
Abstract
The quality and yield of rice are influenced by vascular bundles and highly significant differences about size and number of vascular bundles can be found among rice subspecies. In this study, we divided a series of recombinant inbred lines, which were derived from a cross between the indica variety ‘Luhui 99’ and the japonica variety ‘Shennong 265’ into pre-indica, intermediate and pre-japonica groups using subspecies-specific markers. We completed an in-depth analysis of vascular bundles characteristics at different panicle positions. The results showed that the number of large vascular bundles (LVBs) and the ratio of the number of LVBs to the number of primary branches (V/R ratio) of the three groups was ranked as pre-indica > intermediate > pre-japonica. However, a large variation in the number of LVBs and V/R ratio was observed inside each group. Increases in the LVBs and the V/R ratio did not result in increased numbers of primary branches, but led to an increase in the number of secondary branches. With the increase in the frequency of indica-allelic markers, the differences in the grain setting rate and 1000-grain weight among the upper, middle and lower panicle sections became significant. Thus, we concluded that crosses between subspecies, followed by selection of the vascular bundle related traits and indica pedigree, can optimize the number of LVBs and the frequency of indica alleles. The simultaneous increase in both LVB and primary branch numbers, which might lead to greater rice yields, and may be a goal for breeders and scientists.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice is a staple food crop for more than half of the world’s population. Because of the continuing growth of the global population and the decrease in the availability of arable land, increasing grain yield is an important goal of rice breeders. The two subspecies of cultivated rice are, Oryza sativa ssp. indica and O. sativa ssp. japonica, but they have marked difference on morphology, and ecological requirements. The indica sub species is mainly distributed in lower latitudes while japonica is planted mostly in higher latitudes (Garris et al. 2005).
Because of rice research and the increasing demand of society, the use of indica × japonica crosses has become an important strategy in rice breeding. The traditional geographical distribution of indica and japonica varieties has changed, as has the frequency of indica alleles in these varieties. Consequently, an in-depth analysis on the different characteristics in the filial generations of indica × japonica crosses will generate information necessary to improve rice breeding efforts. The vascular bundle system provides mechanical support for the plant and is closely associated with panicle architecture (Zhang et al. 2002). Rice quality and yield are influenced by the number, size and characteristics of vascular bundles, which are responsible for transporting photosynthetic products, mineral nutrients and moisture into grains. Highly significant differences in vascular bundles were found within rice subspecies and were significantly correlated with panicle traits (Fukuyama et al. 1999). The number of vascular bundles at neck panicles in indica is greater than that in japonica, and the number of these vascular bundles is significantly positively correlated with the number of rachis branches (Ling et al. 1982). The ratio of LVBs at panicle neck to the number of LVBs at the top second internode of the stem exceeded 0.5 in indica and pre-indica. However, the results for japonica and pre-japonica were the opposite. Additionally, the ratio of the number of LVBs to the number of primary branches was nearly 1.0 in japonica and pre-japonica, but about 2.0 in indica (Chen et al. 2007).
In this study, we used a series of recombinant inbred lines (RILs), which were derived from a cross between japonica variety ‘Shennong 265’ and indica variety ‘Luhui 99’, to elucidate the characteristics of vascular bundles in the 7th filial generations. To complete this study, we needed to determine the pedigree of each RIL. The Cheng index method, which can discriminate indica and japonica rice varieties based on several morphological characteristics, grain aspect, leaf hair, glume hair, glume color at heading, internode, length of first–second rachis and phenol reaction (Zhang et al. 2011). However, this method was not ideal for determining the pedigree of the filial generations of indica × japonica crosses (Mao et al. 2009). Instead, we used a fast and accurate alternative method involving subspecies-specific DNA markers, to determine the pedigrees of filial generations of indica × japonica crosses (Shen et al. 2004; Sun et al. 2012; Wang et al. 2006). Using 34 subspecies-specific insertion/deletion (indel) and 56 subspecies-specific indel and subspecies-specific intron length polymorphism (SSILP) markers, we divided the RILs into pre-japonica, intermediate and pre-indica groups based on the frequency of indica alleles in each line. We also determined the vascular bundles characteristics and the relationships between panicles and vascular bundles at different panicle positions of the filial generations. The results from this study will provide a theoretical foundation of vascular bundle characteristics for indica × japonica crossbreeding.
Materials and methods
Plant materials
Two hundred of RILs of the F7 generation derived from a cross between ‘Shennong 265’ (a japonica variety) and ‘Luhui 99’ (an indica variety) were used in this study. Two japonica varieties (‘Nipponbare’ and ‘SA’) and two indica varieties (‘9311’ and ‘Zhongjia’) were also used in this study to compare the differences in vascular bundles between subspecies.
Field experiments
All plant materials were grown in a rice paddy field at the experimental farm of the Shenyang Agricultural University, Shenyang, China (N 41°80′, E 123°44′) during the summer of 2014. Seeds of the RILs were sown in a seedling nursery on 24 April 2014, and transplanted on 23 May. The RILs were arranged in a randomized block design with three replications, and each replication included at least 24 plants. The RILs were transplanted with 30 cm × 15 cm spacing. The soil was OrthicAcrisols at pH 7.2, 24.5 g kg−1 organic C, 0.120 % total N, 0.159 % total P and 3.126 % total K. The day before transplanting, 75 kg N ha−1 as urea, 150 kg diammonium phosphate ha−1 and 75 kg KCl ha−1 were applied. Nitrogen topdressings were applied 7 days after transplanting (75 kg ha−1 as urea). The total amounts of N, P2O5 and K2O values were 165, 69 and 45 kg ha−1, respectively.
DNA extraction and subspecies-specific genotyping
We sampled the leaves of eight plants in bulk for each RIL 3 weeks after transplanting. Genomic DNA was extracted from freshly frozen leaves using the CTAB method (Doyle 1991). In total, 90 sets of subspecies-specific indel and SSILP markers in indica and japonica rice cultivars were used to analyze the proportion of indica alleles in the genome of each RIL. Clone names and physical distances for 34 indel and 56 SSILP markers were obtained from the International Rice Genome Sequencing Project based on previous studies (Lu et al. 2009; Sun et al. 2012; Zhao et al. 2009). Additionally, we previously built a linkage map using Mapchart software based on the physical distance (Sun et al. 2012). We performed PCR reactions according to previously described procedures (Shen et al. 2004; Sun et al. 2012; Wang et al. 2006; Zhao et al. 2009) using primer listed in Online Resource 1. Amplicons of markers were resolved in 1.0 % agarose gels, stained with ethidium bromide and visualized under UV light. To determine the indica-allelic frequency, we used all 90 sets of markers for RIL genotyping. After capturing the image of electrophoretic patterns, we counted the number of bands identical to the ‘Luhui99’ band. The ratio of the number of these bands to the total number of markers (i.e., 90) was considered to indicate the indica-allelic frequency for the RIL line. We calculated the indica-allelic frequency for all the 200 RILs using this method.
Evaluation of vascular bundles
We investigated vascular bundles prior to the flowering stage. In order to avoid edge effect, eight plants at the center of each block were sampled after the heading stage, and the number of LVBs and small vascular bundles (SVBs) at the top second internode of the stem and at the panicle neck were observed in free-hand sections (~1-mm thick) using a microscope (ZEISS AXIO, Germany). We also examined vascular bundles in different areas of the panicle by dividing the panicle into upper, middle and lower sections. The three primary branches at the top of the panicle were designated the upper section, the three primary branches at the bottom of the panicle were designated the lower section, and the remaining primary branches in the middle of the panicle were designated the middle section. The V/R ratio was the ratio of LVBs in the neck panicle to the number of primary rachis branches, and the LVB ratio was the ratio of LVBs in the panicle neck to those at the top second internode of the stem. Each data set was collected by three replications, and the value was used in comparative analysis. Correlation analysis and Student’s test were performed using Excel 2013 (Microsoft, USA), and Duncan’s new multiple range method was used for statistical analysis among pre-indica, intermediate and pre-japonica group using IBM SPSS Statistics 22.0 (IBM, USA). Images of transverse sections of the panicle neck and the second node from the top of the stem were screened using Qcapture Pro 7 Image and Analysis Software (QImaging, Canada) after counting the number of vascular bundles.
Results
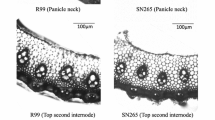
Table 1 summarizes the date regarding the rachis branches and vascular bundles within subspecies. The indica varieties have similar values of SVBs at the panicle neck, but twice the number of LVBs compare with japonica varieties. The differences in the vascular bundles between ‘Shennong265’ and ‘Luhui99’ are shown in Fig. 1. The value of LVB at top second internode in ‘Shennong265’ is similar to that of ‘Luhui99’, whereas the number of LVBs at panicle neck in ‘Shennong265’ was significantly smaller than that of ‘Luhui99’. The significant difference in the number of LVBs between indica and japonica leads to significant differences in the parameters related to LVBs, such as the ratio of LVBs to SVBs at the panicle neck, LVB ratio and the V/R ratio. The LVB ratio exceeded 0.5 in indica varieties, but was below 0.5 in japonica varieties. The V/R ratio of indica was significantly higher than that of japonica varieties. There were no significant differences in the numbers of primary and secondary branches between indica and japonica.

Difference in vascular bundles at the panicle neck and second-to-top internode between ‘Shennong265’ and ‘Luhui99’. a Transverse section of the panicle neck and second-to-top internode. Scale bars 1 mm. b Number of small and large vascular bundles in ‘Shennong265’ and ‘Luhui99’. Values are mean ± SD (n = 8). *Significant at P < 0.05 (Student’s t test)
The subspecies-specific indel and SSILP markers were distributed among the 12 chromosomes in the rice genome, with an average of 7.5 markers on each chromosome. Figure 2 shows the electrophoretic pattern of three indica varieties and three japonica varieties using an indel marker (R11M23) and a SSILP marker (RI05173). The difference between the indica and japonica type bands can be easily distinguished by electrophoresis. The differences in the length of the PCR products for remaining indel and SSILP markers for ‘Shennong265’ and ‘Luhui99’ are provided in Online Resource 1. We genotyped for all the 200 RILs using the subspecies-specific indel and SSILP markers. The indica-allelic frequency of RILs was calculated as the ratio of the number of indica type bands to the total number of subspecies-specific indel and SSILP markers for each lines. After calculating the indica-allelic frequency for all 200 RILs, we separated the RILs into three groups: pre-indica-type lines (frequency of indica-allelic markers >0.6), intermediate-type lines (0.4 < frequency of indica-allelic markers <0.6) and pre-japonica-type lines (frequency of indica-allelic markers <0.4). The frequency of indica-allelic markers in RILs showed a normal distribution (Fig. 3).

Electrophoretic pattern of two subspecies specific markers, R11M23 (indel) and RI05173 (SSILP) in ‘Shennong265’, ‘Luhui99’ and RIL1 to RIL18. All the 90 sets of subspecies specific markers was used for the genotyping of 200 RILs (figure not shown)

Frequency of indica-allelic markers in RIL lines
Table 2 summarizes the differences in the characteristics of rachis branches and vascular bundles in filial generations of the cross between indica and japonica. The number of primary branches, the number of secondary branches, the number of LVBs in the top second stem and the SVBs in the panicle neck showed a trend as follows: pre-indica < intermediate > pre-japonica. However, the number of LVBs, the ratio of the number of large vascular bundles at panicle neck to the number of small vascular bundles at panicle neck (LVB/SVB ratio), LVB ratio and the V/R ratio had the following trend: pre-japonica < intermediate < pre-indica. A correlation analysis between the frequency of indica-allelic markers and the characteristics of vascular bundles suggested that the frequency had a significantly positive correlation with the number of LVBs at the panicle neck, the LVB/SVB ratio, LVB ratio and the V/R ratio. However, the relationships observed among the pre-indica, intermediate and pre-japonica groups were not observed inside each group. The results for the vascular bundle-related traits in the RILs almost covered the full range of variations inside each group (Fig. 4).

Correlations between the frequency of indica-allelic markers and the vascular bundle characteristics. No. of LVB refers to the number of larger vascular bundles at panicle neck, LVB/SVB ratio refers to the ratio of the number of large vascular bundles at panicle neck to the number of small vascular bundles at panicle neck, LVB ratio refers to the ratio of the number of large vascular bundles at the panicle neck to the number of large vascular bundles at the top second internode of the stem, V/R ratio refers to the ratio of the number of large vascular bundles at the panicle neck to the number of primary branches
We divided the panicle into upper, middle and lower sections, and examined the vascular bundles. As indicated in Table 3, the middle section had more secondary branches than the upper and lower sections in the pre-indica, intermediate and pre-japonica groups. The number of LVBs showed the trend lower > middle > upper sections, whereas the middle section of the panicle had more SVBs than the upper and lower sections. The lower section had the highest V/R ratio of all three groups. For the primary branches, the middle section had a higher grain setting rate and greater 1000-grain weight than the upper and lower sections in pre-japonica and intermediate groups, whereas the upper sections had higher values for these two traits compared with middle and lower sections in pre-indica groups. The average number of grains per primary branch was greater in the lower sections than in the upper and middle sections of the panicle. For the secondary branches, the grain setting rate showed the trend upper > middle > lower sections in all three groups. The upper section had a greater 1000-grain weight than the middle and lower sections in the pre-japonica and intermediate groups, and the middle sections had the highest 1000-grain weight in the pre-indica group. The middle sections had the greatest number of grains per branch among the panicles (Table 3). In the upper and middle sections of panicle, the V/R ratio of the three groups showed a similar range of variation, although for the pre-japonica group the variation was slightly smaller than that of the other two groups. The obvious difference between the pre-japonica groups and other two groups was observed in the lower panicle section of panicle. The V/R ratio of the pre-japonica group was concentrated between 0.8 and 1.4, whereas the V/R ratio of the pre-indica and intermediate groups was highly variable, ranging from 0.8 to 2.4. Although there was no obvious relationship between V/R ratio and indica-allelic frequency, the V/R ratio dramatically increased when the indica-allelic frequency was higher than 0.5 in the lower section of the panicle (Fig. 5).

Correlations between the frequency of indica-allelic markers and the V/R ratio at different panicle positions. V/R ratio refers to the ratio of the number of large vascular bundles at the panicle neck to the number of primary branches
Discussion
The japonica and indica rice subspecies are derived from common wild rice (Oryza rufipogon Griff.). They have adapted to different ecological environments after long-term natural selection and artificial selection. Hence, a significant linkage disequilibrium phenomenon between morphological and genetic differentiation, which was caused by non-random combinations of different alleles, has been observed in the subspecies. The vascular bundles, which provide mechanical support for the plant and transport photosynthetic products into grains, are considerable different within the subspecies. The indica varieties have more LVBs at the panicle neck and higher V/R ratios compared with japonica varieties. The different vascular bundle characteristics in subspecies determined the different breeding strategies for indica and japonica. The generation of higher yielding indica varieties was usually achieved by increasing the numbers of the primary rachis branches and grains, whereas, breeding higher yielding japonica varieties required increasing the panicle number per unit area (Fukuyama et al. 1999). The use of indica × japonica crosses has become more common in rice breeding. Following developments in genetic analysis and gene cloning research, it was determined that there was no linkage between the major genes that controlled the characteristics of subspecies, and that these characteristics were randomly separated in crosses between indica and japonica (Chen et al. 2007; Mao et al. 2009; Yu et al. 2008). In this study, we use RILs derived from a cross between indica variety ‘Luhui 99’ and japonica variety ‘Shennong 265’ to investigate the relationship between the panicles characteristics and vascular bundles at different panicle positions in filial generations of an indica × japonica cross. The whole panicle as well as different panicle positions showed the following trend for the V/R ratio: pre-japonica < intermediate < pre-indica. The V/R ratio in the upper and middle sections were similar and both were lower than the ratio in the lower panicle section (Table 3). The V/R ratio of the pre-japonica groups was concentrated around 1.0, whereas the other two groups had a highly variable V/R ratio. Varieties with a high spikelet density varieties (including the parents line ‘Shennong265’ used in this study) in the northern China generally exhibit a mediocre grain setting rate in the lower section of panicle. The low V/R ratio in the lower section of panicle may be responsible for decreasing the grain setting ratio. However, a large variation in the V/R ratio was observed inside each group. In the pre-japonica group, the highest value of V/R ratio was 1.8, which was much higher than the average V/R ratio for japonica varieties (Fig. 4). These results indicated that we can optimize the combination of V/R ratio and the indica allele frequency to achieve higher yielding varieties by crossing subspecies and selection.
The setting rate of rice grains differs based on their panicle position. Grains located on the upper primary rachis branches show the highest grain setting rate and have the greatest 1000-grain weight. In contrast, there is a time lag for grains located on the secondary rachis branches of the lower primary rachis before the start of elongation, leading to the lowest grain setting rate (Ishimaru et al. 2003). In this study, we found that with an increase in the frequency of indica-allelic frequency and the number of LVBs, the differences in the grain setting rate and 1000-grain weight on primary and secondary branches among the three panicle sections became significant (Table 3). In japonica-type cultivars, each of the LVBs is connected to a primary rachis branch. However, in indica-type cultivars, the number of LVBs in the peduncle is much higher than that of primary branches. Therefore, some of the LVBs are directly connected to the secondary rachis branches (Fukushima and Akita 1997). We also observed that the increase in LVBs did not result in an increase in the number of primary branches. Instead, it led to an increase in the number of secondary branches (Table 3). The higher numbers of LVBs without an accompanying increase in the numbers of primary branches resulted in more LVBs that were directly connected to secondary rachis branches. As the primary branch grains have superior grain setting rates and 1000-grain weights compared with those of the secondary branch grains, a simultaneous increase in both the number of LVBs and number of primary branches, which would increase yields levels in crosses between indica and japonica, may be the goal for breeders and scientists.
References
Chen S, Xu Z, Chen W, Xu H, Liu H, Zhu C, Wang Y, Wang J (2007) The relation between the characters of Vascular Bundle and Panicle in the Filial Generation from Indica and Japonica Rice [J]. Acta Agric Boreali-Sin 5:004
Doyle J (1991) DNA protocols for plants. In: Hewitt GM, Johnston AWB, Young JPW (eds) Molecular techniques in taxonomy. NATO ASI Series, vol 57. Springer, Berlin, pp 283–293
Fukushima A, Akita S (1997) Varietal differences of the course and differentiation time of large vascular bundles in the rachis of rice. Jpn J Crop Sci 66(1):24–28
Fukuyama T, Sasahara H, Fukuta Y (1999) Variation of vascular bundle system corresponds to indica, Tropical-and Temperate-japonica differentiation of Asian Rice (Oryza sativa L.). Breed Sci 49:15–19
Garris AJ, Tai TH, Coburn J, Kresovich S, McCOUCH S (2005) Genetic structure and diversity in Oryza sativa L. Genetics 169:1631–1638
Ishimaru T, Matsuda T, Ohsugi R, Yamagishi T (2003) Morphological development of rice caryopses located at the different positions in a panicle from early to middle stage of grain filling. Funct Plant Biol 30:1139–1149
Ling Q, Cai J, Su Z (1982) The relationship between the number of macro-vascular bundles in culms and the panicle characters in rice plant and its application. J Jiangsu Agric Coll 3:7–16
Lu B-R, Cai X, Xin J (2009) Efficient indica and japonica rice identification based on the InDel molecular method: its implication in rice breeding and evolutionary research. Prog Nat Sci 19:1241–1252
Mao T, Xu H, Guo Y, Zhu C, Chen K, Wang J, Xu Z (2009) Comparative Study on Morphological differentiation and genetic differentiation in filial generation of cross between indica and japonica Rice [J]. Chin J Rice Sci 3:017
Shen Y-J, Jiang H, Jin J-P, Zhang Z-B, Xi B, He Y-Y, Wang G, Wang C, Qian L, Li X (2004) Development of genome-wide DNA polymorphism database for map-based cloning of rice genes. Plant Physiol 135:1198–1205
Sun J, Liu D, Wang J-Y, Ma D-R, Tang L, Gao H, Xu Z-J, Chen W-F (2012) The contribution of inter subspecific hybridization to the breeding of super-high-yielding japonica rice in northeast China. Theor Appl Genet 125:1149–1157
Wang X, Zhao X, Zhu J, Wu W (2006) Genome-wide investigation of intron length polymorphisms and their potential as molecular markers in rice (Oryza sativa L.). DNA Res 12:417–427
Yu T, Jiang W, Ham T, Chu S, Lestari P, Lee J, Kim M, Xu F, Han L, Dai L (2008) Comparison of grain quality traits between japonica rice cultivars from Korea and Yunnan Province of China. J Crop Sci Biotechnol 11:135–140
Zhang Z-H, Li P, Wang L-X, Tan C-J, Hu Z-L, Zhu Y-G, Zhu L-H (2002) Identification of quantitative trait loci (QTLs) for the characters of vascular bundles in peduncle related to indica-japonica differentiation in rice (Oryza sativa L.). Euphytica 128:279–284
Zhang P, Li J, Li X, Liu X, Zhao X, Lu Y (2011) Population structure and genetic diversity in a rice core collection (Oryza sativa L.) investigated with SSR markers. PLoS ONE 6:e27565
Zhao X, Yang L, Zheng Y, Xu Z, Wu W (2009) Subspecies-specific intron length polymorphism markers reveal clear genetic differentiation in common wild rice (Oryza rufipogon L.) in relation to the domestication of cultivated rice (O. sativa L.). J Genet Genomics 36:435–442
Acknowledgments
This work was supported by a grant from the National Natural Science Foundation of China (No. 31430062 and No. 31371587) and a class general financial grant from the China Postdoctoral Science Foundation (Grant Nos: 2014M560211 and 2015T80270, Postdoctoral No: 142541).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Tiansheng Liu and Wenjing Bi have contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Liu, T., Bi, W., Zhang, J. et al. Characterization of the relationship between vascular bundles features and indica-allelic frequency using a seventh filial generations of indica × japonica rice crosses. Euphytica 209, 739–748 (2016). https://doi.org/10.1007/s10681-016-1665-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-016-1665-7