
Abstract
Mutations in the FAS gene are the most common cause of Autoimmune Lymphoproliferative Syndrome (ALPS), and the majority of them affect the intracellular domain of FAS protein, particularly the region termed death domain. However, approximately one third of these mutations affect the extracellular region of FAS and most are stop codons, with very few missense changes having been described to date. We previously described 7 patients with a FAS missense extracellular mutation, C107Y, two in homozygozity and 5 in heterozygosity. We investigated here the mechanistic effects of this mutation and observed that the homozygous patients did not show any FAS surface expression, while the heterozygous patients had diminished receptor expression. Aiming to understand why a missense mutation was abolishing receptor expression, we analyzed intracellular FAS protein trafficking using fluorescent fusion proteins of wild type FAS, two missense extracellular mutants (FAS-C107Y and FAS-C104Y) and one missense change localized in the intracellular region, FAS-D260E. The FAS-C107Y and FAS-C104Y mutants failed to reach the cell surface, being retained at the endoplasmic reticulum, unlike the WT or the FAS-D260E which were clearly expressed at the plasma membrane. These results support haploinsufficiency as the underlying mechanism involved in the pathogenesis of ALPS caused by extracellular FAS missense mutations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Autoimmune Lymphoproliferative Syndrome (ALPS) is a rare human complex immunologic disorder characterized by early onset of adenomegaly and splenomegaly, blood cell-directed autoimmune phenomena, increased risk of developing lymphoma and an elevated number of circulating CD4− CD8− TCRαβ+ T cells, named double negative T (DNT) cells [1, 2]. The molecular defects underlying this disorder affect components of the extrinsic apoptosis pathway, namely FAS, FAS Ligand (FASL) and Caspase 10 genes. Somatic mutations in FAS are also a common cause of ALPS, while about one third of patients remain without any identified molecular defect [3].
FAS (also known as APO-1/CD95) is a prototype member of the death receptor subfamily which has many roles in the immune system, particularly in homeostasis by killing the antigen activated and autoreactive lymphocytes [4–7]. The protein belongs to the tumor necrosis factor receptor superfamily (TNFRSF) and has 3 cysteine rich domains (CRD) in the extracellular region [8, 9]. The first domain is involved in the homotypic interaction of the receptor, termed the pre-ligand assembly domain (PLAD) [10]; the CRD2 and CRD3 are related with FASL binding [11, 12]. The FAS gene (or TNFRSF6) has 9 exons, and is located on the long (q) arm of chromosome 10 at position 24.1 [13–15]. The most common causes of ALPS are germline heterozygous mutations in FAS affecting the intracellular domain (ICD) of the protein, particularly into the death domain. The pathophysiological mechanism in this type of mutations is well explained by a dominant negative effect: the presence of one monomer of mutated FAS ruins the active trimmer impairing normal receptor function [16–18]. About a third of patients, however, harbor mutations in exons 1 to 5 of FAS, which encode for the extracellular domain (ECD) of the protein, being the majority of them nonsense changes. In addition, only 12 missense mutations were described in the extracellular domain but the mechanism by which they cause disease is not clear [19–22].
We have previously described an ECD missense mutation in FAS c.514A>G (p.C107Y) in 7 Argentinean ALPS patients [23]; two homozygous, with a severe clinical phenotype, and 5 unrelated heterozygous. Aiming to contribute to the understanding of the pathophysiological mechanism in ALPS patients carrying ECD missense mutations, we evaluated protein expression and intracellular localization of FAS-C107Y.
Methods
Patient Samples, Apoptosis Assay and FAS Expression
Two families were completely diagnosed in the Immunology Unit of the Hospital de Pediatría J.P. Garrahan. The rest of the patients were referred to the Hospital for laboratory studies. For the apoptosis assay, PBMC were activated for 3 days with phytohemagglutinin, M form (PHA-M, GIBCO). After 72 h, dead cells were removed by density gradient centrifugation. T cells were maintained in 40 U/ml IL-2 until day 9. Fresh medium was replaced at days 5 and 8 of culture. Fas sensitivity was tested using cross-linked anti-CD95 Ab APO-1-3 for 24 h. For quantification, cells were resuspended in a hypotonic solution containing 0.1 % sodium citrate, 0.1 % Triton X-100 (Sigma) and 50 μl/ml propidium iodide (PI, Sigma). The PI fluorescence of individual nuclei was measured with a FACSort flow cytometer (Beckton Dickinson, BD, San Diego, CA) and the percentage of hypodiploid nuclei was evaluated. The data are expressed as the percentage of induced cell death calculated as follows: (apoptosis observed in the patient)/(apoptosis observed in the control) × 100. The in vitro apoptosis observed in the control was greater than 80 %.
Three different anti Fas monoclonal antibodies (Abs) were used to stain activated T cells: unconjugated APO-1-3 (Alexis) and B-10 (Santa Cruz) followed by staining with FITC-coupled goat antibody to mouse immunoglobulin (BD), and DX2 PE-conjugated Ab (BD). For intracellular staining cell, permeabilization (Fix&Perm, Caltag) was required. Fas expression was analyzed on a flow cytometer (FACSort, BD).
Plasmids, Cell Lines and Transfection Protocol
The cDNAs from a FAS wild type control were cloned using TOPO-TA Cloning Kit (Invitrogen) and inserted into the GFP or RFP vectors (pEGFP or pDsRed2-C1, Clontech). In-frame GFP and RFP fusions with FAS were generated by standard PCR cloning techniques; the signal peptide of FAS was introduced in the NH2 terminal of GFP or RFP and the complete FAS protein was introduced in the COOH terminal of the fluorescent protein. The primers used are described in the Appendix 1. The following mutants were obtained by a site-directed mutagenesis: C104Y, C107Y and D260E and the primers are detailed in the Appendix 2. The mutations were numbered considering the reference transcript HGNC:11920. The correct sequence of each plasmid was confirmed by Sanger sequencing (Applied Biosystems 3130 Genetic Analyzer).
The HEKT 293 cell line was obtained from American Type Culture Collection (ATCC; Manassas, VA) and was grown in DMEM (GIBCO) with 10 % FBS supplemented with HEPES, glutamine and antibiotic- antimycotic (GIBCO).
HEK 293T cells were transiently transfected using TransIT-2020 Transfection Reagent (MIRUS) according to the manufacturer’s protocol and harvested 48 h post-transfection. The fluorescence expression was confirmed in an epifluorescence microscope, IN Cell Analyzer 1000 Cellular Imaging and Analysis System (General Electrics).
Western Blot Analysis
The molecular weight of the fusion proteins were calculated with a bioinformatics tool named Expasy (www.expasy.org). The results for the FAS-WT-GFP construction was 70231.98 Da and for RFP-FAS.WT 68668.35 Da. The fusion proteins were obtained from the lysis at room temperature of the transfected HEK cells with RIPA buffer (Tris pH 7.4 10 mM, NaCl 1 mM, EDTA 1 mM, Triton X-100 1 %, SDS 0.1 %, Nonidet Np40 substitute 1 %), sodium ortovanadate and protease inhibitor (SIGMA). The proteins were quantified with BCA Protein Assay Kit (Thermo Scientific). CD95 expression analysis was performed on 4–12 % SDS-PAGE with a polyclonal Ab anti-CD95 (G-9, Santa Cruz Biotechnology Inc.) and a polyclonal Ab anti-GFP (FL-8, Santa Cruz Biotechnology Inc.). Deglycosylation of cell lysates was performed with EndoH (New England Biolabs) according to the manufacturer’s protocol and then blotted with anti-CD95 (G-9, Santa Cruz Biotechnology Inc.).
To observe the behavior of the proteins in a native gel, HEK293T cells were transiently transfected with different fusion proteins, and whole-cell lysates were run on a polyacrylamide gel in non-reducing conditions (NativePAGE™ Novex® Bis-Tris Gel System, Invitrogen). The proteins were then immunoblotted using an anti-FAS Ab (G-9 clone).
Flow Cytometry Analysis in Transfected Cell Lines
HEK 293T cells were transiently transfected with the indicated GFP fusion protein constructs. After 48 h, cells were harvested in phosphate-buffered saline and stained either with an APC-conjugated anti-CD95 (DX2, BD Biosciences), or with Ab anti-GFP (FL-8, Santa Cruz) or with Ab anti-CD95 (G-9, Santa Cruz Biotechnology, Inc.) followed by an APC-conjugated secondary Ab (BD). For the intracellular stain, the cells were treated as describe above. The stained cells were analyzed on a FACSCanto cytometer (BD) and the data were analyzed with the FlowJo software package (Treestar Inc.).
Confocal Microscopy
For Golgi detection, the cell lines were grown in glass-Lab Tek Chamber system (Nunc) and transfected with the GFP-FAS constructions. Cells were fixed with PFA 4 % (UBS) and permeabilized with Triton X-100 0.25 %-PBS 1X, in the same glass recipient. We used a rabbit polyclonal anti-giantin Ab (Abcam), followed by Goat polyclonal Ab to Rabbit-Texas Red (Abcam). For detection of ER compartment, we cotransfected the cell line with GFP-FAS constructions and pDsRed-ER (Clontech) in an equal final amount of DNA (1:1), then the cells were fixed as described above. The slides were mounted with Vectashield Hard Set Mounting Medium with DAPI (Vector Laboratories). Images were analyzed in a Leica TCS SP5 / TCS SP5 X Confocal Microscope, collected using 63× Oil Immersion objective with the NA 1.32 and processed using the image processing software Image J.
Results
Pattern of FAS Expression and Apoptotic Assay in ALPS Patients
The FAS-C107Y mutation was found in 6 unrelated Argentinean families and the ALPS clinical presentation was severe in 2 homozygous brothers from the first family diagnosed, while a mild form of the disease was observed in the remaining 5 heterozygous patients [23]. The CD95–induced apoptosis was clearly reduced in the peripheral blood lymphocytes either from heterozygous patients and FAS C107Y healthy carriers as compared with normal controls; and completely abolished in the homozygous patient’s lymphocytes (Fig. 1a).

Defective apoptotic assay and FAS expression in ALPS patients. a CD95-induced cell death: PBMCs were activated for 5 days (3 days with PHA and 6 days with IL2) and incubated in the presence of Apo1.3. Cell death was determined by the percentage of hypodiploid nuclei corresponding to fragmented DNA, after treating the cells with a propidium iodide hypotonic solution. The results are expressed in % of induced cell death as a function of the concentration of the Ab Apo1.3. Data were represented as means of three separate experiments and SD was <10 %. b FAS expression on activated T cells from C107Y homozygous patient, C107Y heterozygous patient and normal control with Apo1.3 anti FAS antibody. c Comparable Fas expression between permeabilized T cells from a heterozygous patient, by CD95 B-10 clone Ab staining (against C-terminus)
We then investigated cell surface FAS expression, and noticed that it was completely lack in T lymphocytes of the patients with homozygous mutations (Fig. 1b). We also observed a minor but reproducible reduction in cell surface FAS expression in T cells from heterozygous patients or carriers using several antibodies (Fig. 1b and data not shown). Interestingly, when we permeabilized the cells from heterozygous carriers or patients and stained with anti-FAS Ab (clone B-10) directed against the C-terminus of the protein, FAS expression was similar to normal controls (Fig. 1c). This suggested that the total amount of synthesized protein was similar between controls and heterozygous carriers, and we decided to investigate where it was trapped.
FAS Extracellular Mutants Lack Surface Expression in Transfected HEK 293T Cells
In order to study the expression behavior of C107Y we constructed an in vitro model by generating fusion proteins between FAS and fluorescent proteins GFP or RFP (GFP-FAS-WT or RFP-FAS-WT, respectively). We applied site-directed mutagenesis to generate the C107Y mutant, as well as other missense extracellular change C104Y and an intracellular mutant, D260E. The amino acid 104 is other cysteine belonging to the CRD2, and there is a missense mutation in this position previously described in an ALPS patient C104G (named as C88G in [24]). The ICD mutation D260E was observed in another Argentinean ALPS patient (Danielian S. unpublished data).
We performed the experiments tagging the GFP or RFP on the C-terminus or the N-terminus of FAS mutants and the expression of the fusion proteins resulted similar (data not shown). The GFP-FAS or RFP-FAS fusion proteins (N-terminus) were transiently transfected in HEK 293T cell lines; the expression of WT, both extracellular mutants C104Y and C107Y, and D260E were confirmed by epifluorescence microscopy (data not shown) as well as by immunoblotting (Fig. 2a). The level of fusion protein expression was consistent with the protein load in each lane (Fig. 2a).

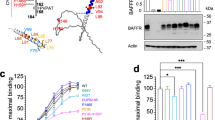
Lack of expression of FAS extracellular mutants at the surface. a Western Blot analysis of FAS expression. The apparent molecular weight of each fusion protein is approximately 70 kDa ( ). Note a band of expression corresponding to endogenous FAS (≈50 kDa) from the HEK 293 cell line (
). Note a band of expression corresponding to endogenous FAS (≈50 kDa) from the HEK 293 cell line ( ). Immunoblot for β-actin is shown as loading control. b Fas expression in transfected cell lines by flow cytometry using an anti FAS Ab (G-9 clone). Analyses were performed by gating on GFP + live cells according to forward- and side-scatter characteristics. The results are representative of three independent experiments. c Fas fusion protein expression in transfected cell lines by flow cytometry using an anti GFP Ab in non permeabilized and permeabilzed cells. Analysis was performed by gating on GFP-transfected and live cell lines
). Immunoblot for β-actin is shown as loading control. b Fas expression in transfected cell lines by flow cytometry using an anti FAS Ab (G-9 clone). Analyses were performed by gating on GFP + live cells according to forward- and side-scatter characteristics. The results are representative of three independent experiments. c Fas fusion protein expression in transfected cell lines by flow cytometry using an anti GFP Ab in non permeabilized and permeabilzed cells. Analysis was performed by gating on GFP-transfected and live cell lines
While the WT and D260E were clearly expressed at the surface (Fig. 2b and c), the extracellular mutants were not expressed at the cell surface. However, FAS proteins could be detected intracellularly by flow cytometry using a polyclonal anti-FAS Ab (G-9) (data not shown) or an anti-GFP Ab (FL-8) (Fig. 2c). Overall, these results are reminiscent of the pattern of expression observed in T lymphocytes from patients and carriers (Fig. 1).
Localization of the FAS Extracellular Mutants
These results suggested that the mutants C104Y and C107Y were probably intracellular retained in transfected HEK 293T cells. In order to evaluate subcellular localization of the mutated proteins, the transfected cell lines were fixed and stained with an Ab against giantin, a resident protein of the Golgi apparatus. No overlapping with the Golgi marker was observed in fluorescent extracellular mutants C104Y and C107Y (Fig. 3a), while FAS-WT and FAS-D260E proteins could be detected at the plasmatic membrane (Fig. 3a). Only a small fraction of these last proteins colocalized with giantin in Golgi, probably in their physiological pathway to the membrane.

FAS extracellular mutants are retained at the ER. a Intracellular staining of transiently transfected cell lines (WT, C104Y, C107Y and D260E) with an anti giantin Ab, a Golgi apparatus marker. Images were analyzed in a Leica TCS SP5 / TCS SP5 X Confocal Microscope, collected using 63× Oil Immersion objective with the NA 1.32. Digital images were analyzed with the image processing software Image J. The images were representative of three different experiments. b HEK293T cells were cotransfected with GFP-FAS and pDsRed-ER plasmids, the latter encodes for a fusion protein that label the ER in living cells. Co-localization is visible as yellow staining in the merged images. Images were analyzed in a Leica TCS SP5 / TCS SP5 X Confocal Microscope, collected using 63× Oil Immersion objective with the NA 1.32. Digital images were analyzed with the image processing software Image J. The images were representative of three independent experiments. c EndoH deglycosylation assay. The cell lysates from transfected HEK cell line and HEK293 cell line mock were deglycosylated with EndoH and then immunoblot with an anti FAS Ab (G-9 clone). Immunoblot for β-actin is shown as loading control. d HEK293T cells were transiently transfected with different fusion proteins, and whole-cell lysates were run on a polyacrylamide gel in nonreducing conditions. The proteins were then immunoblotted using anti-FAS antibody. Note the presence of monomeric ( ) and oligomeric (
) and oligomeric ( ) forms of FAS
) forms of FAS
Subsequently, to evaluate whether the extracellular mutants were retained at the endoplasmatic reticulum (ER), we cotransfected the cell lines with GFP-FAS fusion proteins and a DsRed-ER, which encode for a fusion protein that migrates and localizes into the ER. Again, the WT and D260E plasmids were expressed at the membrane and we could observe co-localization foci on the ER (Fig. 3b). In contrast, FAS-C104Y and FAS-C107Y were retained at this organelle, which could be observed as an overlap between FAS-fusion proteins and the ER marker, by confocal microscopy (Fig. 3b).
To further confirm the localization of both extracellular mutants in the ER and taking into account that the synthetized glycoproteins mature along the exocytic way by incorporating N- glycans, we analyzed the glycosylation pattern of these proteins. The endoglycosidase H (EndoH) enzyme specifically recognizes the glycan residues that are added in the ER: high mannose and hybrid N-glycan’s, but no complex oligosaccharides incorporated later in the pathway. The FAS-WT showed a partially susceptible pattern, with the presence of one band resistant to the enzyme, compatible with the fraction achieving the cell surface (Fig. 3c). However, the extracellular FAS mutated proteins C104Y and C107Y are fully susceptible to EndoH suggesting a lack of complex oligosaccharides in their structure (Fig. 3c). In conjunction, these experiments demonstrate that the C104Y and C107Y FAS mutant proteins are trapped at the ER level, being unable to reach the cell surface.
In order to characterize the molecular nature of the synthetized proteins, we analyzed these fusion proteins by immunoblot in non-denaturing conditions. Indeed, we could evidence conformational changes affecting the ability to form oligomers (Fig.3d). In the electrophoretic profile, C107Y and C104Y presented one band while as expected the WT and the D260E mutant exhibited a high degree of oligomerization, thus presenting lower mobility on the gel (Fig. 3d). The latter probably correspond to mature glycoproteins of FAS achieving the cell membrane and composed by self-associated monomers.
Discussion
In this work we demonstrated that two FAS ECD missense mutations affecting conserved cysteines C104Y and C107Y could not achieve the cell membrane, being retained in the ER.
N-linked glycosylation occurs upon entry of FAS polypeptides into the lumen of the ER and involves the transfer of a carbohydrate moiety to an asparagine residue within a specific amino acid consensus sequence. The predicted sites for CD95 N-glycosylation are the amino acids N118 and N136 [25]. By the EndoH deglycosylation assay we confirmed that the mutants were always glycosylated at the ER. However, as we previously suggested by bioinformatics tools [23] the conserved cysteine change in C107Y disrupted an annotated disulphide bond formation site. Taken together, these results indicate that the mutant proteins once glycosylated in the ER did not fold correctly in order to be transported to the Golgi to follow the secretory pathway.
Misfolded N-glycosylated proteins may have different fates; in most cases they are efficiently disposed of by the ER-associated protein degradation (ERAD) pathway, but, if the degradation process is insufficient, the protein accumulates and may trigger a unfolding protein response (UPR) that protects the cell by enhancing the ER folding and degradation capacity. To evaluate a possible stress generation by accumulated mutants FAS proteins in the ER, we analyzed GRp78 or BiP expression by immunoblot in the HEK cell lines transfected with the GFP-FAS-WT or with the mutants (Simesen de Bielke et al. unpublished data). The resulting electrophoretic profile demonstrated no increase of BiP in HEK 293 cells transfected with FAS-C104Y and FAS-C107Y. Thus, we hypothesize that in vivo mutated FAS proteins would remain shortly in the ER not being rescued by ER chaperones, translocate to the cytosol to be ubiquitinated and degraded via proteasome.
ER retention behavior was also observed in other mutant members from the TNFR superfamily. Extracellular mutants affecting CRD in TNFR1 were also retained in the ER, concomitant with the presence of misfolded mutated proteins [26]. However, while TNFR1 mutants formed abnormal disulfide-linked oligomers trough the PLAD, FAS extracellular mutants failed to oligomerize as efficiently as wild-type proteins. In the same way, CD40 mutations affecting CRD2 resulted in reduced surface expression, being retained in the ER [27]. Interestingly, one of these affecting a conserved cysteine C83R remained stable in this organelle, and increased BiP levels [27], unlike our FAS mutants.
The pathophysiological mechanism of ICD mutations in ALPS have been widely studied in the past and shown to induce dominant-negative interference, disrupting the apoptotic signal in the cell [16, 18]. The receptor is pre-assembled at the surface and the presence of one ICD heterozygous mutant protein leads to the formation of 7 out of 8 homotrimers with at least one mutated monomer, impairing the adequate signaling. A third of ALPS-FAS patients presented ECD mutations and the majority of them carried nonsense, insertion, deletion or open frame splicing site mutations, predicted to abolish FAS expression [19]. Haploinsufficiency has been demonstrated to be the underlying pathophysiological mechanism in these ALPS patients [18, 19, 28, 29]. However, there still remain a small number of patients which carry ECD missense mutations whose pathophysiological mechanism is still debated [3]. We show here that two of these mutations, C104Y and C107Y, affecting the CRD2, induced FAS protein retention at the ER level, preventing cell surface expression, suggesting haploinsufficiency as the main pathological mechanism involved in FAS C107Y patients. In this way, haploinsufficiency could also be suggested from the behavior of homozygous patients’ cells versus those heterozygously mutated in C107Y (patients and healthy carriers) (Fig. 1): in homozygous cells no surface expression nor cell death induction through an anti-FAS Ab could be detected, while an intermediate surface staining and FAS function were exhibited by cells from heterozygous carriers despite a complete intracellular staining shown in the same cells (Fig. 1) in a comparable way with that observed in control cells.
The residual FAS function observed in heterozygous patient’s cells could explain in part the incomplete clinical penetrance observed in our families (5 heterozygous patients and 14 healthy carriers). It has been demonstrated that in low penetrance ECD mutations a somatic event can be present in the second allele form DNT cells associated with ALPS clinical presentation [22]. In this way, at the moment we found a second mutation in DNT cells from the two evaluated patients out of the 5 heterozygous patients (Simesen de Bielke et al. unpublished data). Environmental factors or other genetic abnormalities may also have an impact on clinical penetrance. Further studies in these patients and healthy carriers could elucidate alternative mechanisms involved in the variable clinical expressivity of this disease.
References
Rieux-Laucat F, Le Deist F, Hivroz C, Roberts IA, Debatin KM, Fischer A, et al. Mutations in Fas associated with human lymphoproliferative syndrome and autoimmunity. Science. 1995;268(5215):1347–9.
Fisher GH, Rosenberg FJ, Straus SE, Dale JK, Middleton LA, Lin AY, et al. Dominant interfering Fas gene mutations impair apoptosis in a human autoimmune lymphoproliferative syndrome. Cell. 1995;81(6):935–46.
Oliveira JB, Bleesing JJ, Dianzani U, Fleisher TA, Jaffe ES, Lenardo MJ, et al. Revised diagnostic criteria and classification for the autoimmune lymphoproliferative syndrome (ALPS): report from the 2009 NIH International Workshop. Blood. 2010;116(14):e35–40.
Alderson MR, Armitage RJ, Maraskovsky E, Tough TW, Roux E, Schooley K, et al. Fas transduces activation signals in normal human T lymphocytes. J Exp Med. 1993;178(6):2231–5.
Ju ST, Panka DJ, Cui H, Ettinger R, el-Khatib M, Sherr DH, et al. Fas(CD95)/FasL interactions required for programmed cell death after T-cell activation. Nature. 1995;373(6513):444–8.
Dhein J, Walczak H, Baumler C, Debatin KM, Krammer PH. Autocrine T-cell suicide mediated by APO-1/(Fas/CD95). Nature. 1995;373(6513):438–41.
Strasser A, Jost PJ, Nagata S. The many roles of FAS receptor signaling in the immune system. Immunity. 2009;30(2):180–92.
Itoh N, Yonehara S, Ishii A, Yonehara M, Mizushima S, Sameshima M, et al. The polypeptide encoded by the cDNA for human cell surface antigen Fas can mediate apoptosis. Cell. 1991;66(2):233–43.
Oehm A, Behrmann I, Falk W, Pawlita M, Maier G, Klas C, et al. Purification and molecular cloning of the APO-1 cell surface antigen, a member of the tumor necrosis factor/nerve growth factor receptor superfamily. Sequence identity with the Fas antigen. J Biol Chem. 1992;267(15):10709–15.
Siegel RM, Frederiksen JK, Zacharias DA, Chan FK, Johnson M, Lynch D, et al. Fas preassociation required for apoptosis signaling and dominant inhibition by pathogenic mutations. Science. 2000;288(5475):2354–7.
Orlinick JR, Vaishnaw A, Elkon KB, Chao MV. Requirement of cysteine-rich repeats of the Fas receptor for binding by the Fas ligand. J Biol Chem. 1997;272(46):28889–94.
Starling GC, Bajorath J, Emswiler J, Ledbetter JA, Aruffo A, Kiener PA. Identification of amino acid residues important for ligand binding to Fas. J Exp Med. 1997;185(8):1487–92.
Behrmann I, Walczak H, Krammer PH. Structure of the human APO-1 gene. Eur J Immunol. 1994;24(12):3057–62.
Cheng J, Liu C, Koopman WJ, Mountz JD. Characterization of human Fas gene. Exon/intron organization and promoter region. J Immunol. 1995;154(3):1239–45.
Inazawa J, Itoh N, Abe T, Nagata S. Assignment of the human Fas antigen gene (Fas) to 10q24.1. Genomics. 1992;14(3):821–2.
Jackson CE, Fischer RE, Hsu AP, Anderson SM, Choi Y, Wang J, et al. Autoimmune lymphoproliferative syndrome with defective Fas: genotype influences penetrance. Am J Hum Genet. 1999;64(4):1002–14.
Rieux-Laucat F, Blachere S, Danielan S, De Villartay JP, Oleastro M, Solary E, et al. Lymphoproliferative syndrome with autoimmunity: A possible genetic basis for dominant expression of the clinical manifestations. Blood. 1999;94(8):2575–82.
Vaishnaw AK, Orlinick JR, Chu JL, Krammer PH, Chao MV, Elkon KB. The molecular basis for apoptotic defects in patients with CD95 (Fas/Apo-1) mutations. J Clin Invest. 1999;103(3):355–63.
Kuehn HS, Caminha I, Niemela JE, Rao VK, Davis J, Fleisher TA, et al. FAS haploinsufficiency is a common disease mechanism in the human autoimmune lymphoproliferative syndrome. J Immunol. 2011;186(10):6035–43.
Neven B, Magerus-Chatinet A, Florkin B, Gobert D, Lambotte O, De Somer L, et al. A survey of 90 patients with autoimmune lymphoproliferative syndrome related to TNFRSF6 mutation. Blood. 2011;118(18):4798–807.
Price S, Shaw PA, Seitz A, Joshi G, Davis J, Niemela JE, et al. Natural history of autoimmune lymphoproliferative syndrome associated with FAS gene mutations. Blood. 2014;123(13):1989–99.
Magerus-Chatinet A, Neven B, Stolzenberg MC, Daussy C, Arkwright PD, Lanzarotti N, et al. Onset of autoimmune lymphoproliferative syndrome (ALPS) in humans as a consequence of genetic defect accumulation. J Clin Invest. 2011;121(1):106–12.
Simesen de Bielke MG, Yancoski J, Rocco C, Perez LE, Cantisano C, Perez N, et al. A missense mutation in the extracellular domain of Fas: the most common change in Argentinean patients with autoimmune lymphoproliferative syndrome represents a founder effect. J Clin Immunol. 2012;32(6):1197–203.
Rieux-Laucat F, Le Deist F, Fischer A. Autoimmune lymphoproliferative syndromes: genetic defects of apoptosis pathways. Cell Death Differ. 2003;10(1):124–33.
Shatnyeva OM, Kubarenko AV, Weber CE, Pappa A, Schwartz-Albiez R, Weber AN, et al. Modulation of the CD95-induced apoptosis: the role of CD95 N-glycosylation. PLoS One. 2011;6(5):e19927.
Lobito AA, Kimberley FC, Muppidi JR, Komarow H, Jackson AJ, Hull KM, et al. Abnormal disulfide-linked oligomerization results in ER retention and altered signaling by TNFR1 mutants in TNFR1-associated periodic fever syndrome (TRAPS). Blood. 2006;108(4):1320–7.
Lanzi G, Ferrari S, Vihinen M, Caraffi S, Kutukculer N, Schiaffonati L, et al. Different molecular behavior of CD40 mutants causing hyper-IgM syndrome. Blood. 2010;116(26):5867–74.
Hsu AP, Dowdell KC, Davis J, Niemela JE, Anderson SM, Shaw PA, et al. Autoimmune lymphoproliferative syndrome due to FAS mutations outside the signal-transducing death domain: molecular mechanisms and clinical penetrance. Genet Med. 2011;14(1):81–9.
Roesler J, Izquierdo JM, Ryser M, Rosen-Wolff A, Gahr M, Valcarcel J, et al. Haploinsufficiency, rather than the effect of an excessive production of soluble CD95 (CD95{Delta}TM), is the basis for ALPS Ia in a family with duplicated 3′ splice site AG in CD95 intron 5 on one allele. Blood. 2005;106(5):1652–9.
Acknowledgments
We thank the patients and their families for contributions to the study. We also thank Sundar Ganesan for his collaboration with confocal images. This research was supported by Agencia de Promoción Científica y Tecnológica grant PICT2008 0915.
Authorship Contributions
All authors have contributed in a substantive and intellectual manner to this work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors have no financial conflicts of interest.
Appendices
Appendix 1
PS-NheI-F: 5′ gccctggctagccaggcg 3′
PS-NheI-R: 5′ gatgcagggctagcaagttctgagtc 3′
PM-PstI-F: 5′-gttcttacgtctgcagctagattatcgtc-3′
PM-PstI-R: 5′-gaactgaatttgttgtttttcgctagcgaccaagctttggatttc-3′
Appendix 2
C104Y F: 5′ cttccaaatgcagaagatatagattgtgtgatgaaggacatggc 3′
C104Y R: 5′ gccatgtccttcatcacacaatctatatcttctgcatttggaag 3′
C107Y F: 5′ caaatgcagaagatgtagattgtatgatgaaggacatggcttagaag 3′
C107Y R: 5′ cttctaagccatgtccttcatcatacaatctacatcttctgcatttg 3′
D260E F: 5′ ggtgtcaatgaagccaaaatagaagagatcaagaatgacaatgtcc 3′
D260E R: 5′ ggacattgtcattcttgatctcttctattttggcttcattgacacc3′
Rights and permissions
About this article

Cite this article
de Bielke, M.G.S., Perez, L., Yancoski, J. et al. FAS Haploinsufficiency Caused by Extracellular Missense Mutations Underlying Autoimmune Lymphoproliferative Syndrome. J Clin Immunol 35, 769–776 (2015). https://doi.org/10.1007/s10875-015-0210-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10875-015-0210-0