
Abstract
Hepatic encephalopathy (HE) is a neuropsychiatric complication of acute liver failure or chronic liver injury. Liver dysfunction impairs ammonia detoxification, allowing it to cross the blood-brain barrier (BBB) and disrupt brain function. The hippocampus becomes a crucial target during elevated ammonia levels, causing spatial memory impairment and decreased learning ability. Leuprolide acetate (LA), a GnRH agonist, has been implicated in neuroprotection and neuroregeneration in several regions of the central nervous system (CNS) including hippocampus. In this study, we aim to evaluate the effects of LA treatment on hippocampus of rats with HE induced by portocaval anastomosis (PCA) trough cognitive tests, histology analysis and expression of neuronal recovery marker proteins, such as neurofilament (NF200) and neurabin II, and astrocyte marker glial fibrillary acidic protein (GFAP). Rats were divided into three groups: SHAM, portocaval anastomosis with saline solution (PCA + SS) and portocaval anastomosis treated with LA (PCA + LA). To evaluate learning and spatial memory elevated T-maze (ETM) and Y-maze test (YMT) were respectively used. Results indicated that LA-treated rats performed significantly better in ETM and YMT than untreated rats. Histological analysis of hippocampus showed increased neuron density, nuclear area, and layer thickness in dentate gyrus of PCA + LA group compared to PCA + SS. Additionally, neurabin II and NF200 expression were higher in LA-treated rats, while GFAP expression was elevated in the PCA + SS group compared to control and PCA + LA groups. In conclusion, LA enhances hippocampal neuron recovery and reduces astrogliosis, suggesting its potential as a therapeutic intervention for attenuating hippocampal damage during HE.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Hepatic encephalopathy (HE) is a neuropsychiatric complication of acute liver failure or chronic liver injury (França et al. 2019). Liver failure disrupts ammonia detoxification, allowing it to breach the BBB and interfere with brain function (Chen et al. 2014). Hyperammonemia has been directly implicated in the progression of HE, causing neurotoxicity and activating apoptotic pathways, particularly in neurons, contributing to cell death (Bosoi and Rose 2009). The hippocampus becomes a crucial target during elevated ammonia levels; patients with HE exhibits hippocampal atrophy marked by reduced volume and functional connectivity (García-García et al. 2018; Lin et al. 2019). This atrophy contributes to episodic and spatial memory impairment, decreased learning ability, and altered navigation function (Ochoa-Sanchez et al. 2021). The mechanisms leading to these alterations are poorly understood, and there are no treatments to improve cognitive function (Monfort et al. 2009). Animal models, such portocaval anastomosis, have been used to study the progression of HE and explore new information for potential treatments. This procedure has been demonstrated to induce hyperammonemia, resulting in hippocampus neuron damage and subsequent cognitive impairment. These include diminished learning process during a conditional discrimination task in a Y-maze, as well as reduced abilities in reversal and spatial learning observed in the Morris water maze (Erceg et al. 2005; Cauli et al. 2007; Méndez et al. 2010).
Gonadotropin-releasing hormone (GnRH) is a decapeptide known for regulating the hypothalamus-pituitary-gonad axis (Millar 2005). Interestingly, GnRH receptor (GnRHR) has been identified in the CA1–3 areas and dentate gyrus of the hippocampus, showing its participation in non-reproductive functions (Ferris et al. 2015). Studies on rat hippocampal culture have demonstrated that GnRH administration up-regulates markers of synaptic plasticity, such as synaptophysin and spinophilin, indicating a well-established role for GnRH in hippocampal neuroplasticity and neuroprotection (Schang et al. 2011). Quintanar and Salinas (2008) reported that GnRH enhances the number and length of neuronal process in cortical neurons of rat embryos. In a spinal cord injury model, both GnRH and its analogue, LA, were found to favor the expression of proteins such as neurofilaments and myelin basic protein (Calderón-Vallejo and Quintanar 2012; Díaz-Galindo et al. 2015).
Additionally, treatment with LA has been shown to promote the recovery of hippocampal neurons in an acute brain ischemia-reperfusion injury model (Montoya-García et al. 2023). Based on these findings, LA may attenuate neuronal damage in the hippocampus of rats during HE. This study aims to evaluate the effects of LA treatment on the hippocampus of rats with HE induced by portocaval anastomosis through cognitive tests, histology analysis, and expression of neuronal recovery markers proteins, such as NF200, neurabin II and astrocyte marker GFAP.
Materials and methods
Animals
This study used male Wistar rats weighing 250 to 350 g. The animals were sourced from the Autonomous University of Aguascalientes farm and handled following the Animal Ethics and Welfare Committee of the Autonomous University of Aguascalientes (CEADI/UAA/0025/18). The rats were housed in a light-dark cycle (12 h/12 h) with controlled temperature (~22 °C) and ad libitum access to food and water.
Portocaval anastomosis
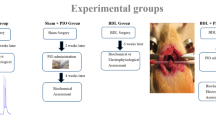
An end-to-side portocaval anastomosis was performed to induce hepatic encephalopathy following the protocol described by Navarro-Gonzalez et al. (2023). Rats were anesthetized with 3.0% isoflurane with a 250 ml/min flow rate during the surgery. Anastomosis of both veins was completed in less than 15 min using the continuous suture technique with an 8-0 polyglycolic acid suture. Post-surgery, all rats received a single injection of 10,000 IU of DEXA-streptovet (penicillin, streptomycin, and dipyrone solution). They were housed in solid-bottomed plastic cages and provided unlimited access to food and water. Additionally, all received 10% glucose-supplemented drinking water for the first 3 days after surgery. The SHAM group (n = 11) underwent the same procedure without cutting vessels after blood flow was suppressed (Fig. 1).
Treatment
The treatment was initiated 4 weeks after PCA surgery. PCA rats were randomly divided into two groups: portocaval anastomosis with saline solution (PCA + SS n = 16), and portocaval anastomosis treated with LA (PCA + LA n = 18). Leuprolide acetate (10 µg/kg i.m) was administered for three consecutive days, followed by subsequent administration every third day for a total of 5 weeks, following the protocol established by Guzmán-Soto et al. (2012), (Fig. 1).

Scheme showing the experimental design
Y-maze test: spatial memory
To evaluate short-term spatial reference memory performance in the final week of the experiment, the animals underwent the Y-maze test following the methodology proposed by Kraeuter et al. (2019). Initially, a 30-min habituation process was conducted in the work area. Subsequently, the rats were placed into the acrylic Y-maze with arms measuring 35 cm long, 10 cm wide, and 25 cm high. During the training session, one arm of the maze was closed, allowing the rats to explore for 10 min. Following this, each rodent was returned to its cage for 1-h intervals. Next, the test session continued, placing each rat back in the maze, with the previously closed arm open (designated as the new arm), allowing the rats to explore for 5 min. The test session was videotaped and analyzed using an automated system (Smart video tracking system, Panlab; Harvard Apparatus). The number of entries in the new arm was compared with entries in the other arms to evaluate spatial memory. A rat exhibiting no preference for either arm during the test session suggests impaired spatial memory, potentially indicating altered hippocampal functioning.
Elevated T-maze test: learning
The elevated T-maze is widely used to assess anxiety, memory, and learning levels in rodents (Asth et al. 2012). This test was implemented following the methodology that González-Torres et al. (2019) proposed. The apparatus used in this study consisted of a T-shaped maze elevated above the floor, with two open opposing arms and one closed arm with an open roof. The procedure comprised two sessions, one for training and another for the test. Before testing, habituation to the workspace was conducted, as mentioned earlier. In the training session, each subject was placed at the end of the closed arm of the maze and allowed to explore for 300 s. The test concluded each time the rat placed all four paws in one of the open arms of the maze. After leaving the closed arm, the rat was placed in isolation in a box for 30 s. This last step was repeated until each rat remained in the closed arm for 300 s or for a maximum of 10 trials. The test session took place 24 h after the training session; the rats were placed in the closed arm, and the time it took them to enter one of the open arms was recorded.
Ammonia measurements
Hepatic encephalopathy has been directly linked to an elevation in systemic ammonia (García-Lezana et al. 2017). Blood ammonium levels were quantified to validate that the portocaval anastomosis model can induce this effect. The ammonia levels were measured using the Ammonia Assay Kit (AA0100, Sigma-Aldrich) according to the manufacturer’s protocols. Three rats per group were used to measure the ammonia levels.
Histological analysis
Under deep anesthesia, rats underwent intracardiac perfusion with a 0.9% saline solution followed by 10% neutral phosphate-buffered formalin. Subsequently, the brains of three animals per group were removed and embedded in the same fixative solution. Dorsal hippocampus sections were made in the bregma region − 3.36 mm, following the coordinates outlined in the Paxinos and Watson atlas (Paxinos and Watson 2006). Hematoxylin-eosin staining was performed on the sections. Photomicrographs were captured using a 40X objective lens with a camera AmScope MU1803 coupled to a Zeiss Axiostar Plus microscope. Two independent blinded observers evaluated nine randomized optical fields of three sections per group per area, analyzing neuron density, thickness, and nuclear area in CA1, CA3, and dentate gyrus (DG). The results of the analysis are presented per visual field.
Western blot
The hippocampus of 5 animals per group was rapidly dissected and homogenized in an extraction cocktail containing protease inhibitors (20mM Tris/HCl, 1 Mm EGTA, 1mM PMSF, 10mM Cl2Mg, 0.6mM aprotinin, 2µM leupeptin). Protein concentrations were determined using the bicinchoninic acid (BCA) method (23227 Thermo Scientific Pierce). Equal amounts of protein (25 µg) were separated on sodium dodecyl sulfate-polyacrylamide gels by electrophoresis and transferred onto polyvinylidene difluoride membranes. The membranes were incubated overnight with the anti-NF200 (1:1000, N0142 Sigma), anti-neurabin II (1:300 A20 Santacruz), anti-GFAP (1:5000, AB7260 ABCAM), and anti-GAPDH (1:5000, G9545 Sigma), with secondary IgG antibody, conjugated with horse-radish peroxidase (HRP) for 2 h at room temperature. Chemiluminescence was used for membrane development, and the signals were visualized using Image LabTM (Bio-Rad). The relative density of the bands was analyzed using FIJI software (ImageJ2). Band densities were normalized using the signal intensity of GAPDH.
Statistical analysis
Results are expressed as mean ± standard error of the mean (SEM). Group data comparisons were conducted using one-way ANOVA with Kruskal-Wallis test post-hoc test for the cognitive tests, Fisher’s LSD as post hoc for histological, and western blot analysis, utilizing GraphPad Prism 8.0 (GraphPad Software, San Diego, CA, USA). Differences were considered significant when p < 0.05.
Results
Body weight, percentage of liver weight and ammonia in serum
The percentage of body weight of both PCA groups significantly decreased compared to the SHAM group. However, the group treated with LA showed less weight loss than the untreated group (PCA + SS: 21.87 ± 6.15%, p < 0.001; PCA + LA: 8.76 ± 2.78%, p < 0.05), (Table 1). Regarding the percentage of liver weight, both the treated and untreated groups demonstrated a significant difference compared with the control group, with a decrease of 58% and 56%, respectively (SHAM: 3.34 ± 0.17%, PCA + SS: 1.94 ± 0.08%, PCA + LA: 1.88 ± 0.08%, p < 0.0001), (Table 1). To quantitatively validate the liver damage caused by the portocaval anastomosis, serum ammonia levels were measured. A significant increase in ammonia was observed in both PCA + SS and PCA + LA groups compared to SHAM group (SHAM: 1.97 ± 0.03 µg/ml, PCA + SS: 4.65 ± 0.99 µg/ml, PCA + LA: 4.04 ± 0.36 µg/ml, p < 0.05), (Table 1). Indicating a state of hyperammonemia in both PCA groups.
Y-maze test: spatial memory
The short-term spatial memory of the different groups was assessed using the Y maze. During this test, the number of entries into the new arm was compared with to those into the other arms. The SHAM group preferred visiting the new arm, indicating spatial memory acquisition. In contrast, the SS group showed a significant reduction in entries into the new arm compared to the SHAM group, suggesting memory impairment (SHAM: 46.31 ± 7.71% vs. PCA + SS: 26.07 ± 2.29%, p < 0.05), (Fig. 2). On the other hand, rats treated with LA exhibited an increase in entries significantly higher than the SS group but not different from the control group, demonstrating an improvement in spatial memory compared to untreated rats (PCA + SS: 26.07 ± 2.29% vs. PCA + LA: 37.6 ± 3.07%, p < 0.05), (Fig. 2).

Short-term spatial memory analysis in the Y-maze test. Percentage of entries to novel arm (%). Experimental groups: SHAM (control) n = 5, portocaval anastomosis rats treated with saline solution (PCA + SS) n = 15, portocaval anastomosis rats treated with leuprolide acetate (PCA + LA) n = 13. Results are shown as mean and SEM. *p < 0.05, determinate by one-way ANOVA with Kruskal-Wallis test post-hoc test
Elevated T-maze test: learning
The T-maze test revealed a significant decrease in the mean latency of scape in the untreated group compared to the treated group (PCA + SS:174.4 ± 54.91s vs. PCA + LA:300 ± 0.0s, p < 0.001), suggesting the inability of these rats to learn the task (Fig. 3A). However, the PCA + LA group showed an increase in the latency of scape in contrast to the SHAM group, indicating that the LA stimulated learning during the test (SHAM: 243.17 ± 31.8s vs. PCA + LA:300 ± 0.0s, p < 0.001), (Fig. 3A). This effect was also evident when evaluating subjects that met the 300s criterion, with LA-treated rats achieving a perfect score, higher than the control group and the SS group (SHAM: 66.67 ± 21.08%, PCA + SS: 20.0 ± 20.0%, PCA + LA: 100.0 ± 0% p < 0.05), (Fig. 3B).

Learning task in elevated T-maze. A Latency to explore open arms in the habituation and test session. B Percentage of rats of each group that achieved the criterion of 300s in the test session (perfect performance). Subjects with perfect performance were assigned 100%, while those without perfect performance were assigned 0%. SHAM (control) n = 6, portocaval anastomosis rats treated with saline solution (PCA + SS) n = 5, portocaval anastomosis rats treated with leuprolide acetate (PCA + LA) n = 7. Results are shown as mean and SEM. *p < 0.05, **p < 0.001, ***p < 0.0001, determinate by one-way ANOVA with Kruskal-Wallis test post-hoc test
Histological analysis
A quantitative analysis of neuron density, layer thickness, and nuclear area was conducted in the CA1, CA3, and DG regions of the hippocampus (Fig. 4A).

Effect of leuprolide acetate on hippocampal areas CA1, CA3, and DG of rats with portocaval anastomosis. A Hippocampus photomicrographs of rats SHAM (control) n = 3, portocaval anastomosis rats treated with saline solution (PCA + SS) n = 3, portocaval anastomosis rats treated with leuprolide acetate (PCA + LA) n = 3. Tissue was stained with hematoxylin-eosin. Scale bars in CA1, CA3, and DG 20 μm. B Neuron density in CA1, CA3 and DG in all groups. C Nuclear area in CA1, CA3, and DG in all grops. D Layer thickness of CA1, CA3, and DG of all groups. Results are shown as mean and SEM. *p < 0.05, **p < 0.001 determinate by one-way ANOVA with Fisher as a post-hoc test
Neuron density
The density of neurons in the CA1 and CA3 areas decreased in both PCA groups compared to the control (SHAM CA1: 36.67 ± 2.82, PCA + SS CA1: 27.0 ± 2.74, PCA + LA: 29.33 ± 1.98, p < 0.05; SHAM CA3: 22.33 ± 2.32, PCA + SS CA3: 14.83 ± 2.15, PCA + LA CA3: 16.17 ± 2.46, p < 0.05), (Fig. 4B). However, in the DG area of the PCA + LA group the number of neurons was significantly higher than in the PCA + SS group but not different from the SHAM group (SHAM: 102.08 ± 3.0, PCA + SS: 86.58 ± 4.77, PCA + LA: 101.83 ± 4.65, p < 0.001), (Fig. 4B).
Nuclear area
The results revealed that the nuclear area in the CA1 and CA3 regions was similar among the three groups (Fig. 4C). Conversely, in the DG of the treated group, the nuclear area significantly increased compared to the untreated group, although it did not differ from the control group (SHAM: 46.89 ± 2.15µm2, PCA + SS: 41.06 ± 1.24µm2, PCA + LA: 49.5 ± 1.68µm2, p < 0.001), (Fig. 4C).
Layer thickness
In the layer thickness of the CA1 area, a significant decrease was observed in the PCA + SS group compared to the SHAM group ((SHAM: 51.15 ± 2.37 μm, PCA + SS: 42.73 ± 2.18 μm, p < 0.05), (Fig. 4D). When the CA3 was analyzed, a reduction was noted in both PCA groups compared to the control (SHAM: 67.45 ± 3.87 μm, PCA + SS: 53.90 ± 2.07 μm, PCA + LA: 53.78 ± 3.9 μm, p < 0.05), (Fig. 4D). However, in the DG, an increase in layer thickness was observed in the treated rats against the untreated group (PCA + SS: 55.41 ± 1.07 μm, PCA + LA: 61.86 ± 2.4 μm, p < 0.05), (Fig. 4D).
Western blot
To evaluate the effect of LA treatment on hippocampal neurons, we analyzed the expression of two proteins involved in neuron structure: neurabin II and NF200. The analysis of neurabin II expression demonstrated a significant decrease in the PCA + SS group compared to the control. There were no differences in the PCA + LA and the other groups; however, we observed a tendency to increase the expression of neurabin II compared to the untreated group (PCA + SS: 45.83 ± 9.36%, PCA + LA: 67.38 ± 8.87%), (Fig. 5). On the other hand, NF200 expression of LA-treated rats showed a significant increase compared to the untreated group, but no difference from the control (PCA + SS: 32.26 ± 9.3%, PCA + LA: 65.25 ± 7.79%, p < 0.05), (Fig. 6).

Western blot analysis of neurabil II and GAPDH expression in the hippocampus of rats with portocaval anastomosis. A Immunoblot representative of neurabin II and GAPDH in SHAM (control) n = 5, portocaval anastomosis rats treated with saline solution (PCA + SS) n = 5 and portocaval anastomosis rats treated with leuprolide acetate (PCA + LA) n = 5. B Densitometric analysis values are expressed as optical density relative to GAPDH per 25 µg of protein. *p < 0.05 determinate by one-way ANOVA with Fisher as a post-hoc test

Western blot analysis of NF200 and GAPDH expression in the hippocampus of rats with portocaval anastomosis. A Immunoblot representative of NF200 and GAPDH in SHAM (control) n = 5, portocaval anastomosis rats treated with saline solution (PCA + SS) n = 5, and portocaval anastomosis rats treated with leuprolide acetate (PCA + LA) n = 5. B) Densitometric analysis values are expressed as optical density relative to GAPDH per 25 µg of protein. *p < 0.05 determinate by one-way ANOVA with Fisher as a post-hoc test
To evaluate the effect of LA treatment on astrocytes, we assessed GFAP, an astrocyte activation marker, in the hippocampus by western blot. The expression of GFAP was significantly higher in the PCA + SS group, while in the PCA + LA group, the expression of GFAP was reduced compared to the PCA + SS group (PCA + SS: 140.7 ± 17.23%, PCA + LA: 92.85 ± 11.73%, p < 0.05), (Fig. 7).

Western blot analysis of GFAP and GAPDH expression in the hippocampus of rats with portocaval anastomosis. A Immunoblot representative of GFAP and GAPDH in SHAM (control) n = 5, portocaval anastomosis rats treated with saline solution (PCA + SS) n = 5, and portocaval anastomosis rats treated with leuprolide acetate (PCA + LA) n = 5. B Densitometric analysis values are expressed as optical density relative to GAPDH per 25 µg of protein. *p < 0.05 determinate by one-way ANOVA with Fisher as a post-hoc test
Discussion
PCA is a surgical model accepted by the International Society for Hepatic Encephalopathy and Nitrogen Metabolism that reproduces some neurological dysfunctions in patients with HE (DeMorrow et al. 2021). As in previous reports, our model accomplished to present a series of alterations characteristic of rats with PCA, including the reduction of body weight, liver atrophy, and hyperammonemia (Vázquez-Martínez et al. 2019; Navarro-Gonzalez et al. 2023; Cauli et al. 2007). We observed decreased spatial memory and learning in the PCA + SS group in both the Y-maze and the elevated T-maze test. This cognitive impairment can be attributed to increased ammonia intake in the brain. Several reports indicate that an elevated amount of this compound can alter cognitive processes. Erceg et al. (2005) reported the diminution in learning capacity during the Y-maze test of both PCA and hyperammonemia rats without liver failure. Additionally, Cauli et al. (2007) demonstrated the reduced ability of PCA rats to learn a conditional discrimination task in the Y-maze test, likely due to inflammation and hyperammonemia affecting the function of the glutamate–NO–cGMP pathway in the brain. Also, it has been reported that chronic hyperammonemia significantly reduces the degree of long-term potentiation and glutamatergic synaptic transmission, both necessary for the establishment of learning and memory (Muñoz et al. 2000; Hernandez-Rabaza et al. 2015; Méndez et al. 2010). Furthermore, it has been described in BLD rats that changes in the electrophysiological firing of pyramidal neurons in the CA1 region may be implicated in cognitive alterations (Tahamtan et al. 2017).
The hippocampal histology revealed reduced thickness and neuron density in the CA1 and CA3 regions of both PCA groups. Previous studies have illustrated decreased hippocampal volume and functional connectivity in cirrhotic patients with HE (Lin et al. 2019). Additionally, reports indicate structural and functional alterations of neurons and astrocytes in the CA1 region in hyperammonemic rats with partial portal vein ligation (PVL), (Tallis et al. 2014). A limited number of studies address the structural changes in neurons that occur in various areas of the hippocampus during HE. However, the main consensus suggests that the key to understanding neurodegeneration during the progression of HE lies within the astrocytes. Astrocytes provide a variety of supportive functions to neurons, including structural support, energy metabolism, delivery of trophic factors, modulation of synaptic transmission, long-distance communication, and involvement in inflammatory processes (Jurga et al. 2021).
Additionally, astrocytes play a vital role in ammonia detoxification since they express glutamine synthetase, an enzyme metabolizing ammonia. The elevation of ammonia in the brain results in a high concentration of intracellular glutamine, causing an ion disturbance and astrocyte swelling (Jaeger et al. 2019). Consequently, ammonia-induced astrocyte swelling may leave the neuronal network vulnerable and without support, leading to altered neuronal integrity and function (Ochoa-Sanchez et al. 2021).
Surprisingly, despite the observed alteration in the CA1 and CA3 areas, rats treated with LA significantly improved cognitive tasks compared to the untreated group. This improvement is likely associated with an increase in the number and size of granular neurons in the DG region of the hippocampus. Montoya-García et al. (2023) reported a significantly higher density of neurons in the DG area in rats with brain ischemia-reperfusion injury treated with LA compared to the injury-only group. Interestingly that the DG area remained intact in the PCA + LA group, as this is the region where neurogenesis occurs (Hainmueller and Bartos 2020). Spatial learning has been described to regulate adult hippocampal neurogenesis by controlling the number of surviving adult-born neurons and accelerating their development (Tronel et al. 2010). Therefore, the effect of LA on spatial memory and learning may be associated with the proliferation and survival of newborn neurons. Furthermore, it is plausible that the GnRH analogue decreases apoptosis in the DG of the hippocampus, contributing to cognitive enhancement in PCA rats treated with LA (Chu et al. 2008). Neurofilaments are expressed in neuronal cells and play an important role in structural support. Three subunits have different molecular weights: light, medium, and heavy (Yuan et al. 2017). The heavy chain of 200 kDa is crucial for interactions with microtubules, association with other proteins, and axonal transport. Disturbances in these neurofilaments can modify the structure of the axon cytoskeleton complex, thereby contributing to the development of neurodegenerative diseases (Porseva et al. 2013). Our study demonstrates decreased NF200 expression in the hippocampus following PCA in untreated rats. These results indicate changes in neuronal shape and extension of neurites, factors that rely on the integrity of the cytoskeleton. Similar findings were observed in rats with partial portal vein ligation, which presented a significant diminution in immunolabeling of NF200 in CA1, indicating a reduction in neurites (Tallis et al. 2014).
In contrast, PCA animals treated with LA showed a significant upregulation in NF200 expression compared to untreated ones but no difference to the control group. Studies have demonstrated that GnRH enhances the expression of NF200 and NF68 in cortical neurons and spinal cord neurons of rat embryos, leading to an increase in both the number and length of neurite outgrowths (Quintanar and Salinas 2008; Quintanar et al. 2016). Furthermore, an elevated expression of NF200 has been confirmed in various injury models treated with GnRH or leuprolide acetate, such as spinal cord injury, experimental autoimmune encephalomyelitis, and brain ischemia reperfusion (Calderón-Vallejo and Quintanar 2012; Guzmán-Soto et al. 2012; Montoya-García et al. 2023). All these findings suggest that GnRH and its analogue AL may act as neurotrophic factors, increasing neurofilament expression.
Dendritic spines are small protrusions from dendritic shafts that receive most of the excitatory input in the CNS (Feng et al. 2000). Neurabin II, also known as spinophilin, is an actin and protein phosphatase-1 (PP1) binding protein highly enriched in dendritic spines. Its localization and functional properties suggest a potential linkage between synaptic transmission and alterations in the structure and function of dendritic spines (Grossman et al. 2004). In our study, we observed a significant decrease in the expression of neurabin II in rats with PCA + SS compared to the control group. Hyperammonemia has been related to a decreased spine density in hippocampal CA1 pyramidal neurons of rats with bile conduct ligation (Chen et al. 2014). Reduced synaptic activity has been associated with loss of dendritic spines and, consequently, impairment in the learning process (Feng et al. 2000). Furthermore, glutamate receptors NMDA and AMPA are associated with synaptic plasticity, which is crucial for memory and learning processes. Deregulation of these receptors can lead to cognitive deterioration and decreased memory (Llansola et al. 2013). During events of increased ammonia, alterations in glutamate transmission have been documented (Llansola et al. 2005, 2013; Cauli et al. 2009). Monfort et al. (2007) observed reduced activation of NMDA and AMPA receptors in hippocampal slices from PCA rats, resulting in altered neurotransmission and impaired communication between neurons. Therefore, our findings of diminished neurabin II expression in the hippocampus of PCA rats may be attributed to altered synaptic transmission, contributing to the cognitive dysfunctions observed during Y-maze and Elevated T-maze tests.
On the other hand, although PCA rats treated with LA showed no significant difference compared to untreated rats, there is a tendency toward an increase in the expression of neurabin II. González-Torres et al. (2019) reported that improved learning in old gonadectomized rats treated with GnRH is related to a greater number of dendritic contacts associated with increased expression in this protein. Previous studies with cultured spinal cord neurons of rat embryos demonstrated that incubation with GnRH promoted more neuritic contacts, consistent with an increased expression of neurabin II (Quintanar et al. 2016). Additionally, it has been shown that, through activation of its receptor in primary cultures of hippocampal cells, GnRH stimulates the expression of two marker genes of neuronal plasticity: synaptophysin and spinophilin (Schang et al. 2011). It leads to increased dendritic spine density and enhanced neural excitation via actions on ionotropic glutamate receptors. Several lines of evidence suggest that GnRH modulates hippocampal function and plasticity, as this structure exhibits the highest density of GnRHR within the brain (Jennes et al. 1988). Therefore, GnRH may exert neuromodulatory actions on the hippocampus (Ferris et al. 2015).
As previously mentioned, astrocytes are crucial in providing tissue structural support and maintaining homeostasis. A commonly used marker for astrocytes is GFAP, an intermediate cytoskeletal protein present in most CNS astrocytes. The main function of GFAP is to mechanically support astrocytes and the structure of the BBB (Jurga et al. 2021). Overexpression of GFAP has been associated with reactive astrogliosis, a characteristic alteration in the morphology and function of astrocytes observed in many neurological disorders such as neurotrauma, ischemic stroke, and neurodegenerative disease (Pekny et al. 2016). Astrogliosis has also been reported in patients with HE and in rats with portal systemic shunting (Agarwal and Mais 2019; Cruz and Duffy 1983). Our results demonstrate an increase in the expression of GFAP in PCA + SS group compared to the control and PCA + LA group. Similar results were observed in rats with hyperammonemia, where immunohistochemical detection of GFAP in the CA1 region of the hippocampus showed an increase in the expression of this protein compared to the SHAM group (França et al. 2019). An examination of astrocytic morphology in PVL rats in the hippocampal CA1 area demonstrated that GFAP-positive astrocytes exhibited an enlarged cell body with more tortuous and thicker processes, indicative of astroglial reaction (Tallis et al. 2014). Considering these reports, it can be suggested that the increased expression of GFAP in the hippocampus of PCA + SS rats may indicate a state of reactive astrogliosis.
Conversely, PCA animals treated with LA exhibited a decrease in GFAP expression, compared to untreated rats. Similarly, a reduction in GFAP was reported in rats with spinal cord injury treated with GnRH, where a diminution of GFAP immunoreactivity in spinal cord tissue was observed, suggesting a reduction in glial activity (Martínez-Moreno et al. 2023). Studies have demonstrated that the suppression of GFAP in animal models leads to decreases in reactive gliosis processes and increases in hippocampal neurogenesis, both under normal conditions and after injury. This indicates that GFAP plays a modulatory role in astrocytic regulation of neurogenesis (Wilhelmsson et al. 2012). Consequently, the reduction in GFAP expression due to LA treatment may contribute to regulating of neurogenesis in the hippocampus of PCA rats, thereby potentially improving cognitive performance. Finally, although all HE models have limitations and do not reproduce exactly all the alterations present in human patients, our study shows that LA improves cognitive function by promoting the recovery of hippocampal neurons and reducing astrogliosis. More studies need to be performed to elucidate the mechanisms and underlying effects of LA. However, based on our results, LA could be a promising treatment to attenuate HE symptoms in humans.
Conclusion
The results reported here demonstrated that treatment with LA in rats with PCA reduces neurodegeneration and astrogliosis, as evidenced by the increased expression of NF200 and neurabin II, along with a decrease in glial activation marker GFAP in the hippocampus, resulting in the restoration of spatial memory and learning functions. Therefore, LA can potentially improve cognitive function in patients with hepatic encephalopathy.
Data availability
No datasets were generated or analysed during the current study.
References
Agarwal AN, Mais DD (2019) Sensitivity and specificity of Alzheimer type II astrocytes in hepatic encephalopathy. Arch Pathol Lab Med 143:1256–1258
Asth L, Lobão-Soares B, André E, Soares V, de Gavioli P EC (2012) The elevated T-maze task as an animal model to simultaneously investigate the effects of drugs on long-term memory and anxiety in mice. Brain Res Bull 87:526–533
Bosoi CR, Rose CF (2009) Identifying the direct effects of ammonia on the brain. Metab Brain Dis 24:95–102
Calderón-Vallejo D, Quintanar JL (2012) Gonadotropin-releasing hormone treatment improves locomotor activity, urinary function and neurofilament protein expression after spinal cord injury in ovariectomized rats. Neurosci Lett 515:187–190
Cauli O, Rodrigo R, Piedrafita B, Boix J, Felipo V (2007) Inflammation and hepatic encephalopathy: Ibuprofen restores learning ability in rats with portacaval shunts. Hepatology 46:514–519
Cauli O, Rodrigo R, Llansola M, Montoliu C, Monfort P, Piedrafita B, El Mlili N, Boix J, Agustí A, Felipo V (2009) Glutamatergic and GABAergic neurotransmission and neuronal circuits in hepatic encephalopathy. Metab Brain Dis 24:69–80
Chen JR, Wang BN, Tseng GF, Wang YJ, Huang YS, Wang TJ (2014) Morphological changes of cortical pyramidal neurons in hepatic encephalopathy. BMC Neurosci 15:1–12
Chu C, Gao G, Huang W (2008) A study on co-localization of FSH and its receptor in rat hippocampus. J Mol Histol 39:49–55
Cruz NF, Duffy TE (1983) Local cerebral glucose metabolism in rats with chronic portacaval shunts. J Cereb Blood Flow Metab 3:311–320
DeMorrow S, Cudalbu C, Davies N, Jayakumar AR, Rose CF (2021) 2021 ISHEN guidelines on animal models of hepatic encephalopathy. Liver Int 41:1474–1488
Díaz-Galindo MDC, Gómez-González B, Salinas E, Calderón-Vallejo D, Hernández-Jasso I, Bautista E, Quintanar JL (2015) Leuprolide acetate induces structural and functional recovery of injured spinal cord in rats. Neural Regen Res 10:1819–1824
Erceg S, Monfort P, Hernández-Viadel M, Rodrigo R, Montoliu C, Felipo V (2005) Oral administration of sildenafil restores learning ability in rats with hyperammonemia and with portacaval shunts. Hepatology 41:299–306
Feng J, Yan Z, Ferreira A, Tomizawa K, Liauw JA, Zhuo M, Allen PB, Ouimet CC, Greengard P (2000) Spinophilin regulates the formation and function of dendritic spines. Proc Natl Acad Sci U S A 97:9287–9292
Ferris JK, Tse MT, Hamson DK, Taves MD, Ma C, McGuire N, Arckens L, Bentley GE, Galea LAM, Floresco SB, Soma KK (2015) Neuronal gonadotrophin-releasing hormone (GnRH) and astrocytic gonadotrophin inhibitory hormone (GnIH) immunoreactivity in the adult rat hippocampus. J Neuroendocrinol 27:772–786
França MER, Ramos RKLG, Oliveira WH, Duarte-Silva E, Araújo SMR, Lós DB, Peixoto CA (2019) Tadalafil restores long-term memory and synaptic plasticity in mice with hepatic encephalopathy. Toxicol Appl Pharmacol 379:114673
García-García R, Cruz-Gómez ÁJ, Urios A, Mangas-Losada A, Forn C, Escudero-García D, Kosenko E, Torregrosa I, Tosca J, Giner-Durán R, Serra MA, Avila C, Belloch V, Felipo V, Montoliu C (2018) Learning and memory impairments in patients with minimal hepatic encephalopathy are associated with structural and functional connectivity alterations in hippocampus. Sci Rep 8:1–13
García-Lezana T, Oria M, Romero-Giménez J, Bové J, Vila M, Genescà J, Chavarria L, Cordoba J (2017) Cerebellar neurodegeneration in a new rat model of episodic hepatic encephalopathy. J Cereb Blood Flow Metab 37:927–937
González-Torres ML, Calderón-Vallejo D, Quintanar JL (2019) Chronic administration of gonadotropin releasing-hormone improves learning in old gonadectomized rats. Neurobiol Learn Mem 157:35–40
Grossman SD, Futter M, Snyder GL, Allen PB, Nairn AC, Greengard P, Hsieh-Wilson LC (2004) Spinophilin is phosphorylated by Ca2+/calmodulin-dependent protein kinase II resulting in regulation of its binding to F-actin. J Neurochem 90:317–324
Guzmán-Soto I, Salinas E, Hernández-Jasso I, Quintanar JL (2012) Leuprolide acetate, a GnRH agonist, improves experimental autoimmune encephalomyelitis: a possible therapy for multiple sclerosis. Neurochem Res 37:2190–2197
Hainmueller T, Bartos M (2020) Dentate gyrus circuits for encoding, retrieval and discrimination of episodic memories. Nat Rev Neurosci 21:153–168
Hernandez-Rabaza V, Agusti A, Cabrera-Pastor A, Fustero S, Delgado O, Taoro-Gonzalez L, Montoliu C, Llansola M, Felipo V (2015) Sildenafil reduces neuroinflammation and restores spatial learning in rats with hepatic encephalopathy: underlying mechanisms. J Neuroinflammation 12:1–12
Jaeger V, Demorrow S, McMillin M (2019) The direct contribution of astrocytes and microglia to the pathogenesis of hepatic encephalopathy. J Clin Transl Hepatol 7:352–361
Jennes L, Dalati B, Michael Conn P (1988) Distribution of gonadrotropin releasing hormone agonist binding sites in the rat central nervous system. Brain Res 452:156–164
Jurga AM, Paleczna M, Kadluczka J, Kuter KZ (2021) Beyond the GFAP-astrocyte protein markers in the brain. Biomolecules 11(9):1361
Kraeuter AK, Guest PC, Sarnyai Z (2019) The Y-maze for assessment of spatial working and reference memory in mice. Methods Mol Biol 1916:105–111
Lin W, Chen X, Gao YQ, Yang ZT, Yang W, Chen HJ (2019) Hippocampal atrophy and functional connectivity disruption in cirrhotic patients with minimal hepatic encephalopathy. Metab Brain Dis 34:1519–1529
Llansola M, Erceg S, Felipo V (2005) Chronic exposure to ammonia alters the modulation of phosphorylation of microtubule-associated protein 2 by metabotropic glutamate receptors 1 and 5 in cerebellar neurons in culture. Neuroscience 133:185–191
Llansola M, Montoliu C, Cauli O, Hernández-Rabaza V, Agustí A, Cabrera-Pastor A, Giménez-Garzó C, González-Usano A, Felipo V (2013) Chronic hyperammonemia, glutamatergic neurotransmission and neurological alterations. Metab Brain Dis 28:151–154
Martínez-Moreno CG, Calderón-Vallejo D, Díaz-Galindo C, Hernández-Jasso I, Olivares-Hernández JD, Ávila-Mendoza J, Epardo D, Balderas-Márquez JE, Urban-Sosa VA, Baltazar-Lara R, Carranza M, Luna M, Arámburo C, Quintanar JL (2023) Gonadotropin-releasing hormone and growth hormone act as anti-inflammatory factors improving sensory recovery in female rats with thoracic spinal cord injury. Front Neurosci 17:1164044
Méndez M, Méndez-López M, López L, Begega A, Ángeles Aller M, Arias J, Arias JL (2010) Reversal learning impairment and alterations in the prefrontal cortex and the hippocampus in a model of portosystemic hepatic encephalopathy. Acta Neurol Belg 110:246–254
Millar RP (2005) GnRHs and GnRH receptors. Anim Reprod Sci 88:5–28
Monfort P, Erceg S, Piedrafita B, Llansola M, Felipo V (2007) Chronic liver failure in rats impairs glutamatergic synaptic transmission and long-term potentiation in hippocampus and learning ability. Eur J Neurosci 25:2103–2111
Monfort P, Cauli O, Montoliu C, Rodrigo R, Llansola M, Piedrafita B, el Mlili N, Boix J, Agustí A, Felipo V (2009) Mechanisms of cognitive alterations in hyperammonemia and hepatic encephalopathy: therapeutical implications. Neurochem Int 55:106–112
Montoya-García R, Fernández-Vargas V, Albor-Martínez KN, Martínez-Martínez A, Hernández-Jasso I, Quintanar-Stephano A, Calderón-Vallejo D, Quintanar JL (2023) Analysis of hippocampus in rats with acute brain ischemia-reperfusion injury treated with leuprolide acetate, an agonist of GnRH. Restor Neurol Neurosci 41:1–7
Muñoz MD, Monfort P, Gaztelu JM, Felipo V (2000) Hyperammonemia impairs NMDA receptor-dependent long-term potentiation in the CA1 of rat hippocampus in vitro. Neurochem Res 25:437–441
Navarro-Gonzalez YD, Ventura-Juarez J, Muñoz-Ortega MH, González-Blas D, Calvillo-Robedo A, Avila-Blanco ME, Valdez-Urias F, Quintanar-Stephano A (2023) Arginine vasopressin deficiency and conivaptan (a V1a–V2 receptor antagonist) treatment reverses liver damage and fibrosis in rats with chronic portocaval anastomosis. Int J Exp Pathol 104:1–14
Ochoa-Sanchez R, Tamnanloo F, Rose CF (2021) Hepatic encephalopathy: from metabolic to neurodegenerative. Neurochem Res 46:2612–2625
Paxinos G, Watson C (2006) The rat brain in stereotaxic coordinates. Academic Press, Cambridge
Pekny M, Pekna M, Messing A, Steinhäuser C, Lee JM, Parpura V, Hol EM, Sofroniew MV, Verkhratsky A (2016) Astrocytes: a central element in neurological diseases. Acta Neuropathol 131:323–345
Porseva VV, Smirnova VP, Korzina MB, Emanuilov AI, Masliukov PM (2013) Age-associated changes in sympathetic neurons containing neurofilament 200 kDa during chemical deafferentation. Bull Exp Biol Med 155:268–271
Quintanar JL, Salinas E (2008) Neurotrophic effects of GnRH on neurite outgrowth and neurofilament protein expression in cultured cerebral cortical neurons of rat embryos. Neurochem Res 33:1051–1056
Quintanar JL, Calderón-Vallejo D, Hernández-Jasso I (2016) Effects of GnRH on neurite outgrowth, neurofilament and spinophilin proteins expression in cultured spinal cord neurons of rat embryos. Neurochem Res 41:2693–2698
Schang AL, Ngô-Muller V, Bleux C, Granger A, Chenut MC, Loudes C, Magre S, Counis R, Cohen-Tannoudji J, Laverrière JN (2011) GnRH receptor gene expression in the developing rat hippocampus: transcriptional regulation and potential roles in neuronal plasticity. Endocrinology 152:568–580
Tahamtan M, Aghaei I, Pooladvand V, Sheibani V, Khaksari M, Shabani M (2017) Characterization of the CA1 pyramidal neurons in rat model of hepatic cirrhosis: insights into their electrophysiological properties. Metab Brain Dis 32:881–889
Tallis S, Caltana LR, Souto PA, Delfante AE, Lago NR, Brusco A, Perazzo JC (2014) Changes in CNS cells in hyperammonemic portal hypertensive rats. J Neurochem 128:431–444
Tronel S, Fabre A, Charrier V, Oliet SHR, Gage FH, Abrous DN (2010) Spatial learning sculpts the dendritic arbor of adult-born hippocampal neurons. Proc Natl Acad Sci U S A 107:7963–7968
Vázquez-Martínez O, Valente-Godínez H, Quintanar-Stephano A, Gasca-Martínez D, López-Cervantes ML, Palma-Tirado L, De Jesús Guerrero-Carrillo M, Pérez-Solís M, Díaz-Muñoz M (2019) Reduced liver lipid peroxidation in subcellular fractions is associated with a hypometabolic state in rats with portacaval anastomosis. Oxid Med Cell Longev 2019:4565238
Wilhelmsson U, Faiz M, De Pablo Y, Sjöqvist M, Andersson D, Widestrand Å, Potokar M, Stenovec M, Smith PLP, Shinjyo N, Pekny T, Zorec R, Ståhlberg A, Pekna M, Sahlgren C, Pekny M (2012) Astrocytes negatively regulate neurogenesis through the Jagged1-mediated notch pathway. Stem Cells 30:2320–2329
Yuan A, Rao MV, Veeranna, Nixon RA (2017) Neurofilaments and neurofilament proteins in health and disease. Cold Spring Harb Perspect Biol 9(4):a018309
Acknowledgements
We thank the MVZ Karen Estefany Sánchez-Hernández, from the Autonomous University of Aguascalientes, for providing the animals for this study, Dr. Ma. Consolación Martínez Saldaña and AQB Sonia Sofía Cruz Muñoz for the support in the histological technique. We also thank Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCYT) for the support in the scholarship 813,968 for doctoral student Brenda Lizeth Gutierrez Esparza.
Funding
This work was supported by the doctoral scholarship 813,968 from Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCYT).
Author information
Authors and Affiliations
Contributions
B.L.G.E participated in material preparation, analysis, interpretation of data, and drafting the work. M.L.G.T, A.Q.S. and J.L.Q. participated in design of the work, analysis, and interpretation of data, drafting, and revising the manuscript.
Corresponding author
Ethics declarations
Ethics approval
This study was reviewed and approved by the Animal Ethics and Welfare Committee of Autonomous University of Aguascalientes (CEADI/UAA/0025/18). All efforts were made to minimize the number of animals used and their suffering during the study in accordance with the Guide for the Care and Use of Laboratory Animals.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gutiérrez-Esparza, B.L., González-Torres, M.L., Quintanar-Stephano, A. et al. Neurological recovery in rats with portocaval anastomosis-induced hepatic encephalopathy treated with leuprolide acetate, a GnRH agonist. Metab Brain Dis (2024). https://doi.org/10.1007/s11011-024-01413-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11011-024-01413-9