
Abstract
The Morrison Formation bears one of the most diverse assemblages of Late Jurassic terrestrial vertebrates worldwide. A recently discovered site in eastern Utah, the Cisco Mammal Quarry (CMQ), shows excellent preservation and small vertebrates (particularly mammals) collected thus far are very diverse. Two isolated petrosals from the CMQ, representing the same taxon, are described here based on CT data. Several plesiomorphic mammalian characters are present, including a horizontal crista interfenestralis, unfloored cavum supracochleare, a perilymphatic foramen, and an open perilymphatic groove. By contrast, a well-developed tractus foraminosus is present for distribution of cochlear nerve fibers and the cochlear endocast makes one full turn, as in early therians such as Prokennalestes. This latter derived condition is unrecorded in the Jurassic; the mammal fauna characteristic of the Morrison is dominated by docodontans, eutriconodontans, dryolestoids (dryolestids and paurodontids), and multituberculates. All currently-known examples of these high-level taxa possess a relatively short, straight or curved cochlea. Therians are not known from the Morrison Formation, but these petrosals invite comparison with some stem therians of South America, the meridiolestidans, which also bear a full coil, but differ from the Morrison petrosals in aspects of external morphology. While the taxonomic affinity of the specimens we describe here remains uncertain, the unique combination of petrosal characters is evidence that our current view of petrosal/inner ear evolution is oversimplified. With the presence of derived and plesiomorphic features character conflict is unavoidable, pointing to a complex interplay of external petrosal osteology under the influence of neurovascular and middle ear evolution and the (perhaps more independent) biophysical demands of inner ear function.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The petrosal is a character-rich skull element unique to mammals and their close relatives (Bast and Anson 1949). While the majority of morphological characters in most recent craniodental analyses are still drawn from the dentition and tooth-bearing elements, the ear region typically accounts for the next largest share of characters (Luo et al. 2017; Bi et al. 2018; Krause et al. 2020). Moreover, many major branches of cranial nerves V–XII have an intimate relationship with the petrosal and basicranium, as does much of the proximal cranial vascular supply. Grooves and foramina in the petrosal can be used to reconstruct the soft tissues associated with them and, by inference, trace morphological transformations expected to be biologically important. Basicranial and petrosal anatomy has been used with some success to infer shifts in major blood vessels in various lineages (Wible 1986, 1987; Rougier et al. 1992; Rougier and Wible 2006), cranial nerve evolution (Presley 1993; Kemp 2009), braincase wall development (Kemp 1983; Hopson and Rougier 1993; Koyabu et al. 2014; Crompton et al. 2018), and inner ear evolution (Meng and Fox 1995a, b; Luo 2001; Luo et al. 2011a, 2016; Schultz et al. 2016). Dr. J. R. Wible has pioneered the incorporation of these characters in the evolutionary study of mammals; we see our paper here as a direct consequence of his influence and a fitting tribute to an exceptional scientist. Extraction of a digital endocast of the bony labyrinth within the petrosal can be used to study morphological evolution and, under certain conditions, auditory capabilities can be estimated (Ekdale 2016; Harper and Rougier 2019) and locomotor habits inferred (Pfaff et al. 2015; Ekdale 2016). In fact, the advent of CT technology has resulted in a resurgence in the study of petrosal anatomy by opening classic and historic specimens to endocast extraction and a great increase in available data.
The Morrison Formation has been foundational for much of our understanding of Mesozoic terrestrial vertebrate evolution since the discovery of the first dinosaurs in Colorado in 1877 (Marsh 1877a, b). The first Jurassic fossil mammal was discovered soon after (Marsh 1878), and the Morrison would yield what remains the most diverse assemblage of Late Jurassic mammals and close relatives worldwide (Kielan-Jaworowska et al. 2004). The classic Como Bluff localities in central Wyoming (Simpson 1929; Prothero 1981) are typical of most of the Morrison record, in that large numbers of specimens from diverse groups have been recovered but preservation is mainly limited to tooth-bearing elements. In contrast, localities within the Fruita Paleontological Area in western Colorado are less taxonomically diverse but preserve much more complete specimens, often 3D and partially articulated or associated (Callison 1987; Engelmann and Callison 1998; Luo and Wible 2005; Kirkland 2006). Despite the long history of collection efforts, only a handful of cranial specimens have been described from the Morrison. Petrosals are known by three specimens: a broken petrosal of uncertain affinities from the Como area (Prothero 1983); an associated petrosal from the eutriconodontan Priacodon, from Fruita (Rougier et al. 1996); and an isolated petrosal, also from Fruita, referred to Dryolestoidea (Hughes et al. 2015). The iconic dryolestid Dryolestes priscus is known by jaws and referred upper dentitions from several quarries in the Morrison Formation (Simpson 1929; Prothero 1981) but no basicranial elements. However, a petrosal of the related species, Dryolestes leiriensis, is known from the slightly older Portuguese locality of Guimarota (Luo et al. 2012); for purposes herein, we assume a uniform petrosal morphology among species of Dryolestes.
Recent exploration of the Morrison Formation in eastern Utah by a joint field crew from the University of Louisville and the Sam Noble Oklahoma Museum of Natural History resulted in the discovery of a new microvertebrate locality in 2015, near the ghost town of Cisco (Davis et al. 2018). The Cisco Mammal Quarry (CMQ) is situated within the Brushy Basin Member and is similar in lithology to the section containing the mammal-bearing sites in the Fruita Paleontological Area (Kirkland 2006; Davis et al. 2018). Excavation is ongoing and much of the collection is still being prepared for study, but preliminary data show rather high taxonomic diversity of mammals; importantly, preservation is good to excellent. Most specimens are recovered as isolated elements, but associated and even partially articulated skeletons of small vertebrates including mammals are not uncommon.
Here, we describe two isolated but well-preserved petrosals from the CMQ, including digital endocasts of the bony labyrinth and fragmentary stapes. The ear region in mammals is radically modified in comparison to other amniotes, especially with regard to the auditory ossicles, vestibule, and cochlea (Kemp 2016; Luo et al. 2016). Modern therians possess a coiled cochlea, and character optimization in recent analyses suggest that a coiled cochlea was present in the last common ancestor of marsupials and placentals (Luo et al. 2016; Bi et al. 2018). Living monotremes, on the other hand, have a straight osseous cochlear canal containing a hook-shaped cochlear duct (Denker 1901; Alexander 1904; Schultz et al. 2016). Stem mammalian groups possess a similarly straight or slightly curved cochlear duct, as do some extinct Crown mammalian lineages (e.g., Ruf et al. 2009; Luo et al. 2011a, b; Hughes et al. 2015; Harper and Rougier 2019). Juramaia is a therian of reported Jurassic age (Luo et al. 2011b). Its ear region is unknown, but its geologic age implies with some certainty that coiling of the cochlea was achieved during the Jurassic in the therian lineage or its stem. Partial cochlear coiling is present in the Early Cretaceous stem therian Vincelestes (Rougier et al. 1992) and in Late Jurassic dryolestoids (Ruf et al. 2009; Luo et al. 2012), and a full coil is achieved in the endemic Late Cretaceous South American meridiolestidans, likely independently (Rougier et al. 2009).
Differences also exist among living and fossil mammal groups in construction of the sidewall of the braincase, which is intimately tied to the petrosal. The anterior lamina of the petrosal forms a substantial portion of the braincase in basal mammaliaforms (e.g., morganucodontans) and some other extinct groups, and this element is expansive in monotremes (e.g., Wible and Hopson 1993; Hopson and Rougier 1993). In contrast, therians lack an anterior lamina of the petrosal, even though the primitive condition is present in basal cladotheres and the stem therian Vincelestes (Rougier et al. 1992; Crompton et al. 2018). The CMQ petrosals are too damaged to provide much evidence of the construction of the lateral braincase wall, beyond establishing the presence of an anterior lamina and its involvement in the formation of a distinct lateral flange. However, the CMQ petrosals provide a novel combination of derived cochlear coiling with a rather primitive tympanic region construction, illustrating the complicated interplay of these character systems during mammal evolution.
The petrosal is a complex region of the skull with a rich tradition of study via comparative morphology and development. Here, we provide a general description to orient the reader. The petrosal is an endochondral element of the mammal skull that develops from the otic capsule and encloses the organs of hearing and balance. Traditionally (De Beer 1929; Bast and Anson 1949), the petrosal is divided for descriptive purposes into two components: the pars cochlearis, which houses the cochlea and saccule; and pars canalicularis, which houses the utricle and semicircular ampullae and canals. Additionally, adjoined to the petrosal proper of most non-therian mammals, including monotremes, the adult petrosal forms part of the lateral wall of the braincase via the anterior lamina, which itself is formed by a combination of appositional bone to the ossified otic capsule and membranous ossification of the embryonic sphenoobturator membrane (Zeller 1989; Rougier et al. 1992; Wible and Hopson 1995). The horizontal portion, forming part of the roof of the tympanic cavity and part of the floor of the cavum epiptericum, is the lateral trough. This region extends posteriorly to the primary facial foramen and conveyed vasculature to the mesocranium and auditory region. The thickened ventral edge of the anterior lamina is the lateral flange, forming part of the lateral boundary of the tympanic region and the inferior portion of the braincase wall. The anterior lamina is moderate-sized in basal mammaliaforms such as Morganucodon and the stem therian Vincelestes, very extensive in some non-therian lineages such as monotremes, multituberculates and eutriconodontans, and lost in therians (see review in Rougier et al. 1992). It is worth mentioning that a rudimentary anterior lamina has been controversially identified in some Paleogene metatherians from South America (Marshall and Muizon 1995; Ladevèze and Muizon 2010), but evaluation of this claim is beyond the scope of this paper. The petrosal, via the lateral trough, lateral flange, and anterior lamina, completed the lateral wall of the braincase and extra-promontorial portions of the rostral ear region.
Institutional abbreviation.—OMNH, Sam Noble Oklahoma Museum of Natural History, Norman, OK.
Materials and Methods
The two isolated petrosals (a right and a left) are nearly identical in morphology and preservation. The Cisco Mammal Quarry is sampled through hand-quarrying of blocks harvested from along the quarry face (Davis et al. 2018), and identification of specimens is typically impossible until some preparation or CT scanning is done. Although we are uncertain how close they were to each other in the quarry, the possibility that these petrosals belong to the same individual cannot be excluded. Following limited mechanical preparation, the specimens were scanned at the University of Texas High-Resolution X-ray CT Facility in Austin, TX, using a Zeiss Xradia, resulting in voxels measuring 7.98 microns for the right petrosal and 9.53 microns for the left petrosal. 3D data visualization and digital segmentation were performed in Avizo 9.1 (Thermo Fisher Scientific).
Description
Both CMQ petrosals were discovered via quarrying and each was split into part-counterpart during discovery, with the break passing through the mastoid region and subarcuate fossa. The halves were glued back together and much of the enclosing rock removed prior to CT scanning. Both petrosals (OMNH 80536, right petrosal; OMNH 80537, left petrosal) are isolated and generally well preserved, including most of the pars cochlearis and pars canalicularis (Figs. 1 and 2; measurements provided in Table 1). We cannot rule out the possibility (and in fact consider it likely) that the petrosals belonged to the same individual skull. Portions of the lateral flange and anterior lamina are present, but this region is heavily damaged and deformed in both specimens, such that the morphology and extent of the anterior lamina cannot be established with confidence, but enough remains to be certain of its presence. The presumed orientation of the petrosals in the skull is based on the well-preserved contact with the basioccipital which, based on character distribution, is inferred to have been oblique, with this suture running parasagittally or slightly diverging posteriorly. The standard ventral (tympanic) and dorsal (endocranial) views are described below; the specimens are too artificially compressed for a separate lateral view to be informative. We follow the description of the petrosal with a description of endocasts of the bony labyrinth, based on virtual 3D renderings. In the process of digitally preparing the endocasts, a partial stapes was discovered within the cochlear canal of each specimen; this will be described last.

CT rendering of OMNH 80536, right petrosal from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ventral (a), dorsal (b), lateral (c), and medial (d) views

CT rendering of OMNH 80537, left petrosal from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ventral (a), dorsal (b), lateral (c), and anterior (d) views. Abbreviations: lf, lateral flange; lt, lateral trough. Asterisk indicates foramen enclosed in lateral flange
Ventral view.—The most prominent features of the petrosal in ventral view are the paroccipital process and the promontorium, which is round but only somewhat inflated (not nearly to the degree seen in therians). Two large openings are present on the posterior end of the promontorium (Fig. 3). The lateral opening, the fenestra vestibuli, would have contacted the footplate of the stapes in life and is somewhat oval shaped. The average stapedial ratio (Segall 1970; Rougier et al. 1996) of the two specimens, a measure of the eccentricity of this opening with a value of 1 being circular, is 1.43 (Table 1). The edges of the fenestra are approximately flush with the surface of the promontorium and therefore a fossula fenestra vestibuli (MacPhee 1981) is lacking, the condition considered primitive for mammals. Two sulci are present along the rim of the fenestra vestibuli. A shallow but distinct notch on the medial edge of the fenestra vestibuli continues anteriorly as a well-defined transpromontorial groove. We interpret this to have accommodated the internal carotid artery, which maintained contact with the promontorium for nearly its entire length (Figs. 4 and 5). On the posteromedial corner of the fenestra vestibuli is a deep sulcus directed medially toward the crista interfenestralis, passing between a thin but sharp dorsal crest and a heavy lip of bone from the back of the promontorium. We interpret that the portion of the internal carotid artery immediately proximal to the stapedial artery would have occupied this sulcus (ica and gica; Figs. 4, 5 and 6). Given the course of the transpromontorial groove and predicted orientation of the petrosal, it would be expected that paired carotid foramina were present in the ventral surface of the basisphenoid, possibly in close proximity to each other. The medial opening at the back of the promontorium is the perilymphatic foramen. This foramen is considerably more elongate than the fenestra vestibuli, the difference in shape being accentuated by the lack of a complete processus recessus (DeBeer 1929). In living mammals, the processus recessus develops during ontogeny to form a partition between the perilymphatic foramen and the perilymphatic duct, resulting in the formation of the fenestra cochleae which is occupied in living mammals by the secondary tympanic membrane (see review in Zeller 1985a). The dorsomedial edge of the perilymphatic foramen narrows considerably and the perilymphatic groove is rather well defined (resembling Priacodon in this regard, Rougier et al. 1996), but there is no evidence on either specimen that a bony partition was present (Figs. 1a, d and 3a), fully enclosing the perilymphatic duct. A cochlear aqueduct is therefore lacking. A small and shallow emargination for the jugular foramen is present along the medial border of the petrosal and cannot be fully distinguished from the notch for the passage of the perilymphatic duct. Based on the preserved margin, the jugular foramen was much smaller than the perilymphatic foramen and there is no evidence of a jugular recess or fossa in the vicinity of the perilymphatic or jugular foramen. The bone posterior to the perilymphatic foramen is smooth and slopes ventrally toward the well-developed caudal tympanic process of the petrosal (CTPP); this surface is bounded medially by a sharp ridge that formed the anterior margin of the robust suture with the exoccipital. Despite the relatively circumscribed space around the perilymphatic foramen, which is encircled by the posterior wall of the promontorium, the crista interfenestralis, and the anterior surface of the CTPP, there is no fossula fenestra cochleae (MacPhee 1981) developed around the perilymphatic foramen. There seems to be no recess for a perilymphatic cistern in the recessus scalae tympani, the immediate vicinity of the foramen, a feature found in monotremes (Zeller 1989).

Illustrations of OMNH 80536 (a, c, e), right petrosal, and OMNH 80537 (b, d, f), left petrosal, both from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ventral (a, b), dorsal (c, d), and lateral (e, f) views. Abbreviations: ace, anterior opening of cavum epiptericum; al, anterior lamina; av, aqueductus vestibuli; ci, crista interfenestralis; cp, crista parotica; cr, crista petrosa; ctpp, caudal tympanic process of petrosal; dag, dorsal ascending groove; ew, epitympanic wing; fi, fossa incudis; fo, foramen ovale; frs, foramen for ramus superior; frt, foramen for ramus temporalis; fv, fenestra vestibuli; gadm, groove for arteria diploëtica magna; goa, groove for occipital artery; gpd, groove for perilymphatic duct; gps, groove for prootic sinus; gss, groove for sigmoid sinus; iam, internal acoustic meatus; jn, jugular notch; lf, lateral flange; lt, lateral trough; ma, mastoid area; pan, pila antotica; pc, prootic canal; pf, perilymphatic foramen; pfc, prefacial commissure; pff, primary facial foramen; pp, paroccipital process; pr, promontorium; ptc, posttemporal canal; saf, subarcuate fossa; sf, stapedius fossa; smn, stylomastoid notch; sr, semilunar recess; th, tympanohyal; tpg, transpromontorial groove; vag, ventral ascending groove

Enlarged illustration of tympanic cavity and surrounding area in OMNH 80537, left petrosal from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ventral view (a); enlarged illustration of internal acoustic meatus of OMNH 80536, right petrosal from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in dorsomedial view (b). Abbreviations: av, aqueductus vestibuli; ci, crista interfenestralis; cp, crista parotica; cpcn, cribriform plate for cochlear nerve; ct, crista transversa; ctpp, caudal tympanic process of petrosal; dag, dorsal ascending groove; er, epitympanic recess; fai, foramen acousticum inferius; fas; foramen acousticum superius; fi, fossa incudis; frs, foramen for ramus superior; frt, foramen for ramus temporalis; fsv, foramen for saccular branch of vestibular nerve; fua, foramen for utriculoampullar branch of vestibular nerve; fv, fenestra vestibuli; gadm, groove for arteria diploëtica magna; gica; groove for internal carotid artery; iam, internal acoustic meatus; jn, jugular notch; lf, lateral flange; pan, pila antotica; pff, primary facial foramen; pp, paroccipital process; pr, promontorium; saf, subarcuate fossa; sf, stapedius fossa; smn, stylomastoid notch; sr, semilunar recess; th, tympanohyal; tpg, transpromontorial groove; vag, ventral ascending groove. Asterisk indicates a pocket of uncertain nature

CT rendering of OMNH 80536, right petrosal (partially opaque) and endocast of bony labyrinth (blue) from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ventral (a), and dorsal (b) views. Oblique lateral view of OMNH 80536 (c) and OMNH 80537 (d) to show fragmentary stapes (red) lodged within the cochlear canal, indicated by asterisks

Reconstruction of cranial neurovasculature in the ear region of OMNH 80536, right petrosal from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ventral (a) and lateral (b) views. Abbreviations: adm, arteria diploëtica magna; fn, hyomandibular branch of facial nerve; gg, geniculate ganglion; gpn, greater petrosal nerve; ica, internal carotid artery; lhv, lateral head vein; oa, occipital artery; ptv, posttrigeminal vein; ri, ramus inferior of stapedial artery; rs, ramus superior of stapedial artery; rt, ramus temporalis; sa, proximal stapedial artery
The lateral flange is well developed though not well preserved in the CMQ petrosals, wherein it is clearly present from the level of the facial foramen backward (see below; Fig. 3a, b). Rostral to this point, broken and displaced fragments suggest the presence of a lateral margin for the lateral trough, but it is uncertain how prominent it was. The presence of a substantial anterior opening for the cavum epiptericum (Fig. 3a) suggests that the alisphenoid likely contributed to form part of the lateral margin of such an aperture and some measure of the lateral wall (described also in other views below); this is the primitive condition, widely distributed among non-mammalian cynodonts (Fourie 1974; Hopson and Barghusen 1986).
Medially, the contact with the basioccipital is well preserved; there is a raised lip of bone along the anterior end of the contact, extending to the level of the anterior pole of the promontorium (Figs. 1d and 3a, b). This is the epitympanic wing and may also represent the position of the suture with the basisphenoid. Numerous vascular foramina, present along the medial edge of the petrosal, communicate with a faint groove curling around the anteromedial margin of the petrosal and converging with the perilymphatic duct posteriorly at the jugular notch. The lack of a distinct singular canal in this region suggests that the inferior petrosal sinus was poorly developed and that a rich intrapetrosal sinus network instead conveyed the blood from the substance of the petrosal.
Between the fenestra vestibuli and the perilymphatic foramen is the crista interfenestralis, which is a prominent ridge jutting posteriorly from the promontorium with its ventral margin oriented horizontally, reaching the CTPP at a point just medial to the base of the tallest preserved part of the paroccipital process (Fig. 3e, f). There is no continuous postpromontorial tympanic sinus, although the postpromontorial spaces on either side of the crista interfenestralis are substantial and distinct, and as such the middle ear space can be divided into two regions (Fig. 3): (1) a lateral tympanic sinus developed lateral to the crista interfenestralis and related to the fenestra vestibuli as a main feature; and (2) a medial tympanic sinus surrounding the perilymphatic foramen and associated structures, which also includes the recessus scalae tympani. Given that the paroccipital process and CTPP likely formed the posterior margin of the tympanic cavity, we would expect that both of these spaces would have converged ventral to the crista interfenestralis and promontorium in life, becoming part of a unified middle ear space. The lateral wall of the crista interfenestralis is quite vertical and bears a well delimited notch, possibly produced by the internal carotid artery which, if it occupied the groove immediately medial to the fenestra vestibuli, must have passed deep and close to the tympanic roof (Fig. 4). The deepest point of the pocket posterior to the fenestra vestibuli, tucked medially against the base of the crista interfenestralis, is the likely position of the fossa for the stapedius muscle (Figs. 3 and 5).
The paroccipital process is robust, very tall, and its tip curls anteriorly, although it is not completely preserved in either specimen (Fig. 3e, f). The paroccipital process extends ventrally well beyond the plane of the promontorium and represents the culmination of a robust thickening of the CTPP. The mastoid area on the ventral and posterior aspect of the paroccipital process is rough and well delimited by a sharp medial boundary and the presumed contact with the squamosal posteriorly and posterolaterally. Along the very extended lateral portion of the CTPP is the deeply incised notch which we interpret as the stylomastoid notch; in life this would have transmitted the hyomandibular ramus of the facial nerve (Figs. 4 and 5). The stylomastoid notch is well separated from the paroccipital process; the CTPP is relatively narrow in the vicinity of the stylomastoid notch, but projects further ventrally and gains in thickness as it approaches the apex of the paroccipital process. The lateral boundary of this notch is formed by a sharp vertical crest; at the dorsal end of this crest is a prominent projection of bone extending toward the fenestra vestibuli. We interpret this as the tympanohyal (Figs. 3 and 5a). The dorsal surface of the tympanohyal is concave and a clear path can be reconstructed for the course of the facial nerve, conforming to the plesiomorphic pattern expected in generalized basal mammals: it entered the ear region via the primary facial foramen, divided, and sent the hyomandibular branch posteriorly and medially underneath the tympanohyal before bending sharply ventrally and laterally to exit the ear region via the stylomastoid notch (see details below; Allin and Hopson 1992; Meng and Hou 2016). The tympanohyal is connected laterally, via a sharp, vertical crest, to a robust platform that is somewhat triangular in ventral view. This platform is a ventral projection of the thickened lateral ends of the CTPP adjacent to the surface we interpret as the facet for the squamosal. Continuing anteriorly from this platform is a sharply defined crest that passes dorsally and curls laterally; this is the crista parotica (Figs. 3 and 5). Given the intimate contact with the tympanohyal at the confluence of the CTPP and the crista parotica, we believe this platform to be a facet for the articulation of the stylohyal. The crista parotica forms a sharp lateral boundary for the roof of the tympanic cavity in the deepest portion of the lateral tympanic sinus, in life accommodating the facial nerve and major ear region vasculature. There is a deep, oval shaped pocket lateral to the crista parotica which is broader anteromedially and becomes deeper and narrower posterolaterally. The deepest posterior portion of this conspicuous fossa is here identified as the fossa incudis and would have housed the crus breve of the incus (MacPhee 1981; Zeller 1989). The anteromedial, broader portion of this pocket is the epitympanic recess for the incudomalleolar articulation (MacPhee 1981). While this pocket is entirely on the petrosal, the posterior and lateral walls of this region were likely in contact with the squamosal, a condition present or predicted in a variety of other Mesozoic mammals (Rougier et al. 1992, 1996; Rougier and Wible 2006); however, it is uncertain if the squamosal formed an actual lateral wall to the fossa incudis and epitympanic recess or simply extended near it.
Several major neurovascular grooves and structures are present lateral and anterolateral to the fenestra vestibuli (Figs. 3 and 5a). The most prominent of these is a round foramen at the bottom of a funnel-shaped fossa. This is the tympanic opening of a canal that can clearly be traced through the petrosal to the internal acoustic meatus; and is readily identified as the primary facial foramen. The widening fossa on the tympanic surface related to the primary facial foramen accommodated the geniculate ganglion of the facial nerve and represents an open cavum supracochleare. In both petrosals the primary facial foramen is exposed and there seems to be no evidence of damage (Figs. 1, 2 and 3); alternatively, the lack of a floor for the geniculate ganglion could reflect a juvenile condition, given that this floor ossifies relatively late in ontogeny (MacPhee 1981; Rougier et al. 1998). Well-defined grooves extend anteriorly and posteriorly from this opening and notch the margins of the cavum supracochleare. The large, posteriorly-directed groove is the sulcus for the hyomandibular branch of the facial nerve; the facial sulcus, which runs independently for a short distance and merges with a somewhat larger, transverse groove imprinted on the roof of the lateral tympanic recess. This second groove disappears beneath bone laterally at the anterior margin of the confluence of the epitympanic recess and lateral flange, and likely held the lateral head vein and a portion of the stapedial artery (Figs. 3, 4, 5 and 6).
Unlike the condition in basal taxa such as morganucodontans and many Mesozoic lineages (e.g., dryolestoids), where the tympanic openings of major vascular channels are fully exposed in ventral view (e.g., Rougier and Wible 2006), the lateral-most corner of the tympanic sinus in our specimens is partially obscured by bone (Fig. 3). Although the lateral flange has been deformed during preservation and the posterolateral region is difficult to visualize, we believe that these foramina in the lateral tympanic region were concealed beneath bone in life. Though not directly exposed to the tympanic cavity, the tympanic opening for the prootic canal is present in the expected position posterolateral to the primary facial foramen (as identified elsewhere by Wible and Hopson 1995) and can be traced vertically to the endocranial surface (described below). The large, transverse vascular groove in the lateral tympanic sinus passes beneath the lateral flange toward the opening of the prootic canal and divides. Of the two divisions, we identify the large, posterolaterally-directed opening as the foramen for the ramus superior of the stapedial artery; anteriorly, a channel continues along the lateral trough of the petrosal. While the lateral trough, lateral flange, and much of the anterior lamina are damaged, it is clear that a portion of the posterior lateral trough was fully enclosed by a curl of bone (Fig. 2d). It is unclear whether this short canal is formed by an inflection of the lateral flange (likely) or is sandwiched between the lateral flange and the roof of the lateral trough, but it was limited in extent and lateral to the position of the primary facial foramen. In the CMQ petrosals, this canal would have held the ramus inferior of the stapedial artery and, perhaps, the posttrigeminal vein if present (the canal is circular in section, so there is no clear osteological suggestion of concomitant vein-artery pair). A small groove is present on the medial margin of the lateral trough, where the trough meets the promontorium; this groove can be followed to a small notch on the anterior margin of the cavum supracochleare (Fig. 3a, b), and we regard this groove as having contained, in life, the greater petrosal branch of the facial nerve (Fig. 6). The notch and the greater petrosal nerve groove leading from it run horizontally at the level of the roof of the lateral trough, a plane considerably ventral to the root of the facial sulcus described above. There is some suggestion from the morphology of the right petrosal that this groove was also enclosed posteriorly, but it is uncertain due to breakage whether the inflection from the lateral flange contacted the lateral trough, it certainly did not reach the promontorium, as in multituberculates (Kielan-Jaworowska et al. 1986; Wible and Hopson 1993; Rougier and Wible 2006). It appears that the greater petrosal nerve was exposed to the tympanic cavity instead of coursing within the bone of the petrosal, and as such a hiatus Fallopii is lacking. The vasculature coursing in the lateral trough would likely have communicated with the cranial cavity via the anterior opening of the cavum epiptericum, which in our specimens is represented by a shallow notch at the rostrolateral aspect of the petrosal (ace, Fig. 3a).
A prominent oblique groove is present along the lateral face of the petrosal (Fig. 3e, f), coursing between the petrosal and, as interpreted, the overlying squamosal; this groove housed a large arteria diploëtica magna (Fig. 4b). Dorsally and medially, a notch present at the suture of these elements indicates the opening of the posttemporal canal. This groove continues dorsally and ventrally and would have been occupied by the occipital artery. As the groove approaches the ear region laterally and anteriorly, it bifurcates, with a larger branch coursing dorsally. This groove ends abruptly, most likely indicating that the vessel became fully enclosed within the squamosal. This groove would have held an intramural orbitotemporal artery, bound for the orbital region. The other division of the presumed arteria diploëtica magna passed anteriorly, lateral to the epitympanic recess, as the ramus superior, occupying the ventral ascending groove (McDowell 1958; Wible and Hopson 1993; Rougier et al. 1996). Following the course of the ramus superior from lateral to medial, toward the ear region, a short section of this vessel would have been fully enclosed by the petrosal. Here, the lateral flange has been completely flattened and its lateral portion is visible in ventral view. A large, oval shaped foramen is present and most likely transmitted an extracranial ramus temporalis (though the foramen appears too large for this vessel to have been the only occupant, based on the scale of neighboring vasculature).
Dorsal view.—The dominant features in dorsal (endocranial) view of the petrosal are the internal acoustic meatus centrally and the subarcuate fossa posteriorly, and on the preserved lateral rim of the specimen, the opening of the prootic canal and, anterior to this, the partially preserved semilunar recess (Fig. 3c, d).
The internal acoustic meatus is shallow and dumbbell-shaped, containing two pairs of openings divided by a low but distinct crest, the crista transversa (Fig. 5b). The upper half of the meatus (foramen acousticum superius) is a circular pit with two round openings. The anterior of these is the primary facial foramen and the posterior, which is slightly larger, is the utriculoampullary foramen. The lower portion of the meatus (foramen acousticum inferius) is anteroposteriorly broad. The round, posterior opening is the saccular foramen, while the remainder of the floor in this region is perforated by a strip of numerous small openings continuing anteriorly to a large circular opening. This is the cribriform plate for the cochlear nerve, indicating the presence of a tractus foraminosus for distribution of the bundles of this nerve. Posterior and somewhat dorsal to the internal acoustic meatus is a very deep, spherical excavation; this is the subarcuate fossa. The endocranial margin of the fossa is formed by the anterior semicircular canal, and this deep pocket would have accommodated the cerebellar paraflocculus in life (Kielan-Jaworowska 1986a, b). The posterior semicircular canal protrudes into the wall of the fossa.
Both specimens are broken dorsally and laterally, with crushing of the anterior lamina leaving the structures here poorly delimited. However, a deep elliptical pocket is present anterior to the level of the internal acoustic meatus; this is the semilunar recess (Fig. 3c, d), which would have housed the semilunar ganglion of the trigeminal nerve. A thick, broad ridge of bone, the crista petrosa, separates the anteromedial and anterolateral portions of the petrosal in dorsal view. A thick and blunt prefacial commissure fully separates the semilunar recess from the facial foramen in the internal acoustic meatus and forms a steep medial wall for the semilunar ganglion. The anterior and lateral walls of the semilunar recess are damaged, but on the better-preserved right petrosal there is enough preserved of the anterior lamina to determine that it was a substantial element in the wall of the braincase (Fig. 3e). The anterior lamina is formed by thin laminar bone extending anteriorly from the edges of the subarcuate fossa and prootic sinus and forming at least a partial lateral wall to the cavum epiptericum. Enclosed in the anterior lamina and its thickened base is an enigmatically large foramen for the ramus temporalis (described above) and perhaps part of the foramen ovale, which would have transmitted the mandibular division of the trigeminal nerve. The evidence of a foramen ovale comes exclusively from the right petrosal, which appears to preserve part of the margin of a relatively large, anterolaterally directed opening from the cavum epiptericum through the side wall of the braincase, close to the lateral flange and just dorsal to the level of the lateral trough (Figs. 1b and 3a).
The right petrosal preserves a distinct broad, dorsal projection of bone at the anterior end of the crista petrosa (Figs. 1 and 3c, e). This dorsal projection reflects a general rise of the anterior pole of the promontorium; it is not very pronounced near the medial edge but becomes more so laterally, where it reaches the level of the crista petrosa. This structure can be interpreted as a broken base for a posterior clinoid process, an ossified pila antotica (this region is missing from the left petrosal). The extent of the pila cannot be determined, but lateral to it the petrosal supports a large space that would have been bounded by the crista petrosa/prefacial commissure medially and by the putative lateral flange/anterior lamina laterally. This large space is the cavum epiptericum, which includes the semilunar recess for the trigeminal ganglion at its most posterior extent. In some early mammals and metatherians there is an incomplete walling between the cavum epiptericum and the cavum supracochleare, leaving a common opening, the fenestra semilunaris. Such a connection is absent in our specimens.
At the anteromedial corner of the petrosal, in the region of the contact with the basisphenoid, is a faint groove that wraps around to the medial border of the petrosal but fades into a series of small, irregular openings (Fig. 1d). As discussed above, we interpret that a well-developed inferior petrosal sinus was not developed; perhaps the anterior groove fed into an intrapetrosal sinus network, but a consistent vascular channel joining the jugular notch was absent. The groove for the perilymphatic duct can be seen opening into the jugular notch and curving toward the intracranial surface. A small foramen is positioned dorsal and posterior to the jugular notch, medial to the subarcuate fossa; this is the opening of the aqueductus vestibuli, containing, in life, the endolymphatic duct (Fig. 3c, d). The petrosal morphology does not make it clear whether an endocranial endolymphatic sinus was developed.
A well-defined groove is present tracing the dorsolateral margin of the subarcuate fossa, running along the bony ridge enclosing the anterior semicircular canal. This groove is incomplete and likely developed along the suture between the petrosal and the missing occipitoparietal elements. The most distinct part of the groove starts medially, near the highest point of the subarcuate fossa, and curves laterally and ventrally to pierce the substance of the petrosal, forming a short and vertical canal just posterior to the semilunar recess. This is the prootic canal, which would have transmitted the prootic sinus from the endocranial to the tympanic surface (Wible and Hopson 1995). A less developed portion of the groove extends posteriorly along the posterior portion of the anterior semicircular canal bulge and subarcuate fossa, becoming the sulcus for the sigmoid sinus, which loses identity toward the jugular foramen (Fig. 3). There is no clear evidence for where a transverse sinus would have joined the system of grooves described above, but given that both limbs—one eventually exiting the skull via the prootic canal and the other likely exiting via the foramen magnum—are distinct, it is likely that it was relatively large. The presence of a small vein continuing from the sigmoid sinus into the jugular foramen is likely but subsidiary to the above-mentioned vessels.
Endocast.—The promontorium in both petrosals is well preserved and likely subjected to little if any deformation, resulting in excellent preservation of the endocast of the bony labyrinth (Figs. 5 and 7). Fine details of the inner wall of the cochlear canal are recorded, and the semicircular canals are complete. The cochlear canal occupies most of the volume of the pars cochlearis but does not reach its anterior pole. The most striking feature of the endocast is that the cochlear canal makes one full turn (Table 1), with the apical portion contacting the base. There is no overlap, with the large majority of the canal coiling on the same plane and only the apical portion slightly projected ventrally. This likely accounts for the rounded but less inflated appearance of the promontorium, as compared with therians. While a slight distal constriction can be seen in both specimens, there is no clear indication of the presence of a lagenar expansion or a separate canal or sulcus for a lagenar nerve (Ruf et al. 2013). Deep grooves are preserved on the cochlear endocast, representing well developed primary and secondary bony laminae. Both spiral laminae extend to roughly the same point along the cochlear coiling, to the slight constriction in the cochlear canal near the apex. The secondary lamina is tall basally but gradually reduces in height distally, becoming quite faint. In life, nerve fibers coming from the cribriform plate would have projected though individual perforations of the tractus foraminosus; however, there is no clear evidence of Rosenthal’s canal for the spiral ganglia, which cannot be attributed to poor preservation or limitations in scan resolution. At the base of the cochlear canal, casts of the apertures of the fenestra vestibuli and the perilymphatic foramen are visible. The latter is positioned immediately medial to the impression for the secondary bony lamina. The confluence of the perilymphatic groove (or its reconstructed occupant, the perilymphatic duct) with the foramen is clearly evident (Fig. 7a, b). The proximal portion of the perilymphatic duct wraps around the cochlear endocast from a point at the base of the primary bony lamina, and bears a slight inflation just prior to merging with the perilymphatic foramen.

CT rendering of bony labyrinth endocast of OMNH 80536, right petrosal from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ventral (a), dorsal (b), lateral (c) views. Abbreviations: amp-a, anterior ampulla; amp-h, horizontal ampulla; amp-p, posterior ampulla; av, aqueductus vestibuli; cc, primary common crus; fv, fenestra vestibuli; pd; perilymphatic duct; pf; perilymphatic foramen; pl, primary bony lamina impression; re, recessus ellipticus; rs, recessus sphericus; scc, secondary common crus; sl, secondary bony lamina impression; tf, tractus foraminosus
The two divisions of the vestibule, the recessus ellipticus and recessus sphericus (which housed the membranous utricle and saccule, respectively), form distinct bulges in the endocast (Fig. 7). Casts of the divisions of the vestibular nerve, passing through the utriculoampullary and saccular foramina, are also evident. The cast of the endolymphatic duct extends from the vestibule near the base of the common crus (Fig. 7). The arrangement and proportions of the semicircular canals and their ampullae are very similar to those described for Dryolestes (Luo et al. 2012). The anterior and posterior semicircular canals unite to form a primary common crus, and the horizontal semicircular canal joins the posterior semicircular canal near its ampulla to form a secondary common crus. The horizontal semicircular canal curves around the medial surface of the posterior ampulla to enter the vestibule near the base of the common crus.
Stapes.—Both petrosals preserve fragments of a likely long and gracile bicrurate stapes (Fig. 8), found lodged just inside their respective fenestra vestibuli (Fig. 5). In each, the footplate is incomplete, with a preserved diameter approximately one-half that of the fenestra (Table 1). The vestibular surface of the footplate is slightly convex. The large, conical base of what we interpret as the anterior crus is present in the center of the preserved footplate, and a crest runs along the external surface of the footplate from this crus toward the base of a smaller, poorly preserved root of the posterior crus at the preserved margin. In addition to the footplate, the left stapes is also represented by the knob-shaped (presumably incomplete) stapedial head at the end of a thin partial crus, and a nearby, similarly thin, isolated rod of bone. We interpret this rod to represent part of the posterior crus. The stapedial foramen between the crura is estimated to be approximately 0.22 mm in diameter as measured between the preserved shafts of the crura on the footplate; given the length of the potential stapedial crura the foramen should have been an elongated ellipse, a morphology thought to be primitive for Mammaliaformes (Gaetano and Abdala 2015). There are isolated bone fragments within the cochlear canal near the right stapes, and while one could represent the stapedial head and another appears to be a partial shaft of a crus, they provide no additional information on morphology. Although the stapes cannot be reconstructed with confidence, it is likely that the crura were delicate and parallel between head and footplate, with a morphology similar to that described for the docodontan Borealestes from the Isle of Skye (Panciroli et al. 2018). However, the footplates are incomplete, so it is uncertain if the stapes was symmetrical or asymmetrical.

CT renderings of stapes of OMNH 80536 (a, b), right petrosal, and OMNH 80537 (c, d), left petrosal, both from CMQ (Upper Jurassic Morrison Formation, Grand County, UT, USA), in ?ventral (a, c) and ?external (b, d) views. Abbreviations: asc, anterior stapedial crus; psc, posterior stapedial crus; sh, stapedial head
Comparisons
While the petrosal is a feature-rich and phylogenetically important element for exploring mammal relationships and character evolution, most groups of early mammals were traditionally, and are still, diagnosed based primarily on dental morphology, making taxonomic referral of isolated petrosals challenging (Martin 1999; Rougier et al. 2001; Lopatin et al. 2005; Li and Luo 2006; Bi et al. 2016). Moreover, the CMQ petrosals exhibit a novel combination of features that cannot be easily reconciled with known or expected petrosal morphologies of taxa represented by dentitions from the CMQ. Most striking is the shape of the endocast: the cochlear duct makes a full turn (Fig. 7). The oldest published occurrence of a fully-coiled cochlear duct is in the eutherian Prokennalestes, from the Aptian-Albian of Mongolia (Wible et al. 2001), nearly 40 Ma later. While a fragmentary lower molar of an indeterminate tribosphenidan has been described from the earliest Cretaceous Lakota Formation (Cifelli et al. 2014), no therians are known from the Jurassic of North America. Based solely on the cast of the bony labyrinth the occurrence of these petrosals in the Upper Jurassic Morrison Formation could indicate the early presence of therians or close therian sister taxa, but their morphology differs from what would be expected based on comparisons with cladotherians (Ruf et al. 2009; Luo et al. 2011a) and the stem therian Vincelestes (Rougier et al. 1992; see Discussion below); more importantly, it contradicts overall optimization of character distribution in most recent high-level phylogenetic trees (Luo et al. 2017; Bi et al. 2018; Huttenlocker et al. 2018; Krause et al. 2020). Alternatively and perhaps more likely, these petrosals suggest that a non-therian lineage independently evolved a full coil of the cochlea, as well as other derived features of the ear region, by the Late Jurassic. Partial coiling of the cochlea has so far been recorded in the stem therian Vincelestes from the Early Cretaceous of Argentina (Rougier et al. 1992; Rougier 1993), and in various members of the Dryolestoidea: ~270° in Late Jurassic Dryolestes and Henkelotherium from Guimarota (Ruf et al. 2009; Luo et al. 2012), and more than one full turn in the Late Cretaceous meridiolestidans from South America (Rougier et al. 2009, 2021). The general incompleteness of the fossil record and the overall taxonomic richness of the Late Jurassic—encompassing almost the entirety of mammaliaform lineages—leave a broad array of relevant groups for which these petrosals must be considered.
The small block that yielded the left petrosal contained a few fragmentary, unidentifiable bone fragments; additionally, a small, isolated multituberculate upper premolar (possibly a P4) was discovered within a few centimeters of the left petrosal. Though limited comparative data are available, the size of this tooth is an appropriate fit for the size to the petrosal (using craniodental proportions from Hahn 1988). However, the morphology of the petrosals and endocasts does not conform easily to any taxa known by dentition, including multituberculates. Admixture of elements from different taxa does occur at this site, and as discussed below there are major differences between these petrosals and described morphology of known multituberculates. We therefore prefer to remain cautious and refrain from formally associating the multituberculate premolar with the petrosals. At present, the taxonomic affinities of the petrosals are better considered on the basis of their intrinsic anatomical information, leaving their taxonomic status uncertain until more complete data become available. Below, we compare the CMQ petrosals with known morphology of contemporaneous or near-contemporaneous members of the most likely candidates (Fig. 9).

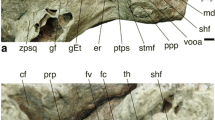
modified from Wible and Hopson 1993) and Dinnetherium (b, modified from Crompton and Luo 1993; Rougier et al. 1996); the docodontan Haldanodon (c, modified from Ruf et al. 2013); the eutriconodontan Priacodon (d, modified from Rougier et al. 1996); the possible gobiconodontid Khoobur 1 (e, modified from Wible and Hopson 1995); the multituberculate ?Catopsalis (f, modified from Wible and Hopson 1993); the meridiolestidan Cronopio (g, modified from Rougier et al. 2011); the stem therian Vincelestes (h, modified from Rougier et al. 1992); the basal eutherian Prokennalestes (i, modified from Wible et al. 2001); the dryolestid Dryolestes (j, modified from Luo et al. 2012); the CMQ petrosal OMNH 80536 (k). Abbreviations: ci, crista interfenestralis; cp, crista parotica; fc, fenestra cochleae; fv, fenestra vestibuli; jn, jugular notch; lt, lateral trough; pc, prootic canal; pp, paroccipital process; pr, promontorium; sff, secondary facial foramen
Right petrosals of Mesozoic taxa with selected features labeled on j; the morganucodontans Morganucodon (a,
Multituberculates.—An isolated multituberculate premolar of appropriate size was found in close association with one of the CMQ petrosals. Because this tooth could potentially be used to refer the CMQ petrosals to a high-level mammalian taxon, multituberculates are perhaps the most important group to consider first. The CMQ petrosals do share a number of generally primitive features with multituberculates, such as an open channel for the perilymphatic duct within the perilymphatic foramen and a horizontal crista interfenestralis (Kielan-Jaworowska et al. 1986; Hurum et al. 1996). However, the rounded promontorium, coiled cochlear canal, and well developed tractus foraminosus for the cochlear nerve differ strongly from known multituberculates.
One of the most distinctive features of the multituberculate petrosal is the medial folding of the lateral flange, contacting the promontorium and enclosing the vasculature that coursed through the lateral trough (Rougier et al. 1992; Wible and Hopson 1995; Wible and Rougier 2000) in a well-delimited posttrigeminal canal (Fig. 9f). The lateral flange is damaged in both CMQ petrosals, but the posterior portion clearly forms an anteroposteriorly-directed canal lateral to the facial foramen that would have enclosed the ramus inferior and, if present, the posttrigeminal vein (Figs. 2d, 4 and 6). This infolding of the lateral flange in the CMQ petrosal is, however, lateral to the position of the facial foramen and therefore unlike multituberculates in that it does not extend medially to the promontorium. The development of a fully enclosed posttrigeminal canal seems to be a condition in Cretaceous cimolodontans, with Jurassic multituberculates (Hahn 1988; Lillegraven and Hahn 1993) possessing a more open facial sulcus. It would be expected that, if the CMQ petrosals were to belong to a multituberculate like Glirodon (Davis et al. 2018), the petrosal morphology would resemble that of their Jurassic European counterparts; the inner ear morphological differences (Lillegraven and Hahn 1993) are dramatic. The condition in the CMQ petrosals could be interpreted as intermediate or incipient, wherein the lateral flange projects medially, enclosing some of the contents of the distal lateral through, but without contacting the promontorium. Overlap to the degree characteristic of multituberculates was not present. Known Jurassic and Cretaceous multituberculate petrosals show a conspicuous fossa anterior to the fenestra vestibuli and medial to the facial foramen, identified as the fossa muscularis major by Kielan-Jaworowska et al. (1986) and Hahn (1988) and as the tensor tympani fossa by Wible and Rougier (2000); the CMQ petrosals lack any indication of a similar structure in the homologous region. The petrosal in Cretaceous and later multituberculates is sunken with respect to the ventral surface of the basicranium, likely to accommodate an enlarged middle ear space possibly mimicking an auditory bulla (e.g., Rougier et al. 2016; Krause et al. 2021), with development of a deep epitympanic recess and jugular fossa. The CMQ petrosals do not show these features. Although the presence of a multituberculate tooth in close proximity to one of the CMQ petrosals requires particular attention to potential multituberculate affinities for the specimens, their overall morphology makes referral of the Morrison petrosals to this group unattractive.
Basal mammaliaforms.—Morganucodontans are well represented in Early Jurassic faunas worldwide, and abundant specimens (including isolated petrosals) of Morganucodon have been recovered from fissure fills in Wales (e.g., Kermack et al. 1981). The morganucodontan Dinnetherium (Jenkins et al. 1983) is known from cranial remains including the petrosal. Isolated teeth from Germany and Britain referred to the group show that it survived until the earliest Cretaceous (Butler et al. 2012; Martin et al. 2019), but well-preserved material from the CMQ (Davis et al. 2018) add a morganucodont to the Morrison fauna. Docodontans are common elements of the Morrison fauna (though perhaps geographically restricted, Foster et al. 2006), but unrepresented by cranial material other than tooth-bearing elements. Docodontans are, however, yet to be documented at CMQ. Haldanodon from Guimarota is closely related to Docodon, is known by much more complete material (Krusat 1980; Lillegraven and Krusat 1991; Ruf et al. 2013), and serves as a good proxy for the expected petrosal morphology for Docodon. While the CMQ petrosals share with basal mammaliaforms exposure of the perilymphatic duct within the perilymphatic foramen and a horizontal crista interfenestralis (Fig. 9a–c), the region posterior to the promontorium is dramatically different in the presence of a well-developed caudal tympanic process and a very tall paroccipital process. Morganucodontans and docodontans retain a robust, primitive jaw articulation where quadrate (incus) and articular (malleus) serve a mechanical function (Kermack et al. 1973; Crompton and Hylander 1986). As such, the short process of the incus is lodged in a deep and well-delimited pit. In contrast, the articulation of the incus with the skull in crown mammals and the CMQ petrosals is represented by a small and shallow pit, the fossa incudis developed lateral to the crista parotica consistent with gracile middle ear ossicles. Moreover, the cochlear canal in basal taxa is short and straight or slightly curved and bears a lagenar expansion. A lagena is absent in the CMQ petrosals and the cochlea makes a full turn; the cochlear nerve is distributed via a tractus foraminosus and both primary and secondary bony laminae are well developed. All these features are absent in morganucodontans and docodontans.
Eutriconodontans.—The eutriconodontan Priacodon is known by a relatively well-preserved petrosal from the Morrison Formation in the nearby Fruita Paleontological Area in Colorado (Rougier et al. 1996; Harper and Rougier 2019; Fig. 9d here). The perilymphatic groove is open to the middle ear cavity lateral to the perilymphatic foramen, but the promontorium is tubular and contains a straight cochlear canal. The lateral wall of the promontorium is steep, lacking a transpromontorial groove, and the lateral trough is open, with vascular structures well exposed in ventral view, differing from the bulbous promontorium and enclosure of the lateral trough in the CMQ petrosals. The petrosal of Priacodon is similar in that it lacks a floor for the cavum supracochleare and the crista interfenestralis is horizontal, interrupting the postpromontorial tympanic sinus and dividing the tympanic cavity into two regions. The internal acoustic meatus is similarly shallow in Priacodon, but there is a single large foramen for the cochlear nerve instead of the cribriform plate present in the CMQ petrosals. A primary bony lamina, well developed in the CMQ petrosals, is absent in Priacodon (Harper and Rougier 2019).
Khoobur petrosals.—Wible et al. (1995) described in detail an isolated petrosal from the Early Cretaceous of Khoobur, Mongolia, mentioning the presence of a second one with moderate differences among them (Rougier et al. 1996). Harper & Rougier (2019) restudied the specimens, including the inner ear anatomy previously inaccessible, within a broader framework of mammalian ear region evolution. The tympanic region morphology of the CMQ and Khoobur petrosal differs strongly (Fig. 9e). The promontorium and the enclosed cochlear endocast are straight, narrow, and accompanied by a steep lateral wall in the Khoobur petrosal, lacking vasculature imprints, a character combination denoting a generalized primitive mammalian morphology; in contrast the Morrison petrosals present a derived condition for all these features. Likewise, the distolateral corner of the Khoobur petrosal appears not to have fully enclosed the arterial branches of the arteria diploëtica magna en route to the tympanic aspect of the middle ear, resulting in a broad and exposed merging of the postpromontorial tympanic sinus and lateral trough. In contrast, the CMQ petrosals have a better developed lateral flange/crista parotica area enclosing, to various degrees, the vascular canals of the ramus superior, ramus inferior, stapedial artery and arteria diploëtica magna.
Not all features are more plesiomorphic in the Khoobur petrosal than in the Morrison specimens. The former has a fully developed postpromontorial tympanic sinus, true fenestra cochleae with complete processus recessus, vertical crista interfenestralis, and a distinct inferior petrosal sinus canal; these presumably derived features shared with advanced stem therians are lacking in the CMQ petrosals. The prominent paroccipital process projects ventrally from the level of the promontorium, is hook-shaped, and projects anteriorly; this feature is shared between the Khoobur and CMQ petrosals and to some extent also with Ornithorhynchus (Wible and Hopson 1993; Wible et al. 1995). The significance of this trait is, however, uncertain.
The systematic attribution of the Khoobur petrosal is not fully settled. Wible et al. (1995) argued for either gobiconodontid or obtuse-angled “symmetrodont” affinities, leaning in favor of a “symmetrodont” affiliation. Harper & Rougier (2019) considered both options viable, but leaned toward gobiconodontid relationships based in part on the presence of a laterally thickened area in the petrosal, likely involved in glenoid buttressing if not in direct articulation with the robust gobiconodontid mandibular condyle. Differences are so pronounced between the Khoobur and CMQ petrosals that non-plesiomorphic similarities are likely to be convergently derived.
“Symmetrodonts”.—Obtuse-angled “symmetrodonts” such as Tinodon are known from the Morrison Formation by tooth-bearing elements only (Simpson 1929). Unnamed derived spalacotheriid “symmetrodonts” have been described from Fruita and Dinosaur National Monument (Engelmann and Callison 1998), and are also known from slightly younger units in Britain and South Dakota (Owen 1871; Ensom and Sigogneau-Russell 2000; Cifelli et al. 2014). Among spalacotherioids, the ear region is only known in the Early Cretaceous zhangheotheriids (Hu et al. 1997; Rougier et al. 2003; Luo et al. 2016), a group which is unknown from North America. Both zhangheotheriids and the CMQ petrosals are plesiomorphic in having an exposed perilymphatic duct within the perilymphatic foramen, but otherwise the ear regions are dissimilar. In Zhangheotherium, the promontorium is very slightly inflated but straight, containing a straight cochlear canal (Luo et al. 2016). The crista interfenestralis is vertical and the postpromontorial tympanic sinus is complete. The cavum supracochleare for the geniculate ganglion is floored, and the lateral trough lacks any enclosure. A secondary bony lamina is described in Zhangheotherium, but the primary lamina is absent (Luo et al. 2016).
Dryolestids and meridiolestidans.—In the broadest sense, three groups can be considered as potential members of the Dryolestoidea: the Dryolestidae, Paurodontidae, and Meridiolestida (Butler 1939; Prothero 1981; Bonaparte 1992; Martin 1999; Rougier et al. 2011, 2012, 2021a, b; Averianov et al. 2013; Averianov and Martin 2015). The interrelationships among these three distinct groups are poorly resolved and alternatives range from all three forming a single monophyletic group to each one of them being successive sister groups to each other as stem lineages to Theria (Rougier et al. 2011, 2012; Martin et al. 2021). Additionally, the Paurodontidae, a common component of Late Jurassic faunas and traditionally best represented by Henkelotherium from Guimarota (Krebs 1991; Jäger et al. 2020), may not be a monophyletic group based on recent analyses (e.g., Martin et al. 2021). The petrosal morphology of Mesozoic dryolestoids is represented by two Late Jurassic specimens from Guimarota (Ruf et al. 2009; Luo et al. 2012) and two members of the endemic South American meridiolestidan radiation from the Late Cretaceous of Argentina (Rougier et al. 2009, 2021). The overall petrosal morphology is similar within this group, with the major exceptions being: 1) the Jurassic taxa have a partially-coiled cochlear canal (~ 270°), whereas it is fully coiled in meridiolestidans; and 2) meridiolestidans have deeper imprints of cranial vasculature. Aside from these points of similarity, dryolestoid petrosals differ from the CMQ petrosals in several features: the fenestra cochleae and perilymphatic duct are separated by a full processus recessus; the postpromontorial tympanic sinus is continuous posterior to a vertical crista interfenestralis; the cavum supracochleare is floored; and the lateral trough lacks a fully enclosed canal, though in the meridiolestidan Cronopio (Rougier et al. 2011) there is at least a partial floor for the ramus superior (Fig. 9g, j). Additionally, a well-developed inferior petrosal sinus canal is present along the petrosal-basisphenoid suture in the meridiolestidans, but not in the CMQ petrosals, which also have a less prominent and differentiated promontorium than the SA taxa.
Vincelestes.—The cranial anatomy of Vincelestes is relatively well known (Bonaparte and Rougier 1987; Hopson and Rougier 1993; Rougier et al. 2021a), with the petrosal/braincase having been described in detail (Rougier et al. 1992). Most phylogenies recover Vincelestes as a stem therian closer to Theria than basal cladotheres like dryolestoids and meridiolestidans, but alternative views exist (Bonaparte 2008; Averianov et al. 2013; Bonaparte and Migale 2015). The petrosal of Vincelestes shows derived features shared by dryolestoids, meridiolestidans, and therians but absent in the CMQ petrosals. Chief among them are the features related to the crista interfenestralis and the postpromontorial tympanic sinus, which is a continuous space in cladotheres, but not in the CMQ petrosals. Here, the space is divided by the horizontal crista interfenestralis (a primitive condition) into two separate realms. Unlike cladotheres, the CMQ petrosals lack a true fenestra cochleae and a processus recessus and therefore the perilymphatic duct was exposed to the middle ear cavity but lodged in a distinct groove leading from the perilymphatic foramen to the jugular foramen. Unlike Vincelestes, Dryolestes, and therians the CMQ petrosals show a prominent and compressed paroccipital process that extends far below (ventral to) the level of the promontorium, a feature present in monotremes, at least some multituberculates, and in one of the taxa represented by the Khoobur petrosals. The petrosals we describe here share with Vincelestes, meridiolestidans, and dryolestoids (but not basal mammaliaforms) a relatively inflated promontorium with prominent vascular groves for a transpromontorial internal carotid artery and stapedial system (Fig. 9h). In Vincelestes and the CMQ petrosals, the separation of the tympanic-bound branches of the arteria diploëtica magna (ramus superior, ramus inferior, and stapedial artery) is enclosed in a medially-folded distal portion of the lateral flange, which in both taxa is pierced by temporal rami traveling through the substance of the lateral flange toward the temporal fenestra.
Vincelestes and the CMQ petrosals differ strongly in the morphology of the internal acoustic meatus. This structure is very deep and tubular in the former, but in the CMQ petrosals it is shallow, with exposure of the foramina of a cribriform plate for entry of branches of cranial nerves VII and VIII into the petrosal—the condition regarded as primitive and retained in therians. The osseous labyrinth offers a sharp contrast between the CMQ petrosals and Vincelestes. Vincelestes shows a “candy cane” cochlear endocast (Rougier 1993; Rougier et al. 2021) where most of the cochlea is straight or slightly curved, with the tip sharply turned and forming an angle of approximately 270°. The CMQ petrosals, instead resembling meridiolestidans (Rougier et al. 2009, 2021; Wible and Rougier 2017), show a complete turn in addition to extensive primary and secondary bony laminae, with the concomitant derived arrangement of cochlear innervation.
Therians.—The oldest described petrosal referred to a therian is assigned to the basal eutherian Prokennalestes, from the Aptian-Albian of Mongolia (Wible et al. 2001). A few features are shared with the CMQ petrosals, such as a full turn of the cochlear canal and presence of a transpromontorial groove, but the petrosals otherwise differ substantially (Fig. 9i). In Prokennalestes, the postpromontorial tympanic sinus is broad and continuous, the cochlear canal is fully separate from the fenestra cochleae, and the hiatus Fallopii (the separate opening for the greater petrosal branch of the facial nerve) is well anterior to the facial foramen, indicating a long intrapetrosal course for this branch and an extensive floor for the cavum supracochleare. Additionally, the lateral trough and lateral flange in the CMQ petrosals are well developed; though a strongly reduced anterior lamina is described in Prokennalestes, the anterior lamina and lateral flange are generally considered absent in therians (Rougier and Wible 2006).
Discussion
Among the derived features of the petrosals from CMQ, perhaps the most striking is the fact that the cochlear canal makes one full turn (Fig. 7). This is a substantially greater degree of coiling than any contemporaneous mammal. Docodontans (Panciroli et al. 2018), eutriconodontans (Rougier et al. 1996; Harper and Rougier 2019), and paulchoffatiid multituberculates (Lillegraven and Hahn 1993) are characterized by a relatively short, straight cochlea, while the cochlea in Dryolestes, a taxon known from both the Morrison Formation and from Guimarota in Portugal (Luo et al. 2012), curves to 270°. Otherwise, a full coil of the cochlear canal is not recorded until the basal eutherian Prokennalestes from the Aptian-Albian of Mongolia. Older therians such as Juramaia have been described (Luo et al. 2011b), and while therian petrosals older than Aptian-Albian are unknown, the character distribution from recent analyses (Bi et al. 2018; Krause et al. 2020) implies that the petrosal and cochlea of Jurassic therians would not be expected to differ substantially from that of Prokennalestes. Importantly, among non-therians, a fully coiled cochlea is known only in meridiolestidan dryolestoids from the Cretaceous of Argentina. Petrosals of Campanian age have been described for Coloniatherium (Rougier et al. 2009), and while the ear region is not yet fully described, well-preserved cranial material of Cronopio extends the record of this morphology to the Cenomanian (Rougier et al. 2011). Recent phylogenetic analyses recover meridiolestidans and Laurasian dryolestids and paurodontids as basal members of the early cladothere radiation (Rougier et al. 2012; Bi et al. 2018; Krause et al. 2020), implying a minimal Jurassic age for all the groups involved. The Jurassic-Early Cretaceous ghost lineage evidenced by the gap in the record of meridiolestidans (Rougier et al. 2021) makes the timing of the origin of a coiled cochlea uncertain for the South American clade within that broad time span.
Ultimately, the CMQ petrosals display conflicting data. The presence of an open perilymphatic duct, tall and horizontal crista interfenestralis, sizeable prootic canal, and ossified pila antotica (Fig. 3) all suggest that this taxon is well removed from cladotheres. However, the cochlear canal makes a full turn and bears an advanced support and distribution system for the cochlear nerve via a complex tractus foraminosus and extensive primary and secondary bony laminae (Figs. 4b and 7). The proximity of an isolated multituberculate premolar cannot be ignored, especially in light of the folding of the lateral flange to partially enclose vasculature and perhaps a small portion of the lateral trough, a feature that can be seen as vaguely similar to the much more extensive medial projection of the lateral flange distinctive of multituberculates. However, no member of this group shows anything resembling the cochlea in the CMQ petrosals, and the morphological details of the lateral trough, facial foramen, and promontorium fail to recover specific similarities between the CMQ and known Jurassic and Cretaceous multituberculate petrosals. The overall evidence leads us to not support multituberculate affinities for the CMQ petrosals. This leaves a number of possibilities regarding the affinities of the CMQ taxon.
First, it could be a Late Jurassic member of a group with known cochlear coiling, such as the meridiolestidans, otherwise known only from South America. The cochlea would be precocious relative to other features of the petrosal, which retain plesiomorphies. However, petrosals of meridiolestidans more closely resemble Dryolestes and Henkelotherium in external morphology than the CMQ petrosals, and these taxa are recovered as closely related to the base of the radiation (Rougier et al. 2012). It is worth pointing out, however, that paurodontids, well known by dentitions from the Morrison Formation (Simpson 1929; Prothero 1981; Bakker and Carpenter 1990), are as yet unknown by fossils preserving the petrosal (Henkelotherium was recovered as a dryolestid in Rougier et al. 2012 and as a dryolestoid by Averianov et al. 2013). Paurodontids appear to be close relatives of meridiolestidans, either contained within a monophyletic group that also includes dryolestids (Rougier et al. 2011) or as therian stem groups (Rougier et al. 2012, 2021b). An additional supported possibility is that paurodontids are paraphyletic stem members of the dryolestoid radiation (Martin et al. 2021), and some of these taxa could have their own trajectory with regard to the ear region. Regardless of the position of the paurodontids among other cladotheres, a petrosal morphology like that in the CMQ would be unexpected and ultimately unlikely. A basal position among cladotheres for Paurodon and relatives (see opposite view in Averianov et al. 2013) would be compatible regarding the morphology of the middle ear region, but the inner ear anatomy would have to be convergent with that of later therians and possibly meridiolestidans. Clustering paurodontids with meridiolestidans reduces inner ear convergences but introduces middle ear character conflict; a substantive amount of homoplastic character evolution would be unavoidable if the CMQ petrosals belong to a paurodontid or any of the other basal cladothere taxa, such as dryolestids and meridiolestidans.
Other taxa with a coiled or partially coiled cochlea include therians and their close relatives. Stem zatherians (Martin 2002), non-therian zatherians (Rougier et al. 1992), and Theria remain unrepresented in the Jurassic of North America. However, they have been recovered from the Late Jurassic to earliest Cretaceous of Western Europe (Simpson 1928; Sigogneau-Russell et al. 2001), and both a stem zatherian and an unidentified tribosphenic mammal have been reported from the earliest Cretaceous of South Dakota (Cifelli et al. 2014). Representation of one or all of these varieties in the Morrison remains a strong possibility. Given the overall primitive ethos of the tympanic region morphology of the CMQ petrosals, identification of them as therians or zatherians would introduce a large number of convergences conflicting with our current view of petrosal surface anatomy and concomitant vasculature and innervation evolution.
Finally, there is the possibility that the CMQ taxon is a multituberculate. The general anatomy of the CMQ petrosals shows little resemblance with that of known Jurassic and Cretaceous multituberculates, although the transmission of vasculature through the substance of the lateral flange and its partial medial projection could be seen as a precursor of the multituberculate condition. However, there is some support for this, as discussed above, but throughout the long, well-sampled record of multituberculates there is no evidence for any coiling of the cochlea or its innervation that could serve as an intermediate between the straight cochlea typical of the group and the full turn, complete with differentiated cochlear nerve canals and long spiral laminae, seen in the CMQ petrosals.
Conclusions
While we remain unsure of the taxonomic affinities of the CMQ petrosals, the implications of this combination of conflicting morphologies are tantalizing. It is certainly feasible that these specimens represent a completely unknown group of mammals, one for which not only the basicranial morphology was hitherto unrecorded, but whose dentition (if it even possessed one) remains to be found. However, given the deep history of work in the Morrison Formation—the unit that provided the first Jurassic mammals from North America (Marsh 1878) and sampled over a span of 140 years—it is most likely that these isolated petrosals belong to a group already known from the Morrison. Among the large collection of mammals recovered to date from the CMQ, most appear to be referable, at least at a high level, to known groups. The most striking and unusual taxon is Fruitafossor, originally known by relatively complete material from the Fruita Paleo Area (Luo and Wible 2005) but now also from the CMQ (Davis et al. 2018). Unpublished data show unequivocally that the ear region of Fruitafossor is radically different from the petrosals described here. This leaves two most likely scenarios: (1) the CMQ petrosals represent an early record of cochlear coiling and derived innervation for a group in which these features are already known (e.g., therians or meridiolestidans), but indicating that the external petrosal morphology is much more primitive than expected; or (2) the CMQ petrosals belong to a group for which cochlear coiling is unrecorded (e.g., multituberculates, eutriconodontans, dryolestids, paurodontids), either representing a lineage that independently derived this feature and its associated innervation pattern, or from which the petrosal is simply yet unknown.
Depending on the position of paurodontids among basal cladotheres, they are a viable possibility for scenarios #1 or #2. If paurodontids are monophyletic with meridiolestidans they could represent basal stock from which meridiolestidans derive their advanced morphology, with dryolestids as intermediate in these features. However, this implies that meridiolestidans evolved from basal dryolestoids that lost primitive features (such as an open perilymphatic duct and horizontal crista interfenestralis) to converge on a general petrosal morphology that is already widespread in other cladotheres. The paurodontid ear region is yet unknown and therefore also fulfills the criterion for scenario #2; however, none of the potential phylogenetic resolutions for them is morphologically congruent with known dryolestid or meridiolestidan ear regions or with the pattern expected to be ancestral to their last common ancestor. Character conflict generated by the non-canonical morphology of the CMQ petrosals is inexorable. Depending on the ultimate taxonomic affinities of the CMQ specimens, this may represent yet another iteration of convergent evolution of cochlear elongation and inner ear coiling in the evolution of Mesozoic mammals.
The CMQ petrosals record for the first time the co-occurrence of a coiled cochlea and a perilymphatic foramen/perilymphatic duct open to the middle ear cavity. Up to now a coiled cochlea has been associated with a relatively derived petrosal morphology which has included a fully ossified processus recessus closing off the duct from the tympanic cavity and forming a neomorphic portion of the fenestra cochleae (DeBeer 1937; Zeller 1985a, b). The reverse has been broadly documented in mammals with a straight or curved cochlea, from partial enclosure of the perilymphatic duct in multituberculates and Priacodon (Rougier et al. 1996; Rougier and Wible 2006) to complete enclosure in dryolestids, Vincelestes, and the Khoobur petrosals (Rougier et al. 1992; Ruf et al. 2009; Luo et al. 2012; Hughes et al. 2015; Harper and Rougier 2019), forming an aqueductus cochleae. Character mapping has long suggested that the formation of a processus recessus long predated the dramatic elongation of the cochlea and its curvature. The CMQ petrosals support a larger degree of plasticity in the interaction of the inner ear and the tympanic cavity than previously documented. It is at present unclear to us the functional implications of a bony enclosure of the perilymphatic duct and its role in auditory performance in early mammals.
References
Alexander G (1904) Entwicklung und Bau des innerens Gehörorgans von Echidna aculeata. Semons Zoologische Forschungsreisen in Australien 3:1–118
Allin EF and Hopson JA (1992) Evolution of the auditory system in Synapsida ("mammal-like reptiles" and primitive mammals) as seen in the fossil record. In: DB Webster, RR Fay, and AN Popper (eds), The Evolutionary Biology of Hearing, 587–614. Springer-Verlag, Inc., New York
Averianov AO, Martin T (2015) Ontogeny and taxonomy of Paurodon valens (Mammalia, Cladotheria) from the Upper Jurassic Morrison Formation of USA. Proc ZIN 319:326–340
Averianov AO, Lopatin AV, Martin T (2013) A new phylogeny for basal Trechnotheria and Cladotheria and affinities of South American endemic Late Cretaceous mammals. Naturwissenschaften 100:311–326. https://doi.org/10.1007/s00114-013-1028-3
Bakker RT, Carpenter K (1990) A new latest Jurassic vertebrate fauna from the highest levels of the Morrison Formation at Como Bluff, Wyoming, with comments on Morrison biochronology. Part III. The mammals: a new multituberculate and a new paurodont. Hunteria 2:2–8
Bast TH and Anson BJ (1949) The Temporal Bone and the Ear. Charles C. Thomas, Springfield
Bi S, Davis B, Meng J, Robinson N, Wang X, Zheng X (2016) A new symmetrodont mammal (Trechnotheria: Zhangheotheriidae) from the Early Cretaceous of China and trechnotherian character evolution. Sci Rep 6:26668. https://doi.org/10.1038/srep26668
Bi S, Cignetti NE, Wang X, Wible JR, Yang S, Zheng X (2018) An Early Cretaceous eutherian and the placental–marsupial dichotomy. Nature 558:390–395. https://doi.org/10.1038/s41586-018-0210-3
Bonaparte JF (1992) Una nueva especie de Triconodonta (Mammalia) de la Formación Los Alamitos, Provincia de Río Negro y comentarios sobre su fauna de mamíferos. Ameghiniana 29:99–110
Bonaparte JF (2008) On the phylogenetic relationships of Vincelestes neuquenianus. Hist Biol 20:81–86
Bonaparte JF, Rougier GW (1987) Mamíferos del Cretácico Inferior de Patagonia. IV Congreso Latinamericano de Paleontología 1:343–359
Bonaparte JF, Migale LA (2015) Protomamíferos y mamíferos Mesozoicos de América del Sur. Fundación de Historia Natural Felix de Azara, Buenos Aires
Butler PM (1939) The teeth of the Jurassic mammals. Proc Zool Soc Lond 109:329–356
Butler PM, Ensom PC, Sigogneau-Russell D (2012) Possible persistence of the morganucodontans in the Lower Cretaceous Purbeck Limestone Group (Dorset, England). Cret Res 33:135–145. https://doi.org/10.1016/j.cretres.2011.09.007
Callison G (1987) Fruita: a place for wee fossils. In: Averett WR (ed) Paleontology and Geology of the Dinosaur Triangle. Museum of Western Colorado, Grand Junction, pp 91–96
Cifelli RL, Davis BM, Sames B (2014) Earliest Cretaceous mammals from the western United States. Acta Palaeontol Pol 59:31–52
Crompton AW, Luo Z-X (1993) Relationships of the Liassic mammals Sinoconodon, Morganucodon, and Dinnetherium. In: Szalay FS, Novacek MJ, McKenna MC (eds) Mammal Phylogeny, Volume 2—Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials. Springer-Verlag, Inc., New York, pp 30–44
Crompton AW, Hylander WL (1986) Changes in mandibular function following the acquisition of a dentary-squamosal joint. In: Hotton N, III, MacLean PD, Roth JJ, Roth EC (eds) The Ecology and Biology of Mammal-like Reptiles. Smithsonian Institution Press, Washington, D. C., pp 263–282
Crompton AW, Bhullar B-AS, Miyamae JA, Musinsky C, Rougier GW (2018) Origin of the lateral wall of the mammalian skull: fossils, monotremes and therians revisited. J Mammal Evol 25:301–313. https://doi.org/10.1007/s10914-017-9388-7
Davis BM, Cifelli RL, Rougier GW (2018) A preliminary report of the fossil mammals from a new microvertebrate locality in the Upper Jurassic Morrison Formation, Grand County, Utah. Geol Intermountain West 5:1–8
DeBeer GR (1937) The Development of the Vertebrate Skull. Clarendon Press, Oxford
DeBeer GR (1929) IX. The development of the skull of the shrew. Philos Trans R Soc B Biol 326, 217:411–480. https://doi.org/10.1098/rstb.1929.0009
Denker A (1901) Zur Anatomie des Gehörorgans der Monotremata. Semon Zool. Forschungsreisen in Australien 3:635–662
Ekdale EG (2016) Form and function of the mammalian inner ear. J Anat 228:324–337. https://doi.org/10.1111/joa.12308
Engelmann GF, Callison G (1998) Mammalian faunas of the Morrison Formation. Mod Geol 23:343–379
Ensom PC, Sigogneau-Russell D (2000) New symmetrodonts (Mammalia, Theria) from the Purbeck Limestone Group, Early Cretaceous of southern England. Cret Res 21:767–779
Foster JR, Madsen SK, Martin JE, Trujillo KC (2006) The Late Jurassic mammal Docodon, from the Morrison Formation of the Black Hills, Wyoming: implications for abundance and biogeography of the genus. New Mexico Mus Nat Hist Sci Bull 36:165–169
Fourie S (1974) The cranial morphology of Thrinaxodon liorhinus Seeley. Ann South African Mus 65:337–400
Gaetano LC, Abdala F (2015) The stapes of gomphodont cynodonts: Insights into the middle ear structure of non-mammaliaform cynodonts. PLoS ONE 10:e0131174. https://doi.org/10.1371/journal.pone.0131174
Hahn G (1988) Die Ohr-Region der Paulchoffatiidae (Multituberculata, Ober-Jura). Palaeovertebrata 18:155–185
Harper T, Rougier GW (2019) Petrosal morphology and cochlear function in Mesozoic stem therians. PLoS ONE 14:e0209457. https://doi.org/10.1371/journal.pone.0209457
Hopson JA and Barghusen H (1986) An analysis of therapsid relationships. In: N. Hotton, III, P.D. MacLean, J.J. Roth, and E.C. Roth (eds), The Ecology and Biology of Mammal-like Reptiles, 83–106. Smithsonian Institution Press, Washington, DC
Hopson JA, Rougier GW (1993) Braincase structure in the oldest known skull of a therian mammal: implications for mammalian systematics and cranial evolution. Am J Sci 293:268–299
Hu Y-M, Li C-K, Luo Z-X, Wang Y-Q (1997) A new symmetrodont mammal from China and its implications for mammalian evolution. Nature 390:137–142
Hughes EM, Luo Z-X, Spaulding M, Wible JR, (2015) Mammalian petrosal from the Upper Jurassic Morrison Formation of Fruita, Colorado. Ann Carnegie Mus 83:1–17
Hurum JH, Kielan-Jaworowska Z, Presley R (1996) The middle ear in multituberculate mammals. Acta Palaeontol Pol 41:253–275
Huttenlocker AK, Grossnickle DM, Kirkland JI, Luo Z-X, Schultz JA (2018) Late-surviving stem mammal links the lowermost Cretaceous of North America and Gondwana. Nature. https://doi.org/10.1038/s41586-018-0126-y
Jäger KRK, Luo Z-X, Martin T (2020) Postcranial skeleton of Henkelotherium guimarotae (Cladotheria, Mammalia) and locomotor adaptation. J Mammal Evol 27:349–372. https://doi.org/10.1007/s10914-018-09457-2
Jenkins FA, Jr., Crompton AW, Downs WR (1983) Mesozoic mammals from Arizona: new evidence on mammalian evolution. Science 222:1233–1235
Kemp TS (1983) The relationships of mammals. Zool J Linn Soc 77:353–384
Kemp TS (2009) The endocranial cavity of a nonmammalian eucynodont, Chiniquodon theotenicus, and its implications for the origin of the mammalian brain. J Vertebr Paleontol 29:1188–1198. https://doi.org/10.1671/039.029.0430
Kemp TS (2016) Non-mammalian synapsids: the beginning of the mammal line. Springer Handbook Aud Res 59:107–137
Kermack KA, Mussett F, Rigney HW (1973) The lower jaw of Morganucodon. J Linn Soc 53:87–175
Kermack KA, Mussett F, Rigney HW (1981) The skull of Morganucodon. Zool J Linn Soc 71:1–158
Kielan-Jaworowska Z (1986a) Brain evolution in Mesozoic mammals. Contributions to Geology, University of Wyoming, Special Paper 3:21–34
Kielan-Jaworowska Z (1986b) Endocranial cast of the Cretaceous eutherian mammal Barunlestes. Acta Palaeontol Pol 31:137–144
Kielan-Jaworowska Z, Poplin C, Presley R, (1986) The cranial vascular system in taeniolabidoid multituberculate mammals. Philos Trans R Soc B Biol 313:525–602
Kielan-Jaworowska Z, Cifelli RL, Luo Z-X (2004) Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. Columbia University Press, New York
Kirkland JI (2006) Fruita Palaeontological Area (Upper Jurassic, Morrison Formation), western Colorado: an example of terrestrial taphofacies analysis. New Mexico Mus Nat Hist Sci Bull 36:67–95
Koyabu D, Endo H, Forasiepi AM, Kimura J, Morimoto N, Ohdachi SD, Sánchez-Villagra MR, Truong Son N, Werneburg I, Zollikofer CPE (2014) Mammalian skull heterochrony reveals modular evolution and a link between cranial development and brain size. Nat Commun 5:3625. https://doi.org/10.1038/ncomms4625
Krause DW, Evans AR, Groenke JR, Hoffmann S, Hu Y, Kirk EC, Rahantarisoa LJ, Rogers RR, Rossie JB, Rougier GW, Schultz JA, von Koenigswald W, Wible JR (2020) Skeleton of a Cretaceous mammal from Madagascar reflects long-term insularity. Nature. https://doi.org/10.1038/s41586-020-2234-8
Krause DW, Hoffman S, Lyson TR, Dougan LG, Petermann H, Tecza A, Chester SGB, Miller IM (2021) New skull material of Taeniolabis taoensis (Multituberculata, Taeniolabididae) from the early Paleocene (Danian) of the Denver Basin, Colorado. J Mammal Evol. https://doi.org/10.1007/s10914-021-09584-3
Krebs B (1991) Das Skelett von Henkelotherium guimarotae gen. et sp. nov. (Eupantotheria, Mammalia) aus dem Oberen Jura von Portugal. Berliner geowiss Abh 133:1–110
Krusat G (1980) Contribução para o conhecimento da fauna do Kimeridgiano da mina de lignito Guimarota (Leiria, Portugal). IV Parte. Haldanodon exspectatus Kuhne & Krusat 1972 (Mammalia, Docodonta). Mem. Serv Geol De Port 27:1–79
Ladevèze S, Muizon C de (2010) Evidence of early evolution of Australidelphia (Metatheria, Mammalia) in South America: phylogenetic relationships of the metatherians from the late Palaeocene of Itaboraí (Brazil) based on teeth and petrosal bones. Zool J Linn Soc 159:746–784. https://doi.org/10.1111/j.1096-3642.2009.00577.x
Li G, Luo Z-X (2006) A Cretaceous symmetrodont therian with some monotreme-like postcranial features. Nature 439:195–200
Lillegraven JA, Krusat G (1991) Cranio-mandibular anatomy of Haldanodon exspectatus (Docodonta; Mammalia) from the Late Jurassic of Portugal and its implications to the evolution of mammalian characters. Contri to Geol, UW 28:39–138
Lillegraven JA, Hahn G (1993) Evolutionary analysis of the middle and inner ear of Late Jurassic multituberculates. J Mammal Evol 1:47–74
Lopatin AV, Averianov AO, Leshchinskiy SV Maschenko EN, Reszvyi AS, Skutchas PP (2005) Early Cretaceous mammals from western Siberia. 1. Tinodontidae. Paleontologiceskij Zurnal 39:523–534
Luo Z-X (2001) Inner ear and its bony housing in tritylodonts and implications for evolution of mammalian ear. In: Jenkins FA, Jr., Owerkowicz T, Shapiro MD (eds) Symposium in Honor of Professor Alfred W. Crompton. Bull Mus Comp Zool 155:621–637
Luo Z-X, Wible JR (2005) A Late Jurassic digging mammal and early mammalian diversification. Science 308:103–107
Luo Z-X, Ji Q, Yuan C-X (2011a) A Jurassic eutherian mammal and divergence of marsupials and placentals. Nature 476:442–445
Luo Z-X, Martin T, Ruf I, Schultz JA (2011b) Fossil evidence on evolution of inner ear cochlea in Jurassic mammals. Proc Royal Soc London 278:28–34
Luo Z-X, Martin T, Ruf I (2012) The petrosal and inner ear of the Late Jurassic cladotherian mammal Dryolestes leiriensis and implications for ear evolution in therian mammals. Zool J Linn Soc 166:433–463. https://doi.org/10.1111/j.1096-3642.2012.00852.x
Luo Z-X, Ekdale EG, Schultz JA (2016) Evolution of the middle and inner ears of mammaliaforms: the approach to mammals. Springer Handbook Aud Res 59:139–174. https://doi.org/10.1007/978-3-319-46661-3_6
Luo Z-X, Grossnickle DM, Ji Q, Liu D, Meng Q-J, Neander AI, Zhang Y-G (2017) New evidence for mammaliaform ear evolution and feeding adaptation in a Jurassic ecosystem. Nature 548:326–329. https://doi.org/10.1038/nature23483
MacPhee RDE (1981) Auditory regions of primates and eutherian insectivores: morphology, ontogeny and character analysis. Contrib Primat 18:1–282
Marsh OC (1877a) Notice of new dinosaurian reptiles from the Jurassic Formation. Am J Sci 14:514–516
Marsh OC (1877b) Notice of a new and gigantic dinosaur. Am J Sci 14:87–88
Marsh OC (1878) Fossil mammal from the Jurassic of the Rocky Mountains. Am J Sci 15:459
Marshall LG, Muizon C, de (1995) Part II: The skull. In: Muizon C, de (ed) Pucadelphys andinus (Marsupialia, Mammalia) from the early Paleocene of Bolivia. Mémoires du Muséum National d'Histoire Naturelle 165, Paris, pp 21–90
Martin T (1999) Dryolestidae (Dryolestoidea, Mammalia) aus dem Oberen Jura von Portugal. Abh der Senck Naturf Gesell 550:1–119
Martin T (2002) New stem-line representatives of Zatheria (Mammalia) from the Late Jurassic of Portugal. J Vertebr Paleontol 22:332–348
Martin T, Averianov A, Jäger KRK, Schwermann AH, Wings O (2019) A large morganucodontan mammaliaform from the Late Jurassic of Germany. Foss Impr 75:504–509
Martin T, Schultz J, Schwermann A, Wings O (2021) A derived dryolestid mammal indicates possible insular endemism in the Late Jurassic of Germany Sci Nat 108. https://doi.org/10.1007/s00114-021-01719-z
McDowell SB (1958) The Greater Antillean insectivores. Bull Am Mus Nat Hist 115:113–214
Meng J, Fox RC (1995a) Osseous inner ear structures and hearing in early marsupials and placentals. Zool J Linn Soc 115:47–71
Meng J, Fox RC (1995b) Therian petrosals from the Oldman and Milk River formations (Late Cretaceous), Alberta, Canada. J Vertebr Paleontol 15:122–130
Meng J and Hou S (2016) Earliest known mammalian stapes from an Early Cretaceous eutriconodontan mammal and implications for transformation of mammalian middle ear. Palaeontol Pol 67:181–196
Owen R (1871) Monograph of the fossil Mammalia of the Mesozoic formations. Monograph of the Palaeontological Soc 33:1–115
Panciroli E, Luo Z-X, Schultz JA (2018) Morphology of the petrosal and stapes of Borealestes (Mammaliaformes, Docodonta) from the Middle Jurassic of Skye, Scotland. Pap Palaeontol. 5:139–156.https://doi.org/10.1002/spp2.1233
Pfaff C, Martin T, Ruf I (2015) Bony labyrinth morphometry indicates locomotor adaptations in the squirrel-related clade (Rodentia, Mammalia). Proc Royal Soc Lond B Biol 282
Presley R (1993) Development and phylogenetic features of the middle ear. In: Szalay FS, Novacek MJ, McKenna MC (eds) Mammal Phylogeny: Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials. Springer-Verlag, New York, pp 21–29
Prothero DR (1981) New Jurassic mammals from Como Bluff, Wyoming, and the interrelationships of non-tribosphenic Theria. Bull Am Mus Nat Hist 167:277–326
Prothero DR (1983) The oldest mammalian petrosals from North America. J Paleontol 57:1040–1046
Rougier GW (1993) Vincelestes neuquenianus Bonaparte (Mammalia, Theria) un Primitivo Mamífero del Cretácico Inferior de la Cuenca Neuquina. Dissertation, Universidad Nacional de Buenos Aires
Rougier GW, Wible JR (2006) Major changes in the ear region and basicranium of early mammals. In: Carrano MT, Gaudin TJ, Blob RW, Wible JR (eds) Amniote Paleobiology: Phylogenetic and Functional Perspectives on the Evolution of Mammals, Birds, and Reptiles. University of Chicago Press, Chicago, pp 269–311
Rougier GW, Hopson JA, Wible JR (1992) Reconstruction of the cranial vessels in the Early Cretaceous mammal Vincelestes neuquenianus: implications for the evolution of the mammalian cranial system. J Vertebr Paleontol 12:188–216
Rougier GW, Hopson JA, Wible JR (1996) Basicranial anatomy of Priacodon fruitaensis (Triconodontidae, Mammalia) from the Late Jurassic of Colorado, and a reappraisal of mammaliaform interrelationships. Am Mus Nov 3183:1–38
Rougier GW, Novacek MJ, Wible JR (1998) Implications of Deltatheridium specimens for early marsupial history. Nature 396:459–463
Rougier GW, McKenna MC, Novacek MJ, Wible JR (2001) Gobiconodonts from the Early Cretaceous of Oshih (Aisle), Mongolia. Am Mus Nov 3348: 1–30
Rougier GW, Ji Q, Novacek MJ (2003) A new symmetrodont mammal with fur impressions from the Mesozoic of China. Acta Geol Sin 77:7–14
Rougier GW, Forasiepi AM, Hill RV, Novacek MJ (2009) New mammalian remains from the Late Cretaceous La Colonia Formation, Patagonia, Argentina. Acta Palaeontol Pol 54:195–212
Rougier GW, Apesteguía S, Gaetano L (2011) Highly specialized mammalian skulls from the Late Cretaceous of South America. Nature 479:98–102
Rougier GW, Apesteguía S, Beck R, Wible JR (2012) The Miocene mammal Necrolestes demonstrates the survival of a Mesozoic non-therian lineage into the late Cenozoic of South America. Proc Nat Acad Sci USA 109: 20053–20058
Rougier GW, Bolortsetseg M, Novacek MJ, Sheth AS, Spurlin BK (2016) Craniodental anatomy of a new Late Cretaceous multituberculate mammal from Udan Sayr, Mongolia. Palaeontol Pol 67:197–248
Rougier GW, Forasiepi AM, Martinelli AG (2021a) Mesozoic Mammals from South America and Their Forerunners. Springer Nature, Switzerland
Rougier GW, Turazzinni GF, Cardozo MS, Harper T, Lires AI, Canessa LA (2021b) New specimens of Reigitherium bunodontum from the Late Cretaceous La Colonia Formation, Patagonia, Argentina and meridiolestidan diversity in South America. J Mammal Evol. https://doi.org/10.1007/s10914-021-09585-2
Ruf I, Luo Z-X, Martin T, Wible JR (2009) Petrosal anatomy and inner ear structures of the Late Jurassic Henkelotherium (Mammalia, Cladotheria, Dryolestoidea): insight into the early evolution of the ear region in cladotherian mammals. J Anat 214:679–693
Ruf I, Luo Z-X, Martin T (2013) Reinvestigation of the basicranium of Haldanodon exspectatus (Mammaliaformes, Docodonta). J Vertebr Paleontol 33:382–400. https://doi.org/10.1080/02724634.2013.722575
Schultz JA, Luo Z-X, Zeller U (2016) Inner ear labyrinth anatomy of monotremes and implications for mammalian inner ear evolution. J Morphol 278:236–263. https://doi.org/10.1002/jmor.20632
Segall W (1970) Morphological parallelisms of the bulla and auditory ossicles in some insectivores and marsupials. Fieldiana 51:169–205
Sigogneau-Russell D, Ensom PC, Hooker JJ (2001) The oldest tribosphenic mammal from Laurasia (Purbeck Limestone Group, Berriasian, Cretaceous, UK) and its bearing on the "dual origin" of Tribosphenida. Comptes Rendus de l'Académie des Sciences 333:141–147
Simpson GG (1928) A Catalogue of the Mesozoic Mammalia in the Geological Department of the British Museum. Trustees of the British Museum, London
Simpson GG (1929) American Mesozoic Mammalia. Mem Peabody Mus 3:1–235
Wible JR (1986) Transformation in the extracranial course of the internal carotid artery in mammalian phylogeny. J Vertebr Paleontol 6:313–325
Wible JR (1987) The eutherian stapedial artery: character analysis and implications for superordinal relationships. Zool J Linn Soc 91:107–135
Wible JR, Hopson JA (1993) Basicranial evidence for early mammal phylogeny. In: Szalay FS, Novacek MJ, McKenna MC (eds) Mammal Phylogeny, Volume 1—Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials. Springer-Verlag, Inc., New York, pp 45–62
Wible JR, Hopson JA (1995) Homologies of the prootic canal in mammals and non-mammalian cynodonts. J Vertebr Paleontol 15:331–336
Wible JR, Rougier GW (2000) The cranial anatomy of Kryptobaatar dashzevegi (Mammalia, Multituberculata), and its bearing on the evolution of mammalian characters. Bull Am Mus Nat Hist 247:1–124
Wible JR, Rougier GW (2017) Craniomandibular anatomy of the subterranian meridolestidan Necrolestes patagonensis Ameghino, 1891 (Mammalia, Cladotheria) from the early Miocene of Patagonia. Ann Carnegie Mus 84:183–251
Wible JR, Dashzeveg D, McKenna MC, Novacek MJ, Rougier GW (1995) A mammalian petrosal from the Early Cretaceous of Mongolia: implications for the evolution of the ear region and mammaliamorph relationships. Am Mus Nov 3149:1–19
Wible JR, McKenna MC, Novacek MJ, Rougier GW (2001) Earliest eutherian ear region: a petrosal of Prokennalestes from the Early Cretaceous of Mongolia. Am Mus Nov 3322:1–44
Zeller U (1985a) Die Ontogenese und Morphologie der Fenestra rotunda und des Aquaeductus cochleae von Tupaia und anderen Säugern. Gegenbaurs Morphol Jahrb, Leipzig 131:179–204
Zeller U (1985b) The morphogenesis of the fenestra rotunda in mammals. Fort der Z 30:153–157
Zeller U (1989) Die Enwicklung und Morphologie des Schädels von Ornithorhynchus anatinus (Mammalia: Prototheria: Monotremata). Abhandlungen der Senck Naturf Ges 545:1–188
Acknowledgements
We dedicate this contribution to John Wible for his leadership in making JME a leading journal in our field and his monumental contributions to our understanding of the evolutionary patterns of the ear region throughout the history of mammals. Dr. Jessie Maisano (UTCT) obtained micro-CT data of the specimens in this study. We thank Simone Hoffmann and an anonymous reviewer for improvements to the manuscript. Funding for this project has come in part from a Discovery Pool grant to BMD from the Canyonlands Natural History Association (Moab, UT). This project has been partially supported by the ASNB Department (BMD and GWR) at the University of Louisville, Louisville, KY. CT scanning and data management support were provided by National Science Foundation grant FRES-1925896 to RLC. Additional support for GWR’s research was provided by PICT 2016 2682, a grant from the Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación, Argentina.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
GWR is an associate editor for the Journal of Mammalian Evolution but was not involved in the evaluation of this manuscript.
Availability of Data
CT data generated and analyzed for the current study are available from the corresponding author on reasonable request.
Rights and permissions
About this article

Cite this article
Davis, B.M., Cifelli, R.L. & Rougier, G.W. Mammalian Petrosals from the Upper Jurassic Morrison Formation (Utah, USA) Reveal Non-canonical Evolution of Middle and Inner Ear Characters. J Mammal Evol 28, 1027–1049 (2021). https://doi.org/10.1007/s10914-021-09586-1
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10914-021-09586-1