
Abstract
Context
With accelerated land-use change throughout the world, increased understanding of the relative effects of landscape composition and configuration on biological system and bioinvasion in particular, is needed to design effective management strategies. However, this topic is poorly understood in part because empirical studies often fail to account for large gradients of habitat complexity and offer insufficient or even no replication across habitats.
Objectives
The aim of this study was to disentangle the independent and interactive effects of landscape composition and landscape configuration on the establishment and spread of invasive insect species.
Methods
We explore a spatially-explicit, mechanistic modeling framework that allows for systematic investigation of the impact of changes in landscape composition and landscape configuration on establishment and spread of invasive insect species. Landscape metrics are used as an indicators of invasive insect establishment and spread.
Results
We showed that the presence of an Allee effect leads to a balance between the effectiveness of spread and invasion success. Spread is maximized at an intermediate dispersal level and inhibited at both low and high levels of dispersal. The landscape, by either increasing or mitigating the dispersal abilities of a species, can lead to a rate of spread under a dispersal threshold for which density and spread is at the highest.
Conclusion
Our study proposes that change in landscape structure is an additional explanation of the highly variable spread dynamics observed in natural and anthropogenic landscapes. Consequently, a landscape-scale perspective could significantly improve spread risk assessment and the design of control or containment strategies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Throughout the world, habitat fragmentation and land-use change are among the most critical threats to biodiversity and ecosystems services (Millennium Ecosystem Assessment 2005; Cardinale et al. 2012). Both processes result in highly heterogeneous landscapes exhibiting differing composition (habitat types and proportion) and configuration (spatial arrangement of habitat types) (Fahrig et al. 2011). Advances in theoretical and empirical approaches to the analysis of range expansion of invasive species have highlighted the importance of such spatial heterogeneity as a critical factor that can influence the invasion process (e.g. Hastings et al. 2005; Bradley 2010; Vilà and Ibáñez 2011; Betts et al. 2014). By interacting with the distribution of invasive species, levels of spatial heterogeneity can either promote the establishment and spread of such species (With 2002; La Morgia et al. 2011; Thies et al. 2011) or, alternatively, can act as a barrier to spread (Jules et al. 2002; Mundt et al. 2011) or host colonization (Zhang and Schlyter 2004). However, it is not fully understood why some new populations spread rapidly across the landscape while others spread slowly or not at all. Whether the variability observed in nature or in experimental ensembles might be accounted for by systematic differences between landscape structures remains an open research question (Richter et al. 2013; Sutherland et al. 2013; Meier et al. 2014).
Research on landscape-level effects on the establishment and spread of invasive species has focused predominantly on the contribution of natural and semi-natural habitat types surrounding urban, forest and agricultural ecosystems. Human based land-use promotes habitat disturbance and human assisted dispersal that can increase propagule pressure and promote the successful establishment and spread of an invading species (Pyšek and Richardson 2010; González-Moreno et al. 2013). For example, the urban landscape supports a diverse fauna ranging from native species to opportunistic species (both native and invasive) which exploit modified habitats (Garden et al. 2006). In contrast, modern methods in agricultural and plantation forestry ecosystems, have generated monocultures that are continuous in their extent and dominated by few crop species, leading to homogenized landscapes (Margosian et al. 2009). Landscape simplification increases the concentration of suitable resources that are available to particular invasive species (Tscharntke et al. 2012; Rigot et al. 2014; Jonsson et al. 2015) and can reduce the number of habitat types that support invasive species’ natural enemies (Chaplin-Kramer et al. 2011; Cardinale et al. 2012). Furthermore, the fragmentation of wild-land habitat resulting from agricultural and urban development also affects the spread of invasive species (Harper et al. 2005; La Morgia et al. 2011; González-Moreno et al. 2014). Urban forests and parklands represent an increasing percentage of our remaining near-natural habitats. Because of their proximity to sites of introduction and their (often) large ratio of edge to interior habitat, they are prime habitat for introduced plant and animal species which can then spread into less urbanised areas (Martin et al. 2008). Landscape disturbance features such as hiking trails or roads can also promote invasion spread, in part by creating disturbed edges where invasive species can easily establish (Christen and Matlack 2006).
Synthesis of research across multiple taxa and habitat types offers strong evidence for landscape composition shaping the establishment and spread of an invasive species (e.g. Tscharntke et al. 2012; Robledo-Arnuncio et al. 2014). Empirical studies are strongly supported by simulation models showing that the local density of invasive species is determined not only by local habitat quality, but also by the spatial structure of the habitat in the surrounding landscape (With 2002, 2004). Understanding the effect of the existing distribution of suitable/unsuitable habitat on the establishment and spread of invasive species is therefore critical for planning strategies to eradicate or contain an invasive species. However, the outcome of such spatial analysis remains limited by our ability to carry out a quantitative synthesis of landscape effects on the spread of a species. In particular, empirical studies often fail to account for large gradients of habitat complexity (e.g. patch size, shape, juxtaposition, inter-patch connectivity or habitat aggregation) and offer insufficient or even no replication across habitat. This deficiency is in part because of an unavoidable trade-off between spatial scope, sampling intensity and accuracy (Robledo-Arnuncio et al. 2014).
Spatially-explicit models, on the other hand, have provided a good basis for generating replicated landscape patterns with partially controllable spatial properties (With 2004). These models traditionally focus on the effect of a limited number of landscape attributes, such as habitat density or patch connectivity (With 2002; Sebert-Cuvillier et al. 2008; Smith et al. 2014; Morel-Journel et al. 2015), where connectivity is classically defined as the degree to which landscape features facilitate or impede the rate of movement of species between habitat patches. Depending on the species considered, these different landscape features have been shown to modify dispersal rates between habitat patches (Calabrese and Fagan 2004) or the pattern of dispersion (Jonsen and Taylor 2000). Theoretical studies, such as that by Hanski and Gaggiotti (2004) suggest that as connectivity increases, it facilitates the formation of a metapopulation which can increase the persistence of local populations through source-sink dynamics. On the other hand, Smith et al. (2014) and Morel-Journel et al. (2015) suggested that increasing connectivity during dispersal allows more effective spread, but simultaneously can decrease population density at the source, which can accentuate demographic stochasticity and Allee effects. Although there is a substantial literature on demographic analysis of invasive populations that helps shed light on specific life-history traits contributing to invasion, only a minority of spatially-explicit models pose demographic processes as essential to a general understanding of the invasion process. Nevertheless, many studies report that the magnitude and direction of landscape effects on each species depends on that species life-history sensitivities to environmental disturbance as well as it’s dispersal abilities (Guisan and Thuiller 2005; Huntley et al. 2010; Schurr et al. 2012; Robledo-Arnuncio et al. 2014). Clearly, there is a need for more integrative approaches that link demographic processes and dispersal strategies to the landscape to better understand the consequences of increasing anthropogenically driven land-use change, and global environmental and climatic change on species spread (Bocedi et al. 2014; Robledo-Arnuncio et al. 2014; Wang and Jackson 2014).
The primary aim of this study was to disentangle the independent and interactive effects of landscape composition and landscape configuration on the establishment and spread of invasive insect species. Critical to this effort is the development of a spatially-explicit, mechanistic model to forecast the spread of species in relation to landscape structure. We use the available modelling platform, MDiG (Pitt 2008; Pitt et al. 2009) to examine how the rate of range expansion across patchy landscapes is determined by the amount and spatial patterns of suitable habitat. The initial parameters of the model were based on the natural history of the European gypsy moth (Lymantria dispar). The species is native to the temperate forests of Europe and is a notorious insect defoliator, occasionally causing extensive tree mortality. A European strain was accidentally introduced to North America near Boston, USA in 1869 and has subsequently invaded much of the susceptible forest of north-eastern America (Tobin and Blackburn 2007). As part of the campaign to slow the European gypsy moth spread across the United States, intensive monitoring efforts have been focused around the invasion front. As a result, the spread of the European gypsy moth across North America is, perhaps, the most thoroughly studied biological invasion and offers unique insight into processes such as Allee effects in time and space, and comprehensive investigations of the interactions between individual movement to mate finding and links between landscape structure and movement (Tobin et al. 2015). We first tested the relative importance of the quantity of suitable habitat, its configuration, as well as their interaction, as predictors of patterns of density and rate of spread of invasive species. We then identified a generic core of landscape features that significantly affect biological invasion success. Our study proposes that change in landscape structure is an additional explanation of the highly variable spread dynamics observed in natural and anthropogenic landscapes. Consequently, a landscape-scale perspective could significantly improve spread risk assessment and the design of control or containment strategies.
Material and methods
A spatially-explicit, stochastic spread model
The central part of the modelling framework is provided by a spatially-explicit, mechanistic model, MDiG, that represents current understanding of the ecology of invasive insect spread and establishment (Pitt 2008; Pitt et al. 2009). The initial parameters of the model were based on the natural history of the European gypsy moth (Lymantria dispar). These parameters have been used in several studies considering European gypsy moth population dynamics and spread in complex patch-matrix landscapes (With 2002; Liebhold and Bascompte 2003; Nesslage et al. 2007), including the MDIG framework (Lustig et al. 2017). A sensitivity analysis evaluating the influence of slight changes to these base parameters was then used to examined individual and joint effects of parameters and ranked parameters based on their influence on the dynamics of simulated spread. Therefore the range of parameters tested in the present study are not limited to the European gypsy moth but include those for other insect species and sub-species, including for example, the Asian and Japanese gypsy moths.
To initialize a simulation, individuals were located on a square lattice (simulation arena) comprised of suitable and unsuitable habitat types. Following Johnson et al. (2006) and Liebhold and Bascompte (2003), the local population density (density per raster cell) was approximated by a deterministic Allee logistic growth model:
where C is the Allee threshold, r the intrinsic growth rate and K the carrying capacity. The values of these parameters were based on previous estimates reported in Liebhold and Bascompte (2003) and Johnson et al. (2006)from capture-release data collected from 1988 to 2004 at the invasion front (Table 1).
Spatially-explicit rules determine the pattern of local dispersal and generic long distance dispersal spread. For every occupied raster cell (or habitat patch), a proportion of the individuals spread evenly to the surrounding patches. In addition, a Poisson process is used to generate how many long distance dispersal events originate from each habitat patch (Pitt 2008; Lustig et al. 2017). The frequency of these long-distance dispersal events correspond to the mean of the underlying Poisson distribution and was set to 0.05. The distance travelled by the individuals is approximated by a Cauchy probability distribution as it allows for long, rare dispersal distances which may occur naturally (wind dispersal for example) or which may represent human assisted dispersal (Hastings et al. 2005). Following Johnson et al. (2006), we fixed the median dispersal distances of long-distance dispersal events (median of the Cauchy distribution) at five raster cells. The direction of dispersal was sampled from a uniform distribution (Pitt 2008).
Generating and quantifying landscape heterogeneity
We generated binary (suitable, unsuitable habitat) landscape patterns for 10 km cell size raster grids of extents \(1280 \times 1280\) km\(^2\) to approximate local movements of larvae and male adults gypsy moth (Jankovic and Petrovskii 2013). We used the computer program Qrule 4.2 (Gardner 1999) to generate a gradient of landscape complexity, in which fragmentation (measured as the degree of spatial autocorrelation) and proportion of habitat cover can be systematically and independently controlled (Gardner and Urban 2007). Qrule uses a midpoint displacement algorithm (Saupe 1988) to generate multi-fractal maps in which the degree of spatial autocorrelation among adjacent cells (H) can be controlled. We generated landscape patterns in a full factorial design across a five-steps gradient in spatial autocorrelation [\(H \in (0.1,1)\) in increments of 0.2, 0 being close to random and 1 being completely clustered] and a six-steps gradient in proportion of suitable habitat habitat cover [\(P \in (5,95\%)\) in 10% increments] with 10 replicate landscapes for each of the 35 factor combinations (Table 1; Lustig et al. 2015).
For each sample landscape, we calculated landscape metrics using the computer program FRAGSTATS (McGarigal et al. 2012). These metrics provide simple quantitative measurements of the composition and configuration of a landscape. The landscape metrics were defined for suitable habitat cover only and are commonly referred to as class-level metrics. McGarigal et al. (2012) categorized these metrics into five groups corresponding to the aspect of landscape structure emphasized. These include patch area/density/edge, shape, aggregation, contrast, core, isolation/proximity and connectivity metrics (Table 1). In addition, we included the percentage of the suitable habitat (PLAND), considered the most universal measure of landscape composition, to enable a comparison between the relative effect of landscape configuration, landscape composition and their potential interactions.
Sensitivity analysis
Insect species can be placed along a ‘slow-fast life-history continuum’, where changes in population growth rate arise mainly from variability in reproductive rates in fast species and in survival rates for slow species (Herrando-Pérez et al. 2012). The position of a species along this continuum could also reflect the propensity of a species to invade a new environment. Similarly, changes in dispersal distances, frequencies of dispersal events, carrying capacity and Allee threshold are also expected among species. We used MdiG to broadly capture this variation, by creating nine different combinations of species’ life-history attributes and dispersal abilities in a mono-factorial design (Table 1). The main reason for doing this was to evaluate how change in the intrinsic growth rate, carrying capacity, Allee effect threshold, and the frequency and median distance of long-distance dispersal events affect the dynamics of spread, and to quantify a general relationship across life-history attributes, with changes to landscape variables. We varied each parameter over three values (Table 1) and studied the effects of these changes on the dynamics of the simulated invasion. For each landscape, we allowed the species to expand its range for 30 years, one model time step represents one year, and we replicated the simulations 25 times to account for dispersal stochasticity. We recorded four response variables over time: the population density d (number of individuals in the population), the rate of spread—ROS (number of new cells occupied per simulation run), the average dispersal distance—avDist, and the maximum dispersal distance—maxDist.
Statistical analysis
To investigate the role of landscape metrics as indicators of invasive insect spread, we used a multi-model inference framework (Burnham and Anderson 2002). A multi-model analysis provides a means to simultaneously compare multiple regression models, to evaluate how well each is supported by data, and potentially to identify the best supported model(s). This approach accounts for the uncertainty associated with a single model by providing confidence interval coverage of variable estimates. Statistical analyses were performed using the R statistical system v 3.2.2 (R Development Core Team 2013).
We used generalized linear models (GLMs) with a Gaussian error distribution. Four response variables were considered: the population density—d, the rate of spread—ROS, the average dispersal distance—avDist, and the maximum dispersal distance—maxDist, respectively. The predictor variables included 84 metrics of landscape configuration and composition (Table 1). Prior to conducting the multivariate regression, we reduced the collinearity among landscape metrics by selecting independent groups of metrics identified in Lustig et al. (2015). We further assessed the degree of mulicollineraty among predictor variables in each group by calculating the generalized variance inflation factor (GVIF) for all predictors (Dormann et al. 2013). Predictors giving high GVIF were identified by calculating the pair-wise correlation between all predictors using a non-parametric Spearman’s rank correlation. We removed highly correlated variables from the analysis until all GVIF values were smaller than 10 (Table 2).
We then performed a multimodel regression model procedure based on the Akaike Information Criterion (AICc) with a correction for finite sample sizes (Burnham and Anderson 2002. Landscape metrics were mean-centred to facilitate the interpretation of the main effect and the outputs were log transformed to improve the normal distribution of each model’s residuals. First, we identified the landscape variables that significantly improved model performance (lowest AICc values) such that they individually accounted for 95% of the cumulative sum of Akaike weights (Akaike weights represent the likelihood of a model) were retained. Second, we calculated the average parameter estimates of the model, and their 95% confidence intervals, using a model averaging algorithm. The relative importance of each landscape metric as a predictor of invasive spread was determined, based on the sum of Akaike weights, with 1 being the most important (the metric variable is present in all model candidates for each species scenario) and 0 the least important. Possible predictor variables were considered to contribute to model performance improvement if their AICc weight summation was relatively high (\(w>0.7\)). Model selection for mixed models was conducted using ‘lme4’ package (Bates et al., 2015) and ‘MuMIn’ package for model-averaging of coefficients (Bartoń 2016).
Results
Aspects of landscape structure influencing the spread of invasive insects
The average population density, the rate of spread, the mean dispersal distance and the maximum dispersal distance were strongly correlated to three landscape metrics: proportion of suitable habitat in the landscape (PLAND)—the most common measure of habitat composition; edge density (ED)—total length of borders between habitat patches in landscape, and normalized shape index (nLSI)—a simple measure of habitat patch shape and clumpiness. These three landscape metrics collectively represent a gradient from areas with few, small and clustered habitat patches towards an area with high inter-dispersion and covered by a large amount of habitat with elongated patches. The effect of the proportion of suitable habitat in the landscape (PLAND) was the most important variable affecting all aspects of spread (Table 3). Increase in density, rate of spread, mean dispersal distance and maximum dispersal distance were all positively related to proportion of suitable habitat in the landscape, while these four components of spread were negatively related to the normalized shape index. An increase in population density, rate of spread and maximum dispersal distance were positively related to edge density (ED), while average dispersal distance was negatively related to edge density.
The percentage of like adjacencies (PLADJ)—which shows the frequency with which different pairs of habitat patches appear side-by-side in the landscape, and the splitting index (SPLIT)—which characterize the disconnection between suitable habitat patches, significantly affected population density and rate of spread. In addition, the mean circumscribing circle (CIRCLE_AM)—a measure of patch elongation, the connectivity index (CONNECT), and the amount of disjunct core areas (DCDA) also had a significant effect on population density and rate of spread. Although the effects were significant, the actual effect size of the last three landscape metrics on population density were small, with lower summed Akaike weights (0.01, 0.01 and 0.02 respectively) and model partial slope coefficient near 0 (Table 3).
The core metrics, in particular, the median and standard deviation of the disjunct patch area, DCORE_MD and DCORE_SD, only showed a significant effect on the mean dispersal distance (avDist) and the maximum dispersal distance (maxDist). A number of shape metrics (FRAC_CV, SHAPE_MD, PARA_MD), core metrics (CAI_CV and DCAD) and aggregation/connectivity metrics (CONNECT and SPLIT) had a significant effect on dispersal distance. However, the effect size of these configuration metrics were relatively small as shown by lower summed Akaike weights and a model partial slope coefficient near 0 (Table 3).
Furthermore, we found significant interactions between the proportion of suitable habitat in the landscape (PLAND) and landscape configuration metrics, which were stronger and better supported for rate of spread and dispersal distances than for population density (Table 3). The average influence of the proportion of suitable habitat (PLAND) on rate of spread, dispersal distance and population density decreased when the degree of fragmentation in the landscape (SPLIT) and edge density (ED) increased. In contrast, the average influence of the proportion of high-quality habitat (PLAND) on rate of spread and dispersal distances was enhanced when the related circumscribing circle (CIRCLE_AM)—a measure of patch elongation, increased. These interactions predicted that the marginal increase of the proportion of suitable habitat within a landscape is on average less when the habitat is highly fragmented.
We also found significant effects of the interactions between the proportion of high-quality habitat (PLAND) and core metrics (DCORE_MD, DCORE_SD, CAI_CV and DCAD) on the maximum dispersal distance, and between the proportion of high-quality habitat (PLAND) and measure of patch aggregation (PLADJ) on the rate of spread. However, the effect of these latter interactions were relatively weak, with lower summed Akaike weights (Table 3).
Balancing effectiveness of spread and invasion success

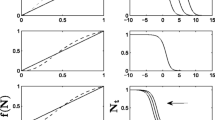
Balancing effectiveness of spread and invasion success in heterogeneous landscapes. Results are shown for all parameter values tested. The top panels show how population density (d) and rate of spread (ROS) respond to different ranges of mean dispersal distances for three different scenarios: without Allee effect, with weak Allee effect and with strong Allee effect. Above a dispersal threshold (red dotted line), growth and rate of spread were considerably reduced or not observed. The bottom panel represents the probability of observing a mean dispersal distance higher than the dispersal threshold that can limit growth and spread as a function of the proportion of suitable habitat (PLAND)
When an Allee effect was absent, an increase in the proportion of suitable habitat (PLAND) resulted to an increase in the mean dispersal distances, and subsequently to an increase in the rate of spread and population density (Fig. 1).
In the presence of an Allee effect, the range of dispersal distances that allow for optimal spread was considerably reduced compared to the case without an Allee effect. At lower realised mean dispersal distances (avDist), spread was limited as evidenced by a lower realized mean rate of spread, but population density increased as indicated by a local maxima in the population density curve at an average mean dispersal of 0.2 m (Fig. 1). A two-fold increase in the mean dispersal distance leads to the highest rate of spread and population density. The range of permissible rate of spread and population density was dramatically reduced when the mean dispersal distance went over a certain dispersal threshold (Fig. 1). These results suggested that invasive insect species subjected to an Allee effect may present two different growth patterns when arriving to a new environment. On one hand, species that have limited dispersal abilities will maximise their populations locally but will fail to establish a population over a large area. On the other hand, species that have high dispersal abilities may spread but face the added risk of not establishing or going extinct.
The results also showed that an increase in the proportion of suitable habitat (PLAND) in the landscape resulted in an increase in the probability of observing a mean dispersal distance higher than the dispersal threshold that can limit growth and spread (Fig. 1). In a landscape with more than 70% of suitable habitat, 5% of the simulations resulted in a mean dispersal distance higher than the dispersal threshold. In a landscape with more than 80% of suitable habitat, 10% of the simulations resulted in a mean dispersal distance higher than this dispersal threshold. Although a high proportion of suitable habitat would lead to more successful spread (highest mean dispersal distances) when the source population density is high enough, it may also reduce spread success and population growth when the source population is too small. This result provides new insights into the proportion of suitable habitat as an additional explanation to the highly variable spread rate observed in both natural and anthropogenic landscapes.
Discussion
The influence of landscape structure on the spread of invasive insect species is generally acknowledged but is often difficult to quantify. In this study, we investigated the effect of landscape structure as a predictor of patterns of spread using the European gypsy moth as a model invasive insect species. We found that both landscape composition and landscape configuration influenced population density and rate of spread in significant and sometimes interactive ways.
Landscape metrics as indicators of spread
We showed that the density and rate of spread of invasive insects are more impacted by the proportion of suitable habitat in the landscape than by habitat configuration. This has significant implications for the prediction and management of invasive insect species in general. In all species scenarios studied here, the increase in density and rate of spread of the species was positively related to an increasing proportion of suitable habitat in the landscape. The direct effect of the amount of suitable habitat on invasive insect species density and spread has been reported in several studies, particularly in agroecosystems (Chaplin-Kramer et al. 2011). Our results suggest that such an effect may be caused by a greater aggregation of suitable habitat patches (as measured by nLSI, PLADJ, SPLIT) and a slight decrease in edge density. Different spread rates in different landscapes can also be caused by variation in connectivity of habitat patches, in habitat patch elongation and by variation in the ratio of edge-to-interior habitat. In other words, landscapes containing larger patches of suitable habitat with simple, circular or square shape are expected to be more conducive to the rapid spread of invasive insects. Previous research also confirms that the metrics identified in this study that characterize habitat density (With 2002; Sebert-Cuvillier et al. 2008; Tscharntke et al. 2012), connectivity [e.g. (With 2002; Sebert-Cuvillier et al. 2008; Wang et al. 2011; Morel-Journel et al. 2015)], path aggregation (With and King 1999b; Wang et al. 2011) and edge density (Radeloff et al. 2000; With 2002; Tscharntke et al. 2012) are important aspects of the landscape that can either influence or exacerbate the spread of insects.
Overall, we found that the main effect of the configuration of habitats at a landscape scale had less impact on total density and rate of spread of the invasive insect species when compared to the main effect of the amount of habitat. Such a finding is consistent with and confirms the finding of many studies that habitat homogenisation is one of the key drivers of global increase in pest and disease establishment and spread (Jonsson et al. 2015). Nevertheless, we recognized that population density and rate of spread of invasive insect species can be significantly affected by complex interactions between the composition and configuration of the landscape in the introduction area.
Fragmentation versus proportion of suitable habitat in the landscape
We showed that the variation in density and spread of invasive insects was best explained by an interaction between the proportion of suitable habitat in the landscape (PLAND) and the splitting index. Moreover, we found evidence of a significant interaction between the proportion of suitable habitat in the landscape and respectively, the effects of edge density and average patch elongation on rate of spread and dispersal distances.
Habitat split is a measure of habitat fragmentation designed to quantify the extent to which habitat patches are disconnected in space (Jaeger 2000). Our results contrast with theoretical studies that suggest that suitable habitat connectivity at the introduction site is a determining factor of patterns of spread (Morel-Journel et al. 2015). On the contrary, we found that invasive spread might be limited more by the size and distribution of non-habitat gaps than by a measure of habitat connectivity (rate of movement between habitat patches). Ultimately, the notion of patch aggregation is embedded within the notion of connectivity such that, when habitat split decreases, connectivity also increases. Our results are in agreement with the results of With and King (1999a) and Wang et al. (2011) who also highlighted the critical role of non-habitat gaps in reducing the spread of species over the landscape.
On the other hand, the effect of edge density and habitat patch elongation was particularly challenging to assess. In this study, a decrease in edge density led to an increase in population density and rate of spread of the gypsy moth, but a slight decrease in density. Other studies have found that depending on whether species are moving primarily within or between habitat types, the micro-environments between habitat fragments may critically increase (Radeloff et al. 2000; With 2002; Tscharntke et al. 2012) or decrease (Tscharntke et al. 2012) the potential of spread. Ewers et al. (2007) further demonstrated that an increase in habitat shape complexity is associated with an increase in the amount of edge habitat that results in a higher edge density. Large patches are more likely to be of a complex shape. We found that as shape complexity increases, patches become less circular, and the probability of population increase and spread of an invasive insect species decreases.
Overall, the results demonstrated that population increase and spread of invasive insect species can be significantly affected by the combined landscape based components: suitable habitat amount, habitat patch aggregation and habitat shape complexity. We found that population density and rate of spread were positively correlated with landscape metrics that increase with decreasing habitat fragmentation, and negatively correlated with metrics which increase with increasing habitat fragmentation. To our knowledge, most theoretical studies that have focused on the spread of invasive insect species at the colonization front have also found a non-negligible effect of the spatial arrangement of habitat on local density and rate of spread (With 2002; Morel-Journel et al. 2015). However, Fahrig et al. (2011) and Tscharntke et al. (2012) proposed that species respond idiosyncratically to habitat fragmentation versus habitat loss, which may suggest that highly mobile species can tolerate habitat fragmentation as long as the amount of total habitat is sufficient. Their finding suggests that the configuration of the landscape might not always be important for predicting the density and rate of spread of invasive insect species.
Impact of landscape structure on population dynamics
Clearly, the overall outcome of spread critically depends on several life-history characteristics and species dispersal ability as well as landscape factors. As reflected by Fig. 1, our results support two important hypothesis about population spread and growth in a heterogeneous environment. First, in the absence of an Allee effect, and in the absence of other influences (predation, for example), a population can spread and grow regardless of the proportion of suitable habitat. Increasing the proportion of suitable habitat in the landscape increases the overall population density and rate of spread exponentially (Fig. 1; Shigesada and Kawasaki (1997)) Second, in the presence of an Allee effect, a population experiences a balancing effect between the effectiveness of spread and the establishment of a persistent population (Fig. 1; Smith et al. (2014)): population growth and spread are maximized at intermediate dispersal abilities but inhibited at high dispersal abilities. Species that have a high intrinsic rate of increase quickly exceed the size of population that is vulnerable to Allee effects, ensuring a higher probability of persistence of the introduced propagule(s) (Morel-Journel et al. 2015).
Furthermore, Smith et al. (2014) used a programmed Allee effect in bacteria to demonstrate that an increase in the number of suitable habitat patches can result in more prolific spread but simultaneously increases the chances that each population could fail to establish due to Allee effect. Our simulations based on insect life-history parameters are in broad agreement with these results. We found evidence of a dispersal threshold above which population spread and growth can be inhibited. Additionally, the results of this study echo the ‘all or nothing’ strategy proposed by Heimpel and Asplen (2011) and Morel-Journel et al. (2015), where they propose that a population would either go extinct quickly or form a stable population expanding across highly connected landscape. Therefore, species with an Allee effect follow idiosyncratic dynamics of spread that can be significantly affected by the structure of the landscape. This study further revealed that by either allowing or preventing the dispersal of a species from the introduction site (or effective containment) complex interactions between the composition or configuration of the landscape may push these species into a range of dispersal that allows optimal spread and growth. Specifically, the results in this study showed that in a landscape with more than 70% of suitable habitat, 5% of the simulations resulted in a mean dispersal distance higher than a dispersal threshold that can limit growth and spread. Such an observation may offer an additional explanation to the highly variable spread dynamics observed in both natural and anthropogenic landscapes. It can also explain why, during the early stages of an outbreak or incursion, spread can be initially slow manifesting itself as a time lag followed by an increase over time. Species must establish a population density that can exceed the size vulnerable to Allee effects before effective spread will occur.
There is a lively current debate about the importance of investing pest-management resources into reducing habitat quality to limit the dispersal abilities of an invasive organism, for example by decreasing the extent of suitable habitat or managing the spatial aggregation of habitat patches across the landscape (With 2004). Such habitat manipulation is consistent with the management practice of creating “barrier zones” at invasion fronts where suppression activities (poisoning/trapping) are employed to create areas of fragmented habitat that disrupt the movement of organisms across the landscape and slow the rate of expansion. Indeed, such a technique was used by the U.S Forest Service to control the spread of the European gypsy moth across North America (Sharov and Liebhold 1998; With 2004). However, Nesslage et al. (2007) reported a lack of a typical threshold response to a declining proportion of habitat, using empirical data on the European gypsy moth invasion wave front across Michigan (1985–1996). Nesslage et al. (2007) further suggested that the concept of creating areas of fragmented habitat as “barrier zones” to slow the invasion wavefront may be of limited use for good dispersers like the European gypsy moth. Using simulation models in this present study, we also found that the rate of spread and population density of several invasive insect species did not exhibit a typical threshold response to declining proportion of habitat. Instead, in situations where the proportion of suitable habitat in the landscape is already above a critical threshold for species dispersal and persistence, our results suggested that further manipulation of the habitat may push these species into a range of dispersal that allows optimal spread. In addition, we found dispersers are likely to be influenced not only by the proportion of habitat in the landscape but also by the spatial aggregation and structural connectivity of habitat patches. Such results create challenges for designing management strategies focused on reducing the spread of invasive insects but that also work across species with very different behaviours and in different landscapes. This is because the degree of habitat fragmentation and the proportion of habitat in the landscape can have very different impacts depending on the dispersal behaviour of the species.
Conclusion
This study offers new insights for management programs that aim to either eradicate or in the case that strategy fails, limit or contain the spread of an invasive insect species. Strong evidence was found for landscape composition and configuration shaping the successful establishment and rate of spread of a species. Adopting a strategy that systematically reduces the dispersal ability of a new introduced species by reducing dispersal between suitable habitat patches, for example (Brown et al. 2006), might inadvertently help them thrive by pushing the species into a range of dispersal that allows optimal spread and growth. We therefore recommend that management strategies for containment or eradication of invasive insect species set spatial priorities of control at the landscape scale. These spatial priorities should account for specific landscape characteristics, the stage of the invasion and the life-history characteristics of the species. The analysis in this study provides a basis for the development of a core set of structural landscape metrics as indicators of invasive insect spread. However, optimal choice of metrics will require case specific exploration of their indicator values as there is no one-case-fits-all strategy that can ensure optimal risk assessment and management control for all species and landscapes.
References
Bartoń K (2016) MuMIn: multi-model inference, version 1.15.6. URL: https://cran.r-project.org/web/packages/MuMIn/index.html
Bates D, Maechler M, Bolker B, Walker S, Christensen RHB, Singmann H, Dai B, Eigen adn Grothendieck G (2015) lme4: linear mixed-effects models using ’Eigen’ and S4, version 1.1-10. https://cran.r-project.org/web/packages/lme4/index.html
Betts MG, Fahrig L, Hadley AS, Halstead KE, Bowman J, Robinson WD, Wiens JA, Lindenmayer DB (2014) A species-centered approach for uncovering generalities in organism responses to habitat loss and fragmentation. Ecography 37(6):517–527
Bocedi G, Palmer SC, Pe’er G, Heikkinen RK, Matsinos YG, Watts K, Travis JM (2014) Rangeshifter: a platform for modelling spatial eco-evolutionary dynamics and species’ responses to environmental changes. Methods Ecol Evol 5(4):388–396
Bradley BA (2010) Assessing ecosystem threats from global and regional change: hierarchical modeling of risk to sagebrush ecosystems from climate change, land use and invasive species in Nevada, USA. Ecography 33(1):198–208
Brown GP, Phillips BL, Webb JK, Shine R (2006) Toad on the road: use of roads as dispersal corridors by cane toads (Bufo marinus) at an invasion front in tropical Australia. Biol Conserv 133(1):88–94
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Calabrese JM, Fagan WF (2004) Lost in time, lonely, and single: reproductive asynchrony and the Allee effect. Am Nat 164(1):25–37
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP (2012) Biodiversity loss and its impact on humanity. Nature 486(7401):59–67
Chaplin-Kramer R, O’Rourke ME, Blitzer EJ, Kremen C (2011) A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol Lett 14(9):922–932
Christen D, Matlack G (2006) The role of roadsides in plant invasions: a demographic approach. Conserv Biol 20(2):385–391
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, Marquéz JRG, Gruber B, Lafourcade B, Leitão PJ (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36(1):27–46
Ewers RM, Thorpe S, Didham RK (2007) Synergistic interactions between edge and area effects in a heavily fragmented landscape. Ecology 88(1):96–106
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin JL (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14(2):101–112
Garden J, Mcalpine C, Peterson A, Jones D, Possingham H (2006) Review of the ecology of Australian urban fauna: a focus on spatially explicit processes. Aust Ecol 31(2):126–148
Gardner RH (1999) RULE: map generation and a spatial analysis program. Landscape ecological analysis. Springer, New York, pp 280–303
Gardner RH, Urban DL (2007) Neutral models for testing landscape hypotheses. Lands Ecol 22(1):15–29
González-Moreno P, Pino J, Gassó N, Vilá M (2013) Landscape context modulates alien plant invasion in Mediterranean forest edges. Biol Invas 15(3):547–557
González-Moreno P, Diez JM, Ibáñez I, Font X, Vilà M (2014) Plant invasions are context-dependent: multiscale effects of climate, human activity and habitat. Divers Distrib 20(6):720–731
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8(9):993–1009
Hanski IA, Gaggiotti OE (2004) Ecology, genetics and evolution of metapopulations. Elsevier, San Diego
Harper KA, Macdonald SE, Burton PJ, Chen J, Brosofske KD, Saunders SC, Euskirchen ES, Roberts D, Jaiteh MS, Esseen PA (2005) Edge influence on forest structure and composition in fragmented landscapes. Conserv Biol 19(3):768–782
Hastings A, Cuddington K, Davies KF, Dugaw CJ, Elmendorf S, Freestone A, Harrison S, Holland M, Lambrinos J, Malvadkar U (2005) The spatial spread of invasions: new developments in theory and evidence. Ecol Lett 8(1):91–101
Heimpel GE, Asplen MK (2011) A ‘Goldilocks’ hypothesis for dispersal of biological control agents. BioControl 56(4):441–450
Herrando-Pérez S, Delean S, Brook BW, Bradshaw CJ (2012) Density dependence: an ecological Tower of Babel. Oecologia 170(3):585–603
Huntley B, Barnard P, Altwegg R, Chambers L, Coetzee BW, Gibson L, Hockey PA, Hole DG, Midgley GF, Underhill LG, Willis SG (2010) Beyond bioclimatic envelopes: dynamic species’ range and abundance modelling in the context of climatic change. Ecography 33(3):621–626
Jaeger JA (2000) Landscape division, splitting index, and effective mesh size: new measures of landscape fragmentation. Lands Ecol 15(2):115–130
Jankovic M, Petrovskii S (2013) Gypsy moth invasion in North America: a simulation study of the spatial pattern and the rate of spread. Ecol Complex 14:132–144
Johnson DM, Liebhold AM, Tobin PC, Bjørnstad ON (2006) Allee effects and pulsed invasion by the gypsy moth. Nature 444(7117):361–363
Jonsen ID, Taylor PD (2000) Fine-scale movement behaviors of calopterygid damselflies are influenced by landscape structure: an experimental manipulation. Oikos 88(3):553–562
Jonsson M, Straub CS, Didham RK, Buckley HL, Case BS, Hale RJ, Gratton C, Wratten SD (2015) Experimental evidence that the effectiveness of conservation biological control depends on landscape complexity. J Appl Ecol 52(5):1274–1282
Jules ES, Kauffman MJ, Ritts WD, Carroll AL (2002) Spread of an invasive pathogen over a variable landscape: a nonnative root rot on Port Orford cedar. Ecology 83(11):3167–3181
La Morgia V, Malenotti E, Badino G, Bona F (2011) Where do we go from here? Dispersal simulations shed light on the role of landscape structure in determining animal redistribution after reintroduction. Lands Ecol 26(7):969–981
Liebhold A, Bascompte J (2003) The allee effect, stochastic dynamics and the eradication of alien species. Ecol Lett 6(2):133–140
Lustig A, Stouffer DB, Roigé M, Worner SP (2015) Towards more predictable and consistent landscape metrics across spatial scales. Ecol Indic 57:11–21
Lustig A, Worner SP, Pitt JPW, Doscher C, Stouffer DB, Senay SD (2017) A modelling framework for the establishment and spread of invasive species in heterogeneous environments. Ecol Evol 10.1002/ece3.2915
Margosian ML, Garrett KA, Hutchinson JS (2009) Connectivity of the American agricultural landscape: assessing the national risk of crop pest and disease spread. BioScience 59(2):141–151
Martin PH, Canham CD, Marks PL (2008) Why forests appear resistant to exotic plant invasions: intentional introductions, stand dynamics, and the role of shade tolerance. Front Ecol Environ 7(3):142–149
McGarigal K, Cushman S, Ene E (2012) FRAGSTATS v4: spatial pattern analysis program for categorical and continuous maps. http://www.umass.edu/landeco/research/fragstats/fragstats.html
Meier ES, Dullinger S, Zimmermann NE, Baumgartner D, Gattringer A, Hülber K (2014) Space matters when defining effective management for invasive plants. Divers Distrib 20(9):1029–1043
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being. Millennium ecosystem assessment. Island Press, Washington, DC
Morel-Journel T, Girod P, Mailleret L, Auguste A, Blin A, Vercken E (2015) The highs and lows of dispersal: how connectivity and initial population size jointly shape establishment dynamics in discrete landscapes. Oikos 0:01–009
Mundt CC, Sackett KE, Wallace LD (2011) Landscape heterogeneity and disease spread: experimental approaches with a plant pathogen. Ecol Appl 21(2):321–328
Nesslage GM, Maurer BA, Gage SH (2007) Gypsy moth response to landscape structure differs from neutral model predictions: implications for invasion monitoring. Biol Invas 9(5):585–595
Pitt JPW (2008) Modelling the spread of invasive species across heterogeneous landscapes. PhD thesis, Lincoln University, New Zealand
Pitt JPW, Worner SP, Suarez AV (2009) Predicting argentine ant spread over the heterogeneous landscape using a spatially explicit stochastic model. Ecol Appl 19(5):1176–1186
Pyšek P, Richardson DM (2010) Invasive species, environmental change and management, and health. Ann Rev Environ Res 35:25–55
Radeloff VC, Mladenoff DJ, Boyce MS (2000) The changing relation of landscape patterns and jack pine budworm populations during an outbreak. Oikos 90(3):417–430
Richter R, Dullinger S, Essl F, Leitner M, Vogl G (2013) How to account for habitat suitability in weed management programmes? Biol Invas 15(3):657–669
Rigot T, van Halder I, Jactel H (2014) Landscape diversity slows the spread of an invasive forest pest species. Ecography 37(7):648–658
Robledo-Arnuncio JJ, Klein EK, Muller-Landau HC, Santamaría L (2014) Space, time and complexity in plant dispersal ecology. Mov Ecol 2(1):16
Saupe D (1988) Algorithms for random fractals. The science of fractal images. Springer, New York, pp 71–136
Schurr FM, Pagel J, Cabral JS, Groeneveld J, Bykova O, O’Hara RB, Hartig F, Kissling WD, Linder HP, Midgley GF, Schröder B (2012) How to understand species’ niches and range dynamics: a demographic research agenda for biogeography. J Biogeogr 39(12):2146–2162
Sebert-Cuvillier E, Simon-Goyheneche V, Paccaut F, Chabrerie O, Goubet O, Decocq G (2008) Spatial spread of an alien tree species in a heterogeneous forest landscape: a spatially realistic simulation model. Landsc Ecol 23(7):787–801
Sharov AA, Liebhold AM (1998) Model of slowing the spread of gypsy moth (Lepidoptera: Lymantriidae) with a barrier zone. Ecol Appl 8(4):1170–1179
Shigesada N, Kawasaki K (1997) Biological invasions: theory and practice. Oxford University Press, Oxford
Smith R, Tan C, Srimani JK, Pai A, Riccione KA, Song H, You L (2014) Programmed Allee effect in bacteria causes a tradeoff between population spread and survival. Proc Nat Acad Sci USA 111(5):1969–1974
Sutherland WJ, Freckleton RP, Godfray HCJ, Beissinger SR, Benton T, Cameron DD, Carmel Y, Coomes DA, Coulson T, Emmerson MC (2013) Identification of 100 fundamental ecological questions. J Ecol 101(1):58–67
Thies C, Haenke S, Scherber C, Bengtsson J, Bommarco R, Clement LW, Ceryngier P, Dennis C, Emmerson M, Gagic V, Hawro V (2011) The relationship between agricultural intensification and biological control: experimental tests across europe. Ecol Appl 21(6):2187–2196
Tobin PC, Blackburn LM (2007) Slow the spread: a national program to manage the gypsy moth. General Technical Report-Northern Research Station, USDA Forest Service (NRS-6)
Tobin PC, Liebhold AM, Anderson E (2015) Estimating spread rates of non-native species: the gypsy moth as a case study. In: Venette RC (ed) Pest Risk Modelling and Mapping for Invasive Alien Species. CAB International and USDA, Wallingford, pp 131–144
Tscharntke T, Tylianakis JM, Rand TA, Didham RK, Fahrig L, Batary P, Bengtsson J, Clough Y, Crist TO, Dormann CF, Ewers RM (2012) Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biol Rev 87(3):661–685
Vilà M, Ibáñez I (2011) Plant invasions in the landscape. Landsc Ecol 26(4):461–472
Wang L, Jackson DA (2014) Shaping up model transferability and generality of species distribution modeling for predicting invasions: implications from a study on Bythotrephes longimanus. Biol Invas 16:1–25
Wang Z, Wu J, Shang H, Cheng J (2011) Landscape connectivity shapes the spread pattern of the rice water weevil: a case study from Zhejiang, China. Environ Manag 47(2):254–262
With KA (2002) The landscape ecology of invasive spread. Conserv Biol 16(5):1192–1203
With KA (2004) Assessing the risk of invasive spread in fragmented landscapes. Risk Anal 24(4):803–815
With KA, King AW (1999a) Dispersal success on fractal landscapes: a consequence of lacunarity thresholds. Landsc Ecol 14(1):73–82
With KA, King AW (1999b) Extinction thresholds for species in fractal landscapes. Conserv Biol 13(2):314–326
Zhang QH, Schlyter F (2004) Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric For Entomol 6(1):1–20
Acknowledgements
Thanks are due to all involved, including Ursula Torres, Mariona Roige and Marona Rovira Capdevila for their help with the interpretation of the results. Work was supported by Bio-Protection Research Centre, Canterbury, New Zealand.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Lustig, A., Stouffer, D.B., Doscher, C. et al. Landscape metrics as a framework to measure the effect of landscape structure on the spread of invasive insect species. Landscape Ecol 32, 2311–2325 (2017). https://doi.org/10.1007/s10980-017-0570-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-017-0570-3