
Abstract
Context
Species distributions are influenced by habitat conditions and ecological processes at multiple spatial scales. An understanding of the importance of habitat characteristics at different spatial scales is important when developing biodiversity conservation measures.
Objectives
We investigated the effect of habitat characteristics or amount at three spatial scales on the occurrence of saproxylic (=dead wood-dependent) beetles.
Methods
Saproxylic beetles were sampled under the bark of dead wood in a managed forest landscape in central Sweden. We modelled the occurrence probability in dead wood items of 44 species (all species occurring in >2 % of the items), based on dead wood item characteristics, forest stand characteristics, and habitat connectivity (i.e. area of potentially suitable forest stands in the surrounding of each stand), using hierarchical Bayesian regression.
Results
For the majority of species, dead wood item characteristics (especially tree species and whether standing or downed) were more important than measured stand characteristics and habitat connectivity. Whether the stands were clear-cuts, mature forests, or reserves affected some species, whereas the stand-level amount of dead wood per hectare was not important for any species. Habitat connectivity improved the occurrence models for about a half of the species, but there were both positive and negative relationships, and they were generally weak.
Conclusions
Forest management should include creation and retention of a high diversity of dead wood to sustain habitat for all species. In a forest-dominated landscape, the spatial distribution of dead wood is of little importance for common saproxylic beetle species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Species distributions are influenced by habitat conditions and ecological processes at multiple spatial scales (Wiens 1989; Levin 1992). Human modifications of natural systems also occur at several scales, and a big challenge in conservation biology is to identify the appropriate spatial scales at which conservation actions most likely will balance the negative effects of human impacts (Wiens 1989; Saunders et al. 1991). An understanding of the relative importance of key habitat characteristics at different spatial scales is therefore important when developing biodiversity conservation measures in human-modified landscapes.
Commercial logging has turned large regions of the natural boreal forest ecosystem of Europe and North America into forest landscapes dominated by monospecific even-aged stands, where the multi-aged and structurally more diverse natural forest stands are rare and scattered (Esseen et al. 1992; Bergeron et al. 2002). This has raised concerns about how to efficiently extract forest products and still maintain biodiversity. For conservation measures to be applied at the scale where they are most probable to improve persistence of naturally occurring species, knowledge of the relative importance of key habitat characteristics at different spatial scales on species occurrence is needed. However, for forest species such knowledge is generally limited (reviews of epiphytic lichens: Nascimbene et al. 2013, and dead-wood associated species: Sverdrup-Thygeson et al. 2014; see however bird studies, e.g., Saab 1999; Rolstad et al. 2000).
About 20 % of all multicellular organisms in boreal forests are saproxylic, i.e. dependent on dead wood, or on other saproxylic organisms (Stokland et al. 2012). Fungi and beetles are the largest groups among the saproxylic organisms. These organisms have been given much attention in nature conservation, because intensive forest management strongly decreases the quantity and diversity of coarse dead wood (Siitonen 2001). For instance, more than a half of all red-listed forest species in Fennoscandia are saproxylic. Saproxylic species are often associated with certain characteristics of the dead wood items (e.g. Lindhe et al. 2005; Berglund et al. 2011). At the stand scale, a higher amount of dead wood have been found to increase species richness (Lassauce et al. 2011), but its effect on the probability of occurrence of individual species has rarely been studied. Since forest habitats are dynamic, species occurrence relies on colonisation from dispersal sources in the surrounding landscape (Ranius et al. 2014). Consequently, the composition of the surrounding landscape may influence the occurrence patterns of saproxylic organisms (e.g. Gu et al. 2002; Ranius et al. 2010). Most studies that investigate the effect of variables at various spatial scales on saproxylic organisms in forests have, however, either ignored the importance of habitat connectivity (Stenbacka et al. 2010; Berglund et al. 2011) or focused on single species (e.g. Sverdrup-Thygeson and Midtgaard 1998; Schroeder et al. 2007; Jackson et al. 2012; Rubene et al. 2014). Therefore, more knowledge is needed on the relative importance of dead wood quality, stand characteristics, and surrounding landscape on a larger range of saproxylic species.
Hierarchical Bayesian modelling could be used for fitting species distribution models with complex multilevel structures to properly model the influence of different explanatory variables at their hierarchical level and account for different sources of variation across different spatial scales (Gelman and Hill 2007). Recently, such approaches have been used for quantifying habitat requirements of individual species in whole communities, based on the relative influence of local and larger-scale explanatory variables. Such analyses have been done on saproxylic fungi (Berglund et al. 2011; Nordén et al. 2013) but to our knowledge not on any other forest-dwelling groups.
The aim of this study was to investigate the relative importance of habitat characteristics at multiple spatial scales for explaining the occurrence of saproxylic beetles. Specifically, we modelled the occurrence probability of individual beetle species on dead wood items, based on characteristics of dead wood items, stand characteristics, and habitat connectivity, using the hierarchical Bayesian framework. To evaluate the relative importance of spatial scale, we compared average differences in deviance information criterion (DIC) between models that included different sets of variables.
Methods
Study landscape and stand selection
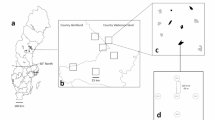
The study was conducted in a 20,000 ha study landscape (Fig. 1) in the Swedish province of Hälsingland (Fig. 1; 62°N, 16°E), situated in the southern boreal vegetation zone (Sjörs 1999). Data on dead wood amounts are available from this landscape (Ekbom et al. 2006). The forest land in the study landscape is owned by one forest company, Holmen Skog AB. Since the 1950s, the forest has been managed more intensively and harvested at thinnings and by clear-felling. Therefore, the landscape is today mainly composed of even-aged management blocks (stands) covering the entire rotation period of about 100 years. There are also three large, legally protected nature reserves, covering 3.1 % of the landscape. Norway spruce (Picea abies (L.) Karsten) and Scots pine (Pinus sylvestris L.) dominate the forests, followed by birch (Betula pendula Roth and B. pubescens Ehrh.) and aspen (Populus tremula L.). Productive forest (i.e. with a potential forest growth >1 m3 ha−1) covers 83.0 % of the landscape. In the managed stands, deciduous trees rarely constitute >20 % of the standing volume.

Location of the 56 forest stands where beetles were sampled, in a study landscape situated in central Sweden. The distance to the nearest neighbour was, on average, 965 m
We surveyed beetles in 56 forest stands, which all were productive forests dominated by Scots pine or Norway spruce. We used databases from the forest owner to randomly select these stands, interspersed across the study landscape and classified into three stand types (Table 1). (1) “Clear-cuts” were 3–7 years old canopy-open stands; (2) “mature” stands (≥60 years old) were canopy-closed, managed stands; and (3) “reserves” were canopy-closed, unmanaged forests that are legally protected. These three stand types are potentially important habitat for saproxylic beetles living under bark, since they are characterized by large volumes of dead wood with bark, compared to managed stands 8–59 years old, where the dead wood recruitment is low and most of the remaining dead wood items have lost their bark (Ekbom et al. 2006). Due to large sizes of the included three reserves (427, 242, and 82 ha), we divided them into 6, 6, and 2 equally sized sub-areas, respectively. The sub-areas were treated as individual stands in the analyses, and this treatment was supported by low levels of spatial autocorrelation of the residuals from the best full models (see "Statistical analyses" section) for all species tested (Moran’s I, pmin = 0.07). More mature stands were surveyed than clear-cuts (27 vs. 15), which reflects the difference in total area between these two stand types in the study landscape.
Beetle sampling
We aimed at sampling saproxylic beetles in 10 dead wood items per forest stand. We only selected items with a diameter >10 cm and a length >0.5 m with bark left, but avoided the youngest decay stage in which bark beetles are typically dominating. In each forest stand, the selection was done randomly from logs (downed CWD) and snags (standing CWD) that occurred within one of four 20 m × 100 m sampling rectangles. If fewer than 10 dead wood items were found in this way, we searched in the remaining parts of the stand for suitable dead wood items. If there still were fewer than 10 available items, we sampled all available items. During 2 years (2002 and 2003), 383 dead wood items were sampled. For each item we sieved 1 m2 of bark through a coarse net (Wikars et al. 2005), and the resulting fine fraction was placed into Tullgren funnels (30 cm wide, 8 mm mesh size) where beetles were extracted under a lamp (Southwood and Henderson 2000). We used 60 W light bulbs as heat and light source, and extraction lasted for at least 24 h. We identified all adult Coleoptera to species or genus level, but in the analyses we only included species known to be obligate saproxylic (Dahlberg and Stokland 2004). Nomenclature of beetles follows Silfverberg (2004).
Explanatory variables
We measured variables that may affect the occurrence of saproxylic beetles on three spatial scales: dead wood items, stands, and surrounding landscape. For each dead wood item we recorded five explanatory variables: diameter, tree species, position (standing or downed), degree of shade, and decay stage. We estimated the degree of shade on a four-level scale: exposed to direct sunlight during (1) >75 %, (2) 50–75 %, (3) 25–50 %, and (4) <25 % of the day. The decay stage was estimated on a six-level scale based on the hardness of the wood (Siitonen and Saaristo 2000).
We analysed three stand variables: stand size, amount of coarse dead wood, and stand type. We obtained stand size using databases from the forest owner, and amount of coarse dead wood (both logs and snags) from a previous study in the same forest stands (Ekbom et al. 2006). For coarse dead wood amount, we used volume (m3) dead wood ha−1, including dead wood items with a diameter >10 cm (7 cm for deciduous trees) and a length >0.5 m. We included stand type because clear-cuts, mature stands, and reserves differ from each other regarding, e.g., forest age and exposure to sun and wind (Table 1).
At the landscape scale, we estimated the amount of habitat in the surrounding landscape by summing the area of all stand types defined as habitat (i.e. clear-cuts, mature stands, and reserves, since they have larger volumes of dead wood with bark; Ekbom et al. 2006) within four buffer circles (500, 1000, 2000, and 4000 m radius, respectively) around each target stand. We used a measure based only on habitat availability since that could potentially be used in forest planning, whereas connectivity measures that require data about individual species’ occurrence patterns and biology are not feasible in most landscapes due to lack of information. We used a buffer metric, since for saproxylic beetles it performs equally well as measures that up-weight closer patches (Ranius et al. 2010). The estimation of surrounding habitat amount was done in ArcMAP 10, and the maximum radius (4000 m) was the maximum distance for which we had available information of stand characteristics for all surrounding stands. Six of the analysed species were identified as canopy-closed species, since they either did not occur in clear-cuts or were associated with closed stands according to statistical analyses (i.e. negative 95 % credible intervals for clear-cut did not overlap 0; Fig. 2). For these six species, we used the summed area of only mature stands and reserves as the habitat connectivity variable. In the analyses, we tested the four spatial scales (radii) for the habitat connectivity one by one in separate models. For stands situated spatially close, the buffer circles overlapped, and we hence to some extent psedo-replicate the connectivity measure. However, that is a minor problem as the spatial autocorrelation was low for the model residuals see "Study landscape and stand selection" section).

Estimates of parameters in Bayesian generalized linear models for occurrence probability of saproxylic beetle species on individual dead wood items. The modes (short vertical lines), 50 % (thick horizontal lines) and 95 % (thin horizontal lines) credible intervals are shown for the full models including or excluding habitat connectivity. Grey lines indicate that the 95 % credible interval includes 0, black lines that it does not. Asterisk denotes that parameter estimates are, for visibility reasons, scaled by a factor 10. 1 and 2 denote effects of habitat connectivity at 500 and 4000 m, respectively. For the categorical variables “tree species”, “dead wood position”, and “stand type”, only parameters for categories other than the reference category (birch, downed, and reserves, respectively) are given. Dead wood amount was also tested, but that is not shown here since it was not included in the final model for any of the species. Standard deviation of random error (σ) is the parameter associated with unexplained between-stand variation
We checked all continuous explanatory variables for cross-correlations prior to the analyses. The highest Pearson correlation coefficient, r = 0.3, was found between stand size and dead wood amount ha−1. For tests of associations between categorical and continuous variables, we performed one-way ANOVAʼs with Tukey’s honestly significant differences post hoc test. Reserve stands were larger than both clear-cuts and mature stands, and their dead wood amounts ha−1 were also higher (Table 1). Individual dead wood items were less shaded in clear-cuts compared to both mature stands and reserves, and more shaded in mature stands than in reserves (Table 1). Dead wood diameter was greater in clear-cuts compared to both mature stands and reserves (Table 1).
Statistical analyses
We modelled occurrence probability of individual beetle species based on dead wood characteristics, stand characteristics, and habitat connectivity, while accounting for the hierarchical structure of the data. Analyses were conducted for the most frequent 44 species, which had occurrences in at least eight (>2 %) of the sampled dead wood items. Specifically, we analysed the probability of occurrence on individual dead wood items, using Bayesian hierarchical generalized linear models (Gelman and Hill 2007) with a logit link function (logistic regression) and varying intercepts. The hierarchical Bayesian framework enables the utilization of explanatory variables measured at the stand level (i.e. at the higher hierarchical level), as they are used to model the stand-specific intercepts (Gelman and Hill 2007). We assumed a Bernoulli probability distribution of the binary response variable (yij; species presence/absence) and modelled species occurrence probability on dead wood item i in stand j, i.e. P(yij = 1) as:
where α j is the stand-specific intercept (see below), X ijk is the dead wood item-level explanatory variable k for dead wood item i in stand j and β k is the dead wood item-level effect-size parameter of explanatory variable k (n in total). The stand-specific intercepts (α j ) were modelled as:
where σα is the standard deviation of a normal distribution with a mean (\(\mu_{{\alpha_{j} }}\)) modelled based on the stand-level explanatory variables as:
where γ is an intercept parameter, Z jm is the stand-level explanatory variable m for stand j and ρ m the associated effect-size parameter (h in total). Hence, the intercepts vary between stands and σ α determines the between-stand variation (here called random error). The landscape-scale variable (habitat connectivity at four spatial scales) was treated as a stand variable in the model, but was added separately at the end of the model-building procedure.
We constructed hierarchical Bayesian models for each species using different sets of variables, but always with a hierarchical structure (i.e. with varying intercepts). First, we parameterized one model containing only dead wood variables (henceforth, dead wood model). Then we selected the model with the lowest deviance information criterion (DIC) among models with all combinations of the five dead wood variables. DIC is analogous to the Akaike information criterion (AIC), and is well-suited for Bayesian hierarchical modelling (Spiegelhalter et al. 2002). Second, we repeated the same model selection procedure, but with only stand variables included (henceforth, stand model). Third, we constructed full models with both dead wood and stand variables included in the model selection procedure. The variables included in these full models could be only dead wood, only stand, both dead wood and stand, or no variables, depending on the species tested. To evaluate the relative importance of dead wood and stand variables, we compared the average differences in DIC between the hierarchical null model (i.e. including stand identity as a random factor but no explanatory variables) and the dead wood, stand, and full model, respectively. Finally, we tested whether adding habitat connectivity at four spatial scales, one by one, to the full models improved the models by reducing DIC.
We estimated the posterior distributions of the Bayesian model parameters in Eqs. (1) and (2), using two Monte Carlo Markov chains of 610,000 iterations each. We discarded 10,000 iterations as ‘burn-in’ and then saved every 60th iteration to accumulate 10,000 values from each chain (i.e. 20,000 in total). To improve convergence of the chains and simplify the interpretation of the models, we centred all variables (i.e. subtracted the mean from each measured value) and also standardized (i.e. dividing each measured value by 2 sd) the continuous variables (Gelman and Hill 2007). For categorical dead wood (tree species and position) and stand (stand type) variables, we excluded categories in which the species was not found. Consequently, we excluded birch for seven species, clear-cuts for three species, and both birch and clear-cuts for one species (Table A1).
We used uninformative prior distributions for all model parameters. We used normal distributions with mean = 0 and variance = 1000 for all effect size parameters and the intercept γ, while σα was drawn from a uniform distribution between 0 and 100. To evaluate convergence, we visually inspected the trace plots and used the Gelman-Rubin diagnostic (Gelman and Hill 2007). Convergence (R < 0.1) was reached for all estimated parameters. We summarized the posterior distribution of estimated parameters by calculating the distribution mode and Bayesian 50 and 95 % credible intervals. For the analyses, we used the statistics software JAGS (Plummer 2003) and R 2.14.0 (R Development Core Team 2011).
Results
Characteristics of dead wood items were more important for explaining species’ occurrence probability than characteristics measured of the forest stands, as judged by the average reduction in DIC between the hierarchical null models and the dead wood (16.7) and stand (1.7) models, respectively (Fig. 3). When adding habitat connectivity, DIC was reduced by 1.9. The average reduction in DIC between the hierarchical null models and the full models was close to the average reduction between the hierarchical null models and the dead wood models (Fig. 3).

Change in DIC (±SE) between models including explanatory variables at different spatial scales and a null model with only the random stand effect included. Variables included in the full models can be only dead wood, only stand, both dead wood and stand, or no variables (see Fig. 2 for species-specific details). hc means habitat connectivity
Tree species and position (i.e. standing or downed) were the variables that were important for the largest number of species; they were included in the final occurrence models for 22 and 21 beetle species, respectively (Fig. 2). However, among the studied species there were no specialists; none occurred only in one tree species or only in either standing or downed dead wood. The majority of the beetles affected by tree species (18 of 21) were associated with conifers (spruce or pine), whereas only three species were associated with birch (i.e. the negative 95 % credible intervals for both spruce and pine did not overlap 0; Fig. 2). Furthermore, there were eight species that did not occur on birch at all, and consequently the effect of birch was not tested for them (Table A1). Degree of shade, diameter, and decay stage were included in the full models for 13, 10, and 9 species, respectively (Fig. 2).
Stand type was the most important stand characteristic for explaining species occurrence, and was included in the final model for seven species. Clear-cuts had a negative effect on several species: four species did not occur on clear-cuts at all (Table A1), and the occurrence probability of two species was lower on clear-cuts compared to reserves (Fig. 2). One species was associated with reserves, whereas for two species the occurrence probability was higher in mature stands compared to reserves (Fig. 2). Stand size affected very few species, whereas the amount of dead wood did not affect any of the species.
Adding habitat connectivity to the full model improved the models for 24 species, but in most cases the 95 % credible interval included 0 (Fig. 2). For 11 species the relationship was positive, whereas for 13 species it was negative.
Discussion
Relative importance of spatial scales
We found that characteristics of the dead wood items were more important than characteristics measured of the forest stand and surrounding landscape for explaining the occurrence of relatively common saproxylic beetles in a managed boreal forest landscape (Fig. 3). Thus, this beetle community are mainly conforming to what in metacommunity ecology is referred to as the species sorting view, which is defined by the close link between species distributions and local conditions (acting directly or by altering competitive abilities) together with sufficient availability of dispersal sources (Leibold et al. 2004). However, our result may not only be a consequence of the species’ biology, but may also reflect that it is easier to measure characteristics relevant for saproxylic species at a dead wood item scale rather than at a stand and landscape scale. The characteristics we used are representative for what is typically measured in biodiversity monitoring and surveys. For that reason our outcome is still relevant for management and conservation, suggesting that strategies should be based more on characteristics of dead wood items rather than stand and landscape characteristics. It should be noted that this study only includes the 44 relatively common saproxylic species, and for rarer and more specialised species, for which the habitat is more fragmented, habitat connectivity is expected to be more important (Fahrig 1998; Nordén et al. 2013).
There were correlations between some characteristics of the dead wood items and stand type (Table 1); however, we believe that these correlations have minor influence on our main conclusions since the characteristics that influenced the largest number of species (tree species and position) did not differ between different stand types. Perhaps stand characteristics would have an overall slightly higher relative importance if shade and diameter was not included at the lower hierarchical level.
Effects of characteristics of dead wood items
The characteristics of dead wood items were important for explaining occurrence of the majority of the saproxylic beetles (Fig. 2). This agrees with earlier studies of saproxylic beetles (see however Wikars 2002; Ulyshen and Hanula 2009; Jackson et al. 2012; ) and fungi (Stokland and Kauserud 2004; Berglund et al. 2011). For both beetle larvae and fungi, development takes place in one single dead wood item, which can explain why the conditions in individual logs are important for the recruitment of adult beetles and fruiting bodies of fungi. The most important characteristics of the dead wood items for explaining species occurrence in the present study were tree species, position, decay stage, and degree of shade. Even though there were no true specialists, many species occurred more frequently in certain types of dead wood. These dead wood characteristics may reflect microclimatic conditions (moisture and temperature) as well as nutrient supply (for instance, availability of fungi), and have been shown important for explaining occurrence of saproxylic organisms also in earlier studies (e.g. Ranius and Jansson 2000; Jonsell and Weslien 2003; Lindhe et al. 2005; Saint-Germain et al. 2007; Berglund et al. 2011). The direction of the impact of dead wood characteristics varied among species, which suggests that a high heterogeneity of microhabitats may increase the diversity of saproxylic species (Davies et al. 2008).
Effects of stand characteristics
Even if dead wood characteristics explained most of the variation in the occurrence patterns (Fig. 3) also stand characteristics were important; for instance, stand type influenced the occurrence probability of 20 % of the species. This was mainly because species occurred in lower frequency on clear-cuts compared to the canopy-closed mature and reserve stands. This agrees with earlier findings of similarities in saproxylic beetle communities among mature managed and old-growth boreal stands, but divergent species composition in clear-cuts (McGeoch et al. 2007; Stenbacka et al. 2010; Hjältén et al. 2012). One reason for this divergence is the difference in sun exposure, which affects saproxylic beetles (Similä et al. 2002; Lindhe et al. 2005). Species dependent on forest cover continuity, dead wood, and large trees have been found to be more species rich in unmanaged forests than in managed ones (Paillet et al. 2010). The relatively weak effect of management in our study may be due to that there are relatively small differences in dead wood amounts between mature managed stands and reserves (Table 1) compared to the differences that often occur between old-growth forests and forests that have been managed by clear-felling since a long time (Siitonen 2001).
We found no effect of dead wood amount per hectare and only a weak effect of stand size on species occurrence probability per dead wood item. In many studies, higher amounts of dead wood increase species richness and probability of occurrence of saproxylic organisms per forest stand (Lassauce et al. 2011 and references therein). The positive effects of the amount of dead wood on species richness reported in the literature could in most cases be explained by a sampling effect alone, i.e. by the fact that a larger amounts of dead wood will contain more individuals and this will imply more species (Fahrig 2013). This is the case when window traps are used to collect saproxylic beetles, since they capture beetles from a larger volume of dead wood if situated at a spot with a higher density of dead wood. Our study is one of a few in which the amount of dead wood sampled was standardized, which is necessary when disentangling the island effect (i.e. higher species densities on larger habitat islands) and the sampling effect (Fahrig 2013). For saproxylic beetles, such standardized samples are obtained by searching through certain amounts of dead wood (using, for instance, bark sieving and extraction as in the present study) and when using emergence traps (e.g., Wikars et al. 2005). An island effect is expected according to the island biogeography theory (predictions about species richness; MacArthur and Wilson 1967) and the resource concentration hypothesis (predictions about population densities; Root 1973). Our results imply that there is no island effect; however, other studies of saproxylic beetles have revealed an island effect, since they have observed a positive effect of habitat amount at the stand scale on the probability of species occurrence per dead wood item (Komonen et al. 2000; Ranius 2002; Sahlin and Schroeder 2010; Victorsson and Jonsell 2013). These studies have mainly focused on species specialised to certain dead wood types with a highly fragmented distribution, while in the present study we analysed the 44 most frequently occurring species in a wide range of dead wood types. Also, a study conducted in the same area as the present study, focusing on certain redlisted saproxylic beetle species, suggested that some species are demanding regarding amounts of certain qualities of dead wood at a local scale (Rubene et al. 2014), but these species were too rare to be analysed in the present study. The lack of relationship in the present study may be explained by the fact that forest stands with at least some dead wood present occurred relatively continuously in the landscape. Consequently, there are many dispersal sources for the relatively common species that were included in the present study. This makes the amount of dispersal sources within each forest stand a less critical factor. Another possible reasons for the weak effect of current dead wood amounts is that saproxylic species richness may be better explained by other factors which are difficult to measure, such as the historical continuity of dead wood. Some studied indicates that historical continuity is important for rare and threatened saproxylic beetles (Nilsson and Baranowski 1997; Siitonen and Saaristo 2000), but little is known about its effect on more common species. It should also be remembered that in the present study, stand size and dead wood amounts differed between the three stand types, and the weak effect could therefore also be because including stand type in the model removes some of the variation in these two explanatory variables. However, this potential bias is still only valid for a few species; only six species had any stand characteristics that did not overlap zero in their final model.
Effects of habitat connectivity
Habitat connectivity affected the occurrence of many species; however, the effect was usually weak and there were nearly as many negative as positive relationships (Fig. 2). The occurrence of both negative and positive effects suggests that the spatial location of the dead wood items had some effect on species’ occurrence; however, the spatial pattern was not clearly associated with habitat density. We had expected a clearer positive relationship, due to higher colonization rates when there are higher habitat density, and thus larger dispersal sources nearby (Thomas et al. 1992). One reason could be that we mainly analyse rather common species. Several other studies of saproxylic beetles, which have shown clearer positive effect of habitat connectivity, have focused on species specialised to habitats that are more fragmented in comparison to the present study (Økland et al. 1996; Ranius et al. 2010, 2014; Götmark et al. 2011; Bergman et al. 2012). It could be that since all species in our study occur in managed forest, and the study landscape is dominated by managed forest, the landscape is not very fragmented for these species. At such low level of habitat fragmentation, habitat quality has generally a greater influence than habitat connectivity on species occurrence patterns (Fahrig 1998; Andrén 1999). Among saproxylic fungi, specialised species have indeed been found to be more sensitive to habitat fragmentation than generalist species since they respond more negatively to connectivity (Nordén et al. 2013). Another reason for the weak effect in the present study could be that the importance of habitat connectivity may be underestimated when analysing snapshot data in landscapes where habitat conditions change over time (Hodgson et al. 2009). In our study landscape, the area covered by older forest has clearly decreased during the last 50 years, and therefore the current species occurrence patterns may to some extent reflect historical habitat connectivity (Schroeder et al. 2007). Thirdly, we measured connectivity as the amount of habitat in the surroundings, while a measure that includes information on habitat quality or species’ occurrences would reflect the amount of dispersal sources better (Ranius et al. 2010). An advantage with the measure we used is that it better reflects what could potentially be used in management, since it only requires data that are widely available.
Implications for conservation
We found that for the occurrence of the more common saproxylic beetle species’, the quality of dead wood items is more important than their spatial location. The habitat requirements regarding dead wood characteristics (i.e. tree species, position, decay stage and degree of shade) differed among species. Therefore, conservation measures aiming at mitigating negative impacts of forestry should aim at creating not only large amounts, but also a high diversity of dead wood. Attempts have been made to identify “thresholds” in the dead wood amounts that should be exceeded for sustaining biodiversity (Müller and Bütler 2010). However, due to the lack of relationships between amount of dead wood per stand and probability of occurrence per dead wood item, our study does not lend support for any such thresholds at a forest stand level. Our study only included more common species, but it may be that rarer species is more demanding (cf. Penttilä et al. 2004). To some extent, our outcome may also be because we lack detailed data on the amount of dead wood that is suitable for each species. In that sense our study is more similar to the situation for practitioners, who do not have detailed data about all individual species’ occurrence patterns and biology. In our study landscape, the amount of dead wood with certain qualities is probably a key factor to allow persistence of the saproxylic fauna. However, in forest habitats that are more fragmented and for rare and demanding species, high concentration of habitat may be important for species’ occurrence (e.g. Ranius et al. 2010; Bergman et al. 2012).
References
Andrén H (1999) Habitat fragmentation, the random sample hypothesis and critical thresholds. Oikos 84:306–308
Bergeron Y, Leduc A, Harvey BD, Gauthier S (2002) Natural fire regime: a guide for sustainable management of the Canadian boreal forest. Silva Fenn 36:81–95
Berglund H, Hottola J, Penttilä R, Siitonen J (2011) Linking substrate and habitat requirements of wood-inhabiting fungi to their regional extinction vulnerability. Ecography 34:864–875
Bergman K-O, Jansson N, Claesson K, Palmer MW, Milberg P (2012) How much and at what scale? Multiscale analyses as decision support for conservation of saproxylic oak beetles. For Ecol Manag 265:133–141
Dahlberg A, Stokland J (2004) Vedlevande arters krav på substrat—en sammanställning och analys av 3600 arter. Skogsstyrelsen, Jönköping
Davies ZG, Tyler C, Stewart GB, Pullin AS (2008) Are current management recommendations for saproxylic invertebrates effective? A systematic review. Biodivers Conserv 17:209–234
Ekbom B, Schroeder LM, Larsson S (2006) Stand specific occurrence of coarse woody debris in a managed boreal forest landscape in central Sweden. For Ecol Manag 221:2–12
Esseen P-A, Ehnström B, Ericson L, Sjöberg K (1992) Boreal forests—the focal habitats of Fennoscandia. In: Hansson L (ed) Ecological principles of nature conservation. Elsevier Science Publishers, Amsterdam, pp 252–325
Fahrig L (1998) When does fragmentation of breeding habitat affect population survival? Ecol Model 105:273–292
Fahrig L (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. J Biogeogr 40:1649–1663
Gelman A, Hill J (2007) Data analysis using regression and multilevel/hierarchical models. Cambridge University Press, Cambridge
Götmark F, Åsegård E, Franc N (2011) How we improved a landscape study of species richness of beetles in woodland key habitats, and how model output can be improved. For Ecol Manag 262:2297–2305
Gu W, Heikkilä R, Hanski I (2002) Estimating the consequences of habitat fragmentation on extinction risk in dynamic landscapes. Landscape Ecol 17:699–710
Hjältén J, Stenbacka F, Pettersson RB, Gibb H, Johansson T, Danell K, Ball JP, Hilsczański J (2012) Micro and macro-habitat associations in saproxylic beetles: implications for biodiversity management. PLoS One 7:e41100
Hodgson JA, Moilanen A, Thomas CD (2009) Metapopulation responses to patch connectivity and quality are masked by successional habitat dynamics. Ecology 90:1608–1619
Jackson HB, Baum KA, Cronin JT (2012) From logs to landscapes: determining the scale of ecological processes affecting the incidence of a saproxylic beetle. Ecol Entomol 37:233–243
Jonsell M, Weslien J (2003) Felled or standing retained wood—it makes a difference for saproxylic beetles. For Ecol Manag 175:425–435
Komonen A, Penttilä R, Lindgren M, Hanski I (2000) Forest fragmentation truncates a food chain based on an old-growth forest bracket fungus. Oikos 90:119–126
Lassauce A, Paillet Y, Jactel H, Bouget C (2011) Deadwood as a surrogate for forest biodiversity: meta-analysis of correlations between deadwood volume and species richness of saproxylic organisms. Ecol Indic 11:1027–1039
Leibold MA, Holyoak M, Mouquet N, Amaresekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzalez A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613
Levin SA (1992) The problem of pattern and scale in ecology. Ecology 73:1943–1967
Lindhe A, Lindelöw Å, Åsenblad N (2005) Saproxylic beetles in standing dead wood density in relation to substrate sun-exposure and diameter. Biodivers Conserv 14:3033–3053
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton, p 203
McGeoch M, Schroeder M, Ekbom B, Larsson S (2007) Saproxylic beetle diversity in a managed boreal forest: importance of stand characteristics and forestry conservation measures. Divers Distrib 13:418–429
Müller J, Bütler R (2010) A review of habitat thresholds for dead wood: a baseline for management recommendations in European forests. Eur J For Res 129:981–992
Nascimbene J, Thor G, Nimis PL (2013) Effects of forest management on epiphytic lichens in temperate deciduous forests of Europe—a review. For Ecol Manag 298:27–38
Nilsson SG, Baranowski R (1997) Habitat predictability and the occurrence of wood beetles in old-growth beech forests. Ecography 20:491–498
Nordén J, Penttilä R, Siiitonen J, Tomppo E, Ovaskainen O (2013) Specialist species of wood-inhabiting fungi struggle while generalists thrive in fragmented boreal forests. J Ecol 101:701–712
Økland B, Bakke A, Hågvar S, Kvamme T (1996) What factors influence the diversity of saproxylic beetles? A multiscaled study from a spruce forest in southern Norway. Biodiver Conserv 5:75–100
Paillet Y, Bergès L, Hjältén J, Ódor P, Avon C, Bernhrdt Römermann M, Bijlsma R-J, deBruyn L, Fuhr M, Grandin U, Kanka R, Lundin L, Luque S, Magura T, Matesanz S, Mészáros I, Sebastià M-T, Schmidt W, Standvár T, Tóthmérész B, Uotila A, Valladares F, Vellak K, Virtanen R (2010) Biodiversity differences between managed and unmanaged forests: meta-analysis of species richness in Europe. Conserv Biol 24:101–112
Penttilä R, Siitonen J, Kuusinen M (2004) Polypore diversity in managed and old-growth boreal Picea abies forests in southern Finland. Biol Conserv 117:271–283
Plummer M (2003) Jags: a program for analysis of Bayesian graphical models using Gibbs sampling. DSC working papers, Austrian Association for Statistical Computing, Vienna
R Development Core Team (2011) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0
Ranius T (2002) Influence of stand size and quality of tree hollows on saproxylic beetles in Sweden. Biol Conserv 103:85–91
Ranius T, Jansson N (2000) The influence of forest regrowth, original canopy cover and tree size on saproxylic beetles associated with old oaks. Biol Conserv 95:85–94
Ranius T, Johansson V, Fahrig L (2010) A comparison of patch connectivity measures using data on invertebrates in hollow oaks. Ecography 33:971–978
Ranius T, Bohman P, Hedgren O, Wikars L-O, Caruso A (2014) Metapopulation dynamics of a beetle species confined to burned forest sites in a managed forest region. Ecography 37:797–804
Rolstad J, Løken B, Rolstad E (2000) Habitat selection as a hierarchical spatial process: the green woodpecker at the northern edge of its distribution range. Oecologia 124:116–129
Root RB (1973) Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecol Monogr 43:95–124
Rubene D, Wikars L-O, Ranius T (2014) Importance of high quality early-successional habitats in managed forest landscapes to rare beetle species. Biodivers Conserv 23:449–466
Saab V (1999) Importance of spatial scale to habitat use by breeding birds in riparian forests: a hierarchical analysis. Ecol Appl 9:135–151
Sahlin E, Schroeder LM (2010) Importance of habitat patch size for occupancy and density of aspen-associated saproxylic beetles. Biodivers Conserv 19:1325–1339
Saint-Germain M, Drapeau P, Buddle CM (2007) Host-use patterns of saproxylic phloeophagous and xylophagous Coleoptera adults and larvae along the decay gradient in standing dead black spruce and aspen. Ecography 30:737–748
Saunders DA, Hobbs RJ, Margules CR (1991) Biological consequences of ecosystem fragmentation—a review. Conserv Biol 5:18–32
Schroeder LM, Ranius T, Ekbom B, Larsson S (2007) Spatial occurrence of a habitat-tracking saproylic beetle inhabiting a managed forest landscape. Ecol Appl 17:900–909
Siitonen J (2001) Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example. Ecol Bull 49:11–41
Siitonen J, Saaristo L (2000) Habitat requirements and conservation of Pytho kolwensis, a beetle species of old-growth boreal forest. Biol Conserv 94:211–220
Silfverberg H (2004) Enumeratio nova Coleopterorum Fennoscandiae, Daniae et Baltiae. Sahlbergia 9:1–111
Similä M, Kouki J, Martikainen P, Uotila A (2002) Conservation of beetles in boreal pine forests: the effects of forest age and naturalness on species assemblages. Biol Conserv 106:19–27
Sjörs H (1999) The background: geology, climate and zonation. Acta Phytogeographica Suecica 84:5–14
Southwood TRE, Henderson PA (2000) Ecological methods. Blackwell Science, Oxford
Spiegelhalter DJ, Best NJ, Carlin BP, vad der Linde A (2002) Bayesian measures of model complexity and fit. J R Stat Soc B 64:583–616
Stenbacka F, Hjältén J, Hilszczański J, Dynesius M (2010) Saproxylic and non-saproxylic beetle assemblages in boreal spruce forests of different age and forestry intensity. Ecol Appl 20:2310–2321
Stokland J, Kauserud H (2004) Phellinus nigrolimitatus—a wood-decomposing fungus highly influenced by forestry. For Ecol Manag 187:333–343
Stokland JN, Siitonen J, Jonsson BG (2012) Biodiversity in dead wood. Cambridge University Press, Cambridge
Sverdrup-Thygeson A, Midtgaard F (1998) Fungus-infected trees as islands in boreal forest: spatial distribution of the fungivorous beetle Bolitophagus reticulatus (Coleoptera, Tenebrionidae). Ecoscience 5:486–493
Sverdrup-Thygeson A, Gustafsson L, Kouki J (2014) Spatial and temporal scales relevant for conservation of dead-wood associated species: current status and perspectives. Biodivers Conserv 23:513–535
Thomas CD, Thomas JA, Warren MS (1992) Distributions of occupied and vacant butterfly habitats in fragmented landscapes. Oecologia 92:563–567
Ulyshen MD, Hanula JL (2009) Habitat associations of saproxylic beetles in the southeastern United States: a comparison of forest types, tree species and wood postures. For Ecol Manag 257:653–664
Victorsson J, Jonsell M (2013) Effect of stump extraction on saproxylic beetle diversity in Swedish clear-cuts. Insect Conserv Divers 6:483–493
Wiens JA (1989) Spatial scaling in ecology. Funct Ecol 3:385–397
Wikars LO (2002) Dependence on fire in wood-living insects: an experiment with burned and unburned spruce and birch logs. J Insect Conserv 6:1–12
Wikars LO, Sahlin E, Ranius T (2005) A comparison of three methods to estimate species richness of saproxylic beetles (Coleoptera) in logs and high stumps of Norway spruce. Can Entomol 137:304–332
Acknowledgments
We thank Holmen Skog for access to their databases. Matilda Apelqvist, Björn Forsberg, Markus Franzén, David Isaksson, Niklas Jönsson, Mats Larsson, Per Larsson, Carola Orrmalm, Erik Sahlin, Måns Svensson, and Jan ten Hoopen helped with field and laboratory work. Stig Lundberg identified the beetles. Håkan Berglund, Matthew Hiron, Joakim Hjältén, and Tobias Jeppsson provided valuable comments on the manuscript. The study was supported by the Mistra research program Future Forests (to TR).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Ranius, T., Johansson, V., Schroeder, M. et al. Relative importance of habitat characteristics at multiple spatial scales for wood-dependent beetles in boreal forest. Landscape Ecol 30, 1931–1942 (2015). https://doi.org/10.1007/s10980-015-0221-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-015-0221-5