
Abstract
In Fennoscandian boreal forests, aspen (Populus tremula) is one of the most important tree species for biodiversity. In this study we explore how occupancy and density of beetles associated with dead aspen are related to habitat patch size and connectedness in a 45,000 ha boreal managed forest landscape in central Sweden. Patch size was estimated as amount of breeding substrate and connectedness as crown cover of living aspen in the surrounding landscape. The beetles were sampled by sieving of bark or by inspection of species-characteristic galleries in 56 patches with dead aspen. Six of nine aspen-associated species (Xylotrechus rusticus, Ptilinus fuscus, Mycetophagus fulvicollis, Cyphaea curtula, Homalota plana and Endomychus coccineus) showed a positive significant relationship between habitat patch size and occupancy. For all these species, except C. curtula, there was also a significant positive relationship between patch size and density. Connectedness was not retained as a significant variable in the analyses. Species not defined as aspen-associated constituted a significantly larger proportion of the total density of individuals of saproxylic beetles in smaller habitat patches than in larger patches. Richness of aspen-associated species was positively related to habitat patch size. Efforts in the managed forest should be directed towards preserving and creating larger patches of living and dead aspen trees and increasing the amount of aspen at the landscape level.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Habitat loss and fragmentation due to human land use are major threats to biodiversity (Ehrlich 1988; Baille et al. 2004). Decreasing habitat patch size and increasing patch isolation negatively affect species richness (MacArthur and Wilson 1967; Drakare et al. 2006; Watling and Donelly 2006). Also the occupancy (i.e., if present or absent) of individual species, that underlie the diversity patterns, has been demonstrated to be affected by habitat patch size and isolation. With decreasing patch size and increasing isolation the probability that a patch is occupied may decrease (Prugh et al. 2008). In a synthesis including 1,015 bird, mammal, reptile, amphibian and invertebrate population networks, patch area was a better predictor of occupancy than isolation: area alone accounted for 13% of the deviance in occupancy versus 3% accounted for isolation alone. A positive relationship between habitat patch size and occupancy may be caused by smaller patches having lower immigration rates and higher extinction rates (Root 1973, Hanski 1999). However, it could also just be an effect of small habitat patches being samples of larger ones, because even if the density is the same in small and large patches, species occurring at low densities will be absent from small patches more often. This has been referred to as the random sample hypothesis (Andrén 1996). Rejection of the random sample hypothesis requires density data from both small and large patches.
In Fennoscandia, most of the forests have been under production management since the nineteenth or twentieth century (Östlund et al. 1997; Kouki et al. 2001), which has resulted in a lower amount of coarse woody debris (CWD) compared to natural forests (Linder and Östlund 1998). Surveys in Fennoscandian forests suggest that the volume of CWD has been reduced by 90–98% because of forestry (Siitonen 2001). In accordance with this, populations of many saproxylic (dependent on dead wood) species have declined (Gärdenfors 2005). In the present study we analyse the occurrence of saproxylic beetles associated with aspen (Populus tremula). In Fennoscandian boreal forests, aspen is one of the most important tree species for biodiversity; for instance, in Finland it harbours a higher proportion of critically endangered species than any other tree species (Tikkanen et al. 2006). In Fennoscandia, aspen has been regarded as a pest species and was killed with herbicides until recently. In the longer perspective there is also a concern for a low regeneration rate of aspen. Fire suppression (Zackrisson 1985) and high browsing pressure (Angelstam et al. 2000; Kouki et al. 2004) have been suggested as explanations for the low rate of aspen regeneration (but see Zakrisson et al. 2007). Thus there is a risk for a future bottleneck for aspen-associated species even in areas that currently contain mature aspen stands (Kouki et al. 2004). Aspen has an aggregated growth pattern because of its ability to generate asexually by suckering and as a consequence aspen CWD is spatially clustered in patches differing in size from single dead trees to large groups of dead trees.
In this study we analyse the importance of habitat patch size, and other patch characteristics, and habitat connectedness, for the occupancy and density of individual saproxylic beetles associated with aspen. We also test if the proportion of the summed beetle individuals constituted by other species (i.e., not aspen-associated) are related to patch size. This was done because the density of generalist species may increase in smaller patches which may negatively affect habitat specialists (Ås 1993; Burke and Nol 2000; Henle et al. 2004).
Materials and methods
Study area
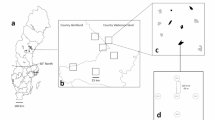
The study was carried out in a managed forest landscape north-west of Delsbo in the province of Hälsingland, central Sweden (62°N, 13°E). The landscape is situated in the mid-boreal vegetation zone (Ahti et al. 1968). The size of the study landscape is 45,000 ha. The forest is dominated by Scots pine (Pinus silvestris L.) and Norway spruce (Picea abies Karst.). One forest company (Holmen Skog AB) almost exclusively owns the land, and the study landscape is also included in the same management unit, and therefore the management regime is similar over the entire area. Since 1998, management follows the FSC-standard (Anon. 2000). During 1995 and 2000 most living aspen trees were retained at final cuttings (hereafter denoted clearcuts). After 2000, this green-tree retention was mainly limited to leaving groups of living trees (P.G. Jacobsson pers. comm.). Based on the interpretation of aerial photography (see below) living aspen trees covered 1.33% of the forest area in the landscape and 0.98% of the total area.
Patch selection
On the 30th of May 2002, 27,500 ha of the 45,000 ha study landscape were photographed using infrared aerial photography at a height of 4,600 m. The photographs were taken just after leaf-flush, to enable identification of aspen. Crown cover of mature aspen (canopy height ≥20 m) was interpreted from the photographs. Polygons were formed for areas interpreted as mature aspen and these polygons were classified based on percentage of aspen crown cover.
Based on the interpretation of the aerial photographs and on information from the forest company 42 patches with aspen were selected within the photographed landscape and an additional 14 patches outside the photographed landscape. Of these 32 were situated on clearcuts and 24 in closed forest stands. The clearcut sites had been cut during the last 13 years. The minimum distance between selected patches was at least 500 m except in two cases. Delimitation of forest patches was based on aspen crown cover polygons. For patches on clearcuts the whole clearcut area was included in the patch size (since aspen trees may have been cut during final felling).
Estimation of habitat connectedness
Crown cover of living mature aspen trees was used as an estimate of habitat connectedness for saproxylic beetles associated with aspen. Thus, we assumed that there is a correlation between the crown cover of living aspen and the amount of dead aspen trees. For the 42 patches within the photographed landscape, the habitat connectedness (area of aspen crown cover) in the surrounding landscape was calculated at 3 different spatial scales (buffers with radius of 500 m, 750 m and 1,000 m) using ESRI ArcMap (v. 8.2). For intersections partly outside the aerial photography area the mean aspen crown cover was calculated for the part located inside the photographed area, which was then assumed to be of equal composition as the part outside. Only patches where at least 60% of the intersection was inside of the aerial photography area were used in the calculations. Mean values and ranges of connectedness are listed in Table 1.
Measurements of aspen CWD in patches
All aspen CWD with a diameter ≥10 cm, and standing CWD with a height ≥50 cm were recorded for each patch. Diameter, length for lying CWD, height for snags and percentage of surface area covered by bark (in 10% classes) was measured for each individual CWD object. For a more detailed description of the CWD estimates see Sahlin and Ranius (2009). As we made a total survey of aspen CWD at every patch, we could obtain direct estimates of the bark areas and volumes per patch. Mean values and ranges for patch variables are listed in Table 1.
Beetle sampling
We used direct sampling of beetles in aspen CWD. With direct sampling we can be sure that recorded species really have colonised the CWD in our sampled patches and it is also possible to estimate the proportion of occupied CWD objects within patches. Two methods of direct sampling were used: (i) recording presence of species-specific emergence holes and larval galleries of Xylotrechus rusticus, Saperda perforata and Ptilinus fuscus on all dead wood objects of aspen in each patch, and (ii) sieving of 0.5 m2 bark on 10 randomly selected dead aspen trunks per patch. In patches with less than 10 aspen CWD objects with enough bark for a 0.5 m2 sample, all available objects were sieved (but still a 0.5 m2 sample per object). Whenever possible, both standing and lying CWD objects were sieved. The sieving was done in late September to early October in 2002–2004. Autumn is ideal for sampling by sieving because it is the period when the highest number of beetle species is present under the bark (Wikars et al. 2005). Sieved samples were extracted using Tullgren funnels (30 cm wide, 8 mm mesh size) (New 1998). 60 W light bulbs were used as heat and light source, and extraction lasted for at least 24 h. All adult beetles, along with the larvae of Endomychus coccineus, were determined to species.
Analyses
Only saproxylic beetles were included in the analyses. The species were divided into two groups: aspen-associated and generalist species. All species that according to Palm (1959) and Dahlberg and Stokland (2004) prefer aspen CWD over other tree species were classified as aspen-associated. The remaining species were classified as generalist species (i.e., species preferring other tree species than aspen).
Single species analyses were conducted for nine aspen-associated species that fulfilled the requirement of being recorded from at least ten CWD objects. Analyses of species richness included all 13 aspen-associated species that were recorded in the study. The analyses were conducted in three steps. (i) Both patch variables and habitat connectedness were included as explanatory variables for occupancy of single species and species richness. This analysis included only the 42 patches for which connectedness data (i.e., aspen crown cover) was available. (ii) Only patch size, defined as area with or without bark on aspen CWD, was included as explanatory variable because this was the dominant significant variable in the first analysis. In addition to occupancy also densities (proportion colonised CWD objects and densities of individuals) within patches was related to patch area. These analyses included all the 56 sampled patches (i.e., also those for which connectedness data was lacking). (iii) Total densities of individuals of aspen-associated and generalist species were related to habitat patch size.
The influence of patch characteristics and connectedness for occupancy of each one of the nine species was analysed with logistic regression and for total aspen-associated species richness with multiple linear regression. Many of the patch characteristics were correlated with each other (Table 2). Of the five variables reflecting habitat patch size (VOLUME, BARK, NOBARK, CWDNUMBER, AREA; see Table 1 for explanations) BARK and NOBARK were chosen because they best describe the available habitat amount for the studied aspen-associated species. All species except one (Ptilinus fuscus) mainly develop under the bark (Freude et al. 1967, 1969; Švacha and Danilevsky 1988; Koch 1989, 1992; Klausnitzer 2001). For P. fuscus the mantle area without bark (NOBARK) was used instead because it breeds in wood without bark (Palm, 1959, Freude et al. 1969). Thus, the patch characteristics included in the regressions were BARK or NOBARK, STANDING (percentage of standing CWD) and CC/FOR (clearcut or forest patch). The habitat connectedness measures were included one at a time in the regressions because they were strongly correlated with each other. In the measures of habitat connectedness the habitat amounts in the focal patches were not included. All proportion values (STANDING, A500, A750 and A1000) were arcsine square-root transformed.
The influence of habitat patch size alone (expressed as area of bark (BARK) or without bark (NOBARK) on aspen CWD) on occupancy was analysed with logistic regression, while influence on proportion of occupied objects and density of individuals was analysed with Kendall’s tau B. Estimates of mean proportion of occupied objects and mean densities included also patches without occurrences. Density data was only available for species sampled by sieving. Relationship between patch size and species richness was analysed with linear regression. We also used volume of aspen CWD as a predictor because of its earlier use in studies of species richness in saproxylic species.
Results
A total of 4,091 individuals belonging to 140 saproxylic beetle species, were collected by sieving. Of these, 1,018 individuals (25%) belonging to 10 species (7%) were aspen-associated (i.e., prefer aspen over other tree species) (Table 3). Three additional aspen-associated species were recorded from galleries on CWD objects (Table 3). Three of the species are included in the Swedish red list (C. curtula, M. fulvicollis and T. asperatus), all in the category near threatened (Gärdenfors 2005).
Patch characteristics and habitat connectedness
There was a significant positive relationship between habitat patch size (area with or without bark on aspen CWD) and occupancy of A. bicolor, H. plana, P. fuscus, M. fulvicollis, C. ferrugineum and E. coccineus (Table 4). In addition, patch type (forest or clearcut) was significantly related to occupancy of X. rusticus, favouring clearcuts. Also species richness was positively related to habitat patch size (Table 4). Connectedness was not retained as a significant variable for any of the species or for species richness.
Patch size alone (extended data set)
Six of the nine species (H. plana, C. curtula, P. fuscus, E. coccineus, M. fulvicollis and X. rusticus) showed a significant positive relationship between habitat patch size and occupancy for the extended data set (Table 5, Fig. 1). For all these species, except C. curtula, there was also a significant positive relationship between patch size and proportion of occupied CWD objects (Table 5, Fig. 2). Density of individuals was significantly related to patch size for H. plana, E. coccineus and M. fulvicollis (Table 5). Species richness was significantly related to patch size defined as area bark and as volume of aspen CWD (Figs. 3 and 4).

Occupancy (proportion of occupied patches) in relation to habitat patch size for: A Homalota plana, B Cyphaea curtula, C Ptilinus fuscus, D Endomychus coccineus, E Mycetophagus fulvicollis and F Xylotrechus rusticus. Habitat patch size is defined as the area of bark on aspen CWD. N = 8 for <10 m2, N = 13 for 10–20 m2, N = 12 for 20–50 m2, N = 15 for 50–100 m2 and N = 8 for 100–180 m2. For P. fuscus habitat patch size is defined as surface area without bark on aspen CWD. N = 10 for <10 m2, N = 6 for 10–20 m2, N = 15 for 20–50 m2, N = 12 for 50–100 m2 and N = 13 for 100–400 m2

Mean proportion of occupied CWD objects and densities per patch in relation to patch size for: A Homalota plana, B Ptilinus fuscus, C Endomychus coccineus, D Mycetophagus fulvicollis and E Xylotrechus rusticus. Habitat patch size is defined as the area of bark on aspen CWD for all species except P. fuscus. For P. fuscus habitat patch size is defined as surface area without bark on aspen CWD. For number of replicates see legend of Fig. 1

Relationship between habitat patch size, expressed as bark area on aspen CWD, and number of aspen-associated saproxylic beetle species. Each point represents one habitat patch. P < 0.001, N = 56

Relationship between habitat patch size, expressed as volume of aspen CWD, and number of aspen-associated saproxylic beetle species. Each point represents one habitat patch. P < 0.001, N = 56
Generalist species
For the mean density of the summed individuals of generalist species (i.e., the saproxylic species that were not defined as aspen-associated) there was no significant relationship with habitat patch size (Kendall’s tau B = −0.020, P = 0.829, N = 54). The mean density of aspen-associated species was close to significantly related to patch size (B = 0.182, P = 0.054). Generalist species constituted a significantly larger proportion of the total density in smaller habitat patches than in larger patches (B = 0.224, P = 0.018) (Fig. 5).

Relationship between habitat patch size, expressed as bark area on aspen CWD, and the mean proportion of the total density of saproxylic beetles constituted by aspen-associated beetles
Discussion
In this study we demonstrate that habitat patch size, measured as area with or without bark on aspen CWD, influence occurrence patterns of saproxylic aspen-associated beetles. For a majority of the studied species there was a significant positive relationship between habitat patch size and density (proportion of occupied CWD objects and density of individuals). For these species we can thus reject the random sample hypothesis as an explanation for the observed positive relationship between occupancy and patch size.
Three processes that may have contributed to the observed density–area relationships are: (i) Higher extinction risk in small than in large patches resulting in a higher proportion of empty small patches. (ii) Patch size per se may influence immigration and emigration rates. The resource concentration hypothesis, formulated by Root (1973) predicts that specialist herbivores should reach higher densities in large patches as a result of larger emigration rates from smaller patches and larger immigration rates into larger patches. Later studies have explored how the rate of successful migration into habitat patches differing in size may be a function of search behaviour, mode of patch detection and mobility (e.g., Bowman et al. 2002; Hambäck and Englund 2005; Englund and Hambäck 2007). (iii) The density of generalist species may increase in smaller patches which may negatively affect habitat specialists (Ås 1993; Burke and Nol 2000; Henle et al. 2004).
Our data support the hypothesis that extinctions are likely to occur frequently in smaller patches of aspen CWD. Based on the densities in occupied patches the total number of individuals in a patch with 20 m2 bark (maximum size in the second smallest class in Fig. 2) should be only 20–40 for M. fulvicollis, H. plana and E. coccineus. In addition, in such small patches suitable breeding substrate may not be continuously available. Regarding search behaviour, mode of patch detection and dispersal capacity, nothing is known about the studied species. We found some support for the hypothesis that generalist species have negatively influenced the colonisation of aspen-associated species in the smaller habitat patches. The proportion of the total density of saproxylic beetle individuals constituted by generalist species was positively related to habitat patch size. Smaller patches, in which the probability for local extinctions of aspen-associated species can be expected to be high, may be colonised rapidly by generalist species, which could result in a competitive disadvantage for the aspen-associated species.
Based on modelling and empirical studies (including birds and mammals) a threshold value of about 10–30% habitat, below which true fragmentation effects on population persistence will occur, has been suggested (Andrén 1994; Fahrig 1998). The habitat amount in the present study is much lower than this threshold. If we use the percentage of the basal area of living forest constituted by aspen as a measure of habitat amount (no data available for aspen CWD), aspen constitutes less than 0.5% in our study landscape (Anon. 2009). In addition, our study landscape currently holds much lower amount of habitat compared to some other regions in Sweden that contain more than 15% of aspen (Anon. 2009) and in which areas known to host a rich fauna of aspen-associated saproxylic beetles are located (Lundberg and Gustafsson 1995). Our study area has also experienced a 70% decrease in area of old deciduous forest from 1967 to 2002 (data based on interpretation of aerial photographs from 1967 and the aerial photographs of this study) (L.-O. Wikars pers. comm.).
The species demonstrating a positive relationship between patch size and proportion of occupied objects or density differs regarding preferences for breeding material. One species, P. fuscus, breeds in CWD without bark while the other species utilise CWD with bark (Freude et al. 1967; Freude et al. 1969; Švacha and Danilevsky 1988; Koch 1989; Koch 1992; Klausnitzer 2001). Three of the species (A. bicolor, E. coccineus and M. fulvicollis) inhabits fungi-infected substrate. The variety of species responding to patch size implies that our results may hold also for other aspen-associated beetles.
Only a few earlier studies have explored the relationship between habitat patch size and the densities of saproxylic insect species. In a study by Ranius (2002, see also Ranius and Fahrig 2006) the percentage of occupied hollow oak trees increased with increasing number of hollow oaks per stand for three (Osmoderma eremita, Elater ferrugineus and Tenebrio opacus) of eleven studied species. In a study by Komonen et al. (2000) the proportion of fruiting bodies of the fungus Fomitopsis rosea colonised by the moth Agnathosia mendicella was significantly lower in fragments of old-growth spruce swamp forests than in large continuous stands. In contrast, there was no relationship between storm gap size and percentage of colonised storm-felled spruces or densities of colonising individuals of the bark beetle Ips typographus (Eriksson et al. 2005; Schroeder 2010). The bark beetle system differs from the other systems (including the present study) by a much higher habitat amount. Spruce is one of the dominant tree species in Sweden while there are only isolated stands of hollow oaks (Ranius 2002) and old-growth spruce swamp forests. In addition, the dispersal rate of O. eremita (dispersal rate not studied for the other species) (Ranius and Hedin 2001) is low while it is high for I. typographus (Forsse and Solbreck 1985).
The positive relationship between richness of aspen-associated species and habitat patch size is a result of both demographic processes and environmental heterogeneity. The positive relationship for occupancy for many of the species has clearly contributed to the observed relationship for species richness. The beetle species have somewhat different substrate preferences (Sahlin and Ranius 2009) which in combination with a higher heterogeneity of aspen CWD in larger patches (with more sampled CWD objects) should result in higher species richness in larger patches. Also in a study by Ranius (2002), on saproxylic beetles inhabiting hollow oaks, there was a positive relationship between species richness and patch size (number of hollow oaks in each stand).
The sieving regime (all CWD objects with bark sieved if ten or less, a maximum of ten if more than ten CWD objects with bark) means that estimates of occupancy for single species and species richness are more precise for small habitat patches than for larger patches. If a higher proportion of the CWD objects had been sieved in the larger patches the positive relationships between patch size and occupancy and patch size and species richness could have been even stronger. For the three species recorded from presence of galleries no bias exists because all CWD objects were inspected. In contrast to occupancy and species richness, the proportion of occupied objects and densities of beetle individuals are not influenced by sample effort.
Connectedness was not retained as a significant predictor of either occupancy or species richness. The stronger effect of patch size compared with connectedness may be caused by: (i) crown cover of living aspen trees may be a poor estimate of aspen CWD, (ii) our connectedness measure gives no information about which habitat patches that are occupied and thus can function as sources of immigrants (Prugh et al. 2008), (iii) most species were not strictly associated to aspen but could also reproduce in other tree species, (iv) the amount of deciduous forest (including aspen) has decreased strongly in the last decades and occurrence patterns may thus still be more related to historical habitat distribution than to current, and (v) the tested scales may have been too small.
Management implications
This study shows that the density of aspen-associated beetle species increases with habitat patch size. Thus, efforts in the managed forest should be directed towards creating larger patches of living and dead aspen trees and to increase the amount of aspen at the landscape level. Living aspen trees in such patches should be retained during thinning and clearcutting operations and care should be taken not to destroy already existing aspen CWD (Hautala et al. 2004; Ranius and Kindvall 2004). The latter could be achieved by careful planning of logging and scarification operations to avoid destruction of aspen CWD. When creating new aspen CWD (e.g., high stumps or cut trees) localities with already existing aspen CWD should be favoured over localities without aspen CWD because this will result in on average larger patches of aspen CWD.
Abbreviations
- CWD:
-
Coarse woody debris
References
Ahti T, Hämet-Ahti L, Jalas J (1968) Vegetation zones and their sections in Northwestern Europe. Annales Botanici Fennici 5:169–211
Andrén H (1994) Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355–366
Andrén H (1996) Population responses to habitat fragmentation: statistical power and the random sample hypothesis. Oikos 76:235–242
Angelstam P, Wikberg P-E, Danilov P, Faber WE, Nygrén K (2000) Effects of moose density on timber quality and biodiversity restoration in Sweden, Finland, and Russian Karelia. Alces 36:133–145
Anonymous (2000) Svensk FSC-standard för certifiering av skogsbruk. Svenska FSC-Rådet, Uppsala
Anonymous (2009) Forestry statistics 2009. Official statistics of Sweden. Swedish University of Agricultural Sciences, Umeå
Ås S (1993) Are habitat islands islands? Woodliving beetles (Coleoptera) in deciduous forest fragments in boreal forest. Ecography 16:219–228
Baille J, Hilton-Taylor C, Stuart SN (2004) 2004 IUCN red list of threatened species: a global assessment. International Union for Conservation of Nature, Cambridge, UK
Bowman J, Cappuccino N, Fahrig L (2002) Patch size and population density: the effect of immigration behaviour. Conserv Ecol 6:9
Burke DM, Nol E (2000) Landscape and fragment size effects on reproductive success of forest-breeding birds in Ontario. Ecol Appl 10:1749–1761
Dahlberg A, Stokland JN (2004) Vedlevande arters krav på substrat. Rapport No. 7. Skogsstyrelsen, Jönköping
Drakare S, Lennon JJ, Hillebrand H (2006) The imprint of the geographical, evolutionary and ecological context on species-area relationships. Ecol Lett 9:215–227
Ehrlich PR (1988) The loss of diversity: causes and consequences. In: Wilson EO (ed) Biodiversity. National Academic Press, Washington, DC, pp 21–27
Englund G, Hambäck PA (2007) Scale dependence of immigration rates: models, metrics and data. J Anim Ecol 76:30–35
Eriksson M, Pouttu A, Roininen H (2005) The influence of windthrow area and timber characteristics on colonization of wind-felled spruces by Ips typographus (L.). For Ecol Manage 216:105–116
Fahrig L (1998) When does fragmentation of breeding habitat affect population survival? Ecol Modell 105:273–292
Forsse E, Solbreck C (1985) Migration in the bark beetle Ips typographus L.: duration, timing and height of flight. Zeitschrift für angewandte Entomologie 100:47–57
Freude H, Harde WK, Lohse GA (1967) Die Käfer Mitteleuropas, Band 7. Goecke & Evers Verlag, Krefeld
Freude H, Harde WK, Lohse GA (1969) Die Käfer Mitteleuropas, Band 8. Goecke & Evers Verlag, Krefeld
Gärdenfors U (2005) The 2005 red list of Swedish species. ArtDatabanken, SLU, Uppsala
Hambäck PA, Englund G (2005) Patch area, population density and the scaling of migration rates: the resource concentration hypothesis revisited. Ecol Lett 8:1057–1065
Hanski I (1999) Metapopulation ecology. Oxford University Press, Oxford
Hautala H, Jalonen J, Laaka-Lindberg S, Vanha-Majamaa I (2004) Impacts of retention felling on coarse woody debris (CWD) in mature boreal spruce forests in Finland. Biodivers Conserv 13:1541–1554
Henle K, Davies KF, Kleyer M, Margules C, Settele J (2004) Predictors of species sensitivity to fragmentation. Biodivers Conserv 13:207–251
Klausnitzer B (2001) Die Larven der Käfer Mitteleuropas, Band 6. Spektrum Akademisher Verlag GmbH, Heidelberg
Koch KC (1989) Die Käfer Mitteleuropas - Ökologie, Band 1. Goecke & Evers Verlag, Krefeld
Koch KC (1992) Die Käfer Mitteleuropas - Ökologie, Band 3. Goecke & Evers Verlag, Krefeld
Komonen A, Penttilä R, Lindgren M, Hanski I (2000) Forest fragmentation truncates a food chain based on a old-growth forest bracket fungus. Oikos 90:119–126
Kouki J, Löfman S, Martikainen P, Rouvinen S, Uotila A (2001) Forest fragmentation in Fennoscandia: linking habitat requirements of wood-associated threatened species to landscape and habitat changes. Scand J For Res Suppl 3:27–37
Kouki J, Arnold K, Martikainen P (2004) Long-term persistence of aspen—a key host for many threatened species—is endangered in old-growth conservation areas in Finland. J Nat Conserv 12:41–52
Linder P, Östlund L (1998) Structural changes in three mid-boreal Swedish forest landscapes, 1885–1996. Biol Conserv 85:9–19
Lundberg S, Gustafsson B (1995) Catalogus Coleopterorum Sueciae. Naturhistoriska Riksmuseet, Stockholm
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
New TR (1998) Invertebrate surveys for conservation. Oxford University Press, Oxford
Östlund L, Zackrisson O, Axelsson AL (1997) The history and transformation of a Scandinavian boreal forest landscape since the 19th century. Can J For Res 27:1198–1206
Palm T (1959) Die Holz- und Rindenkäfer der Süd- und Mittelschwedischen Laubbäume. Opuscula Entomologica Supplementum XVI, Lund
Prugh R, Hodges KE, Sinclair ARE, Brashares JS (2008) Effect of habitat area and isolation on fragmented animal populations. PNAS 105:20770–20775
Ranius T (2002) Influence of stand size and quality of tree hollows on saproxylic beetles in Sweden. Biol Conserv 103:85–91
Ranius T, Fahrig L (2006) Targets for maintenance of dead wood for biodiversity conservation based on extinction thresholds. Scand J For Res 21:201–208
Ranius T, Hedin J (2001) The dispersal rate of a beetle, Osmoderma eremita, living in tree hollows. Oecologia 126:363–370
Ranius T, Kindvall O (2004) Modelling the amount of coarse woody debris produced by the new biodiversity-oriented silvicultural practices in Sweden. Biol Conserv 119:51–59
Root RB (1973) Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecol Monogr 43:95–124
Sahlin E, Ranius T (2009) Habitat availability in forests and clearcuts for saproxylic beetles associated with aspen. Biodivers Conserv 18:621–628
Schroeder LM (2010) Colonisation of storm gaps by the spruce bark beetle—influence of gap and landscape characteristics. Agric For Entomol (in press)
Siitonen J (2001) Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example. Ecol Bull 49:11–41
Švacha P, Danilevsky ML (1988) Cerambycoid Larvae of Europe and Soviet Union (Coleoptera, Cerambycoidea)—part 2. Acta Universitatis Carolinae - Biologica 31:121–284
Tikkanen OP, Martikainen P, Hyvärinen E, Junninen K, Kouki J (2006) Red-listed boreal forest species of Finland: associations with forest structure, tree species, and decaying wood. Ann Zool Fenn 43:373–383
Watling JI, Donelly MA (2006) Fragments as islands: a synthesis of faunal responses to habitat patchiness. Conserv Biol 20:1016–1025
Wikars LO, Sahlin E, Ranius T (2005) A comparison of three methods to estimate species richness of saproxylic beetles (Coleoptera) in logs and high stumps of Norway spruce. Can Entomol 137:304–324
Zackrisson O (1985) Some evolutionary aspects of life history characteristics of broadleaved tree species found in the boreal forest. In: Hägglund B, Peterson G (eds) Broadleaves in boreal silviculture-An obstacle or an asset?. Department of Silviculture, Swedish University of Agricultural Sciences, Umeå, pp 17–36
Zakrisson C, Ericsson G, Edenius L (2007) Effects of browsing on recruitment and mortality of European aspen (Populus tremula L.). Scand J For Res 22:324–332
Acknowledgements
Atte Komonen and Thomas Ranius provided valuable comments on the manuscript. Support for this study came from the projects ‘Conservation of Biodiversity in Managed Forests’ (grants to Stig Larsson and Kjell Danell) financed by the Faculty of Forestry at The Swedish University of Agricultural Sciences (SLU). Holmen Skog AB generously provided maps, aerial photographs and hosted our field study. P.G. Jacobsson and Ola Kårén from Holmen Skog AB provided important information about the study landscape. Kalle Sahlin, Kurt Sahlin, Lena Wedmo, Robert Wedmo, Jan ten Hoopen and David Isaksson assisted in the field.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sahlin, E., Schroeder, L.M. Importance of habitat patch size for occupancy and density of aspen-associated saproxylic beetles. Biodivers Conserv 19, 1325–1339 (2010). https://doi.org/10.1007/s10531-009-9764-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-009-9764-6