
Abstract
Mammals can serve as an indicator of global climate change impacts on species’ distributions due to the wide range of ecological niches they utilize. Tropical Asia encompasses several biodiversity hotspots, is the largest reservoir of mammalian diversity on earth, and has already experienced the extinction of several mammal species either regionally or locally. Global climate change could become a significant driver of species extinction, either directly or synergistically with other factors, such as habitat loss, agricultural expansion, overexploitation, and land use change. Despite the variability of climatic regimes across tropical Asia, the potential impacts of climate change on continental-scale distributions of mammals have not been examined. To address this issue, we developed habitat suitability models for four threatened large mammals (Ursus thibetanus, Elephas maximus, Hoolock hoolock, and Panthera tigris tigris), across their entire distributions in Asia. We used presence-only distribution records and nine bioclimatic and environmental variables and built species-specific habitat suitability models using a maximum entropy algorithm (MaxEnt). We used a moderate and an extreme climate scenario (RCP6.0 and RCP8.5) and three time steps: current, 2050, and 2070. Our results suggest that changes in annual precipitation, annual mean temperature, precipitation, and temperature seasonality could reduce suitable habitat for these mammals and therefore increase their extinction risks. However, several patches of stable habitat are projected to persist through the late twenty-first century, and these climate change refugia areas can be managed as an important strategy for conservation of the mammal species and the maintenance of biodiversity in the face of ongoing climate change. In this context, we recommend the following steps for the conservation of global mammal populations: (i) define the spatial extent (local, regional, or continental scale) of the target mammals, (ii) identify and prioritize climate change refugial areas following ecological niche models or other methods based on biological data, and (iii) implement management actions by analyzing current management tools and the strategies required (e.g., habitat restoration or assisted migration for prioritized species) to achieve long-term conservation goals.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
There is growing observational evidence that global climate change is having a significant impact on species distributions, phenology, and vegetation dynamics, and could become a major cause for species extinction in concert with other global change drivers, such as agricultural expansion, overexploitation, habitat destruction and fragmentation, land use change, and invasive species (Thomas et al. 2004; Brook et al. 2008; Pacifici et al. 2015; Franklin et al. 2016). Terrestrial plant communities (such as forests, woodlands, shrublands, and grasslands) provide natural habitat for many animal species, and climate change-driven shifts in vegetation distribution could have cascading effects on the distribution of wildlife (Butt et al. 2015; Franklin et al. 2016). Mammals can serve as an indicator of climate change impacts on wildlife populations due to the wide range of ecological niches they exploit (Ceballos and Ehrlich 2002). Results of previous studies showed that extinction risk is greater in large mammals than small mammals (Cardillo et al. 2005). This is driven by a combination of extrinsic (environmental) factors and intrinsic species traits, such as small geographic range, low population density, slow life history, low reproductive rates, and large body size (Fisher and Owens 2004; Davidson et al. 2009). Therefore, future loss of large mammals due to climate change acting synergistically with other extinction drivers, such as habitat loss, land use change, poaching, and hunting, could be far more rapid than expected (Fig. 1) (Cardillo et al. 2005).

A schematic representation of different extinction drivers (climate change, habitat loss, deforestation, land use change, hunting, and poaching) for threatened large mammals in tropical Asia (for details see Table A1)
The population density of forest vertebrates largely depends on climatic factors, elevation, floristic composition, and net primary productivity (Galetti et al. 2009). However, the abundance of large mammals is primarily associated with both habitat fragmentation and hunting pressure (Michalski and Peres 2007; Peres and Palacios 2007). Despite several studies attempting to elucidate the effects of climate change on the distribution of large mammals (Levinsky et al. 2007; Alamgir et al. 2015; Pokharel et al. 2016), the interactions between anthropogenic disturbances and environmental variables remain poorly understood, especially in complex human-dominated biomes such as tropical forests (Galetti et al. 2009). Rondinini et al. (2011) developed habitat suitability models to assess large-scale distribution of terrestrial mammals based on their habitat relationships, with a focus on land cover, elevation, and hydrological features variables. Their study demonstrated the importance of fine-resolution distribution data for the development of global conservation strategies for mammals. Levinsky et al. (2007) evaluated the potential impact of climate change (excluding non-climatic factors such as land-use, biotic interactions, human interference, etc.) on the distributions and species richness of 120 native terrestrial non-volant European mammals and reported that 1 or 5–9% respectively, of European mammals risk extinction, while 32–46 or 70–78%, respectively, may be severely threatened (lose > 30% of their current distribution) under two IPCC future climatic scenarios (B1 and A2 scenarios). To comprehensively assess the impacts of climate change on the distribution of mammals, it is important that all contributing factors of vulnerability are taken into account to inform conservation actions effectively (Butt et al. 2016).
Of the four forest biomes (tropical, subtropical, temperate, and boreal), tropical forests are the richest biologically and contain the highest number of threatened species (Corlett and Lafrankie 1998; Brook et al. 2008; Butler and Laurance 2008). For instance, it has been estimated that mammal species are approximately seven times more numerous (http://www.iucnredlist.org/) within tropical biodiversity hotspots, compared with non-tropical hotspots (Myers et al. 2000). Southeast Asia encompasses four biodiversity hotspots and several of the most species-rich ecoregions (Olson and Dinerstein 1998; Myers et al. 2000). It has the highest relative rate of deforestation of any tropical region (Hansen et al. 2013), and could lose three quarters of its original forests and half of its biodiversity by 2100 (Sodhi et al. 2004). In Southeast Asia, 13 mammal species have already experienced 83% habitat loss (Ceballos and Ehrlich 2002), of great concern as this region holds the highest reservoirs of biodiversity on earth and is home to one of the highest concentrations of endemic species (Sodhi et al. 2004). South Asia represents approximately 10% of the world’s mammalian diversity, and includes 502 species belonging to 215 genera and 14 orders (Srinivasulu and Srinivasulu 2012). Approximately 32 mammal species have become extinct regionally or locally in South Asia due to habitat loss and fragmentation, and other extinction drivers such as land use change and climate change (Fig. 1) (Srinivasulu and Srinivasulu 2012). With an annual forest loss increment of 2101 km2 in tropical Asia (from 2000 to 2012) (Hansen et al. 2013), the region’s mammal populations are losing their natural habitats (Srinivasulu and Srinivasulu 2012). The extent of habitat loss in concert with global climate change is increasing the extinction risks of the large mammals (Sala et al. 2000; Thomas et al. 2004).
Habitat suitability models or species distribution models have been widely used in ecology to detect the climatically suitable habitat of mammals and inform conservation planning (Guisan and Zimmermann 2000; Elith and Leathwick 2009; Franklin 2010; Rondinini et al. 2011). Despite mammals being among the most intensively studied taxa, lack of detailed large-scale information on their potential distribution under future climate scenarios may hinder conservation efforts (Rondinini et al. 2011). In Asia, most studies of climate change impacts on mammal distributions focus on the local scale and do not consider the entire distribution ranges of the species (e.g., Loucks et al. 2010; Trisurat et al. 2012; Alamgir et al. 2015; Pokharel et al. 2016). Thus, continental-scale studies in Asia are limited, despite the conservation significance of mammal diversity in the region (Catullo et al. 2008). In this study, we have addressed this research gap by modeling the habitat suitability of four threatened large mammals under different climate scenarios across their entire distribution range.
The unique and globally endangered Bengal tiger (Panthera tigris tigris), Asian elephant (Elephas maximus), Western Hoolock Gibbon (Hoolock hoolock), and vulnerable Asiatic black bear (Ursus thibetanus) are naturally distributed in different Asian forest ecosystems (Table 1 and Table A1 for details), and their main threats are the combined effects of habitat loss, forest fragmentation, human interference, hunting, and global climate change (Loucks et al. 2010; Srinivasulu and Srinivasulu 2012; Alamgir et al. 2015; IUCN 2016). These species are already extinct locally in several countries, and the remaining disjunct populations are declining (Sala et al. 2000; Thomas et al. 2004; IUCN 2016). They play key roles in their forest ecosystems (e.g., grazing, predation, and seed dispersal), and are important for ecosystem function, such as in relation to food chains and food webs (Franklin et al. 2016). We focused on these threatened large mammals of Asian continent as these species are of highest conservation concern and typically targeted by international conventions (Secretariat of the CBD 2010).
The aim of the paper was to assess the four species’ vulnerability to global climate change and examined the importance of mean and seasonal climate, topography, land use/land cover, and maximum green vegetation fraction of landscape variables for habitat suitability for the mammals under different climate scenarios and for different time periods. This allowed us to identify the potential extinction risks for each species, with implications for conservation planning. Finally, global recommendations are presented to provide adaptive management and conservation strategies for the studied species, and suggestions for how these methods and analyses could be applied to other tropical regions (i.e., in Africa and Americas), using different mammal species and forest types.
2 Materials and methods
2.1 Ecology of study species
The four mammals occupy a variety of forested habitats, grasslands, cultivated and secondary forests across Asia (Fig. 2 and Table A1 for details). They play important ecological roles, and help maintain ecosystem health and diversity (Franklin et al. 2016). For instance, the Asian elephant and Bengal tiger are considered keystone species, and their presence in the forests is an indicator of ecosystem well-being. The Asiatic black bear, Asian elephant, and Western hoolock gibbon rely on tree flowering and fruiting, and also on the shoots, forbs, and leaves of many plants (Corlett and Lafrankie 1998; IUCN 2016). Bengal tiger is at the apex of the food chain and maintains the balance between prey herbivores and the vegetation upon which they feed (IUCN 2016). Although no rigorous population estimates exist for these mammals, recent studies suggest that the current populations of these species are declining (Table 1), which may lead to local or regional extinction in the near future (IUCN 2016). Temperature and rainfall variations (e.g., drought, heavy rainfall) have the potential to affect the phenology of tropical and subtropical Asian forests through significant perturbations to the timing of fruit, seed, and flower availability, with cascading effects on the distribution and population dynamics of large mammals (Parmesan 2006).

The current distribution ranges of the threatened large mammals in different landscapes across Asian continent: (1) Asiatic black bear (Ursus thibetanus), (2) Asian elephant (Elephas maximus), (3) Western hoolock gibbon (Hoolock hoolock), and (4) Bengal tiger (Panthera tigris tigris) (for details see Table A1) (Source: IUCN 2014)
2.2 Species distribution and environmental data
We obtained occurrence records for the four mammals from the terrestrial mammals’ data of the IUCN Red List (IUCN 2014), and the Global Biodiversity Information Facility (GBIF, <http://www.gbif.org/>). We compiled the datasets and cleaned the occurrence records by removing overlapping locations or spurious points after reviewing the literature for each species (e.g., Khan 2008). Finally, we used 398, 160, 52, and 75 records for Asiatic black bear, Asian elephant, Western hoolock gibbon, and Bengal tiger, respectively, to model the distributions (Fig. 2).
The direct effects of human activities such as large-scale industrial logging, habitat destruction and fragmentation, illegal logging, and overexploitation are the primary contemporary drivers of tropical forest biodiversity loss along with the indirect effects of anthropogenic climate change (Hansen et al. 2013; Pacifici et al. 2015). Therefore, it is important to include habitat destruction variables along with climate variables in the modeling. Land use (description of land in terms of its socio-economic purpose, e.g., agriculture, forestry, residential, etc.) and land cover (physical and biological cover of earth’s surface, e.g., forests, agricultural areas, wetlands, water bodies, etc.) variables are expected to change over shorter timescales. For instance, Sohl (2014) used land use/land cover (LULC) projections data produced for the conterminous USA, with annual LULC maps from 1992 to 2100 for four Intergovernmental Panel on Climate Change (IPCC) Special Report on Emissions Scenarios (SRES) for a bird distribution modeling. Although reliable projections of LULC are not available for tropical Asia, future values of climate change are predicted for the next several decades by general circulation models (GCM) (Hijmans et al. 2005). Stanton et al. (2012) suggested that combining the important static variables in the model along with the dynamic climate variables showed better result than excluding them (static variables). In this study, we modeled the distribution of threatened large mammals using climatic variables only, and combining the unchanging or static environmental variables with the projected climate variables.
We initially considered 19 bioclimatic variables (11 temperature and 8 precipitation metrics) from the WorldClim database (Hijmans et al. 2005). All the bioclimatic layers were 1 km resolution. In addition, we also included three other static environmental variables: elevation (ELV) (Hijmans et al. 2005), land use/land cover (LULC) (Arino et al. 2012), and annual maximum green vegetation fraction (MGVF) (Broxton et al. 2014) in a different model to compare the model variations. As there is no robust dataset on LULC for tropical Asia, we included LULC variable from the default Global Land Cover Map for 2009 data (300 m resolution; 21 LULC classes) (Arino et al. 2012). The 1 km MODIS-based MGVF data are based on 12 years (2001–2012) of normalized difference vegetation index (NDVI) data (Broxton et al. 2014). Vegetation cover influences the land-atmosphere exchanges of water, energy, momentum, and carbon, and is widely used in global models along with many other applications such as studies of land cover change (Broxton et al. 2014; Dai et al. 2003). MGVF describes the vegetation abundance, i.e., green vegetation fraction (vs. non vegetated area) for each land cover class for each year (Broxton et al. 2014). We assume including the LULC and MGVF variables in the model may capture the deforestation scenarios in the study region.
We selected two of the four representative concentration pathway (RCP) scenarios: RCP6.0, a stabilization-without-overshoot pathway to 6 W m−2 by 2100, corresponds to a peak in greenhouse gases by 2060 and RCP8.5, a rising radiative forcing pathway resulting in 8.5 W m−2 by 2100, which reflects high levels of energy demand and greenhouse gas emissions without climate change policies (Moss et al. 2010). We used data for current conditions (the average for 1950–2000) and projected climate data for the time periods of 2050 (the average for 2041–2060) and 2070 (the average for 2061–2080) from the HadGEM2-ES global circulation model (Hijmans et al. 2005).
2.3 Exploratory data analysis
As this study aimed to understand which variables were driving distributions, we applied Spearman’s rank correlation to test for collinearity between variables at each level. Dormann et al. (2013) suggest that a threshold of 0.7 is the most common in ecology (i.e., if a pair of variables has a correlation coefficient > 0.7, then they should be considered proxies of one another). We applied this general rule and removed 13 of the variables (Table A2). We used nine predictor variables for habitat suitability modeling: annual mean temperature; mean diurnal range; isothermality; temperature seasonality; annual precipitation; precipitation seasonality; elevation; land use/land cover; and annual maximum green vegetation fraction. The elevation variable is important because the mammals occupy different topographic features of the landscapes. We used the land use/land cover and maximum green vegetation fraction (the average data of 2001–2012) variables to predict current distributions of the species. However, there are no models for the future estimates of these two variables for each climate scenarios; we projected the distributions to 2050 and 2070, with and without these ‘fixed’ variables.
2.4 Habitat suitability models
We built the models of the relationship between each species’ occurrences and the climatic conditions using the niche modeling software MaxEnt version 3.3.3 k (Phillips et al. 2006; Phillips and Dudík 2008). The MaxEnt (maximum entropy) algorithm has been shown to perform well, even with low sample sizes, and has the advantages over other species distribution models in that it is designed to operate without data on true absences (Phillips et al. 2004; Elith et al. 2006; Phillips and Dudík 2008). To improve the robustness of the model extrapolation, we created a bias file layer and defined MaxEnt Background selection by limiting the sampling locations from where they were selected (Phillips et al. 2009). This limits the background point to areas that we assume were surveyed for the mammals, and provides MaxEnt with a background file with the same bias as the presence locations (Phillips et al. 2009). We generated the MaxEnt models from a cross-validation on the data and setting the default background points to 10,000, regularization multiplier to 1, and maximum iterations to 500. We also used the jackknife test to measure the variable importance to the models. We validated the models using the threshold-independent area under the receiver operating curve (AUC) metric (value ranges between 0 and 1.0) that describes the fit of the model to the test data and gives strong model discrimination ability for predicting changes in species distribution under future climate scenarios (Phillips et al. 2006; Phillips and Dudík 2008). An AUC value greater than 0.75 indicates that the model has good discrimination ability in accurately identifying the potential distribution of a species (Elith et al. 2011). We used the ten percentile training presence logistic threshold to define the minimum probability of suitable habitat (Phillips et al. 2006) and account for sampling error: we defined suitable habitat to include 90% of the data used to develop the model.
We generated the MaxEnt models for the threatened mammals in two different settings: (a) incorporating the bioclimatic variables only as predictor variables, and (b) including the three other environmental variables (i.e., elevation, land use/land cover, and annual maximum green vegetation fraction) along with bioclimatic variables. As the mammals have wide distributions across different ecosystems in Asia, variations in topographic heterogeneity, deforestation scenarios, and land use change are likely to have a significant influence on the distributions, even for a given set of climatic conditions (Hansen et al. 2013; Wilson et al. 2013).
3 Results
Overall, the MaxEnt models performed well in predicting habitat suitability for the threatened mammals across Asia. Mean AUC values across all models (for models that includes climatic variables only: 0.80 ± 0.05; and models including all variables: 0.82 ± 0.04) fell within the range of good performance (Table A3).
There were some similarities found in the relative contributions of the predictor variables that influence the spatial distribution of the large mammals in the study region (Fig. 3). The key bioclimatic predictor variable for the Asiatic black bear, Asian elephant, and Bengal tiger was annual precipitation (BIO12) for both models, i.e., models with climatic variables only and with all variables. In contrast, the key variable for the Western hoolock gibbon was mean diurnal range (BIO2) (Fig. 3). Precipitation and temperature seasonality coupled with annual mean temperature and elevation features may also influence the distribution of Asiatic black bear as they were important variables in the model. The important contribution of temperature seasonality along with annual mean temperature, maximum green vegetation fraction, and elevation or topographical variables in the model indicates that variation in these variables may influence the distribution of Asian elephant in the region (Fig. 3). The relative contribution of annual precipitation, precipitation seasonality, elevation, and isothermality were important in both models for the Western hoolock gibbon (Fig. 3). Annual mean temperature, land use/land cover change, and precipitation seasonality variables were also important for the distribution of Bengal tiger.

Summary of the bioclimatic and environmental variables used in the habitat suitability models and their percent contribution to each model: a models with only bioclimatic variables as predictors and b models with all variables. Annual precipitation (BIO12) was the most regulatory variable found in both models that influence the habitat suitability of all the mammals
Both MaxEnt models (for ‘climate variables’ and ‘all variables’) revealed a consistent pattern of predicted habitat suitability for all four mammals, i.e., range contraction of their natural habitat (Figs. 4, 5, 6, and 7 and Fig. A1–4). Our models predicted that climatically suitable habitat conditions for the threatened large mammals will decline across Asia. However, Western hoolock gibbon is likely to gain climatically suitable habitat outside of its current natural habitats. The projected impacts of climate change on the habitats of Asiatic black bear are severe under both RCP scenarios, with a 38% decline by 2070 under RCP8.5 for the model with climate variables only, and 40% for the model with all variables (Fig. 8). The model with climate variables only indicates a decline of up to 59% of suitable climate space for the Asian elephant by 2070 under RCP8.5. However, the model with all variables indicates a relatively low percentage (5%) of decline in habitat suitability for Asian elephant. This may be due to the wide variety of ecosystems (grasslands, tropical evergreen, semi-evergreen, dry and moist deciduous, dry thorn forests) the Asian elephant occupies, with an elevation ranging from sea level to 3000 m across tropical Asia (Table A1). The habitat suitability of Bengal tigers will decline up to 14% across Asia by 2070 under RCP8.5 indicated by the model with climatic variables. However, the model with all variables indicates that Bengal tigers may gain some climate space (1%) by 2070 under RCP8.5, with relatively low declines in habitat suitability by 2050 and 2070 under RCP6.0. In contrast to the other three mammals, Wester hoolock gibbon will gain suitable climatic conditions in all climatic scenarios for both models: up to 12% by 2070 under RCP8.5 for the model with climatic variables and up to 20% for the other model (Fig. 8). All models revealed the likely range contraction of climatically suitable natural habitats of the threatened large mammals. However, climatically suitable range expansion outside of their natural habitats may provide potential for species migration.

The potential habitat suitability for Asiatic black bear (Ursus thibetanus) across its entire distributions in Asia: a species occurrences across Asia and mapped current habitat suitability for bear and b–e the four projected habitat suitability for bear in different scenarios. The models indicate that the likely habitat suitability for bear will decline under both climate scenarios (RCP6.0 and RCP8.5) by 2050 and 2070. Changes in annual precipitation, precipitation and temperature seasonality, and annual mean temperature may influence the distribution of Asiatic black bear

The predicted habitat suitability for Asian elephant (Elephas maximus) across its entire distributions in Asia: a species occurrences across Asia and mapped current habitat suitability for elephant and b–e the four projected habitat suitability for elephant in different scenarios. The models indicate that the Asian elephant are likely to face extinction risk under both climate scenarios (RCP6.0 and RCP8.5) by 2050 and 2070. The key bioclimatic variables that influence the distribution of Asian elephant are annual precipitation, temperature seasonality, and annual mean temperature

The projected habitat suitability for Western hoolock gibbon (Hoolock hoolock) across Asia: a species occurrences across Asia and mapped current habitat suitability for gibbon and b–e the four projected habitat suitability for gibbon in different scenarios. The models indicate that the habitat suitability of gibbon is likely to contract under both climate scenarios (RCP6.0 and RCP8.5) by 2050 and 2070. However, there is a shift in the distribution for gibbon outside of its native ranges where assisted migration of the species can reduce the species extinction risks. The key bioclimatic variables that influence the distribution of gibbon are mean diurnal range, annual precipitation, isothermality, and precipitation seasonality

The potential habitat suitability for Bengal tiger (Panthera tigris tigris) across Asia: a species occurrences across Asia and mapped current habitat suitability for tiger and b–e the four projected habitat suitability for tiger in different scenarios. The models indicate that the habitat suitability of tiger is likely to contract under both climate scenarios (RCP6.0 and RCP8.5) by 2050 and 2070. However, there is a shift in the distribution for tiger outside of its native ranges. The key bioclimatic variables that influence the distribution of tiger are annual precipitation, annual mean temperature, and precipitation seasonality

The climatically suitable habitat conditions for the threatened mammals by 2050 and 2070 under RCP6.0 and RCP8.5: a models with bioclimatic variables only and b models with all variables. Results of both models suggest that the habitat suitability of Asiatic black bear (Ursus thibetanus), Asian elephant (Elephas maximus), and Bengal tiger (Panthera tigris tigris) will decline across Asia except Western hoolock gibbon (Hoolock hoolock) which will likely to gain climatically suitable habitat outside of its natural habitats
4 Discussion
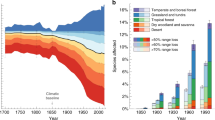
Our results suggest that global climate change could severely impact the distributions of threatened large mammals across Asia, with contraction and shifts in climatically suitable habitat conditions. The projected changes in annual precipitation and annual mean temperature and changes in seasonal climate (precipitation and temperature regimes) could be the key regulatory factors for the mammals’ distributions in tropical Asia. In addition to increasing climate stress, land use change and other anthropogenic factors may drive the distribution of mammals and lead them to become extinct, either locally or regionally in Asia (Thomas et al. 2004; Hansen et al. 2013; Franklin et al. 2016). Visconti et al. (2016) found that the extinction risks of terrestrial carnivore species increases for 8–23% depending on assumptions about species responses to climate change, which is consistent with our findings.
4.1 The influences of climatic variability on mammals’ distribution
Climatic regimes in the Asian tropics are highly diverse and can be divided into three zones: the marginal tropics (mean temperature of the coldest month < 18 °C; low seasonal temperatures may limit the growth of plants); the monsoon tropics (mean rainfall of the driest month < 50 mm; water availability limits plant growth); and the aseasonal tropics (temperature and water supply are adequate for growth year round) (Corlett and Lafrankie 1998). Climate change is already impacting vegetation in this region through an influence on phenology (Corlett and Lafrankie 1998). The Asiatic black bear, Asian elephant, and Western hoolock gibbon occupy a large variety of ecosystems across Asia and rely on periodically available plant resources for their survival (Corlett and Lafrankie 1998; IUCN 2016).
The habitat of the Asiatic black bear includes both broad-leaved and coniferous forests and they mostly occur in the marginal and monsoon tropics (Corlett and Lafrankie 1998). The bear moves to different habitats and elevations seasonally for tracking changes in food abundance and relies on fruit at different times of the year (Izumiyama and Shiraishi 2004). Seasonal low temperatures drive annual fruiting phenology in the Indo-Malayan subtropics and variations in temperature and precipitation seasonality, coupled with annual precipitation, annual mean temperature, and elevation features, are important for Asiatic black bear distribution (Corlett 1998). Asian elephant browse a wide variety of ecosystems and in South India, 70% of their diet comes from dry season browsing, while in the wet season grasses make up about 55% of their diet (Sukumar 1992). Although the annual diet of Asian elephant is dominated by grass (84%), dry deciduous forest species also contribute a considerable amount (Baskaran 1998). Variation in seasonal temperature, annual mean temperature, and annual precipitation such as drought or heavy rains can lead the plants to flower and fruit drop, with therefore potentially significant effects on elephant populations (Gunarathne and Perera 2014). As the elephants mostly rely on crops and grass rather than wild fruits, other factors such as roads, poaching, and conflicts with humans may also be important for their distribution.
The Western hoolock gibbon is a frugivorous species found in the tropical evergreen, semi-evergreen, mixed deciduous, and subtropical broad leaf forests of India, Bangladesh, and Myanmar (IUCN 2016). Ting et al. (2008) revealed that fruit production in tropical regions was related most strongly to evapotranspiration. Therefore, seasonal changes in climate (e.g., mean diurnal range, annual precipitation, precipitation seasonality, isothermality) leading to variations in evapotranspiration could affect fruiting phenology, with potential effects for the gibbon species (Butt et al. 2015).
The loss of highly suitable habitat for Bengal tiger is associated with flooding resulting from heavy rainfall in Nepal’s Chitwan district (Carter et al. 2013), and is consistent with our modeling results. Increasing annual mean temperatures and variation in precipitation seasonality, such as drought or heavy rain, are likely to affect the phenology of tropical evergreen, dry deciduous, moist deciduous, mangrove, subtropical, temperate uplands, and alluvial grasslands across India, Bangladesh, Nepal, and Bhutan. This may affect the population of prey herbivores and thereby the Bengal tiger, with disruption to ecosystem food webs.
4.2 Modeling approach and limitations
Our modeled responses to global changes may be overoptimistic for the studied mammals in tropical Asia because we did not account for all threats to mammals, especially hunting, poaching, and human-wildlife conflicts which are major threats to the species considered here. In addition, we used the static LULC and MGVF variables as reliable projections are not available for tropical Asia. However, assessing the biodiversity consequences of climate change is complicated due to the uncertainty of the degree, rate, and nature of projected climate change (IPCC 2007), and the interaction of climate change effects with biotic factors (competition, trophic relationships, dispersal abilities, etc.) and stressors (land use, habitat fragmentation, etc.) (Wiegand et al. 2005). In contrast, predicting spatially explicit maps for LULC change is difficult as deforestation may spread unexpectedly to areas that are currently pristine, and forests may be allowed to regrow in previously cleared areas (Asner et al. 2010). We acknowledge that our correlative approach of modeling based on dynamic bioclimatic and static LULC variables for the studied species are not a representative subset of all mammals in tropical Asia. However, it can form a basis for the mammal studies in tropical Asian region. Although human land use remains the main driver of present day species extinction and habitat loss (Hoffmann et al. 2010), our models suggest that climate change is projected to become equally or more important in the coming decades for mammals distribution in tropical Asia (Fig. 5). It is difficult to rely on a single scientific approach for the conservation policy and management of the threatened mammals in tropical Asia given the underlying assumptions of that approach are under debate. Additional research is needed to assess the optimum combination of covariates (e.g., LULC change, climate change, biotic factors, and other variables such as hunting, poaching, and human-wildlife conflicts) using different methods (rather than relying on one single method) and how covariate choice impacts results.
4.3 Extinction risks of the mammals
The current population trend of these threatened large mammals is negative, and there are multiple pathways (e.g., habitat loss, fragmentation, human interference, poaching, hunting, and global climate change) to extinctions for these species (Davidson et al. 2009; IUCN 2016). Although no rigorous population estimates exist for Asiatic black bear for the whole continent, a study in Bangladesh suggests that the distributions of this species is highly fragmented/patchy and it is ‘Critically Endangered’ according to IUCN (2000) guidelines (Garshelis et al. 2008; Islam et al. 2010). Sport hunting and trading of Asiatic black bears in Japan, South Korea, China, Vietnam, and several other countries is increasing the extinction risk for this species (IUCN 2016). One estimate for the global population size of Asian elephant was 41–52,000, of which more than 50% occurred in India (Choudhury et al. 2008). However, a more recent study reported a significant decrease in the population of Asian elephant in India (Puyravaud et al. 2017), and Alamgir et al. (2015) reported that there is likely to be a 38% loss in suitable habitat in Bangladesh for the remaining Asian elephant populations (300–350) in the near future.
The scenarios for Western hoolock gibbon populations (approximately 300) are extreme in Bangladesh, with 100% habitat loss and therefore possible extinction by 2070 under RCP8.5 (Alamgir et al. 2015). Sanderson et al. (2010) reported a 41% decline in population and occupied area for Bengal tiger in India. It has been estimated that tiger habitat and tiger populations in the Sundarbans are likely to reach a critical threshold at sea level rise between 24 and 28 cm above the year 2000 baseline; beyond 28 cm, the remaining tiger habitat in Bangladesh’s Sundarbans would decline by 96%, and the number of breeding individuals would be reduced to fewer than 20 (Loucks et al. 2010). Horev et al. (2012) reported that the entire population of Bengal tigers in India is likely to go extinct in 21.5 years as six tigers are being poached annually. The number of extinct mammal species in South Asian countries is greatest for Bangladesh (11 species), followed by Afghanistan (7), Pakistan (5), Bhutan (3), Nepal (3), India (2), and Sri Lanka (1) (Srinivasulu and Srinivasulu 2012). Extinction rates are usually high in large mammals due to the interaction between small geographic ranges and slow reproductive rates (Cardillo et al. 2005; Davidson et al. 2009), and our results also suggest that there will be declines in the suitable habitat for the threatened large mammals of Asia, which may lead to local or regional extinction with the current rates of population decrease.
4.4 Implications for conservation planning
The habitat of the threatened large mammals occurs in a variety of land management regimes (e.g., protected areas, reserved forests, multiple land-use areas) across tropical Asia (Carter et al. 2013; IUCN 2016). The habitat preferences differ among mammal species. For instance, Bengal tigers prefer habitats with more grasslands and higher landscape connectivity in Chitwan district of Nepal (Carter et al. 2013), whereas Asian elephants prefer areas close to a permanent source of fresh water as they need 80–200 l of water a day for drinking and bathing (Shoshani and Eisenberg 1982). Results from previous studies suggest that the area of highly suitable habitat for Bengal tigers has decreased inside the park over 20 years in the Chitwan district of Nepal, while outside the park habitat suitability increased, especially from 1999 to 2009 (Carter et al. 2013). The distribution range of all these large mammals across tropical Asia is not limited to protected areas (PAs) and areas outside PAs are subject to development projects that may be a problem for the conservation of these mammals (Sathyakumar 2006).
The findings of our study inform the suitability of habitats for these threatened large mammals in different climatic scenarios inside and outside the PAs and can inform conservation planning. Our models predict more than 50% of climatically suitable habitat conditions for all species will occur outside of their natural habitats (Fig. 4, 5, 6, and 7 and Fig. A1–4). Different efforts to reduce habitat degradation outside PAs, and to increase the number and/or area of PAs considering the habitat range of these species, would be highly beneficial for species conservation (IUCN 2016). For instance, China, India, and several other countries have already established a number of PAs within the range of Asiatic black bears (Chape et al. 2003). In addition, establishing travel corridors between existing PAs could also be an option to account for the projected shifts in the distributions of mammal habitat under global climate change, and facilitate species’ movement (Chape et al. 2003). The future distributions of the Asian elephant and Bengal tiger depend upon the conservation of large areas of suitable habitat by securing additional habitat, as their distribution is now highly clumped in disjunct areas (Walston et al. 2010; Puyravaud et al. 2017).
Habitat suitability models that predict the impact of climate change on species distributions frequently contrast scenarios of unconstrained and no dispersal with the caveat that, in reality, most species will show a range of dispersal distances which fall between these two assumptions (Broennimann et al. 2006). Therefore, the importance of dispersal in enabling species to keep pace with changing climates could be a useful tool for conservation planning. However, the quantitative descriptions of dispersal for the studied species were not included in the models due to the lack of robust data. The shifts in the distribution of the threatened large mammals’ suitable climate space revealed in this study could be used to inform assisted migration as a management strategy for aiding species in reaching newly suitable locations as climate changes (Hällfors et al. 2016). However, poaching, hunting, and human-wildlife conflicts also increase the extinction risks of mammals. Although these variables were not included in our models, the conservation needs of the threatened large mammals may vary depending on the intensity of these variables. For instance, if poaching is worse in some areas, then different conservation measures would be needed, such as improved legislation and law enforcement regarding poaching, hunting, and human-wildlife conflicts. Monitoring of conservation interventions as part of adaptive management, and reliable estimation of population size and trends, are also required for the success of mammal conservation (IUCN 2016). In addition, increasing connectivity of suitable habitats between PAs that are too small to maintain viable populations in isolation, as well as conservation outside PAs, would be beneficial for mammal conservation in tropical Asia (Trisurat et al. 2012).
4.5 Global strategy recommendations for the management of mammal populations in a changing climate
Our study predicted climatically suitable habitat conditions for all species both within and outside of their natural habitats across Asia (Fig. 4, 5, 6, and 7 and Fig. A1–4). Analysis of habitat maps from current to 2070 indicate that several patches of stable habitat are projected to persist through the late twenty-first century for all species, suggesting that these areas could function as climate change refugia (locations where taxa survive periods of regionally adverse climate) in the future. However, habitat loss leading to fragmentation in tropical Asia, as well as increasing pressure from human activities, will present conservation challenges for large mammals (e.g., Li et al. 2016). Managing climate change refugia can be an important strategy for the conservation of mammal species and maintaining biodiversity under ongoing climate change (Gavin et al. 2014; Morelli et al. 2016). Our models suggest that refugia and nearby non-refugia habitats could be reconnected (by protecting or enhancing connectivity corridors and restoring or protecting nearby non-refugial habitats) to improve long-term access to refuges (Morelli et al. 2016). Despite the conservation appeal of buffer regions, climate change refugia are not necessarily long-term solutions (Magris et al. 2014). They function best when coupled with contingency plans, such as tracking geographic shifts in refugial habitats to keep pace with climate change, or maintaining captive propagation, or zoos for future re-introduction (Morelli et al. 2016). However, adaptive management strategies incorporating climate change refugia might be worthwhile for highly valued targets, such as iconic or endemic large mammals (e.g., Asian elephant, Bengal tiger, Western hoolock gibbon, and Asiatic black bear). Challenges currently include questions about the scale at which climate change refugia should be identified and managed, uncertainty about the duration of their effectiveness, and lack of clarity over how to incorporate the requirements of multiple mammal species or other resources that will respond to climate change in different ways. Effective climate adaptation strategies must encompass targets that are spatially diverse, temporally dynamic, and multi-faceted (Morelli et al. 2016). We recommend the following steps for the management of refugia and conservation of the threatened global mammal populations: (i) define the spatial extent (local, regional, or continental scale) of the target mammals, (ii) identify and prioritize climate change refugial areas following ecological niche models or other methods based on biological data, and (iii) implement management actions by analyzing current management tools and the strategies required (e.g., habitat restoration or assisted migration for prioritized species) to achieve long-term conservation goals (Morelli et al. 2016). Future research should focus on exploring the opportunities and challenges for effective implementation of climate change refugia.
5 Conclusions
Most studies on mammal habitat suitability in Asian countries focus on the local scale and do not consider the entire distribution ranges of the species. This can hinder conservation efforts. However, habitat suitability models can assess the vulnerability of threatened Asian mammals with patchy distributions in different ecosystems and in areas that have undergone extensive disturbance. Our models revealed the likely range contraction of climatically suitable natural habitats of the threated large mammals across Asia. However, the projected suitable conditions both within and outside of the natural habitats of the mammals could function as climate change refugia in the future. The findings of our models can inform conservation planning for these threatened large mammals under global climate change. Managing climate change refugia can be an important adaptation strategy for the conservation of global mammal populations under ongoing climate change. We recommend that proper habitat management of the existing protected areas, and increasing the number and connectivity of protected areas, could reduce the extinction risks of these threatened mammals. Future research should focus on the spatial prediction of these mammals within and outside of protected areas, looking for previously unrecorded populations, prey density, poaching incidents, dispersal capabilities of species, and conflicts with humans, updating models, and planning for conservation.
References
Alamgir M, Mukul SA, Turton SM (2015) Modelling spatial distribution of critically endangered Asian elephant and Hoolock gibbon in Bangladesh forest ecosystems under a changing climate. Appl Geogr 60:10–19
Arino O, Ramos Perez J, Kalogirou V, Bontemps S, Defourny P, Van Bogaert E (2012) Global land cover map for 2009 (GlobCover 2009). European Space Agency & Université Catholique de Louvain
Asner GP, Loarie SR, Heyder U (2010) Combined effects of climate and land-use change on the future of humid tropical forests. Conserv Lett 3:395–403
Baskaran N (1998) Ranging and resource utilization by Asian elephant (Elephas maximus Linnaeus) in Nilgiri biosphere reserve, South India. Ph. D. thesis, Bharathidasan University, Tamil Nadu
Broennimann O, Thuiller W, Hughes G, Midgley GF, Alkemade JR, Guisan A (2006) Do geographic distribution, niche property and life form explain plants' vulnerability to global change? Glob Chang Biol 12:1079–1093
Brook BW, Sodhi NS, Bradshaw CJ (2008) Synergies among extinction drivers under global change. Trends Ecol Evol 23:453–460
Broxton PD, Zeng X, Scheftic W, Troch PA (2014) A MODIS-based global 1-km maximum green vegetation fraction dataset. J Appl Meteorol Climatol 53:1996–2004
Butler RA, Laurance WF (2008) New strategies for conserving tropical forests. Trends Ecol Evol 23:469–472
Butt N, Seabrook L, Maron M, Law BS, Dawson TP, Syktus J, McAlpine CA (2015) Cascading effects of climate extremes on vertebrate fauna through changes to low-latitude tree flowering and fruiting phenology. Glob Chang Biol 21:3267–3277
Butt N, Whiting S, Dethmers K (2016) Identifying future sea turtle conservation areas under climate change. Biol Conserv 204:189–196
Cardillo M, Mace GM, Jones KE, Bielby J, Bininda-Emonds OR, Sechrest W, Orme CDL, Purvis A (2005) Multiple causes of high extinction risk in large mammal species. Science 309:1239–1241
Carter NH, Gurung B, Vina A, Campa H III, Karki JB, Liu J (2013) Assessing spatiotemporal changes in tiger habitat across different land management regimes. Ecosphere 4:1–19
Catullo G, Masi M, Falcucci A, Maiorano L, Rondinini C, Boitani L (2008) A gap analysis of southeast Asian mammals based on habitat suitability models. Biol Conserv 141:2730–2744
Ceballos G, Ehrlich PR (2002) Mammal population losses and the extinction crisis. Science 296:904–907
Chape S, Blyth S, Fish L, Fox P, Spalding M (2003) 2003 United Nations list of protected areas. IUCN, Cambridge (RU). UNEP, Geneva (Suiza)
Choudhury A, Lahiri Choudhury DK, Desai A, Duckworth JW, Easa PS, Johnsingh AJT, Fernando P, Hedges S, Gunawardena M, Kurt F, Karanth U, Lister A, Menon V, Riddle H, Rübel A, Wikramanayake E (IUCN SSC Asian Elephant Specialist Group) (2008) Elephas maximus. The IUCN Red List of Threatened Species 2008:e.T7140A12828813. https://doi.org/10.2305/IUCN.UK.2008.RLTS.T7140A12828813.en. Downloaded on 28 September 2016
Corlett RT (1998) Frugivory and seed dispersal by vertebrates in the oriental (Indomalayan) region. Biol Rev Camb Philos Soc 73:413–448
Corlett RT, Lafrankie JV Jr (1998) Potential impacts of climate change on tropical asian foreststhrough an influence on phenology. Climatic Change 39:439–453
Dai Y, Zeng X, Dickinson RE, Baker I, Bonan GB, Bosilovich MG, Denning AS, Dirmeyer PA, Houser PR, Niu G (2003) The common land model. Bull Am Meteorol Soc 84:1013–1023
Davidson AD, Hamilton MJ, Boyer AG, Brown JH, Ceballos G (2009) Multiple ecological pathways to extinction in mammals. Proc Natl Acad Sci 106:10702–10705
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, Marquéz JRG, Gruber B, Lafourcade B, Leitão PJ (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46
Elith J, Leathwick JR (2009) Species distribution models: ecological explanation and prediction across space and time. Annu Rev Ecol Evol Syst 40:677–697
Elith J, Graham CH, Anderson RP, Dudík M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57
Fisher DO, Owens IP (2004) The comparative method in conservation biology. Trends Ecol Evol 19:391–398
Franklin J (2010) Mapping species distributions: spatial inference and prediction. Cambridge University Press, Cambridge
Franklin J, Serra-Diaz JM, Syphard AD, Regan HM (2016) Global change and terrestrial plant community dynamics. Proc Natl Acad Sci 113:3725–3734
Galetti M, Giacomini HC, Bueno RS, Bernardo CS, Marques RM, Bovendorp RS, Steffler CE, Rubim P, Gobbo SK, Donatti CI (2009) Priority areas for the conservation of Atlantic forest large mammals. Biol Conserv 142:1229–1241
Garshelis DL, Steinmetz R (IUCN SSC Bear Specialist Group) (2008) Ursus thibetanus. The IUCN red list of threatened species 2008: e.T22824A9391633. https://doi.org/10.2305/IUCN.UK.2008.RLTS.T22824A9391633.en. Downloaded on 28 September 2016
Gavin DG, Fitzpatrick MC, Gugger PF, Heath KD, Rodríguez-Sánchez F, Dobrowski SZ, Hampe A, Hu FS, Ashcroft MB, Bartlein PJ, Blois JL, Carstens BC, Davis EB, de Lafontaine G, Edwards ME, Fernandez M, Henne PD, Herring EM, Holden ZA, W-s K, Liu J, Magri D, Matzke NJ, McGlone MS, Saltré F, Stigall AL, Tsai Y-HE, Williams JW (2014) Climate refugia: joint inference from fossil records, species distribution models and phylogeography. New Phytol 204:37–54
Guisan A, Zimmermann NE (2000) Predictive habitat distribution models in ecology. Ecol Model 135:147–186
Gunarathne R, Perera G (2014) Climatic factors responsible for triggering phenological events in Manilkara hexandra (Roxb.) Dubard., a canopy tree in tropical semi-deciduous forest of Sri Lanka. Trop Ecol 55:63–73
Hällfors M, Aikio S, Fronzek S, Hellmann J, Ryttäri T, Heikkinen R (2016) Assessing the need and potential of assisted migration using species distribution models. Biol Conserv 196:60–68
Hansen MC, Potapov PV, Moore R, Hancher M, Turubanova S, Tyukavina A, Thau D, Stehman S, Goetz S, Loveland T (2013) High-resolution global maps of 21st-century forest cover change. Science 342:850–853
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978
Hoffmann M, Hilton-Taylor C, Angulo A, Böhm M, Brooks TM, Butchart SH, Carpenter KE, Chanson J, Collen B, Cox NA (2010) The impact of conservation on the status of the world’s vertebrates. Science 330:1503–1509
Horev A, Yosef R, Tryjanowski P, Ovadia O (2012) Consequences of variation in male harem size to population persistence: modeling poaching and extinction risk of Bengal tigers (Panthera tigris). Biol Conserv 147:22–31
IPCC (2007) Climate change 2007: impacts, adaptation and vulnerability. Contribution of working group II to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Islam A, Muzaffar SB, Aziz MA, Kabir MM, Uddin M, ChaNma S, Chowdhury SU, Rashid MA, Chowdhury GW, Mohsanin S (2010) Baseline survey of bears in Bangladesh 2008–2010. Wildlife Trust of Bangladesh
IUCN (2000) 2000 IUCN Red List of Threatened Species. IUCN, Gland, Switzerland and Cambridge, UK
IUCN (2014) The IUCN Red List of Threatened Species. Version 2014.1. http://www.iucnredlist.org. Downloaded on 28 September 2015
IUCN (2016) The IUCN Red List of Threatened Species. Version 2016.2. http://www.iucnredlist.org. Downloaded on 21 July 2016
Izumiyama S, Shiraishi T (2004) Seasonal changes in elevation and habitat use of the Asiatic black bear (Ursus thibetanus) in the northern Japan alps. Mammal Study 29:1–8
Khan MMH (2008) Protected areas of Bangladesh: a guide to wildlife: Nishorgo program, wildlife management and nature conservation Circle, Bangladesh Forest Department
Levinsky I, Skov F, Svenning J-C, Rahbek C (2007) Potential impacts of climate change on the distributions and diversity patterns of European mammals. Biodivers Conserv 16:3803–3816
Li J, McCarthy TM, Wang H, Weckworth BV, Schaller GB, Mishra C, Lu Z, Beissinger SR (2016) Climate refugia of snow leopards in high Asia. Biol Conserv 203:188–196
Loucks C, Barber-Meyer S, Hossain MAA, Barlow A, Chowdhury RM (2010) Sea level rise and tigers: predicted impacts to Bangladesh’s Sundarbans mangroves. Clim Chang 98:291–298
Magris RA, Pressey RL, Weeks R, Ban NC (2014) Integrating connectivity and climate change into marine conservation planning. Biol Conserv 170:207–221
Michalski F, Peres CA (2007) Disturbance-mediated mammal persistence and abundance-area relationships in Amazonian forest fragments. Conserv Biol 21:1626–1640
Morelli TL, Daly C, Dobrowski SZ, Dulen DM, Ebersole JL, Jackson ST, Lundquist JD, Millar CI, Maher SP, Monahan WB (2016) Managing climate change refugia for climate adaptation. PLoS One 11:e0159909
Moss RH, Edmonds JA, Hibbard KA, Manning MR, Rose SK, Van Vuuren DP, Carter TR, Emori S, Kainuma M, Kram T (2010) The next generation of scenarios for climate change research and assessment. Nature 463:747–756
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GA, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Olson DM, Dinerstein E (1998) The global 200: a representation approach to conserving the Earth’s most biologically valuable ecoregions. Conserv Biol 12:502–515
Pacifici M, Foden WB, Visconti P, Watson JE, Butchart SH, Kovacs KM, Scheffers BR, Hole DG, Martin TG, Akçakaya HR (2015) Assessing species vulnerability to climate change. Nat Clim Chang 5:215–224
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Evol Syst 37:637–669
Peres CA, Palacios E (2007) Basin-wide effects of game harvest on vertebrate population densities in Amazonian forests: implications for animal-mediated seed dispersal. Biotropica 39:304–315
Phillips SJ, Dudík M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161–175
Phillips SJ, Dudík M, Schapire RE (2004) A maximum entropy approach to species distribution modeling. Proceedings of the twenty-first international conference on machine learning. ACM Press, New York, pp 655–662
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Phillips SJ, Dudík M, Elith J, Graham CH, Lehmann A, Leathwick J, Ferrier S (2009) Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol Appl 19:181–197
Pokharel KP, Ludwig T, Storch I (2016) Predicting potential distribution of poorly known species with small database: the case of four-horned antelope Tetracerus quadricornis on the Indian subcontinent. Ecol Evol 6:2297–2307
Puyravaud JP, Davidar P, Srivastava RK, Wright B (2017) Modelling harvest of Asian elephants Elephas maximus on the basis of faulty assumptions promotes inappropriate management solutions. Oryx 51:506–512
Rondinini C, Di Marco M, Chiozza F, Santulli G, Baisero D, Visconti P, Hoffmann M, Schipper J, Stuart SN, Tognelli MF (2011) Global habitat suitability models of terrestrial mammals. Philos Trans R Soc Lond B Biol Sci 366:2633–2641
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Sanderson EW, Forrest J, Loucks C, Ginsberg J, Dinerstein E, Seidensticker J, Leimgruber P, Songer M, Heydlauff A, O’Brien T (2010) Setting priorities for tiger conservation: 2005–2015. Tigers of the world: the science, politics, and conservation of Panthera tigris. William Andrew Publishing, Boston, pp 143–161
Sathyakumar S (2006) The status of Asiatic black bears in India. Understanding Asian bears to secure their future. Japan Bear Network, Ibaraki, Japan, pp 12–19
Secretariat of the CBD (2010) Conference of the parties 10 decision X/2. Strategic Plan for Biodiversity 2011-2020, 1–13
Shoshani J, Eisenberg JF (1982) Elephas maximus. Mamm Species Arch 182:1–8
Sodhi NS, Koh LP, Brook BW, Ng PK (2004) Southeast Asian biodiversity: an impending disaster. Trends Ecol Evol 19:654–660
Sohl TL (2014) The relative impacts of climate and land-use change on conterminous United States bird species from 2001 to 2075. PLoS One 9:e112251
Srinivasulu C, Srinivasulu B (2012) South Asian Mammals. South Asian mammals. Springer, p 9–98
Stanton JC, Pearson RG, Horning N, Ersts P, Reşit Akçakaya H (2012) Combining static and dynamic variables in species distribution models under climate change. Methods Ecol Evol 3:349–357
Sukumar R (1992) The Asian elephant: ecology and management. Cambridge University Press, Cambridge
Thomas CD, Cameron A, Green RE, Bakkenes M, Beaumont LJ, Collingham YC, Erasmus BF, De Siqueira MF, Grainger A, Hannah L (2004) Extinction risk from climate change. Nature 427:145–148
Ting S, Hartley S, Burns K (2008) Global patterns in fruiting seasons. Glob Ecol Biogeogr 17:648–657
Trisurat Y, Bhumpakphan N, Reed DH, Kanchanasaka B (2012) Using species distribution modeling to set management priorities for mammals in northern Thailand. J Nat Conserv 20:264–273
Visconti P, Bakkenes M, Baisero D, Brooks T, Butchart SH, Joppa L, Alkemade R, Di Marco M, Santini L, Hoffmann M, Maiorano L, Pressey RL, Arponen A, Boitani L, Reside AE, Van Vuuren DP, Rondinini C (2016) Projecting global biodiversity indicators under future development scenarios. Conserv Lett 9:5–13
Walston J, Robinson JG, Bennett EL, Breitenmoser U, da Fonseca GA, Goodrich J, Gumal M, Hunter L, Johnson A, Karanth KU (2010) Bringing the tiger back from the brink—the six percent solution. PLoS Biol 8:e1000485
Wiegand T, Revilla E, Moloney KA (2005) Effects of habitat loss and fragmentation on population dynamics. Conserv Biol 19:108–121
Wilson JW, Sexton JO, Jobe RT, Haddad NM (2013) The relative contribution of terrain, land cover, and vegetation structure indices to species distribution models. Biol Conserv 164:170–176
Acknowledgements
This research was funded by International Postgraduate Research Scholarship (IPRS) and The University of Queensland (UQ) Centennial Scholarship to the first author. We would like to acknowledge the Research Grant from School of Earth and Environmental Sciences, The University of Queensland for funding this research. We also thank the anonymous reviewers for their excellent comments on the earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 17431 kb)
Rights and permissions
About this article

Cite this article
Deb, J.C., Phinn, S., Butt, N. et al. Adaptive management and planning for the conservation of four threatened large Asian mammals in a changing climate. Mitig Adapt Strateg Glob Change 24, 259–280 (2019). https://doi.org/10.1007/s11027-018-9810-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11027-018-9810-3