
Abstract
The Rideau Canal (Ontario, Canada) was constructed in the early 1830s, primarily as a means to transport military personnel, but now is primarily recreational. The construction of the canal and associated flooding, as well as other land-use changes, likely impacted lakes within the system, however, long-term monitoring data are not available. Furthermore, recent environmental changes, including accelerated climate warming, are affecting lake ecosystems. Shallow, macrophyte-dominated Lake Opinicon, which is part of the canal system, has been impacted by other various catchment disturbances over the past ~ 200 years. A previous diatom-based paleolimnological study conducted on the lake, examining a core collected in 1995, found that the diatom responses to a host of large-scale catchment disturbances were moderate compared to nearby deeper lakes. A more recent diatom-based study conducted on a 2019 core (the same core used in the present study) found similar results; however, over the most recent ~ 25–30 years a striking shift in diatom assemblage composition was documented, coinciding with increased regional climate warming. Nothing is known concerning long-term changes within the primary consumers linked to the array of disturbances. Here, we examined changes in cladoceran assemblages over the past ~200 years, using the 2019 sediment core, to track their response to various environmental stressors including climate warming. We found that pelagic Bosmina and Daphnia species began to increase in the early nineteenth century, consistent with the flooding of the lake during canal construction. The most ecologically notable changes in the cladoceran record, however, occurred in the most recent sediments. These were characterized by marked declines (often to trace abundances) in several littoral taxa, concurrent with a further increase in the relative abundance of small, pelagic Bosmina spp. This most recent compositional shift was consistent with accelerated regional climate warming and associated limnological changes, decreased total phosphorus (TP) concentrations, and changing food sources. These changes in primary consumers will likely cascade throughout the food web.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Rideau Canal is a 202-km long waterway that connects the cities of Ottawa and Kingston, ON. Following five years of construction, the canal was completed in 1832. Originally designed for the transport of military personnel and equipment, it now primarily serves recreational purposes (Bergman et al. 2022). Prior to canal construction, the surrounding vegetation was characterized by mixed woodlands and wetlands (Karst and Smol 2000). However, forests along the watershed experienced severe deforestation as the military required timber for the construction of numerous locks and damming of the waterways (CHPC 1996). Extensive flooding of surrounding lakes was required to provide navigable depths for boat transport, increasing the depths of the impacted lakes, including Lake Opinicon (L. Opinicon), by ~ 2–3 m (Forrest et al. 2002).
Numerous ecologically significant disturbances within the catchment area of the Rideau Canal system have exerted a considerable influence on water quality over the past ~ 200 years. Paleolimnological evidence indicates that many lakes within this system were significantly affected by canal construction (Christie and Smol 1996; Forrest et al. 2002). For example, deeper lakes such as Upper Rideau Lake, experienced increased nutrient loading (Forrest et al. 2002; Balasubramaniam 2022), while nearby shallow and macrophyte-dominated L. Opinicon was largely resistant to the effects of canal construction and other cultural disturbances, likely due to its shallow nature and extensive macrophyte cover (Karst and Smol 2000; Balasubramaniam et al. 2023), as described by Scheffer et al. (1993).
More recently, accelerated anthropogenic climate warming has become an important stressor affecting this region. Mean annual air temperature (MAAT) data collected since 1873 at the Kingston (Ontario) weather station, located ~ 60 km south of L. Opinicon, indicated an increase of ~ 2.34 °C in the past 148 years, that has markedly accelerated in recent decades (Nelligan et al. 2016; Balasubramaniam et al. 2023). With increasingly warmer conditions, many lakes in Ontario are experiencing longer ice-free seasons, resulting in earlier onset and longer duration of thermal stratification (Stainsby et al. 2011). With accelerated warming, L. Opinicon has shifted over the last ~ 20 years from a polymictic to a dimictic system (albeit often weakly stratified) with concurrent marked changes in diatom assemblage composition (Balasubramaniam et al. 2023). While paleolimnological data are available for long-term changes in diatoms (Karst and Smol 2000; Balasubramaniam et al. 2023) and chironomids (Little and Smol 2000) in L. Opinicon, little is known about how primary consumers, such as Cladocera, have responded to past cultural disturbances, as well as climate-related changes.
Cladocera are key components of aquatic food webs, occupying an intermediate trophic position between top-down regulators (e.g., fish and invertebrate predators) and bottom-up factors (e.g., nutrients, algae) (Korosi and Smol 2011). Cladoceran species composition is a strong environmental indicator of changes in habitat and trophic status (Nevalainen and Luoto 2017), chemical characteristics of lakes (DeSellas et al. 2008), top-down interactions with predators (Korosi et al. 2008), water-level changes (Wang et al. 2020), and climate warming (Thienpont et al. 2015; Labaj et al. 2018). Cladocera are also excellent paleolimnological indicators as their chitinous exoskeletons are often identifiable to the species-level and preserve well in lake-sediment records (Korhola and Rautio 2001).
Based on previous paleolimnological studies using diatoms (Karst and Smol 2000; Balasubramaniam et al. 2023), we anticipated that, during the period of canal construction (including the flooding of L. Opinicon), the increased water depth and the doubling in the length of the lake, together with other catchment disturbances (e.g., land clearance, settlement, increased agricultural activities), would be the main drivers of cladoceran assemblage changes in the earlier part of the sediment record. It has often been reported that with increased TP loading resulting from land clearance, pelagic cladoceran taxa may increase along with increases in primary production (Jeppesen et al. 2011; Bennion et al. 2015). However, the Cladocera response may be more nuanced in shallow macrophyte-dominated L. Opinicon, as previous diatom-inferred reconstructions found that TP concentrations did not change greatly following these substantial catchment disturbances (Karst and Smol 2000), and overall primary production declined following hydrological changes associated with canal construction (Balasubramaniam et al. 2023). For certain Cladocera taxa, the extensive macrophyte coverage in L. Opinicon may offer a refuge, which can help reduce predation pressure (Timms and Moss 1984; Burks et al. 2001) and provide additional sources of food (Tremel et al. 2000; Basińska et al. 2014). In addition, we anticipated that cladoceran assemblages would register a response to recent accelerated climate warming, as was observed in the diatom record (Balasubramaniam et al. 2023).
Using the same 210Pb-dated sediment core used by Balasubramaniam et al. (2023) for diatom analyses, this study explores long-term environmental changes in cladoceran assemblages in L. Opinicon over the past ~ 200 years in response to multiple environmental stressors, including the construction of the Rideau Canal (~ 1830) and twenty-first century accelerated regional climate warming. Moreover, we compare the timing and nature of the changes in these primary consumers to trends in diatom assemblage composition reported by Balasubramaniam et al. (2023) and explore possible environmental drivers (e.g., habitat changes from increased macrophyte cover, changes in food quality/quantity, and recent warming).
Site description
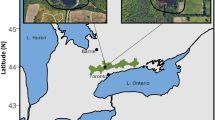
L. Opinicon (44.34°N, 76.19°W) is located approximately 60 km north of Kingston in Eastern Ontario (Fig. 1). It is a small (surface area: 7.85 km2), narrow (< 1.5 km wide), shallow (Zmean: 4.9 m, Zmax: 9.2 m), relatively long (~ 9.5 km), macrophyte-dominated lake (Karst and Smol 2000). Water flows into the lake from Indian, Hart, Warner, and Lower Rock lakes, and flows out to Sand Lake. The southern catchment is underlain by hard-weathering Precambrian Shield bedrock, in contrast to the northern region that is underlain by limestone (Karst and Smol 2000). Detailed algal and limnological data (Agbeti and Smol 1995a, 1995b; Agbeti et al. 1997; Balasubramaniam et al. 2023) have been gathered for this lake for many decades, as it is adjacent to the Queen’s University Biological Station. The lake is currently considered to be oligo-mesotrophic with a total phosphorus (TP) concentration of ~ 11 µg/L (measured in 2019). However, TP concentrations have declined significantly since the 1970s (Balasubramaniam et al. 2023), coinciding with declines in watershed agricultural activities (Warren 1997), improved soil-nutrient retention of secondary growth forests, and improvements in residential septic tank systems (Forrest et al. 2002). The lake has long been considered to be well-mixed, but in the past few decades it has experienced sustained periods of thermal stratification linked to accelerated warming (Balasubramaniam et al. 2023).

Image of L. Opinicon (Ontario, Canada). The locations of the 1994 (Karst and Smol 2000) and the 2019 (current study) sediment cores are labelled with stars. The Queen’s University Biological Station (QUBS), as well as nearby locks, are depicted in the image. Inset: Map of Canada with the approximate location of Lake Opinicon (red box) within the province of Ontario (green shading). Reference: created in R using package ggmap (Kahle and Wickham 2013)
Materials and methods
Sediment-sample collection
A 45-cm-long sediment core was retrieved from L. Opinicon (coring depth: 4.3 m) on October 5th, 2019, using a Glew (1989) gravity corer, as described in Balasubramaniam et al. (2023). The core was sectioned on site using a Glew (1988) vertical extruder into 0.5-cm contiguous intervals for the entire length of the core. Each sediment interval was placed into Whirl–Pak® bags and kept in a cooler until they were transported to the PEARL cold room where they were stored at ~ 4 °C. At the time of coring, Secchi depth and coring depth were recorded, and water samples were collected for chemical analysis. The sediment core was freeze-dried prior to analyses.
210Pb dating
As described in detail in Balasubramaniam et al. (2023), 210Pb gamma spectrometry techniques closely followed the methods outlined by Schelske et al. (1994). Strategically selected intervals from the sediment core (26 intervals) were prepared by weighing ~ 0.3 g of the freeze-dried material into plastic tubes at a height of ~ 2 cm. Based on unsupported 210Pb activity (total 210Pb minus supported 210Pb), a chronology was generated using 214Pb as a proxy for background activities and the constant rate of supply (CRS) model (Appleby 2001), executed through the ScienTissiME package (http://www.scientissime.net/software; Scheer Software Solutions, Barry’s Bay, ON, Canada). In addition, 137Cs was examined as a potential independent chronological marker of the 1963 global moratorium on nuclear weapons testing to validate the 210Pb results. Dates of interest beyond background 210Pb (e.g., 1830 canal construction) were extrapolated using third-order polynomial models (these dates are viewed with caution as associated errors may be high). Detailed CRS dating results for this core are reported in Balasubramaniam et al. (2023).
Instrumental records
Trends in mean annual air temperature (MAAT) measured at the Kingston climate station (44°13′24.000″ N, 76°35′58.000″ W) between 1873 and 2021 were obtained from Environment and Climate Change Canada (ECCC, station ID: 6,104,142 and 6,104,146) using adjusted and homogenized climate data (Vincent et al. 2020). To fill in data gaps in the Kingston temperature record (during WWII and the 1970s), data from the nearby Hartington weather station (ECCC, ID: 6,103,367) were used as MAAT data from these two stations were highly correlated (r = 0.82; p < 0.001; Balasubramaniam et al. 2023). MAAT were presented as anomalies relative to the 1951-1980 mean (Hansen et al. 2010).
Cladoceran analysis
Twenty-three sediment samples were processed for Cladocera subfossil remains following the methods described by Frey (1986) and Korhola and Rautio (2001). We used ~ 0.2 g of freeze-dried sediment for each depth interval and then deflocculated the sediment by heating it in 10% potassium hydroxide (KOH) solution to ~ 80 °C for 30 min. We then passed each sample through a sieve (38 μm) and rinsed with deionized water to remove small particles and KOH. Ethanol and safranin were used to preserve and stain the samples. We applied one aliquot (100-µL each) of the concentrated remains to each slide followed by glycerin jelly and a coverslip.
The identification of cladoceran subfossils generally followed Korosi and Smol (2012a, b). We identified cladoceran remains under 200× or 400× magnification using a DMR2 microscope (Leica® Microsystems, Wetzlar, Germany) and bright-field optics. We counted all cladoceran remains including carapaces, headshields, and postabdominal claws and used the most abundant remain to calculate the number of individuals per taxa. Coverslips were scanned in their entirety for all identifiable remains and a minimum of 70 individuals for each interval were enumerated and expressed as % relative abundance. We combined Bosmina longirostris O. F. Müller and Eubosmina longispina Leydig into Bosmina spp. for analyses. Daphnia spp. included the Daphnia longispina complex O. F. Müller and the Daphnia pulex complex Leydig. Chydorus spp. included Chydorus brevilabris Frey, Chydorus piger Sars, and Chydorus gibbus Leach. As well, we combined Alona affinis Leydig, Alona circumfimbriata Megard, and Alona quadrangularis O. F. Müller into Alona spp., and Alonella nana Baird and Alonella excisa Fisher into Alonella spp. for analyses.
Relative abundances of the most common cladoceran taxa (greater than 2% in at least two intervals) were displayed stratigraphically and ordered by habitat (i.e., pelagic, littoral, and benthic). To quantitatively display the stratigraphic succession of cladoceran assemblages over time, taxa were organized from left to right in the figure based on increasing species scores from a canonical correspondence analysis (CCA) constrained to sediment core depth (Janssen and Birks 1994). The resulting species scores approximate each taxon’s weighted average mean or “optimum” in relation to core depth (age) and were subsequently ordered from lowest to highest, providing a numerical means to order taxa stratigraphically (Janssen and Birks 1994). Cladocera stratigraphic zones were established through constrained incremental sum of squares (CONISS) cluster analysis using the program TGView version 2.0.2 (Grimm 1991), with important zones identified by broken-stick analysis (Bennett 1996) using the vegan package (Oksanen et al. 2015) in R (version 3.6.1, R Development Core Team 2018).
VRS-chlorophyll-α trends
Visible reflectance spectroscopy-inferred chlorophyll-α (VRS-Chla) analysis was used to examine trends in whole-lake primary production, as explained in detail in Balasubramaniam et al. (2023). Briefly, following methods described in Wolfe et al. (2006) and Michelutti et al. (2010), approximately 0.2 g of freeze-dried sediment from each sediment interval of the L. Opinicon core was sieved through a 125-µm mesh, placed into glass cuvettes, and processed with a Model 6500 series Rapid Content Analyzer (FOSS NIRSystems, Inc.). Trends in chlorophyll a concentrations and its main diagenetic products were inferred from spectral absorbance values of wavelengths between 650 and 750 nm, using log-transformed data from Michelutti et al. (2010). Long-term trends in VRS-Chla were expressed as Z-scores.
Comparison among paleo proxy trends and regional warming over past ~ 150 years
Detrended correspondence analysis (DCA) determined that the cladoceran gradient length of 0.7 SD for the L. Opinicon record was relatively short (i.e., < 2.0 SD; Birks 1995) and therefore a linear regression technique would be appropriate. The major patterns of variation in the cladoceran assemblages from the 2019 L. Opinicon core were summarized with principal component analysis (PCA) and plotted against core depth/time, using the default options in C2 version 1.5 (Juggins 2007). To compare trends among sedimentary proxies and regional MAAT records, PCA axis 1 (PC1) sample scores for cladocerans (this study), PC1 sample scores for diatoms (Balasubramaniam et al. 2023), ratios of benthic and pelagic cladoceran taxa (B:P), and Z-scores of VRS-Chla (the value of each sediment interval standardized within the core to a mean of zero and a standard deviation of one) were plotted against sediment core depth/age. The relationship between Cladocera and diatom PC1 sample scores, as well as between PC1 scores from both proxies and MAAT anomalies, were assessed using Spearman rank correlation analysis. A PCA was also used to provide a graphical trajectory of change in the cladoceran composition over time by plotting sample scores of PC1 and PC2.
Results
A detailed summary of the 210Pb dating of this sediment core is provided in Balasubramaniam et al. (2023). Below we describe the changes in paleolimnological proxies with respect to the significant Cladocera zones based on a broken stick model applied to stratigraphic zones that were established using constrained incremental sum of squares (CONISS) cluster analysis. The two sub-zones (3a and 3b) were not identified as significant by the broken stick model.
Zone 1: 44–21 cm (bottom of core to canal construction to early 1800s)
From the bottom of the core to the early 1800s, the cladoceran assemblage was mainly dominated by littoral taxa, particularly Alona spp., which remained relatively stable at ~30% relative abundance for the majority of the zone but began decreasing slightly around 22 cm (early 1800s; Fig. 2). Chydorus spp. decreased from ~ 25% relative abundance at the bottom of this zone to ~ 10% at 24 cm. The remainder of littoral species remained relatively stable throughout the zone. Meanwhile, pelagic taxa, such as Bosmina spp., remained low from the bottom of the core (~ 10% relative abundance) to 22 cm (~ 25% relative abundance; Fig. 2). Daphnia spp. were either absent or only present in trace amounts in this zone.

Cladocera stratigraphic profile showing the percent relative abundances of the most common Cladocera taxa enumerated in the 2019 L. Opinicon core. The figure is scaled by depth with the corresponding CRS-estimated 210Pb dates (including errors) presented as a secondary axis. Dates in bold have been extrapolated and are viewed with caution: italicized dates were interpolated. Cladocera are ordered from left to right based on their axis one species scores from a canonical correspondence analysis constrained to depth. Horizontal black lines depict significant zones (dashed lines are sub-zones) determined by constrained incremental sum of squares (CONISS) and are represented as the total sum of squares (TSS)
Zone 2: 21–13 cm (early 1800s to early 1900s)
Zone 2 was characterized by decreased relative abundances of some littoral species, especially after canal construction ca. 1830 (Fig. 2). Alona spp. began decreasing from ~ 25% relative abundance at 22 cm (early 1800s) to ~ 15% at 14 cm (ca. 1900). Chydorus spp. remained relatively stable at ~ 15% relative abundance. The remainder of the littoral species had subtle declines in relative abundances, except for Alonella spp. that exhibited a subtle increasing trend. Bosmina spp. began increasing subtly during Zone 2, from 15% in Zone 1 to ~ 20% during Zone 2 (Fig. 2). Daphnia spp. remained at relatively low abundances with a slight jump to ~ 5% at 16 cm (ca. 1875).
Zone 3a: 13–5 cm (early 1900s to mid-1970s)
Zone 3a was characterized by a distinct increase in Daphnia spp. and further increases in Bosmina spp. that were concurrent with changes in the relative abundance of many littoral species (Fig. 2). For example, Daphnia spp., which were in low relative abundance in earlier intervals, increased at the beginning of this zone (12 cm; ca. 1921) to the end of this zone (6 cm; ca. 1970 ± 6.9) reaching peak relative abundances of ~ 15%. Chydorus spp. and Alona spp. remained relatively stable in this zone whereas Alonella spp., Ophryoxus gracilis Sars, Kurzia latissimi Kurz, Eurycerus spp. Baird and Camptocercus spp. Baird continued to decrease.
Zone 3b: 5 cm to top of the core (mid-1970s to 2019)
The shift to subzone 3b was characterized by the almost complete disappearance of several littoral taxa including Acroperus harpae Baird, Kurzia latissima, and Ophryoxus gracilis at 4 cm (ca. 1970 ± 6.9): these taxa did not recover in the remainder of the sediment record (Fig. 2). Alona spp. decreased to ~ 8% at 2 cm (ca. 2009 ± 1.2) but increased to ~ 25% in the uppermost core interval. Chydorus spp. increased to ~ 25% at the top of the core (ca. 2019 ± 0.7), whereas Eurycercus spp. and Camptocercus spp. decreased to below ~ 5% in the most recent sediment intervals of the core. Pelagic Bosmina spp. continued to increase in this zone, reaching a peak relative abundance of ~ 45% at 2 cm (ca. 2009 ± 1.2), whereas Daphnia spp. decreased to 5% relative abundance at the top of the core (2019 ± 0.7).
Comparison among paleo proxy trends and regional warming over past ~ 150 years
The main cladoceran trends in the L. Opinicon sediment record, summarized using both PC1 sample scores (Fig. 3A) and ratios of benthic to pelagic taxa (Fig. 3B), tracked similar temporal responses to diatoms (PC1 samples scores; Fig. 3C). Cladocera PC1 sample scores were significantly correlated to diatom PC1 sample scores (r = 0.88, p = 0.003). In addition, regional air temperature (MAAT) trends (Fig. 3E) were significantly correlated to the Cladocera (r = 0.82, p = 0.011) and diatom (r = 0.65, p = 0.02) PC1 sample scores. Cladocera PC1 (ʎ = 0.54) generally follows the trends in Bosmina spp., Daphnia spp., and Chydorus spp. (Fig. 2) with relatively high species scores on PCA axis 1 (2.48, 0.98, 0.79, respectively). Increases in Bosmina spp. and Daphnia spp. between ca. 1940 to the ~1980s correspond to a clear increase in pelagic taxa (Fig. 3b), the lowest values in VRS-Chla (Fig. 3D), and a shift towards warmer and more variable temperature anomalies (Fig. 3E). PC1 diatom sample scores remained relatively stable during this period (Fig. 3C), when assemblages were dominated by a variety of benthic fragilarioid taxa (Balasubramaniam et al. 2023).

Comparison of L. Opinicon proxy data and regional temperature trends. A Cladocera assemblage changes summarized by principal component analysis axis one sample scores (PC1); B Ratio of benthic to pelagic Cladocera sums; C Diatom assemblage changes summarized by PC1; D VRS-Chla concentrations presented as Z-scores; and E Mean annual air temperature (MAAT) data from the Kingston, ON weather station where the negative anomalies (black bars) are values below the baseline mean (1951–1980) and positive anomalies (red bars) are values above the mean. The horizontal dashed lines are the significant cladoceran assemblage zones determined by constrained incremental sum of squares (CONISS) and the broken-stick model
Following the 1980s, MAAT anomalies shift to consistently positive values (i.e., temperatures were above the 30-year baseline; Fig. 3E). Cladoceran PC1 scores are highest during this period, reflecting further declines in littoral taxa and increases in pelagic species (Fig. 3A). Diatom PC1 scores show a pronounced change (Fig. 3C) that reflects the onset of planktonic diatom accumulation for the first time in the core (Balasubramaniam et al. 2023) and an increase in VRS-Chla, stabilizing around 2000 (Fig. 3D).
Discussion
Over the past ~ 200 years, L. Opinicon and its catchment have experienced a multitude of environmental stressors, including canal construction and land-use change. Whilst compositional changes in cladoceran assemblages coincided with early deforestation and flooding, the most pronounced ecological shift in cladoceran assemblages occurred from the ~ 1970s to present-day, with a decrease in previously plentiful littoral taxa and increases in Bosmina spp. These changes are discussed in detail below and compared to the Balasubramaniam et al. (2023) diatom record to explore the possible impacts of environmental drivers (e.g., habitat changes, changes in food quality/quantity, and response to recent warming) on biological assemblages in L. Opinicon.
Past lake conditions: pre-Rideau Canal (1827–1832) to ca.1995
Prior to the construction of the canal, cladoceran assemblages were dominated by littoral species with pelagic species accounting for < 20%. However, canal construction and extensive flooding led to an increase in depth of shallow L. Opinicon and an extension of its length from 4.3 to 9.5 km. Flooding not only expanded the already extensive littoral zone (80% of the lake < 4 m deep; Little and Smol 2000) but also, with deepening, provided more habitat for pelagic species such as Bosmina spp., whose relative abundance doubled following canal construction. These early large-scale catchment disturbances (complete deforestation, flooding with canal construction) were also evident in the diatom record when tychoplanktonic Aulacoseira ambigua, that dominated the pre-canal sediments, abruptly decreased to trace abundances with a concurrent increase in benthic diatoms (Balasubramaniam et al. 2023). The post-flooding extension of the littoral zone and expansion of macrophytes led to increased benthic diatom habitat availability in this shallow system. Concurrent with this diatom shift, the cladoceran record registered an increase in littoral Chydorus species. Certain Cladocera taxa, such as Chydorus, are excellent at using the complex macrophyte habitat for survival and can adapt well to the variety of food sources available in macrophyte-rich littoral zones (Tremel et al. 2000; Basińska et al. 2014). For example, macrophytes can serve as a daytime refuge for Cladocera taxa (Timms and Moss 1984; Lauridsen et al. 1996) against predation when lakes are in clear water states. The advantages that macrophyte coverage offer cladocerans are even more important in shallow systems such as L. Opinicon, as the capacity to evade predation through vertical migration is greatly diminished (Jeppesen et al. 1997).
Diatom-inferred TP concentrations did not change substantially following early catchment activities at L. Opinicon (mean TP = 17–23 μg/L; Karst and Smol 2000) but were relatively high during the 1970s (based on monitoring data). During this time of higher TP, Daphnia spp. reached its peak in relative abundances, consistent with studies that have found a positive relationship between TP and the growth rate of Daphnia spp. (Urabe et al. 1997; Labaj et al. 2021). However, this relationship is likely indirect as TP can impact the quantity and quality of algal food for Daphnia spp. (Scheuerell et al. 2002). Bosmina spp., on the other hand, have been associated with more oligotrophic conditions (Labaj et al. 2021) and are specialized to effectively feed on smaller food sources (DeMott and Kerfoot 1982). The continued increase in Bosmina spp. in our sediment record between the 1940s and 1970s, despite relatively high TP concentrations, is consistent with the lowest concentrations in VRS-Chla during this time. During this period, the lake bottom was 80% covered in macrophytes (Crowder et al. 1977). In contrast to deeper lakes within the Rideau Canal system, such as Upper Rideau Lake, shallow L. Opinicon has likely resisted some symptoms of cultural eutrophication (Karst and Smol 2000; Balasubramaniam 2022). Because of the large macrophyte abundance, L. Opinicon is believed to be in a clear-water equilibrium, with the dense growth of macrophytes acting to supress algal activity (Scheffer et al. 1993).
With the exception of Chydorus spp., the most distinct compositional change in the cladoceran record identified by CONISS occurs at a later point (ca. 1920) than the largest change registered in the diatom record (ca. 1850). This difference in the two indicator species may be for a variety of reasons. Previous multi-proxy paleolimnological studies have shown that Cladocera tend to have muted or delayed responses to environmental stressors, compared to diatoms (Rühland et al. 2014; Hargan et al. 2016). In general, Cladocera tend to have lower taxa richness than diatoms and assemblages are comprised of greater numbers of generalist species (Kurek et al. 2011; Rühland et al. 2014; Simmatis et al. 2021). Therefore, Cladocera taxa may not respond as quickly or as clearly to limnological changes associated with climate warming. Cladocera are also impacted by both top-down and bottom-up factors, whereas diatoms are strongly impacted by bottom-up factors and thus diatoms may respond faster to climate warming (Sweetman et al. 2008; Rühland et al. 2014; Hargan et al. 2016; Reavie 2020). The gradual increase in pelagic taxa (including both Bosmina spp. and Daphnia spp.) and a compensatory decline in most littoral taxa may be a response to the onset of warmer conditions in the twentieth century (Fig. 3E) with longer open water periods that would have been favourable for pelagic cladoceran taxa.
Recent lake conditions: ~ 1995 to 2019
Relative to the response to canal construction during the nineteenth century in deeper lakes in the Rideau Canal system, the sedimentary cladoceran and diatom records from L. Opinicon showed a muted response to the large-scale catchment disturbances (complete deforestation, flooding with canal construction compared to deeper lakes in the system; Graves et al. unpublished data). However, both proxies underwent distinct compositional changes during the past ~ 25 years. In particular, both proxies registered notable increases in planktonic taxa that likely reflect changes associated with accelerated regional climate warming including longer and warmer ice-free periods, decreased wind speeds, and a shift from a well-mixed water column to sustained periods of thermal stratification (Balasubramaniam et al. 2023). Indeed, regional temperature trends clearly show a shift to warmer conditions with consistently positive MAAT anomalies during this recent period (Fig. 3E) that were significantly correlated to cladoceran (r = 0.82, p = 0.011) and diatom (r = 0.65, p = 0.02) PC1 trends. The continued decline during the past ~ 25 years of several littoral Cladocera taxa, including Alonella spp., Eurycercus spp., Camptocercus spp., Acroperus harpae, Kurzia latissima, and Ophryoxus gracilis, and an increase in pelagic Bosmina spp., has often been linked to warming-induced limnological changes. For example, recent increases in Bosmina spp. have been widely reported in many temperate and high latitude lake ecosystems as a response to earlier ice-off periods in the spring, higher production, and longer open-water seasons in recent decades (Conboy and English 2010; Jeppesen et al. 2014; Thienpont et al. 2015; Armstrong and Kurek 2019).
In addition to accelerated regional warming, recent changes in cladoceran assemblages may be associated with declines in food sources. For example, VRS-Chla increased from the 1970s to the early 2000s (although still lower than the pre-canal period) but has remained relatively stable thereafter (in the past ~ 20 years). Based on measured limnological data, L. Opinicon also experienced decreasing lakewater TP concentrations from the 1970s (mean TP [1971–1988] = 22.86 μg/L) to the present (mean TP [2002–2019] = 14.08 μg/L) and the lake is now considered to be oligo-mesotrophic (Balasubramaniam et al. 2023). The lower TP concentrations and relatively stable algal biomass may partly explain the increasing trend in Bosmina spp. and lower abundances of Daphnia spp., as Bosmina spp. tend to outcompete Daphnia spp. when food resources shift to lower concentrations and smaller sizes (DeMott and Kerfoot 1982). Also consistent with declining TP concentrations, Bosmina spp. are more disturbance-tolerant (Jiang et al. 2014) and require less phosphorus than the larger bodied Daphnia spp. (Schulz and Sterner 1999; Reissig et al. 2015).
An additional recent stressor that has been impacting lakes along the Rideau Canal system is the introduction of invasive zebra mussels (Dreissena polymorpha Pallas). Zebra mussels were first noted in L. Opinicon in the mid-1990s (Watson 2000) and, as detailed in Balasubramaniam et al. (2023), populations increased substantially until ~ 2013, but have since declined markedly. Similar to the conclusions reached by Balasubramaniam et al. (2023) for the diatom assemblage changes, it is unlikely that the stratigraphic changes in cladocerans were linked to zebra mussel invasion. First, the cladoceran changes predate the arrival of zebra mussels. Second, planktonic taxa such as Bosmina spp. continue to increase towards the surface, even though mussel populations have declined markedly over the last decade or so. Overall, changes to cladoceran communities appear to indicate that warming since the 1970s, followed by recent acceleration over the past ~ 25 years, has had a notable impact on L. Opinicon ecology.
Conclusions
During the past ~ 200 years, various catchment and other lake disturbances have occurred within L. Opinicon. Whilst compositional changes in cladoceran assemblages have occurred throughout the lake’s post-industrial history, the most pronounced ecological shift occurred after the ~ 1970s. These changes are similar in nature to those previously recorded in diatom assemblages. Based on cladoceran assemblage compositional changes, and comparisons to regional meteorological data, as well as the diatom and VRS-Chla records from this same sediment core, it seems likely that these most recent shifts in primary consumers are linked to climate warming as well as changing food resources and declining nutrient concentrations. Collectively, these data show that shallow, macrophyte-dominated lakes, such as L. Opinicon, despite experiencing relative stability even when subjected to large-scale nineteenth century catchment disturbances, are now changing rapidly in response to recent climate warming and associated limnological changes. Our ongoing research includes a multi-proxy study of other lakes within the Rideau Canal system (especially deeper lakes than L. Opinicon) to gain further insights into how various catchment disturbances and recent climate warming have impacted these waterbodies and whether lakes with different morphologies are impacted similarly.
References
Agbeti MD, Smol JP (1995a) Chrysophyte population and encystment patterns in two Canadian lakes. J Phycol 31:70–78
Agbeti MD, Smol JP (1995b) Winter limnology: a comparison of physical, chemical and biological characteristics in two temperate lakes during ice cover. Hydrobiologia 304:221–234
Agbeti MD, Kingston JC, Smol JP, Watters C (1997) Comparison of phytoplankton succession in two lakes of different mixing regimes. Arch Hydrobiol 140:37–69
Appleby PG (2001) Chronostratigraphic techniques in recent sediments. In: Last WM, Smol JP (eds) Tracking Environmental Change Using Lake Sediments. Volume 1: Basin Analysis, Coring, and Chronological Techniques. Kluwer Academic Publishers, Dordrecht, pp 171–203
Armstrong Z, Kurek J (2019) Sensitivity and response of low-nutrient lakes to post twentieth century environmental change in New Brunswick, Canada. J Paleolimnol 61:85–98
Balasubramaniam K, Rühland KM, Smol JP (2023) A diatom-based paleolimnological re-assessment of previously polymictic Lake Opinicon, Ontario (Canada): crossing an ecological threshold in response to warming over the past 25 years. J Paleolimnol 69:37–55
Balasubramaniam K (2022) Have lakes within the Rideau Canal system responded to impacts of recent anthropogenic stressors over the past ~25 years: A diatom-based paleolimnological re-assessment. MSc thesis. Queen’s University, Kingston
Basińska AM, Antczak M, Świdnicki K, Jassey VEJ, Kuczyńska-Kippen N (2014) Habitat type as strongest predictor of the body size distribution of Chydorus sphaericus (O. F. Müller) in small water bodies. Int Rev Hydrobiol 99:382–392
Bennett KD (1996) Determination of the number of zones in a biostratigraphical sequence. New Phytol 132:155–170
Bennion H, Davidson TA, Sayer CD, Simpson GL, Rose NL, Sadler JP (2015) Harnessing the potential of the multi-indicator palaeoecological approach: an assessment of the nature and causes of ecological change in a eutrophic shallow lake. Freshw Biol 60:1423–1442
Bergman JN, Beaudoin C, Mistry I, Turcotte A, Vis C, Minelga V, Neigel K, Lin HY, Bennett JR, Young N, Rennie C, Trottier LL, Abrams AEI, Beaupre P, Glassman D, Blouin-Demers G, Garant D, Donaldson L, Vermaire JC, Smol JP, Cooke SJ (2022) Historical, contemporary, and future perspectives on a coupled social-ecological system in a changing world: Canada’s historic Rideau Canal. Environ Rev 30:72–87
Birks HJB (1995) Quantitative paleoenvironmental reconstructions. In: Maddy D, Brew JS (eds) Statistical modeling of Quaternary science data: issue 5 of Quaternary Research Association Technical Guide. Quaternary Research Association, Chicago, pp 161–255
Burks RL, Jeppesen E, Lodge DM (2001) Littoral zone structures as Daphnia refugia against fish predators. Limnol Oceanogr 46:230–237
Canadian Heritage Parks Canada (CHPC) (1996) Working Towards a Shared Future, Rideau Canal Management Plan
Christie CE, Smol JP (1996) Limnological effects of 19th century canal construction and other disturbances on the trophic state history of Upper Rideau Lake, Ontario. Lake Reserv Manage 12:448–454
Conboy MA, English P (2010) Freeze up and thaw dates for Lake Opinicon, 1985–2010. Accessed from https://opinicon.wordpress.com/2010/12/06/freeze-up-and-thaw-dates-for-lake-opinicon-1985-2010/
Crowder AA, Bristow JM, King MR, Vanderkloet S (1977) Distribution, seasonality, and biomass of aquatic macrophytes in Lake Opinicon (Eastern Ontario). Nat Can 104:441–456
DeMott WR, Kerfoot CW (1982) Competition among cladocerans: nature of the interaction between Bosmina and Daphnia. Ecology 63:1949–1966
DeSellas AM, Paterson AM, Sweetman JN, Smol JP (2008) Cladocera assemblages from the surface sediments of south-central Ontario (Canada) lakes and their relationships to measured environmental variables. Hydrobiologia 600:105–119
Forrest F, Reavie ED, Smol JP (2002) Comparing limnological changes associated with 19th century canal construction and other catchment disturbances in four lakes within the Rideau Canal system, Ontario, Canada. J Limnol 61:183–197
Frey DG (1986) Cladocera analysis. In: Berglund BE (ed) Handbook of Holocene palaeoecology and palaeohydrology. Wiley, New York, pp 667–692
Glew JR (1988) A portable extruding device for close interval sectioning of unconsolidated core samples. J Paleolimnol 1:235–239
Glew JR (1989) A new trigger mechanism for sediment samplers. J Paleolimnol 2:241–243
Grimm EC (1991) TILIA v2. 0. B. 4 and TGView v2. 0 (computer software) Illinois State Museum, Research and Collections Center Springfield, United States
Hansen JR, Ruedy R, Sato M, Lo K (2010) Global surface temperature change. Rev Geophys 48:RG4004
Hargan KE, Nelligan C, Jeziorski A, Rühland KM, Paterson AM, Keller W, Smol JP (2016) Tracking the long-term responses of diatoms and cladocerans to climate warming and human influences across lakes of the Ring of Fire in the far North of Ontario, Canada. J Paleolimnol 56:153–172
Janssen CR, Birks HJB (1994) Recurrent groups of pollen types in time. Rev Palaeobot Palyno 82:165–173
Jeppesen E, Jensen JP, Søndergaard M, Lauridsen T, Pedersen LJ, Jensen L (1997) Top-down control in freshwater lakes: the role of nutrient state, submerged macrophytes and water depth. In: Kufel L, Prejs A, Rybak JI (eds) Shallow Lakes ’95, vol 119. Developments in Hydrobiology. Springer, Dordrecht, pp 151–164
Jeppesen E, Noges P, Davidson TA, Haberman J, Noges T, Blank K, Lauridsen TL, Søndergaard M, Sayer C, Laugaste R, Johansson LS, Bjerring R, Amsinck SL (2011) Zooplankton as indicators in lakes: a scientific-based plea for including zooplankton in the ecological quality assessment of lakes according to the European Water Framework Directive (WFD). Hydrobiologia 676:279–297
Jeppesen E, Meerhoff M, Davidson TA, Trolle D, Søndergaard M, Lauridsen TL, Beklioglu M, Brucet S, Volta P, Gonzalez-Bergonzoni I, Nielsen A (2014) Climate change impacts on lakes: an integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J Limnol 73:84–107
Jiang X, Yang W, Xiang X, Niu Y, Chen L, Zhang J (2014) Cyanobacteria alter competitive outcomes between Daphnia and Bosmina in dependence on environmental conditions. Fund Appl Limnol 184:11–22
Juggins S (2007) C2 Version 1.5 User Guide. Software for ecological and palaeoecological data analysis and visualisation. Newcastle University
Kahle D, Wickham H (2013) ggmap: Spatial Visualization with ggplot2. R J 5:144–161
Karst TL, Smol JP (2000) Paleolimnological evidence of limnetic nutrient concentration equilibrium in a shallow, macrophyte-dominated lake. Aquat Sci 62:20–38
Korhola A, Rautio M (2001) Cladocera and other branchiopod crustaceans. In: Smol JP, Birks HJ, Last WM (eds) Tracking Environmental Change Using Lake Sediments, vol 4. Zoological Indicators. Springer, Dordrecht, pp 5–41
Korosi JB, Smol JP (2011) Distribution of cladoceran assemblages across environmental gradients in Nova Scotia (Canada) lakes. Hydrobiologia 633:83–99
Korosi JB, Smol JP (2012a) An illustrated guide to the identification of cladoceran subfossils from lake sediments in northeastern North America: part 1—the Daphniidae, Leptodoridae, Bosminidae, Polyphemidae, Holopedidae, Sididae, and Macrothricidae. J Paleolimnol 48:571–586
Korosi JB, Smol JP (2012b) An illustrated guide to the identification of cladoceran subfossils from lake sediments in northeastern North America: part 2—the Chydoridae. J Paleolimnol 48:587–622
Korosi JB, Paterson AM, DeSellas AM, Smol JP (2008) Linking mean body size of pelagic Cladocera to environmental variables in Precambrian Shield lakes: a paleolimnological approach. J Limnol 67:22–34
Kurek J, Weeber RC, Smol JP (2011) Environment trumps predation and spatial factors in structuring cladoceran communities from Boreal Shield lakes. Fish Aquat Sci 68:1408–1419
Labaj AL, Michelutti N, Smol JP (2018) Cladocera in shallow lakes from the Ecuadorian Andes show little response to recent climate change. Hydrobiologia 822:203–216
Labaj AL, Jeziorski A, Kurek J, Bennett JR, Cumming BF, DeSellas AM, Korosi JB, Paterson AM, Sweetman JN, Thienpont JR, Smol JP (2021) Environmental drivers of cladoceran assemblages at a continental scale: a synthesis of Alaskan and Canadian datasets. Freshw Biol 66:949–967
Lauridsen T, Pedersen LJ, Jeppesen E, Sønergaard M (1996) The importance of macrophyte bed size for cladoceran composition and horizontal migration in a shallow lake. J Plankton Res 18:2283–2294
Little JL, Smol JP (2000) Changes in fossil midge (Chironomidae) assemblages in response to cultural activities in a shallow, polymictic lake. J Paleolimnol 23:201–212
Michelutti N, Blais JM, Cumming BF, Paterson AM, Rühland KM, Wolfe AP, Smol JP (2010) Do spectrally inferred determinations of chlorophyll a reflect trends in lake trophic status? J Paleolimnol 43:205–217
Nelligan C, Jeziorski A, Rühland KM, Paterson AM, Smol JP (2016) Managing lake trout lakes in a warming world: a paleolimnological assessment of nutrients and lake production at three Ontario sites. Lake Reserv Manage 32:315–328
Nevalainen L, Luoto TP (2017) Relationship between cladoceran (Crustacea) functional diversity and lake trophic gradients. Funct Ecol 31:488–498
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin RP, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2015) Vegan: community ecology package. R package version 2.2–1. http://CRAN.Rproject.org/package=vegan
Reavie ED (2020) Paleolimnology supports aquatic management by providing early warnings of stressor impacts. Lake Reserv Manag 36:210–217
Reissig M, Quiemaliños C, Modenutti B, Balseiro E (2015) Prey C: P ratio and phosphorus recycling by a planktivorous fish: advantages of fish selection towards pelagic cladocerans. Ecol Freshw Fish 24:214–224
Rühland KM, Hargan KE, Jeziorski A, Paterson AM, Keller W, Smol JP (2014) A multi-trophic exploratory survey of recent environmental changes using lake sediments in the Hudson Bay Lowlands, Ontario, Canada. Arct Antarct Alp Res 46:139–158
Scheffer M, Hosper SH, Meijer ML, Moss B, Jeppesen E (1993) Alternative equilibria in shallow lakes. Trends Ecol Evol 8:275–279
Schelske CL, Peplow A, Brenner M, Spencer CN (1994) Low-background gamma counting: applications for 210Pb dating of sediments. J Paleolimnol 10:115–128
Scheuerell MD, Schindler DE, Litt AH, Edmondson WT (2002) Environmental and algal forcing of Daphnia production dynamics. Limnol Oceanogr 47:1477–1485
Schulz KL, Sterner RW (1999) Phytoplankton phosphorus limitation and food quality for Bosmina. Limnol Oceanogr 44:1549–1556
Simmatis B, Paterson AM, Edwards BA, Smol JP (2021) Linking changes in sedimentary Cladocera assemblages to limnological variables in 67 Sudbury (Ontario, Canada) lakes reflecting various degrees of metal smelter impacts. Fish Aquat Sci 78:1455–1469
Stainsby EA, Winter JG, Jarjanazi H, Paterson AM, Evans DO, Young JD (2011) Changes in the thermal stability of Lake Simcoe from 1980 to 2008. J Great Lakes Res 37:55–62
Sweetman JN, LaFace E, Rühland KM, Smol JP (2008) Evaluating the response of Cladocera to recent environmental changes in lakes from the Central Canadian Arctic Treeline Region. Arct Antarct Alp Res 40:584–591
Thienpont JR, Korosi JB, Cheng ES, Deasley K, Pisaric MF, Smol JP (2015) Recent climate warming favours more specialized cladoceran taxa in western Canadian Arctic lakes. J Biogeogr 42:1553–1565
Timms RM, Moss B (1984) Prevention of growth of potentially dense phytoplankton populations by zooplankton grazing, in the presence of zooplanktivorous fish, in a shallow wetland ecosystem. Limnol Oceanogr 29:451–673
Tremel B, Frey SE, Yan ND, Somers KM, Pawson TW (2000) Habitat specificity of littoral Chydoridae (Crustacea, Branchiopoda, Anomopoda) in Plastic Lake, Ontario, Canada. Hydrobiologia 432:195–205
Urabe J, Clasen J, Sterner RW (1997) Phosphorus limitation of Daphnia growth: is it real? Limnol Oceanogr 42:1436–1443
Vincent LA, Hartwell MM, Wang XL (2020) A third generation of homogenized temperature for trend analysis and monitoring changes in Canada’s climate. Atmos Ocean 58:173–191
Wang D, Kong L, Wang J, Huang L, Liu Y, Chen G (2020) Spatiotemporal patterns of cladoceran community responses to water level variation in Haixi Lake, southwest China. Inland Waters 10:267–282
Warren S (1997) Hub of the Rideau: a history of South Crosby township. The Local Architectural Conservation Advisory Committee, Township of South Crosby, Leeds County, Ontario
Watson K (2000) Rideau Canal Waterway–zebra mussel sightings. http://www.rideau-info.com/local/zebra.html. Accessed 20 Aug 2023
Wolfe AP, Vinebrooke RD, Michelutti N, Rivard B, Das B (2006) Experimental calibration of lake-sediment spectral reflectance to chlorophyll a concentrations: methodology and paleolimnological validation. J Paleolimnol 36:91–100
Acknowledgements
This research was funded by the Natural Sciences and Engineering Research Council of Canada by Discovery Grants to JPS and AMP and the Rideau Lakes Environmental Foundation (RLEF). We thank the staff at the Queen’s University Biological Station (QUBS) for logistical support, and the Ontario Ministry of the Environment, Conservation and Parks, Dr. David Phillipp for sharing his knowledge of historical zebra mussel abundances in Lake Opinicon, and Brigitte Simmatis for coordinating sediment core collection at QUBS. We also thank Maarten van Hardenbroek and two anonymous reviewers for constructive comments that helped strengthen the manuscript.
Funding
This study was funded by the Natural Sciences and Engineering Research Council of Canada (Discovery) awards to JPS and AMP.
Author information
Authors and Affiliations
Contributions
ELG, KB, KMR, AMP, and JPS conceived and designed the study. KB assisted with collecting and dating the sediment core. ELG identified and counted the cladocerans and prepared the figures. AMP and JPS provided the funding. All authors co-wrote and reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
We declare no conflict of interests with any commercial or other interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
10933_2024_319_MOESM1_ESM.pdf
Supplementary Figure 1: Principal component analysis (PCA) showing axis 1 (λ = 0.54) and 2 (λ = 0.15) sample scores from L. Opinicon. Time markers show significant dates including the beginning of the record (2019), period of increased regional warming (1970), and when the Rideau Canal was constructed (1830) (PDF 12 KB)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Graves, E.L., Balasubramaniam, K., Rühland, K.M. et al. Changes in cladoceran assemblage composition linked to early nineteenth century canal construction, land-use changes, and recent climate change in a macrophyte-dominated Ontario lake. J Paleolimnol 72, 49–61 (2024). https://doi.org/10.1007/s10933-024-00319-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-024-00319-x