Ecological and biogeographical studies of Neotropical non-marine ostracods are rare, although such information is needed to develop reliable paleoecological and paleoclimatic reconstructions for the region. An extensive, yet little explored South American area of paleoclimatic interest, is the arid-semiarid ecotone (Arid Diagonal) that separates arid Patagonia from subtropical/tropical northern South America, and lies at the intersection of the Pacific and Atlantic atmospheric circulation systems. This study focused on the Laguna Llancanelo basin, Argentina, a Ramsar site located within the Arid Diagonal, and was designed to build a modern dataset using ostracods (diversity, spatial distribution, seasonality, habitat preferences) and water chemistry. Cluster and multivariate analysis of the data indicated that salinity is the most significant variable segregating two ostracod groups. Limnocythere aff. staplini is the only species that develops abundant populations in the saline ephemeral Laguna Llancanelo during almost all seasons, and is accompanied by scarce Cypridopsis vidua in summer. The latter species is abundant in freshwater lotic sites, where Ilyocypris ramirezi, Herpetocypris helenae, and Cyprididae indet. are also found in large numbers. Darwinula stevensoni, Penthesilenula incae, Heterocypris incongruens, Chlamydotheca arcuata, Chlamydotheca sp., Herpetocypris helenae, and Potamocypris smaragdina prefer freshwater lentic conditions (springs), with C. arcuata and Chlamydotheca sp. found only in the Carapacho warm spring, which has a year-round constant temperature of ~20 °C. Seasonal sampling was necessary because some taxa display a highly seasonal distribution. Species that were recorded have either subtropical or Patagonian affinities, although a few taxa are endemic or common to both regions. These data can serve as modern analogues for reconstructing the late Quaternary history of the area, and to investigate the extent and position of the arid/semiarid ecotone (Arid Diagonal) during past glacial/interglacial cycles.
Ecological and biogeographical studies of non-marine ostracods from the Neotropical bioregion are rare, despite the wide application of ostracods in paleoclimate and paleoecological reconstructions. Databases such as the European (OMEGA, Horne et al. 2011) and North America databases (NACODe: North America Combined Ostracode Database; originally named NANODe (North American Non-marine Ostracode Database) (Forester et al. 2005; Curry et al. 2012) are compiling data on the presence/absence of ostracod species and associated hydrological/geochemical and climate conditions, to develop calibration datasets for paleo-reconstructions. The effort to build comprehensive databases will not be complete until data from the Southern Hemisphere are incorporated, particularly from South America, where the number of recorded ostracod species is low compared to other continents, including Australia and Africa (Martens et al. 2008).
Ostracod assemblages are useful to determine past hydrological conditions [salinity, temperature, water energy (De Deckker and Forester 1988; Curry 1999)] and water depth (Mourguiart et al. 1998), and when coupled with trace-element and oxygen-isotope analyses, enable estimation of past salinity, temperature and pH conditions (Chivas et al. 1983, 1985, 1986a, b; Gouramanis et al. 2010). In Argentina, Schwalb et al. (2002) studied ostracod taxa and isotopes from two Patagonian lakes, providing a database that can be applied to paleo-reconstructions. Other recent papers, involving a variety of approaches, have been published on studies in Patagonia (Cusminsky et al. 2005, 2011; Ballent and Díaz 2011; Ramón Mercau et al. 2012), the Pampas inland and coastal wetlands, including Isla Martín García, eastern Argentina (Cusminsky et al. 2006; Liberto et al. 2012; Ramón Mercau et al. 2014), and the Altiplano of northern Argentina (Laprida et al. 2006; Díaz and Lopretto 2011; Palacios-Fest et al. 2016). As part of this project in central-western Argentina, D’Ambrosio et al. (2015, 2016a) published a description of the soft parts of Ilyocypris ramirezi Cusminsky and Whatley (1996), a taxon originally described from Patagonian Quaternary sediments using only hard parts, and a revised description of Herpetocypris helenae Müller, 1908, which included soft and hard parts and was reported for the first time in the Neotropical region.
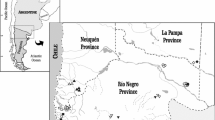
This study focuses on ostracods and water chemistry from a system of wetlands in west-central Argentina, namely the Laguna Llancanelo basin (~35° S Lat), located about 70 km east of the Andes range, where the mountains produce a topographic barrier that restricts continued easterly flow of winds and humidity. This situation changes south of ~40° S latitude, where the altitude of the Andes diminishes and the westerlies from the Pacific Ocean are the dominant wind through Patagonia, reaching the Pampean area. The wetlands are within the Arid Diagonal (Bruniard 1982), an extensive semiarid-arid area that receives <250 mm of annual precipitation, and that traverses South America approximately following the Río Colorado-Desaguadero basins in Argentina and continuing northwest to northern Perú (Fig. 1). Northeast of the Arid Diagonal the climate is subtropical, with annual precipitation >600 mm, mostly affected by the Atlantic Ocean summer monsoon. South of the Arid Diagonal the climate is arid, with annual precipitation <200 mm, typical of Patagonia, and influenced by the Pacific Ocean (Piovano et al. 2009).
Fig. 1
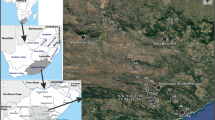
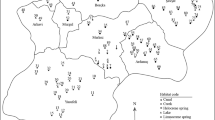
Location of sampling sites in Mendoza province, Argentina. 1–4 Laguna Llancanelo (Llancanelo Lake), 5 Bañado Los Menucos (Los Menucos spring), 6 Bañado Carilauquen (Carilauquen spring), 7 Bañado Carapacho (Carapacho warm spring), 8 Bañado La Porteña (La Porteña spring), 9 Río Malargüe (Malargüe River), 10 Arroyo El Chacay (El Chacay creek), 11 Arroyo El Alamo (El Alamo creek), 12 Arroyo El Mocho (El Mocho creek), 13–17 Delta del Río Malargüe (Malargüe River delta), 18 Arroyo El Malo (El Malo creek). In the inset map of South America, LL is the position of Laguna Llancanelo, SdB is Salina del Bebedero (San Luis province) and the shaded grey area (AD) is the Arid Diagonal
Modern ecological studies from the wetlands connected to Laguna Llancanelo dealt with malacofauna (Ciocco and Scheibler 2008; Scheibler and Ciocco 2011, 2013), plankton (Peralta and Fuentes 2005; D’Ambrosio et al. 2016b) and the ostracods I. ramirezi (D’Ambrosio et al. 2015) and H. helenae (D’Ambrosio et al. 2016a). Paleolimnological research undertaken at other sites within the Arid Diagonal (Salina del Bebedero and Río Atuel) used charophytes (García 1999) and molluscs (De Francesco 2010; De Francesco and Hassan 2009).
This is the first comprehensive study of ostracod diversity, water chemistry and seasonal data from the distinctive ecosystems of the Laguna Llancanelo area. This modern dataset provides ostracod/environment relationships across a broad environmental gradient, and is applicable to reconstruction of paleoecological and paleoclimate conditions in wetlands from the Arid Diagonal and elsewhere. The dataset also has the potential to shed light on changes in the behaviour of this ecotone during glacial/interglacial times, in response to weakening or strengthening of Pacific or Atlantic atmospheric circulation patterns.
A larger paleolimnological project in the Llancanelo area is currently being completed, including geochemistry of modern and fossil ostracod valves. Cores obtained in the lake basin are up to 10 m long and span the past 30 ka. Results from continuing “paleo” studies will be published separately and are beyond the scope of this paper.
Study site
The Llancanelo wetlands are located in southwest Mendoza Province, Argentina (35°–36.5°S, 68.5°–70°W) at 1330 m a.s.l., and close to the Cordillera de los Andes (Fig. 1). The drainage basin is endorheic, developed within the Huarpes depression in the northern section of the Payenia Volcanic Province (Ramos and Folguera 2011), an area characterized by the presence of more than 800 back-arc volcanic cones, some as young as 5 ka. Input of volcanic ash in the area, however, commonly derives from the Cordillera de los Andes, and layers of volcanic ash are found in the banks of creeks and around springs.
Sampling sites
Eighteen localities in the Laguna Llancanelo area were selected for sampling, but two sites were consistently dry (Arroyo El Alamo and Arroyo El Malo), so 16 sites (lotic, lentic, temporary, permanent, fresh to mesosaline ecosystems) were sampled seasonally for water and ostracod analyses (Fig. 1, Electronic Supplementary Material [ESM] Table S1).
Lentic, permanent, fresh and saline environments
Laguna Llancanelo (Fig. 1, sites 1–4) is a Provincial Reserve and a Ramsar site because of its important ecological role. It is the largest wetland within the arid area north of Patagonia, a refuge for endemic flora and fauna, and the nesting site for large colonies of flamingoes. The lake is ephemeral, shallow and saline, with a mean water depth of 0.3–1.7 m. Its surface area is ~28,000 ha, with large changes in lake area at annual, inter-annual and decadal scales (Isla et al. 2005). The lake is fed by permanent and ephemeral streams, groundwater and cold and warm springs—all fresh. The principal water source is from Andean snowmelt in spring and comes mainly from the Malargüe River.
According to Ostera and Dapeña (2003) the water is a chloride–sulfate–sodium type, with mean conductivity of 10.8 mS cm−1 (Peralta and Fuentes 2005). The high temperature and scarce rainfall produce salt flats in the shallowest areas of the lake.
There are several types of springs (bañados) around the lake (Fig. 1, sites 5–8). These are developed in depressions where groundwater emerges and forms large wetlands, commonly connected to the lake by small streams. Los Menucos rheocrene spring (site 5) is located in the northwest of the lake and has a mean water depth of 0.3 m. Carilauquen limnocrene spring (site 6) is located southwest of the lake; the water is predominantly sulfate calcic and has a mean conductivity of 0.959 mS cm−1 (Peralta and Fuentes 2005) and mean water depth of 0.2 m, though deeper areas reach 1.6 m (Ciocco and Scheibler 2008). The Carapacho helocrene spring (site 7) is the only warm spring in the area, located on the southwest side of the lake. The spring’s temperature is constant during the four seasons, around 19–20 °C; this is also the deepest wetland in the area, up to 4 m deep, although the mean water depth is 1 m and sampling was done close to the shore. La Porteña limnocrene spring (site 8) is in the southeast part of the lake, with a mean water depth of 0.5 m.
Lotic permanent environments
Río Malargüe (site 9) originates in the Cordillera de los Andes at 3000 m a.s.l. and receives input from numerous tributaries as it flows to Laguna Llancanelo, producing maximum discharge in spring and summer. Arroyo El Chacay (site 10) is a creek that originates in the Andes and flows eastward, joining the Río Malargüe close to the lake.
Lotic temporary environments
Arroyo El Alamo (site 11) is a tributary of Arroyo El Chacay and was dry during the sampling period. Accordingly, there are no data from this site. Arroyo El Mocho (site 12) originates in the Andes, and is a second-order tributary of the Río Malargüe, joining it in the lower section of its course. This creek displayed highly variable inter-season flow during sampling, holding much water during autumn and winter, but being almost dry in summer. The Río Malargüe delta (sites 13–17) forms where this river joins the lake in its northwest corner, after receiving water from Arroyos El Chacay and El Mocho. The delta is developed in a lowland area known as “Bajo Llancanelo” and holds water only temporarily because of marked seasonal fluctuations. Arroyo El Malo (site 18), which used to discharge into the northeast section of the lake, is now permanently dry, although it was an important watercourse in the past, its paleo-channel showing a connection to the Río Salado, which runs toward the north of the Llancanelo basin.
Materials and methods
Sampling procedures
Samples of ostracods and water were taken seasonally from 16 sites (September 2009–September 2010), and several physical and chemical variables of the waters were measured in situ at the same time. Ostracod samples were collected in shallow areas (about 20–40 cm deep) by washing aquatic plants (three times at each site to recover ostracod species that live close to macrophytes) and surrounding surface sediments (upper 3 cm) in a bucket, then rinsing the sediments in a 100-µm sieve. Samples were divided in two aliquots, one stored in 70% ethanol and the other air-dried.
In the field, a Horiba U-10 multimeter was used to measure pH, temperature, dissolved oxygen (DO) and conductivity. To calculate salinity, the conversion table from conductivity (envco) was used. Salinity classification used the ranges and terms proposed by Hammer et al. (1983).
Water samples were collected in bottles (500 ml) from the water surface, avoiding bubbling, after rinsing the bottles with water of the corresponding wetland. Once hermetically sealed, the bottles were stored in darkness and kept cold (4 °C). In the Geochemistry Laboratory, University of Wollongong, Australia, the water was filtered at 2.5 µm retention (Filtech 1803), and kept in bottles pre-washed with 10% HNO3 prior to chemical analysis using an Agilent 7500 ce Inductively Coupled Plasma Mass Spectrometer (ICP-MS). The interference-free isotopes 48Ca, 86Sr, 25Mg, 39K and 138Ba were analyzed.
Ostracod assemblage analyses
In the laboratory, each dried sample was washed again with tap water on a 100-μm sieve. Given the paleolimnological objectives of this work, we focused on detailed study of the hard parts, valves and carapaces, as these are the only parts of the ostracod that preserve in sediments and form the basis for the taxonomy of fossil taxa. Carapaces and valves were inspected using a stereo-microscope to separate those in perfect condition (in some cases with soft parts still attached), from those showing signs of dissolution or transport. Only ostracods showing pristine preservation were counted to be sure that specimens were either alive at the time of collection or had died very recently. The ostracods were sorted and valves stored dry on micro-paleontological slides. Identification at the genus and species level was performed using local literature and general references commonly used for ostracod systematics, with the abbreviations Cp (carapace), dv (dorsal view), ev (external view), H (height of valve), iv (internal view), lv (lateral view), L (length of valve), LV (left valve), RV (right valve), and vv (ventral view) used for descriptions of the valves. Valves were photographed and measured using scanning electron microscopy (SEM) (JEOL JSM 6360 LV at Universidad Nacional de La Plata, JEOL JSM 6610 LV at IANIGLA, Mendoza, Argentina, Philips XL30 at Royal Belgian Institute of Natural Sciences, Brussels, Belgium and JEOL JSM 6490 LA at University of Wollongong, Australia). The material is deposited in the carcinological collection of the Museo de La Plata, La Plata (Argentina) (MLP numbers). For statistical analyses, samples were standardized as individuals per gram of dry sediment.
Database: ostracods, environmental variables and water chemistry
Construction of a modern database, using data from the Llancanelo area, included sampling sites with geo-referenced positions, ecological variables (pH, salinity, conductivity, DO, temperature), and chemical analysis of the water (Ba, Ca, Mg, Na, Sr, K), collected seasonally (Table S1, ESM).
Statistical analyses
To explore relationships between ostracod associations and each type of environment, cluster analyses, outlined by constrained incremental sum of squares analysis and unconstrained-type analysis and square root transformation (CONISS), were carried out using Tilia and TiliaGraph programs (TGView 2.0.2; Grimm 2004). In addition, similarity analysis was used to segregate environmental variables that control the distribution of ostracods.
To investigate the relationship between species composition and environmental variables over the studied period, multivariate ordination techniques from the CANOCO program (version 4.5) were used (Pielou 1984; ter Braak 1986). The method of redundancy analysis (RDA) was selected because the lengths of the gradients of explanatory variables were short (ter Braak and Smilauer 2002).
Redundancy Analysis (RDA) was implemented to investigate which environmental and water chemistry factors affected ostracod distribution, with the analyses including only those environmental variables with a variance inflation smaller than 10, to mitigate the effect of multi-colinearity (ter Braak and Verdonschot 1995). The environmental data were standardized, then 11 ostracod species and 11 environmental variables from 16 sites collected in four seasons (72 samples) were statistically analysed. The statistical significance of variability for each parameter, and the general meaning of ordination were tested with a Monte Carlo test (499 permutations, p < 0.01), with three variables (salinity, Ba and Sr) being used for the posterior analysis.
Results
Water chemistry
The progressive, largely evaporative chemical evolution of the water bodies, from creeks, springs and delta inflow, through to a saline lake, is accompanied by a progressive increase in the relative amount of sodium and potassium, as demonstrated on a cation ternary diagram (Fig. 2) and by total salinity. On this diagram, Carapacho warm spring is similar to the cold springs. La Porteña spring, located on the east side of the lake, is somewhat different, with relatively higher magnesium content in all four sampled seasons. Earlier studies on limited datasets (Ostera and Dapeña 2003; Alvarez and Wetten 2004) demonstrated that anion evolution in water bodies of the Llancanelo area progresses from high relative levels of dissolved sulfate to high chloride, with a progressive increase in total salinity. The oxygen isotope values (vs VSMOW) of several of the water bodies span a considerable range, consistent with progressive evaporation. Sampling in February 2001, Ostera and Dapeña (2003) showed that the Andean headwaters (3000 m a.s.l.) of the Río Malargüe have a δ18O value of −16.5‰, with several springs, including Carilauquen, near Laguna Llancanelo having values of −14.9 to −14.4‰, and Laguna Llancanelo a value of +0.5‰.
Fig. 2
Cation diagram, in molar proportions, for solutes from water bodies within the Llancanelo catchment. All four seasonal samples from La Porteña spring plot with a higher relative Mg content, near the centre of the triangle
The solute chemistry of the various water bodies can be further examined using a series of bivariate diagrams. Sodium and potassium, previously grouped together in the triangular plot, are separated in Fig. 3a. The saline lake water demonstrates conservative consistency, with a molar Na:K ratio of 52, as do the less saline ponds, again except for La Porteña (the four more-saline green dots). Among the saline lakes, the four seasonal samples at Cerro Coral (Na = 550–660) are enriched in Na over K. This part of the lake is also fed by a saline spring at the foot of Cerro Coral, which must be adding additional Na. Similar to La Porteña, this is on the east side of the lake, suggesting that the north–south geological fault that is commonly postulated to be along the eastern shoreline, taps deeper water, rather than just near-surface groundwater and rain water.
Fig. 3
Bivariate molar plots (in milliequivalents per litre) of a Na versus K, b Sr versus Ca, c Mg versus Ca, and d Ba versus Ca, for waters within the Llancanelo catchment. Within each panel, the inset diagrams expand the scale for the areas near the origin
Plots of strontium versus calcium (Fig. 3b) and magnesium versus calcium (Fig. 3c) distinguish La Porteña, whereas the saline lake waters demonstrate their depleted barium content on a Ba versus Ca diagram (Fig. 3d), as Ba has been precipitated as BaSO4 and/or BaCO3, upon reaching saturation for these species. In a later section we examine whether these salinity and water chemistry differences exert control on the ostracod assemblages in each water body.
Ostracods
Eleven species of ostracods were identified: Limnocythere aff. staplini Gutentag and Benson, 1962, Chlamydotheca arcuata (Sars, 1901), Chlamydotheca sp., Heterocypris incongruens (Ramdohr, 1808), Cypridopsis vidua (Müller, 1776), Potamocypris smaragdina (Vávra, 1891), Herpetocypris helenae Müller, 1908, Ilyocypris ramirezi Cusminsky and Whatley, 1996, Darwinula stevensoni (Brady and Robertson, 1870), Penthesilenula incae (Delachaux, 1928) and Cyprididae indet. The description and SEM illustration of Limnocythere aff. staplini is presented in the main text because it is the characteristic taxon in the Laguna Llancanelo area, particularly in the lake, where a large population consisting of males, females and juveniles was found. The description, environmental preferences, and SEM illustrations of the remaining ostracod taxa are provided in the ESM. The relative abundance of each taxon at each locality in the different seasons is shown in ESM Fig. S2. ESM Table S1 is the complete modern database arising from this study, including site names, GPS positions, ostracod taxa, ecological variables and water chemistry data collected seasonally. ESM Table S2 compiles maximum and minimum values of the ecological data for each ostracod taxon.
Class Ostracoda Latreille, 1802
Order Podocopida G. O. Sars, 1866
Superfamily Cytheroidea Baird, 1850
Family Limnocytheridae Klie, 1938
Genus Limnocythere Brady, 1867
Limnocythere aff. staplini Gutentag and Benson, 1962
Limnocythere aff. staplini. a ♀ RV ev; b ♀ LV ev; c ♀ RV iv; d ♀ LV iv; e ♂ RV ev; f ♂ LV ev; g ♂ RV iv; h ♂ LV iv; i ♀ LV ev, light microscope illustrations showing the single undivided marginal pore-canals; j ♂ RV ev light microscope illustrations showing the single undivided marginal pore-canals; k ♀ Cp vv; l ♂ Cp dv; m ♀ RV iv, hinge detail; n ♀ RV iv, anterior tooth detail; o ♀ RV iv, posterior tooth detail; scale barsa–m = 100 μm; n–o = 50 μm
Description: Medium size. Carapace reniform to sub-rectangular in lateral view. Valves sub-equal with strong sexual dimorphism. Maximum height situated approximately at the anterior 1/4 of the valves. In lateral view the anterior margin is broadly rounded and the posterior margin is sharply rounded. In dorsal view are depressed anteriorly and more globose in the posterior section. LV overlapping the RV in posterior margin.
Female valve in lateral view is reniform to sub-quadrate. Dorsal margin broadly arched, in some cases with a small hump coinciding with the maximum height in the anterior dorsal margin, anterior and posterior cardinal angles weakly discernible. Ventral margin slightly concave, incurved near the middle, with minimum height of the valve about 2/3 from the anterior margin.
Male valve in lateral view is longer and sub-rectangular. Dorsal margin straight, forming a soft angle when it reaches the anterior and posterior area. Ventral margin straight in the middle section of the valve, subparallel to dorsal margin, rounded towards the anterior and posterior region.
External valve ornamentation is a soft reticulation, in some cases completely absent, more developed in the margin’s proximity. Four to five ribs develop around the anterior margin, starting approximately at the dorso-anterior hump and continuing in the ventral part, whereas in the posterior margin they are visible just around the margin. One median-dorsal transversal sulcus and one or two smooth, very poorly developed tubercles occur approximately at the anterior third of the maximum length. Normal pores scattered on the surface.
Internal view: Anterior and posterior margin rounded, ventral margin slightly curved inwards in the middle section, more marked approximately 2/3 from the anterior margin. Dorsal margin is straight in male and arched in female. Marginal zone well developed in the anterior, ventral and postero-ventral margins, but is two to three times larger in the anterior margin. Marginal pore canals undivided, straight.
The hinge is modified lophodont, antimerodont, consisting in the RV of a smooth groove with two terminal, one anterior and one posterior, medium-size smooth teeth. LV with a central smooth bar with an anterior and posterior smooth socket. Muscle scars as in genus. Juveniles are similar to adult females in contour, but a bit more rounded.
Measurements of adults Male: RV (n = 51), L = 520–665 μm (604 ± 33 μm); H = 242–300 μm (272 ± 15 μm); LV (n = 50), L = 555–681 μm (614 ± 33 μm); H = 240–310 μm (277 ± 15 μm). Female: RV (n = 92), L = 512–662 μm (582 ± 38 μm); H = 259–354 μm (302 ± 22 μm); LV (n = 95), L = 514–684 μm (586 ± 39 μm); H = 258–370 μm (301 ± 22 μm).
Previous record in Argentina The closely related Limnocythere staplini Gutentag and Benson, 1962 was originally described and found in North America (Gutentag and Benson 1962; Delorme 1971). Records of L. staplini in Argentina were later considered L. cusminskyae Ramón Mercau, Plastani and Laprida, 2014 (Ramón Mercau et al. 2014).
Distribution in the Laguna Llancanelo area and ecological data Most (91%) Limnocythere aff. staplini specimens were found in Laguna Llancanelo, a lentic mesosaline shallow waterbody subject to evaporation, with a mean annual salinity of 24.3 g L−1 and maximum salinity of 42.3 g L−1. The remaining 9% were collected from a small fresh to subsaline rheocrene spring (Los Menucos), the Malargüe River delta, which forms small fresh to subsaline pools when joining the lake, and El Chacay and El Mocho creeks, both ephemeral, consisting of fresh to subsaline lentic and lotic sections. The species Limnocythere aff. staplini was absent in the fresh to subsaline helocrene warm spring (Carapacho), in the subsaline limnocrene springs (Carilauquen and La Porteña) and the freshwater lotic system of the Malargüe River.
Remarks Although more than 4500 individuals were studied, only hard parts were used in the description because the soft tissues were degraded. There are nine extant and Quaternary species of Limnocythere recorded in Argentina, but our specimens, utilising the morphology of hard parts and ecological preferences, is closest to L. staplini, known only from North America, although in general shape it is also comparable to L. cusminskyae, which is recorded in Buenos Aires Province, Argentina (Ramón Mercau et al. 2014). It is generally accepted that hard parts of ostracods are genetically determined, in particular the hinge morphology, as valves are the only available remains for fossil ostracod taxonomy (Gutentag and Benson 1962). Because our specimens show close similarity to L. staplini, we considered them to be L. aff. staplini, though the hinges show consistent differences among these three taxa. A taxonomic decision will be made once soft parts are available. It is not clear at the moment if L. aff. staplini is a new species or merely an ecophenotype of L. staplini, which if so, poses interesting biogeographic questions.
It is important to describe the differences in valve morphology among the three taxa. The hinge of RV of Limnocythere staplini has two crenulated terminal teeth and a strong depression in the median section of the dorsal region of the valve, whereas both teeth in L. aff. staplini are smooth and the depression in the dorsal region of the valves is soft or absent. Both teeth of the RV in L. cusminskyae are small, with the posterior being crenulated and the anterior smooth, with marked reticulation, and the presence of denticles on the anterior and posterior margins of the valves, absent in L. aff. staplini. Also, the dorsal margin of the valve in L. staplini and L. aff. staplini continues with a symmetrical posterior curvature, in contrast to the dorsal margin in L. cusminskyae, which has a marked cardinal angle. With respect to ecological variables, L. aff. staplini is closer to L. staplini, preferring lentic saline water bodies.
Statistical analyses
Characterization of the water bodies from the Llancanelo Basin
Environmental variables pH, salinity, dissolved oxygen, temperature, Ca, Mg, Ba, Sr, K and Na concentrations (ESM Table S1), measured seasonally at all sites, were analysed using cluster analysis to visualise factors that are stronger in segregation of the sampled localities (Fig. 5). Two main groups were separated. Group A includes environments with salinities >10.3 g L−1 and group B includes sites with salinity <3.5 g L−1. Group A encompasses all the samples from Laguna Llancanelo. Group B is segregated in two subgroups by K concentration, with subgroup B1 characterized by K concentrations <5 mg L−1, grouping all samples from La Porteña spring, and most samples from Bañado Carapacho (warm spring) and Arroyo El Mocho; subgroup B2, with K content between 5 and 113 mg L−1 contains all samples from Los Menucos spring and most samples from the Río Malargüe delta, Arroyo El Chacay and Carilauquen spring.
Fig. 5
Cluster analysis of each sampling site in each season in relation to environmental variables. LL lake Laguna Llancanelo (Llancanelo Lake), LM spring Bañado Los Menucos (Los Menucos spring), Cari spring Bañado Carilauquen (Carilauquen spring), C w-spring Bañado Carapacho (Carapacho warm spring), LP spring Bañado La Porteña (La Porteña spring), M river Río Malargüe (Malargüe River), EC creek Arroyo El Chacay (El Chacay creek), EM creek Arroyo El Mocho (El Mocho creek), RM delta Delta del Río Malargüe (Malargüe River delta). The sampling site number is in brackets
The relative abundance of ostracod taxa in the different ecosystems and the cluster analysis of their ecological preferences (Fig. 6) show two main components: (1) group A, formed by Chlamydotheca sp. and C. arcuata found only in the Carapacho warm spring, with a constant yearly temperature, and (2) group B, characterized by species with higher representation in seasonally dependent environments. Group B comprises subgroup B1, characterized by species that live almost exclusively in the springs and subgroup B2, characterized by species that live almost exclusively in lotic and lentic environments.
Fig. 6
Cluster analysis of ostracod species and their distribution in percentage in each type of environment
Subgroup B1 is further segregated into B11 with P. incae and P. smaragdina, which are almost exclusively represented in limnocrene and rheocrene spring environments, and B12, with D. stevensoni, which has 68% of its individuals living in the freshwater Bañado Los Menucos spring, characterized by running water and 23% living in the Bañado Carapacho warm spring. Subgroup B2 is further segregated into B21, characterized by species that live almost exclusively in lotic sites, but also present in springs, and in the case of C. vidua, also in the lake, and subgroup B22, represented by L. aff. staplini. Subgroup B21 is further segregated into C and D. Subgroup C segregates further into C1 with H. helenae and H. incongruens, both very plastic species that are present in all freshwater environments and C2, with I. ramirezi, with higher abundance in lotic environments. Subgroup D is characterized by Cyprididae indet., which is present only in lotic environments and C. vidua, with higher affinity to lotic environments, but also present in the saline lake.
In the Redundancy Analysis (RDA) (Fig. 7), the first two canonical axes explain 99.8% of the cumulative variance of species–environment relationships. The first canonical axis and the sum of all canonical axes explain a significant portion of the variance in the ostracod abundance data (p = 0.04; p = 0.04, respectively). The environmental variables that were significantly correlated (p < 0.05) with the canonical axes were salinity, Sr and Ba content. The concentration of K, Na, Ca, and Mg were removed because they were statistically co-linear. Salinity and Sr were correlated with the first axis (r = 0.99 and 0.90, respectively), while Ba was correlated with the second axis (r = 0.77). Redundancy Analysis clearly defines two ostracod assemblages, determined by the environmental conditions. The first axis shows the gradient of salinity, with the highest values of this variable and Sr concentration in the positive sector with Limnocythere aff. staplini. The rest of the species are grouped in the negative quadrant of the first axis, indicating their preference for freshwater, with C. vidua, H. incongruens, H. helenae, I. ramirezi, P. smaragdina, Chlamydotheca sp. and Cyprididae indet. correlated with higher Ba concentration, whereas P. incae, D. stevensoni and Chlamydotheca arcuata show a weak negative relationship with Ba content. It should be noted that both Chlamydotheca species, despite being present only in the Carapacho warm spring, show different preferences for Ba; this is because Chlamydotheca sp. was found during spring and C. arcuata was found in both spring and autumn.
Fig. 7
Redundancy analyses (RDA) diagram showing the correlation among eleven ostracod species and three environmental variables
Taxonomic analysis of ostracods from the Laguna Llancanelo area has extended the distribution range of all taxa, with some species exclusively found in this area. It shows that the assemblages are formed by taxa recorded only in the Arid Diagonal, but with others found in the subtropical Chaco-Pampean and/or Patagonian areas.
The species Limnocythere aff. staplini was the most abundant (>9700 recovered valves), lives mostly in ephemeral saline Laguna Llancanelo, and is described based on hard parts. Limnocythere aff. staplini is very similar to L. staplini, known from North America (Canada, USA) (Gutentag and Benson 1962; Delorme 1971; Curry et al. 2012) and is also comparable to L. cusminskyae from the Argentinian Pampas (Ramón Mercau et al. 2014). Despite the similarities in valve shape and size, the hinges are clearly different, with the female RV having smooth and prominent anterior and posterior teeth in L. aff. staplini, both teeth crenulated in L. staplini, and the anterior tooth smooth and the posterior crenulated, both small, in L. cusminskyae. Delorme (1971) in a revision of the Family Limnocytheridae in Canada, indicated that the shape of the valve of L. staplini is very distinctive, which in part supports the notion that L. aff. staplini is a phenotypic variant, although the hinges are different as is the size, with valves of L. aff. staplini a little longer. Our specimens, however, are shorter compared with L. cusminskyae. A decision as to whether L. aff. staplini is a new species or an ecophenotype of L. staplini must wait until the study of soft tissues provides definitive evidence. Limnocythere aff. staplini, despite the disjunct distribution, has similar ecological and hydrochemical requirements as L. staplini [fresh to saline lentic waters of 0.2–43 g L−1 (Forester 1983) and high concentrations of Ca (Curry et al. 2012), while L. cusminskyae prefers fresh to mesosaline salinity (0.5–20 g L−1) and bicarbonate-rich waters (Ramón Mercau et al. 2014)].
The species Chlamydotheca arcuata, originally described from temporary ponds and creeks (Díaz and Lopretto 2011), was recorded for the first time in a warm spring. The two species of Chlamydotheca, C. arcuata and Chlamydotheca sp. were found only in the Carapacho warm spring, indicating a preference for relatively constant temperature (~20 °C). These taxa, however, were not found during winter, indicating that daylight, i.e. hours of insolation, may be the limiting factor. This confirms the value of seasonal collections when building a modern dataset, as it would not have been possible to establish that Carapacho had a constant, year-round warm temperature if only summer sampling had been done. Nor would it be possible to acquire information on ostracod species preferences and life cycles.
The main environmental factor segregating wetlands in the Llancanelo area is salinity, with the evaporative saline lake set apart from all other freshwater wetlands (Fig. 5). Concentration of K is the factor that further separates the freshwater wetlands. When the analysis includes the ostracod taxa and their abundances, Carapacho warm spring is separated from the rest of the sites because it is the only locality with the two Chlamydotheca species found in the area. The creeks and saline lake are grouped together, separated from the other lentic freshwater sites. This is probably a consequence of the fact that the creeks are ephemeral and water is restricted to very shallow environments in both summer and winter.
The halophile association of Laguna Llancanelo is characterized by abundant Limnocythere aff. staplini, which is present during all four seasons, accompanied by low numbers of C. vidua during the summer months. Limnocythere aff. staplini develops large populations, with 91% of its individuals living in the highly variable Laguna Llancanelo. The species is also present in the rheocrene cold spring and creeks. Seventy-one percent of Cypridopsis vidua specimens were found in the creeks, and it is present in low numbers in Laguna Llancanelo at salinities <29 g L−1. Limnocythere aff. staplini is absent in the warm spring and limnocrene spring and rare at other flowing freshwater sites, but is very abundant in the lake when salinity reaches 42.3 g L−1 during summer. Limnocythere aff. staplini can be found in the lake at different water depths, commonly represented by large numbers of female, male and juvenile individuals. In this study, we also found that L. aff. staplini prefers higher concentrations of K, Na and Ca; and high levels of Sr and Mg (Table EMS S2).
The fresh water to subsaline ostracod assemblage is more diverse, characterized by the presence of Potamocypris smaragdina, Heterocypris incongruens, Chlamydotheca sp., C. arcuata, Penthesilenula incae, Darwinula stevensoni, Herpetocypris helenae, Ilyocypris ramirezi and Cyprididae indet. The cosmopolitan species H. incongruens and D. stevensoni live in the freshwater springs and slow-flowing creeks. Potamocypris smaragdina and P. incae are restricted to freshwater, mostly in lentic environments, although the former is also present in the Río Malargüe delta. They can be considered stenohaline species, as indicated by Ramón Mercau et al. (2012), with P. smaragdina having affinity for lentic or low-current environments, whereas P. incae lives in environments with a constant freshwater supply (Cusminsky et al. 2011). Ilyocypris ramirezi has been found in cold springs and lotic environments, particularly the Río Malargüe delta, in fresh to subsaline (<3 g L−1) waters, but is absent in the lake. This does not support previous records describing it as ‘euryhaline’ (Markgraf et al. 2003; Schwalb et al. 2002; Laprida 2006; Ramón Mercau et al. 2012). The last authors suggested a salinity tolerance from freshwater to mesosaline for this species. The cosmopolitan species D. stevensoni and H. incongruens were found in cold and warm springs, as well as in the creeks, showing that these taxa tolerate a wide temperature range, contrary to Külköylüoglu (2003), who associated D. stevensoni with mid-range temperatures, and Meisch (2000), who indicated a cold-water preference for H. incongruens.
Conclusions
The 11 taxa of ostracods found in saline Laguna Llancanelo and related freshwater wetlands can be considered moderately diverse, and the number compares well with the fifteen taxa described from northern Patagonia and 12 from southern Patagonia (Ramón Mercau et al. 2012). This study shows that Herpetocypris helenae, Limnocythere aff. staplini, Chlamydotheca sp. and the undetermined Cyprididae are restricted to the Laguna Llancanelo area.
The ‘subtropical’ taxon Chlamydotheca arcuata, with exclusive American distribution, and Penthesilenula incae, found in Neotropical areas, were previously recorded in Argentina only in wetlands from the Puna (northwest Argentina) and Patagonia. Potamocypris smaragdina, a taxon with Palearctic–Neotropical distribution and cosmopolitan Darwinula stevensoni and Heterocypris incongruens, have been found in both the Pampean and Patagonian areas, together with the endemic Ilyocypris ramirezi.
This study shows that Limnocythere aff. staplini is a euryhaline taxon, indicative of shallow, saline water bodies that are subject to changes in salinity and temperature, and also related to Na, K, Sr and Ca in the water. Other taxa, except C. vidua, are typically from freshwater, in some cases showing affinity to Ba content (C. vidua, H. incongruens, H. helenae, I. ramirezi, P. smaragdina and Cyprididae indet.). After salinity and ionic composition of the water, temperature and pH are the ecological variables that influence these ostracods. At present, arid conditions characterize the area, with extensive salt crusts deposited around Laguna Llancanelo. These freshwater bodies serve as refuges for species that cannot tolerate higher salinity.
Our data suggest that Limnocythere aff. staplini, Herpetocypris helenae, Chlamydotheca sp. and the Cyprididae indet. are found only in the Arid Diagonal, though more studies are needed to confirm the endemic nature of these taxa. This work provides the first modern database of ostracods and water chemistry (with SEM images, water variables, and seasonal data) from South America. Additional data from other South American regions will provide information to construct a continental dataset that can be applied to ostracods in quantitative paleolimnological and paleoclimatic reconstructions, and can be compared with Northern Hemisphere reconstructions to better understand biogeographic patterns. Despite the preliminary nature of this database, this study provides modern analogue data needed to infer past climate and ecological conditions on the continent, particularly in the Laguna Llancanelo area where a multidisciplinary paleolimnological study is underway.
References
Alvarez A, Wetten C (2004) Hidroquímica y sistemas de flujo subterráneo en áreas próximas al humedal de Llancanelo, tenor natural de hidrocarburos, Mendoza, Argentina. XXXIII Congreso Internacional AIH, 7º ALHSUD, Zacatecas, Actas 32–33
Ballent SC, Díaz AR (2011) Contribution to the taxonomy, distribution and paleoecology of the early representatives of Penthesilenula Rossetti & Martens, 1998 (Crustacea, Ostracoda, Darwinulidae) from Argentina, with the description of a new species. Hydrobiologia 688:125–138. doi:10.1007/s10750-011-0658-8
Chivas AR, De Deckker P, Shelley JMG (1983) Magnesium, strontium and barium partitioning in non-marine ostracode shells and their use in paleoenvironmental reconstructions—a preliminary study. In: Maddocks RF (ed) Applications of Ostracoda. University of Houston Geoscience, Houston, pp 238–249
Chivas AR, De Deckker P, Shelley JMG (1986a) Magnesium and strontium in non-marine ostracod shells as indicators of palaeosalinity and palaeotemperature. Hydrobiologia 143:135–142. doi:10.1007/bf00026656
Chivas AR, De Deckker P, Shelley JMG (1986b) Magnesium content of non-marine ostracode shells: a new palaeosalinometer and palaeothermometer. Palaeogeogr Palaeoclimatol Palaeoecol 54:43–61. doi:10.1016/0031-0182(86)90117-3
Ciocco NF, Scheibler EE (2008) Malacofauna of the littoral benthos of a saline lake in southern Mendoza, Argentina. Fundam Appl Limnol 172:87–98. doi:10.1127/1863-9135/2008/0172-0087
Curry BB (1999) An environmental tolerance index for ostracodes as indicators of physical and chemical factors in aquatic habitats. Palaeogeogr Palaeoclimatol Palaeoecol 148:51–63. doi:10.1016/s0031-0182(98)00175-8
Curry BB, Delorme LD, Smith AJ, Palmer DF, Stiff BJ (2012) The biogeography and physicochemical characteristic of aquatic habitats of freshwater ostracods in Canada and United States. In: van der Meer JJM (ed) Ostracoda as proxies for Quaternary climate change, vol 17. Developments in Quaternary science series. Elsevier, Amsterdam, pp 85–116
Cusminsky GC, Pérez PA, Schwalb A, Whatley R (2005) Recent lacustrine ostracods from Patagonia, Argentina. Rev Esp Micropaleontol 37:431–450
Cusminsky GC, Martínez D, Bernasconi E (2006) Foraminíferos y ostrácodos de sedimentos recientes del estuario de Bahía Blanca, Argentina. Rev Esp Micropaleontol 38:395–410
D’Ambrosio DS, Díaz AR, García A, Claps MC (2015) First description of the soft part anatomy of Ilyocypris ramirezi Cusminsky & Whatley (Crustacea, Ostracoda) from Argentina, South America. Zootaxa 3957:059–068. doi:10.11646/zootaxa.3957.1.4
D’Ambrosio DS, Claps MC, García A (2016a) Zooplankton of a protected and vulnerable wetland system in southern South America (Llancanelo área, Argentina). Int Aquat Res 8:65–80. doi:10.1007/s40071-016-0125-2
D’Ambrosio DS, Díaz AR, García A, Claps MC (2016b) First record and revised description of Herpetocypris helenae G. W. Müller, 1908 (Crustacea, Ostracoda) from the Neotropics (central-west Argentina). Spixiana 39:29–38
De Deckker P, Forester RM (1988) The use of ostracods to reconstruct continental palaeoenvironmental records. In: De Deckker P, Colin JP, Peypouquet JP (eds) Ostracoda in the earth sciences. Elsevier, New York, pp 175–199
De Francesco CG (2010) Moluscos y paleoambientes del centro-oeste de Argentina durante el Cuaternario. In: Zarate M, Gil A, Neme G (eds) Condiciones paleoambientales y ocupaciones humanas durante la transición Pleistoceno-Holoceno y Holoceno de Mendoza. Publicaciones de la Sociedad Argentina de Antropología, Buenos Aires, pp 151–174
De Francesco CG, Hassan GS (2009) The significance of molluscs as paleoecological indicators of freshwater systems in central-western Argentina. Palaeogeogr Palaeoclimatol Palaeoecol 274:105–113. doi:10.1016/j.palaeo.2009.01.003
Forester RM (1983) Relationship of two lacustrine ostracode species to solute composition and salinity: implication for paleohydrochemistry. Geology 11:435–438
Forester RM, Smith AJ, Palmer DF, Curry BB (2005) North American ostracode database “NANODe” version 1. http://www.kent.edu/NANODe/. Accessed 22 May 2016
García A (1999) Quaternary charophytes from Salina del Bebedero, Argentina: their relation with extant taxa and paleolimnological significance. J Paleolimnol 21:307–323. doi:10.1023/A:1008067112334
Gouramanis C, Wilkins D, De Deckker P (2010) 6000 years of environmental changes recorded in Blue Lake, South Australia, based on ostracod ecology and valve chemistry. Palaeogeogr Palaeoclimatol Palaeoecol 297:223–237. doi:10.1016/j.palaeo.2010.08.005
Hammer UT, Shamess J, Haynes RC (1983) The distribution and abundance of algae in saline lakes of Saskatchewan, Canada. Hydrobiologia 105:1–26. doi:10.1007/bf00025173
Isla FI, Ruiz Barlett E, Márquez J, Urrutia A (2005) ENSO signal recognized by NDVI images from Llancanelo shallow lake, Mendoza, Argentina. XVI Congreso Geológico Argentino, La Plata, Actas 4, pp 333–338
Külköylüoĝlu O (2003) Ecology of freshwater Ostracoda (Crustacea) from lakes and reservoirs in Bolu, Turkey. J Freshw Ecol 18:343–347. doi:10.1080/02705060.2003.9663968
Laprida C (2006) Ostrácodos recientes de la llanura pampeana, Buenos Aires, Argentina: ecología e implicancias paleolimnológicas. Ameghiniana 43:181–204
Liberto R, Mesquita-Joanes F, César I (2012) Dynamics of pleustonic ostracod populations in small ponds on the Island of Martín García (Río de la Plata, Argentina). Hydrobiologia 688:47–61. doi:10.1007/s10750-011-0600-0
Markgraf V, Bradbury JP, Schwalb A, Burns SJ, Stern C, Ariztegui D, Gilli A, Anselmetti FS, Stine S, Maidana N (2003) Holocene palaeoclimates of southern Patagonia: limnological and environmental history of Lago Cardiel, Argentina. Holocene 13:581–591. doi:10.1191/0959683603hl648rp
Martens K, Schon I, Meisch C, Horne DJ (2008) Global biodiversity of non-marine Ostracoda (Crustacea). In: Balian EV, Lévȇque C, Segers H, Martens K (eds) Freshwater animal diversity assessment. Hydrobiologia 599:185–193
Meisch C (2000) Freshwater Ostracoda of Western and Central Europe. In: Schwoerbel J, Zwick P (eds) Süsswasserfauna von Mitteleuropa 8/3. Spektrum Akademischer Verlag, Heidelberg, p 522
Mourguiart P, Corrège T, Wirrmann D, Argollo J, Montenegro ME, Pourchet M, Carbonel P (1998) Holocene palaeohydrology of Lake Titicaca estimated from an ostracod-based transfer function. Palaeogeogr Palaeoclimatol Palaeoecol 143:51–72. doi:10.1016/s0031-0182(98)00068-6
Ostera HA, Dapeña C (2003) Environmental isotopes and geochemistry of bañado Carilauquen, Mendoza, Argentina. In: IV South American symposium on isotope geology, Actas 461–464
Palacios-Fest MR, Cusminsky GC, McGlue MM (2016) Late Quaternary lacustrine ostracods (Ostracoda, Crustacea) and charophytes (Charophyta, Charales) from the Puna Plateau, Argentina. J Micropalaeontol 35:66–78
Peralta P, Fuentes V (2005) Fitobentos, fitoplancton y zooplancton litoral del bañado de Carilauquen, Cuenca de Llancanelo, Mendoza, Argentina. Limnetica 24:183–198
Piovano EL, Ariztegui D, Córdoba F, Cioccale M, Sylvestre F (2009) Hydrological variability in South America below the Tropic of Capricorn (Pampas and Patagonia, Argentina) during the last 13.0 ka. In: Vimeux F, Sylvestre F, Khodri M (eds) Past climate variability in South America and surrounding regions, Developments in paleoenvironmental research, vol 14. Springer, Berlin, pp 323–351. doi:10.1007/978-90-481-2672-9-14
Ramón Mercau J, Laprida C, Massaferro J, Rogora M, Tartari G, Maidana NI (2012) Patagonian ostracods as indicators of climate related hydrological variables: implications for paleoenvironmental reconstructions in southern South America. Hydrobiologia 694:235–251. doi:10.1007/s10750-012-1192-z
Ramón Mercau J, Plastani MS, Laprida C (2014) A review of the genus Limnocythere (Podocopida: Limnocytheridae) in the Pampean region (Argentina), with the description of a new species, Limnocythere cusminskyae sp. nov. Zootaxa 3821:026–036. doi:10.11646/zootaxa.3821.1.2
Ramos VA, Folguera A (2011) Payenia volcanic province in the Southern Andes: an appraisal of an exceptional Quaternary tectonic setting. J Volcanol Geotherm Res 201:53–64. doi:10.1016/j.jvolgeores.2010.09.008
Scheibler EE, Ciocco NF (2011) Distribution of macroinvertebrate assemblages along a saline wetland in harsh environmental conditions from Central-West Argentina. Limnologica 41:37–47. doi:10.1016/j.limno.2010.03.001
Scheibler EE, Ciocco NF (2013) Diversity of aquatic insects and other associated macroinvertebrates in an arid wetland (Mendoza Province, Argentina). Rev Soc Entomol Argent 72:41–53
Schwalb A, Burns SJ, Cusminsky G, Kelts K, Margraff V (2002) Assemblage diversity and isotopic signals of modern ostracods and host waters from Patagonia, Argentina. Palaeogeogr Palaeoclimatol Palaeoecol 187:323–339. doi:10.1016/s0031-0182(02)00484-4
ter Braak CJF (1986) Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67:1167–1179. doi:10.2307/1938672
ter Braak CJF, Smilauer P (2002) CANOCO reference manual and CanocoDraw for Windows user’s guide: software for canonical community ordination (version 4.5). Microcomputer Power, Ithaca
ter Braak CJF, Verdonschot PFM (1995) Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat Sci 57:255–289. doi:10.1007/BF00877430
We thank the Dirección de Recursos Naturales de la Provincia de Mendoza for facilitating the research on this Ramsar site. We also thank park ranger Fabricio Ayala for his invaluable help in the field. We thank Silvina Lassa, Nick Mackie and Rafael Urréjola for technical assistance with the SEM. The assistance of David Wheeler and Lili Yu at the University of Wollongong is greatly appreciated. This work was supported by a grant from the Agencia Nacional de Promoción Científica y Tecnológica, FONCYT (Argentina) (Grant Raíces 2006 – PICT 1311), which also enabled DSD to take a visiting position in Wollongong.
Author information
Authors and Affiliations
Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (CCT-Mendoza CONICET), Av. Ruíz Leal, 5500, Ciudad de Mendoza, Provincia de Mendoza, Argentina
D. Sabina D’Ambrosio
GeoQuEST Research Centre, School of Earth and Environmental Sciences, University of Wollongong, Wollongong, NSW, 2522, Australia
Adriana García & Allan R. Chivas
Instituto de Limnología Dr. Raúl A. Ringuelet (CCT-La Plata CONICET), Boulevard 120 y 62, 1900, La Plata, Provincia de Buenos Aires, Argentina
D’Ambrosio, D.S., García, A., Díaz, A.R. et al. Distribution of ostracods in west-central Argentina related to host-water chemistry and climate: implications for paleolimnology.
J Paleolimnol58, 101–117 (2017). https://doi.org/10.1007/s10933-017-9963-1