-dried at 80 °C for 2 days. The number of grains per panicle and setting rate were calculated from the above data. Eight average-sized panicles were taken from each plot to count the number of primary and secondary branches and the number of spikelets per branch.
Results
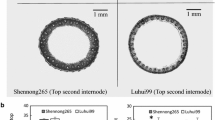
The subspecies-specific INDEL and SSILP markers were distributed among the 12 chromosomes in the rice genome, with an average of 9.9 markers on each chromosome. We calculated the indica allele frequency for all 200 RIL and then separated the RIL into four groups: indica-type lines (indica allele frequency >0.5) with an erect panicle (dep1), indica-type lines with a curved panicle (DEP1), japonica-type lines (indica allele frequency <0.5) with an erect panicle (dep1), and japonica-type lines with a curved panicle (DEP1). The proportion of the four types was 1:0.98:0.98:0.97, which indicated that the separation of the DEP1 locus was not affected by the indica/japonica genetic background. The differences in vascular bundles between R99 and SN265 are shown in Fig. 1.

Differences in vascular bundles in panicle neck and second-to-top internode between R99 and SN265. a Transverse section of panicle neck and second-to-top internode. Scale bars 100 μm. b Number of small and large vascular bundles in R99 and SN265. Values are mean ± SD (n = 10). *Significant at p < 0.05 (Student’s t test)
Table 1 summarizes the vascular bundle-related traits of the four types. Among the indica-type plants, those with the dep1 allele showed significant increases in the numbers of LVB at the neck of the panicle and the second node, the LVB (panicle)/LVB (stem) ratio, and number of SVB at the neck of the panicle, compared with plants with the DEP1 allele. Similar differences were observed between the japonica lines harboring DEP1 and dep1. However, there were no significant differences in the average values of most traits between indica-type and japonica-type lines. The lines with the dep1 allele showed significant differences from those with the DEP1 allele for all of the investigated vascular bundle-related traits. In contrast, the indica-type lines showed a significant difference from japonica-type lines only in the number of LVB at the neck of the panicle, which resulted in significant differences in the LVB/SVB and LVB (panicle)/LVB (stem) ratios. These results indicated that the effects of the DEP1/dep1 allele were much stronger than the effects of the indica/japonica genetic background on vascular bundle-related traits. Furthermore, these results showed that the indica/japonica genetic background mainly affected the number of LVB at the neck of the panicle.
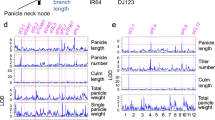
Compared with indica-type lines with the DEP1 allele, those with the dep1 allele showed superior values for almost all of the vascular bundle-related traits. Because the V/R ratio is distinguishable between indica and japonica and is affected by panicle architecture, we investigated the V/R ratio in detail. We separated the panicle into three parts: bottom (three primary branches at the base of the panicle), top (three primary branches at the top of the panicle), and middle (the remaining primary branches in the middle of the panicle). This analysis revealed that dep1 only increased the V/R ratio at the bottom part of the panicle (Fig. 2). The V/R ratio was higher in the indica-type lines than in japonica-type lines at all three parts of the panicle.

Effects of erect-panicle architecture on ratio of vascular bundles to primary rachis branches (V/R ratio). a Top, middle, and bottom of panicle. b Comparison of V/R ratio between erect and curved panicle at top, middle, and bottom of panicle. Values are mean ± SD (n = 10). *Significant at p < 0.05 (Student’s t test)
Table 2 summarizes the yield components for the four types. Among the indica-type lines, those with dep1 showed a significant increase in biomass, grain yield per plant, harvest index, number of panicles per plant, and number of grains per panicle, but a lower setting rate, compared with plants with the DEP1 allele. Comparing the indica and japonica lines with the dep1 allele, the japonica lines showed higher biomass, grain yield per plant, and seed setting rate. Like the vascular bundle-related traits, almost all of the yield components had higher values in indica-type lines with the dep1 allele than in those with the DEP1 allele. The lines with the dep1 allele showed significant differences from lines with the DEP1 allele in all yield component traits except for biomass per plant. The only significant difference between japonica and indica lines was in the setting rate, which was higher in japonica lines. These findings indicated that the effects of the DEP1/dep1 allele on yield components were much stronger than the effects of the indica/japonica genetic background.
Table 3 shows the correlation coefficients between the indica-allelic frequency and all the traits summarized in Tables 1 and 2. As the frequency of the indica alleles increased, the grain yield per plant and the harvest index significantly increased in lines with the dep1 allele; the opposite trend was observed in lines with DEP1 allele, although the differences were not statistically significant. In the plants with the dep1 allele, the number of panicles per plant was significantly negatively correlated with the number of LVB at the panicle neck, the number of LVB at the second-to-top internode, the number of SVB at the panicle neck, and the ratio of the number of LVB at the second-to-top internode to the number of LVB at the panicle neck. In contrast, none of those correlations were significant for plants with the DEP1 allele.
To identify the best genotype/background combination among the RIL (those with the optimum DEP1/dep1 allele, indica-allelic frequency, and V/R ratio to achieve higher yield), we identified the lines showing superior yield performance among the RILs and compared them with the parent lines (Table S2). We found 13 lines with high yields (>45 g per plant). For comparison, the grain yield per plant for the parent lines SN265 and R99 was 40.12 and 40.52 g, respectively. Among the 13 high-yielding lines, three harbored the DEP1 allele and the other 10 harbored the dep1 allele. The indica-allelic frequency of these lines was distributed from 0.40 to 0.79, indicating that an intermediate indica-allelic frequency may combine the advantages of indica and japonica, resulting in higher yield. The V/R ratios of these high-yielding lines were markedly higher than that of SN265. Some of the high-yielding lines had higher V/R ratios than that of R99. Thus, we concluded that the dep1 allele along with an intermediate indica-allelic frequency and a high V/R ratio may be a goal for rice breeding via crosses between indica and japonica subspecies.
Discussion
In rice, the vascular bundle system in the culms and leaf veins provides mechanical support for the plant (Zhang et al. 2002). Panicle architecture is closely related to the number of vascular bundles (Qiao et al. 2011). In this study, we found that the indica/japonica genetic background mainly affected LVB and that the effect was weaker with the dep1 allele than with the DEP1 allele. The DEP1/dep1 allele affected not only the LVB and SVB, but also the number of primary branches. These effects were stronger under the indica genetic background than under the japonica genetic background.
The V/R ratio is an important index of the relationship between vascular bundles and panicle-related traits. The V/R ratio of japonica was around 1.0, whereas that of indica ranged from 1.6 to 2.0. An increase in the V/R ratio was shown to be an effective way to improve the yield of japonica (Fukuyama et al. 1999). Compared with wild type, erect-panicle varieties have more SVB and/or LVB (Qiao et al. 2011). Both panicle architecture and the indica/japonica genetic background affect the V/R ratio. Therefore, a series of RILs derived from the cross between an erect-panicle japonica variety and a curved-panicle indica variety provided the ideal experimental material to analyze the effects of the interaction between the DEP1/dep1 allele and the indica/japonica genetic background on the V/R ratio.
The results of this study showed that both the DEP1/dep1 allele and the indica/japonica genetic background significantly affected the V/R ratio. Regardless of the DEP1/dep1 allele, there was a significant positive correlation between the V/R ratio and the frequency of indica-allelic markers. However, the V/R ratio was distributed over a large range, because it was affected by both the DEP1/dep1 allele and the indica/japonica genetic background (Fig. S3). This result indicated that we can optimize the combination of the DEP1/dep1 allele, the frequency of indica alleles, and the V/R ratio to achieve higher yields via selection of the genetic background.
The erect panicle is considered as a high-yield trait in rice production. The conclusions regarding the genetic effects of DEP1 on plant height, number of panicles, and 1000-grain weight have been consistent among different research groups (Huang et al. 2009; Wang et al. 2009; Zhou et al. 2009). However, there has been no consensus as to whether the erect panicle can increase the yield per plant. Huang et al. (2009) and Wang et al. (2009) demonstrated that the dominant allele at the DEP1 locus led to the erect-panicle architecture under the japonica genetic background, along with a significant increase in grain yield per plant. Zhou et al. (2009) reported the opposite effect of DEP1 under the indica genetic background, consistent with previous studies on the erect-panicle trait under the indica genetic background. In the present study, analyses of RIL with different genetic backgrounds clarified the effects of the genetic background and dep1 on various vascular bundle- and yield-related parameters. The frequency of indica alleles in these RIL ranged from 0.1 to 0.9, which provided a wide range of genetic backgrounds to research the effects of DEP1 under indica and japonica genetic backgrounds. The results showed that, compared with the lines with the DEP1 allele, those with the dep1 allele showed higher grain yield per plant under both indica and japonica genetic backgrounds. As the frequency of indica alleles increased, the grain yield per plant and the harvest index significantly increased in lines with the dep1 allele (Table 3). This result indicated that the dep1 allele has a superior effect under the indica genetic background than under the japonica background. The increase in grain yield per plant resulted from increased harvest index, rather than increased biomass.
Traits associated with the sink–source relationship are major determinants of plant architecture and yield potential in crops (Cui et al. 2003). In lines with the dep1 allele, the increased grain number per panicle enlarged the sink, and the greater number of vascular bundles allowed photosynthates to flow to the sink. However, if the source is unchanged, this may lead to a deficiency in the accumulation of dry matter and/or the undesirable distribution of dry matter, which are reflected by decreases in the setting rate and 1000-grain weight. Increases in the grain yield and harvest index may be caused by other factors related to DEP1, such as the heterotrimeric G protein that regulates nitrogen signaling and leads to increased harvest index and grain yield in plants with the dep1 allele (Sun et al. 2014). Thus, we hypothesize that improvements to photosynthetic efficiency may strengthen the relationship between the sink and source, consequently increasing the yield.
The grain yield of rice is determined by yield components such as the number of grains per panicle, number of panicles per plant, grain weight, and setting rate. In recent molecular studies, several genes related to natural variations in yield-related traits have been isolated from rice. These genes have been shown to increase grain yield in lines with the japonica genetic background in field experiments. For example, APO1 affected the number of grains; SCM2 led to stronger culms, and DEP1 increased yield (Terao et al. 2010; Huang et al. 2009; Ookawa et al. 2010). However, few of the newly cloned genes have been reported to increase grain yield in cultivars with the indica genetic background (Miura et al. 2011). The dep1 allele has been widely used in japonica rice breeding in northern China. Since the 1980s, several high-yielding japonica varieties with dense and erect panicles have been released for commercial production (Wang et al. 2009). Studies on whether yield-related genes that increase grain yield under the japonica genetic background will also increase yield under the indica genetic background will provide insights into gene function and will be useful for breeding high-yielding rice varieties. In this study, dep1 resulted in a superior phenotype under the indica genetic background than under the japonica background. Therefore, the introduction of dep1 into indica varieties may further enhance their grain yields. The data obtained in this study provide a theoretical foundation for the application of erect-panicle architecture in indica rice breeding.
References
Cui K, Peng S, Xing Y, Yu S, Xu C, Zhang Q (2003) Molecular dissection of the genetic relationships of source, sink and transport tissue with yield traits in rice. Theor Appl Genet 106(4):649–658
Doyle J (1991) DNA protocols for plants. In: Hewitt GM, Johnston AWB, Young JPW (eds) Molecular techniques in taxonomy, vol 57. NATO ASI Series. Springer, Berlin, pp 283–293
Fan C, Xing Y, Mao H, Lu T, Han B, Xu C, Li X, Zhang Q (2006) GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet 112(6):1164–1171
Fukuyama T, Sasahara H, Fukuta Y (1999) Variation of vascular bundle system corresponds to indica, tropical-and temperate-japonica differentiation of Asian rice (Oryza sativa L.). Breed Sci 49(1):15–19
Huang X, Qian Q, Liu Z, Sun H, He S, Luo D, Xia G, Chu C, Li J, Fu X (2009) Natural variation at the DEP1 locus enhances grain yield in rice. Nat Genet 41(4):494–497
Khush GS (1999) Green revolution: preparing for the 21st century. Genome 42(4):646–655
Komatsu K, Maekawa M, Ujiie S, Satake Y, Furutani I, Okamoto H, Shimamoto K, Kyozuka J (2003) LAX and SPA: major regulators of shoot branching in rice. Proc Natl Acad Sci 100(20):11765–11770
Li Y, Fan C, Xing Y, Jiang Y, Luo L, Sun L, Shao D, Xu C, Li X, Xiao J (2011) Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet 43(12):1266–1269
Lu B-R, Cai X, Xin J (2009) Efficient indica and japonica rice identification based on the InDel molecular method: its implication in rice breeding and evolutionary research. Prog Nat Sci 19(10):1241–1252
Miura K, Ashikari M, Matsuoka M (2011) The role of QTLs in the breeding of high-yielding rice. Trends Plant Sci 16(6):319–326
Oikawa T, Kyozuka J (2009) Two-step regulation of LAX PANICLE1 protein accumulation in axillary meristem formation in rice. Plant Cell Online 21(4):1095–1108
Ookawa T, Hobo T, Yano M, Murata K, Ando T, Miura H, Asano K, Ochiai Y, Ikeda M, Nishitani R (2010) New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat Commun 1:132
Peng S, Khush GS, Virk P, Tang Q, Zou Y (2008) Progress in ideotype breeding to increase rice yield potential. Field Crops Res 108(1):32–38
Qiao Y, Piao R, Shi J, Lee S-I, Jiang W, Kim B-K, Lee J, Han L, Ma W, Koh H-J (2011) Fine mapping and candidate gene analysis of dense and erect panicle 3, DEP3, which confers high grain yield in rice (Oryza sativa L.). Theor Appl Genet 122(7):1439–1449
Shen Y-J, Jiang H, Jin J-P, Zhang Z-B, Xi B, He Y-Y, Wang G, Wang C, Qian L, Li X (2004) Development of genome-wide DNA polymorphism database for map-based cloning of rice genes. Plant Physiol 135(3):1198–1205
Shomura A, Izawa T, Ebana K, Ebitani T, Kanegae H, Konishi S, Yano M (2008) Deletion in a gene associated with grain size increased yields during rice domestication. Nat Genet 40(8):1023–1028
Song X-J, Huang W, Shi M, Zhu M-Z, Lin H-X (2007) A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet 39(5):623–630
Suh H, Heu M (1978) The segregation mode of plant height in the cross of rice varieties. XI. Linkage analysis of the semi-dwarfness of the rice variety “Tongil”. Korean J Breed 10:1–6
Sun J, Liu D, Wang J-Y, Ma D-R, Tang L, Gao H, Xu Z-J, Chen W-F (2012) The contribution of intersubspecific hybridization to the breeding of super-high-yielding japonica rice in northeast China. Theor Appl Genet 125(6):1149–1157
Sun H, Qian Q, Wu K, Luo J, Wang S, Zhang C, Ma Y, Liu Q, Huang X, Yuan Q (2014) Heterotrimeric G proteins regulate nitrogen-use efficiency in rice. Nat Genet 46(6):652–656
Terao T, Nagata K, Morino K, Hirose T (2010) A gene controlling the number of primary rachis branches also controls the vascular bundle formation and hence is responsible to increase the harvest index and grain yield in rice. Theor Appl Genet 120(5):875–893
Virmani S, Aquino R, Khush G (1982) Heterosis breeding in rice (Oryza sativa L.). Theor Appl Genet 63(4):373–380
Wang X, Zhao X, Zhu J, Wu W (2006) Genome-wide investigation of intron length polymorphisms and their potential as molecular markers in rice (Oryza sativa L.). DNA Res 12(6):417–427
Wang J, Nakazaki T, Chen S, Chen W, Saito H, Tsukiyama T, Okumoto Y, Xu Z, Tanisaka T (2009) Identification and characterization of the erect-pose panicle gene EP conferring high grain yield in rice (Oryza sativa L.). Theor Appl Genet 119(1):85–91
Xu Q, Xu N, Xu H, Tang L, Liu J, Sun J, Wang J (2014) Breeding value estimation of the application of IPA1 and DEP1 to improvement of Oryza sativa L. ssp. japonica in early hybrid generations. Mol Breed 34(4):1933–1942
Yi X, Zhang Z, Zeng S, Tian C, Peng J, Li M, Lu Y, Meng Q, Gu M, Yan C (2011) Introgression of qPE9-1 allele, conferring the panicle erectness, leads to the decrease of grain yield per plant in japonica rice (Oryza sativa L.). J Genet Genom 38(5):217–223
Yuan L (1998a) Hybrid rice breeding for super high yield. In: 21st century, vol 10
Yuan LP (1998b) Hybrid rice breeding in China. In: Advances in hybrid rice technology Philippines: International Rice Research Institute, pp 27–33
Zhang Z-H, Li P, Wang L-X, Tan C-J, Hu Z-L, Zhu Y-G, Zhu L-H (2002) Identification of quantitative trait loci (QTLs) for the characters of vascular bundles in peduncle related to indica–japonica differentiation in rice (Oryza sativa L.). Euphytica 128(2):279–284
Zhao X, Yang L, Zheng Y, Xu Z, Wu W (2009a) Subspecies-specific intron length polymorphism markers reveal clear genetic differentiation in common wild rice (Oryza rufipogon L.) in relation to the domestication of cultivated rice (O. sativa L.). J Genet Genom 36(7):435–442
Zhao X, Yang L, Zheng Y, Xu Z, Wu W (2009b) Subspecies-specific intron length polymorphism markers reveal clear genetic differentiation in common wild rice (Oryza rufipogon L.) in relation to the domestication of cultivated rice (O. sativa L.). J Genet Genom 36(7):435–442
Zhou Y, Zhu J, Li Z, Yi C, Liu J, Zhang H, Tang S, Gu M, Liang G (2009) Deletion in a quantitative trait gene qPE9-1 associated with panicle erectness improves plant architecture during rice domestication. Genetics 183(1):315–324
Acknowledgments
This work was supported by Grants from the National Natural Science Foundation of China (No. 31430062 and 31371587) and a class general financial Grant from the China Postdoctoral Science Foundation (Grant No. 2014M560211, Postdoctoral No. 142541).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Quan Xu and Tiansheng Liu have contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Figure S1
DEP1 allele in R99 and SN265. (a) Schematic representation of DEP1 allele in R99 and SN265. Light gray boxes and dark gray boxes represent UTR region and exons, respectively. Lines separating boxes represent introns. (b) Amino acid sequences of DEP1 alleles in R99 and SN265, gray boxes indicate same alignment. (PPTX 82 kb)
Figure S2
Linkage map of INDEL and SSILP subspecies-specific markers used in this study (PPTX 136 kb)
Figure S3
Relationship between V/R ratio and frequency of indica allele markers in curved- and erect-panicle lines (PPTX 59 kb)
Rights and permissions
About this article

Cite this article
Xu, Q., Liu, T., Bi, W. et al. Different effects of DEP1 on vascular bundle- and panicle-related traits under indica and japonica genetic backgrounds. Mol Breeding 35, 173 (2015). https://doi.org/10.1007/s11032-015-0364-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11032-015-0364-8