
Abstract
Lysophosphatidic acid (LPA) is a simple lipid which is endogenously synthesized from lysophosphatidylcholine (LPC) by autotaxin (ATX). LPA mediates a variety of cellular responses through the binding of G protein-coupled LPA receptors (LPA1 to LPA6). It is considered that LPA receptor-mediated signaling plays an important role in the pathogenesis of human malignancy. Genetic alterations and epigenetic changes of LPA receptors have been detected in some cancer cells as well as LPA per se. Moreover, LPA receptors contribute to the promotion of tumor progression, including cell proliferation, invasion, metastasis, tumorigenicity, and angiogenesis. In recent studies, the activation of LPA receptor-mediated signaling regulates chemoresistance and radiosensitivity in cancer cells. This review provides an updated overview on the roles of LPA receptor-mediated signaling in the regulation of cancer cell functions and its potential utility as a molecular target for novel therapies in clinical cancer approaches.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Lysophosphatidic acid (LPA) is an extracellular lipid which evokes the intracellular signaling via binding to G-protein-coupled LPA receptors. At least six subtypes of LPA receptors (LPA receptor-1 (LPA1) to LPA6) have been determined. LPA signaling via LPA receptors indicates a variety of cellular responses, including cell growth, differentiation, morphogenesis, cell migration and protection from apoptosis (Geraldo et al. 2021; Stoddard and Chun 2015; Aikawa et al. 2015; Yung et al. 2014). The biological functions of the individual LPA receptors are not uniform, dependent on types of cells. It is considered that LPA receptor-mediated signaling plays an important role in the pathogenesis of human disease, such as cardiovascular disease, neuropathic pain, fibrosis and cancer (Lin et al. 2010; Tsujiuchi et al. 2014).
In the 1990s, Xu et al. reported that LPA was highly secreted in serum and ascites in aggressive ovarian cancers (Xu et al. 1995). Subsequently, numerous studies have indicated the involvement of LPA receptors in cancer cell biology as well as LPA per se. Genetic alterations and epigenetic changes of LPA receptor genes occur in some cancer cells. Moreover, LPA receptors participate in the promotion of tumor progression, such as cell proliferation, invasion, metastasis, tumorigenicity and angiogenesis (Lin et al. 2010; Tsujiuchi et al. 2014). In recent studies, the activation of LPA receptor-mediated signaling modulates chemoresistance and radiosensitivity in cancer cells (Ueda et al. 2020; Minami et al. 2019; Okuda et al. 2023). Therefore, it is suggested that LPA receptor-mediated signaling may be a target molecule for novel therapies in clinical cancer approaches. In this review, we provide an updated overview on the current evidence of the roles of LPA receptor-mediated signaling in the regulation of cancer cell functions.
LPA
LPA is a simple lipid and structurally consists of a glycerol, a fatty acid and a phosphate (Geraldo et al. 2021; Stoddard and Chun 2015; Aikawa et al. 2015). In 1970s, LPA is identified as a physiological molecule which modulates platelet aggregation, intracellular calcium release and blood pressure (Tokumura et al. 1978; Gerrard et al. 1979). LPA is found in not only all mammalian cells and tissues, but also in plasma, serum and saliva (Geraldo et al. 2021; Stoddard and Chun 2015; Aikawa et al. 2015). LPA is released from activated platelets and detectable at concentrations of approximately 1 to 5 µM in serum (Eichholtz et al. 1993). LPA is present as a mixture of several fatty acids in vivo condition; unsaturated fatty acids (16:1, 18:1, 18:2 and 20:4) and saturated fatty acids (16:0, 18:0) (Aoki et al. 2008). In human plasma, the most abundant LPA forms are 16:0, 18:2 and 18:1 (Sano et al. 2002) (Fig. 1). It is considered that there are at least two pathways for LPA synthesis. LPA is endogenously synthesized from lysophosphatidylcholine (LPC) by autotaxin (ATX). ATX is widely present in biological fluids, such as plasma, cerebrospinal fluid, synovial fluid and cancer ascites (Stracke et al. 1992). Conversely, membrane-bound phosphatidic acid-preferring phospholipase A1 also catalyzes the conversion of phosphatidic acid (PA) to LPA (Aoki et al. 2008).

Structure of 16:0-LPA. 16:0-LPA is one of the most abundant forms in human plasma (Sano et al. 2002)
LPA receptors
LPA receptors are members of G protein-coupled receptors (GPCR) (Arang and Gutking 2020). So far, six subtypes of LPA receptors have been identified as LPA1/EDG2, LPA2/EDG4, LPA3/EDG7, LPA4/P2Y9/GPR23, LPA5/GPR92 and LPA6/P2Y5 (Choi et al. 2008; Ishii et al. 2009). Additionally, LPA receptors are classified into two groups. LPA1, LPA2 and LPA3 belong to the endothelial cell differentiation gene (Edg) family. Conversely, LPA4, LPA5 and LPA6 have been determined as non-Edg LPA receptors which are the purinergic receptor family. These receptors are structurally distance from other LPA receptors (Choi et al. 2008; Ishii et al. 2009). LPA receptors are coupled to individual sets of G proteins (Gi, Gq, Gs and G12/13) and mediate a large variety of LPA effector functions (Table 1). The effects of each LPA receptor on cellular responses are not equivalent. For instance, LPA1 and LPA2 stimulate cell proliferation, intracellular calcium mobilization, adenylyl cyclase inhibition and phospholipase C activation (Geraldo et al. 2021). LPA3 increases axon branching via the activation of Gq protein in neural cells (Furuta et al. 2012). LPA4 and LPA5 provoke neurite retraction and stress fiber formation of neural cells (Geraldo et al. 2021). LPA6 is involved in the maintenance of human hair growth. In addition, homozygous mutation of LPA6 gene is the cause of hypotrichosis (Pasternack et al. 2008; Shimomura et al. 2009).
Roles of LPA signaling via LPA receptors in the pathogenesis of cancer cells
It has been reported that genetic and epigenetic alterations of LPA receptors are detected in cancer cells as well as LPA per se (Tsujiuchi et al. 2014). LPA is present at high concentrations in blood and ascites from ovarian cancer patients (Xu et al. 1995). In colon and gastric cancer cells, LPA contributes to the modulation of cell proliferation, migration and adhesion (Shida et al. 2003, 2004a). Moreover, ATX overexpression is associated with the promotion of malignant potency during tumor progression in several cancer cells (Samadi et al. 2011; Leblanc and Peyruchaud 2015; van Meeteren and Moolenaar 2007). Mutations of LPAR2 and LPAR4 genes are found in colon cancer cells (Tsujino et al. 2010). LPAR1 and LPAR3 mutations occur in osteosarcoma, while no mutation of LPA receptors is detected in in fibrosarcoma cells (Okabe et al. 2010). In contrast, rodent tumors induced by chemical carcinogens harbor high frequent mutations of Lpar1 gene. Lpar1 gene mutations are detected 46.7% in rat liver tumors induced by N-nitrosodiethylamine (Obo et al. 2009). Moreover, the frequency of Lpar1 gene mutations is 16.7% in adenomas and 41.2% in adenocarcinomas during rat lung carcinogenesis induced by N-nitrosobis(2-hydroxypropyl)amine (Yamada et al. 2009). Mutant LPA1 positively regulates malignant properties of cancer cells (Hayashi et al. 2012; Kato et al. 2012). Aberrant LPA receptor expressions are detected in human cancer cells. LPA2 expressions are significantly higher in thyroid and breast cancer cells than in normal tissues, but not LPA1 expressions (Schulte et al. 2001). The expression levels of LPA2 are increased in breast cancer cells, while no change of LPA1 and LPA3 expressions is observed (Kitayama et al. 2004). LPA1 expressions are elevated and LPA2 expressions are reduced in colorectal cancers, compared with normal surrounding tissues (Shida et al. 2004b). In addition, loss of LPA receptor expressions is due to hyper DNA methylation of the promoter region of LPA receptor genes in colon cancer and osteosarcoma cells (Tsujino et al. 2010; Okabe et al. 2011).
Regulation of cellular functions via LPA receptor-mediated signaling in cancer cells
Pancreatic cancer cells
The cell motile and invasive activities of pancreatic cancer cells are stimulated by LPA1, LPA3 and LPA6. Conversely, LPA2, LPA4 and LPA5 inhibit the cell motility and invasion of pancreatic cancer cells. The activation of matrix metalloproteinase-2 (MMP-2) is elevated by LPA1, LPA2, LPA5 and LPA6 (Fukushima et al. 2017; Komachi et al. 2009; Ishii et al. 2015). In addition, the cell motility is decreased by culturing in low glucose mediums, while LPAR1 and LPAR2 expression levels are increased (Takai et al. 2023). It is well known that MMP-2 activation participates in the promotion of cancer cell invasion and metastasis during tumor progression as well as MMP-9 (Kessenbrock et al. 2010). In colony assay, LPA1, LPA3 and LPA6 enhance the colony formation of pancreatic cancer cells, while LPA4 and LPA5 inhibit (Fukushima et al. 2017; Ishii et al. 2015). ATX-LPA axis promotes tumor progression of pancreatic cancer cells, such as peritoneal seeding and malignant ascites (Jinno et al. 2021).
Gastrointestinal cancer cells
In gastric cancer cells, the cell migration of LPAR1-expressing cells is increased by LPA, but not LPAR2-expressing cells (Shida et al. 2004a). In colon cancer cells, LPA stimulated the cell proliferation, migration and adhesion of LPAR1-expressing cells. In contrast, LPA did not affect the cell migration and adhesion of LPAR2-expressing cells, whereas it increased the cell growth activity (Shida et al. 2003). LPA facilitates the colon cancer cell growth activity through ROCK and STAT-3 pathways (Leve et al. 2018). The cell motility and invasion of colon cancer cells are suppressed by LPA3, LPA4 and LPA6 (Fukui et al. 2012; Takahashi et al. 2017a). While LPA1 forms the large sized colonies of colon cancer cells, the colony formation is decreased by LPA6 (Takahashi et al. 2018a). LPA induces the secretion of angiogenic factors through LPA1 and LPA2 in colon cancer cells (Shida et al. 2003).
Ovarian cancer cells
The cell death is promoted through apoptosis and anoikis by LPA in highly LPA1-expressing cells, while LPA inhibits the cell growth activity (Furui et al. 1999; Fang et al. 2002). The expression levels of vascular endothelial growth factor (VEGF) are associated with LPA2 and LPA3 expression levels in ovarian cancer cells (Fujita et al. 2003). LPA2 knockdown inhibits the production of VEGF as well as LPA3 knockdown (Yu et al. 2008). LPA increases VEGF mRNA expression and protein secretion (Hu et al. 2001). The cell motility and invasion of ovarian cancer cells are elevated by LPA3. In mouse xenograft study, LPA2 and LPA3 enhance tumor growth, ascites formation and metastatic potency to distant organs, resulting in the reduction of the survival rate of mice (Yu et al. 2008).
Bone and soft tissue sarcoma cells
The cell motility and invasion of fibrosarcoma cells are stimulated by LPA2 (Takahashi et al. 2017b). In highly migratory osteosarcoma cells, the cell motile activity is closely associated with LPAR2 gene expression (Takahashi et al. 2018b). The cell motile activity of osteosarcoma cells is stimulated via LPA2-mediated signaling activated by culturing with endothelial cells (Minami et al. 2021). In soft-agar colony formation assay, LPA2 enhances the colony formation activity in fibrosarcoma and osteosarcoma cells (Takahashi et al. 2017b, 2018b). In contrast, LPA1-mediated signaling suppresses pulmonary metastasis of osteosarcoma cells (Takagi et al. 2021). The cell motile and invasive activities are reduced by LPA5 in osteosarcoma and fibrosarcoma cells (Minami et al. 2020a; Dong et al. 2014).
Others
In neuroblastoma cells, the cell motility and invasion are elevated by LPA2 and LPA3. Although wild-type LPA1 suppresses the cell motility and MMP-2 activation, mutant LPA1 has promoting effects. Moreover, mutant LPA1 enhances the colony formation as well as LPA3 (Hayashi et al. 2012; Kato et al. 2012). In lung cancer cells, the cell motile activity is suppressed by LPA1 and LPA2. Conversely, LPA3 stimulates the cell motile activity and inhibited angiogenesis (Ueda et al. 2020; Tanabe et al. 2013). The cell motility and invasion of hepatoma cells are stimulated by LPA3. LPA3 enhances the colony formation of hepatoma cells (Okabe et al. 2013).
Chemoresistance via activation of LPA receptor-mediated signaling
Multidrug resistance is a pharmacological phenomenon of the simultaneous tolerance to functionally and structurally unrelated anticancer drugs and toxic compounds. The acquisition of multidrug resistance is one of the major causes of chemotherapeutic failure during cancer treatment (Hamilton and Rath 2014). It has been reported that LPA receptor-mediated signaling participates in the modulation of chemoresistance of cancer cells. In ovarian cancer cells, LPA1-expressing cells shows the low cell proliferation activity and high cell viability to CDDP, compared with LPA1-unexpressing cells (Furui et al. 1999). The cell survival rate to CDDP is enhanced through LPA2-mediated signaling in fibrosarcoma, osteosarcoma and lung cancer cells (Ueda et al. 2020; Minami et al. 2020b; Kurisu et al. 2022). The cell survival is elevated by LPA3 in hepatoma cells treated with CDDP and doxorubicin (Okabe et al. 2013). In contrast, LPA3 decreases the cell survival rate to CDDP of lung cancer and osteosarcoma cells (Ueda et al. 2020; Kurisu et al. 2022). LPA5 reduces the cell survival to CDDP of osteosarcoma cells (Minami et al. 2020a). The cell survival is decreased by LPA5 in melanoma cells treated with CDDP and dacarbazine (Minami et al. 2019). The cell survival to CDDP is suppressed through apoptosis by LPA4 and LPA6 in osteosarcoma cells (Kurisu et al. 202).
It is considered that activation of ATP-binding cassette (ABC) transporters and drug-detoxifying enzyme facilitates the acquisition of multidrug resistance in cancer cells. ABC transporters act as the efflux pumps of anticancer drugs through the cell membrane. ABC transporters are composed of at least 48 members. In particular, ABCB1, ABCC1, ABCC10 and ABCG2 contribute to the promotion of multidrug resistance. ABC transporters requires abundant ATP production as the energy molecule in cancer cells (Kathawala et al. 2015; Chen et al. 2016; Sau et al. 2010). It is suggested that the depletion of intracellular ATP may result in the suppression of chemoresistance through ABC transporters. In the presence of LPA, the cell survival to CDDP is decreased in ATP-reduced osteosarcoma cells (Kurisu et al. 2022). On the other hand, ATP is used as the substrate for cAMP synthesis by adenylyl cyclase activity (Steegborn 2014). cAMP induces apoptosis and promotes chemoresistance in some types of tumor cells (Zhang et al. 2020; Insel et al. 2012). Gs protein stimulates and Gi protein suppresses the adenylyl cyclase activity (Stoddard and Chun 2015). Therefore, the differential effects of the individual LPA receptors on cell survival to anticancer drugs may be due to the intracellular cAMP accumulation levels in cancer cells. Moreover, Rho family is involved in the regulation of chemosensitivity to CDDP (Mokady et al. 2015). G12/13 protein activates Rho signaling pathway (Geraldo et al. 2021). While RhoA and RhoC have the antiapoptotic effects, resulting in chemoresistance to CDDP, the proapoptosis induced by RhoB activation enhances chemosensitivity to CDDP (Mokady et al. 2015). In the presence of LPA2 agonist, the cell survival to CDDP of fibrosarcoma cells is inhibited by RhoA and RhoC knockdowns (Minami et al. 2020b) (Fig. 2).

Roles of LPA signaling via LPA receptors in the regulation of cell survival to anticancer drugs in cancer cells. Gi protein inhibits and Gs protein stimulates adenylyl cyclase activity. The increased amount of cAMP causes tumor suppression, such as apoptosis. ABC transporters require the abundant ATP to excrete chemotherapeutic agents across cellular membrane. G12/13 protein activates Rho-mediated signaling. RhoA and RhoC have the antiapoptotic effects, resulting in chemoresistance. RhoB induces the proapoptosis in cancer cells treated with anticancer drugs
Involvement of LPA receptor-mediated signaling in radiation sensitivity
Radiation is one of the common treatments for a variety of cancers as well as chemotherapy. Ionizing radiation directly produces DNA damage and induces DNA double-strand breaks, resulting in tumor cell death (Huang and Zhou 2020). The activation of LPA2 promotes the response to DNA damage induced by gamma irradiation. Exposure to gamma irradiation elevates the expression level of LPAR2 gene in intestinal epithelial cells. In mouse models, treatment of gamma irradiation increases the plasma ATX activity and LPA concentrations (Balogh et al. 2015). Furthermore, LPAR2 gene expressions are increased in fibrosarcoma, pancreatic and lung cancer cells irradiated with X-rays. The cell motile activity of pancreatic cancer cells is reduced through LPA2 by X-ray irradiation. On the other hand, the cell survival to X-ray irradiation is enhanced by the activation of LPA2-mediated signaling in pancreatic cancer cells (Okuda et al. 2023).
Cancer stem cell and LPA receptor-mediated signaling
Cancer stem cells (CSCs) are conceptually proposed as a subpopulation of tumor cells exhibiting stem cell properties like self-renewal, sphere-forming and multi-lineage differentiation ability. CSCs are considered to be tumor initiating cells and could be an origin for tumor heterogeneity and involved in metastasis and relapse (Visvader and Lindeman 2008). CSCs has been identified in various types of hematologic and solid cancers (Bonnet and Dick 1997; Hermann et al. 2007; Al-Hajj et al. 2003; Fujii et al. 2009; Honoki et al. 2010). Various mechanisms are implicated in the evasion of CSC from therapy such as enhanced DNA damage repair, altered cell cycle checkpoint control and overexpression of multidrug resistance proteins (Morrison et al. 2011).
In ovarian cancer cells, LPA treatment stimulates the expression of CSC-associated stem cell marker genes, including OCT4, SOX2, ALDH1 and drug transporters. Moreover, LPA promotes CSC-like characteristics: epithelial-to-mesenchymal transition, sphere forming ability and resistance to anti-cancer drugs. Therefore, it is suggested that LPA plays a key role in the therapeutic resistance and tumor progression of ovarian CSCs (Seo et al. 2016).
ATX-LPA signaling axis participates in the interaction between tumor cells and tumor environment through the crosstalk of tumor cells and tumor-associated fibroblasts originated from mesenchymal stem cells (MSCs). Accumulating evidence suggests that MSCs promote in vivo growth of xenograft transplanted tumors as well as animal models (Huang et al. 2013; Tsukamoto et al. 2012). Interestingly, cancer-derived LPA facilitates the differentiation of human MSCs to myofibroblast-like cells (Jeon et al. 2008). Furthermore, LPA signaling promotes the secretion of cytokines like vascular endothelial growth factor and stromal cell-derived factor-1/CXCL12 from MSCs (Ptaszynska et al. 2010; Jeon et al. 2010). Taken together, targeting of ATX-LPA signaling pathway could be a potential therapeutic candidate to overcome the therapy-resistance and disease relapse as well as metastatic spreading in cancer.
Conclusion
In this review, we provide an overview of the pivotal roles of LPA receptor-mediated signaling in the regulation of cancer cell functions. LPA receptor-mediated signaling consists of complicated components of G protein-coupled LPA receptors and involved in several physiological functions of normal and cancer cells. Therefore, further efforts should be attempted to clarify the specific molecular mechanisms linked to LPA receptor-mediated signaling in cancer cells and establish a novel therapeutic approach for cancer treatment.
Data availability
Not applicable.
References
Aikawa S, Hashimoto T, Kano K, Aoki J (2015) Lysophosphatidic acid as a lipid mediator with multiple biological actions. J Biochem 157:81–89
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF (2003) Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci USA 100:3983–3988
Aoki J, Inoue A, Okudaira S (2008) Two pathway for lysophosphatidic acid production. Biochim Biophys Acta 1781:513–518
Arang N, Gutking JS (2020) G protein-coupled receptors and heterotrimeric G proteins as cancer drivers. FEBS Lett 594:4201–4232
Balogh A, Shimizu Y, Lee SC, Norman DD, Gangwar R, Bavaria M, Moon C, Shukla P, Rao R, Ray R, Naren AP, Banerje S, Miller DD, Balazs L, Pelus L, Tigyi G (2015) The autotaxin-LPA2 GPCR axis is modulated by γ-irradiation and facilitates DNA damage repair. Cell Signal 27:1751–1762
Bonnet D, Dick JE (1997) Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med 3:730–737
Chen Z, Shi T, Zhang L, Zhu P, Deng M, Huang C, Hu T, Jiang L, Li J (2016) Mammalian drug efflux transporters of the ATP binding cassette (ABC) family in multidrug resistance: a review of the past decade. Cancer Lett 370:153–164
Choi JW, Lee CW, Chun J (2008) Biological roles of lysophospholipid receptors revealed by genetic null mice: an update. Biochim Biophys Act 1781:531–539
Dong Y, Hirane M, Araki M, Fukushima N, Honoki K, Tsujiuchi T (2014) Lysophosphatidic acid receptor-5 regulates cells motile and invasive avtivities of human sarcoma cell lines. Mol Cell Biochem 393:17–22
Eichholtz T, Jalink K, Fahrenfort I, Moolenaar WH (1993) The bioactive phospholipid lysophosphatidic acid is released from activated platelets. Biochem J 291:677–680
Fang X, Schummer M, Mao M, Yu S, Tabassam FH, Swaby R, Hasegawa Y, Tanyi JL, LaPushin R, Eder A, Jaffe R, Erickson J, Mills GB (2002) Lysophosphatidic acid is a bioactive mediator in ovarian cancer. Biochim Biophys Acta 1582:257–264
Fujii H, Honoki K, Tsujiuchi T, Kido A, Yoshitani K, Takakura Y (2009) Sphere-forming stem-like cell populations with drug resistance in human sarcoma cell lines. Int J Oncol 34:1381–1386
Fujita T, Miyamoto S, Onoyama I, Sonoda K, Mekada E, Nakano H (2003) Expression of lysophosphatidic acid receptors and vascular endothelial growth factor mediating lysophosphatidic acid in the development of human ovarian cancer. Cancer Lett 192:161–169
Fukui R, Tanabe E, Kitayoshi M, Yoshikawa K, Fukushima N, Tsujiuchi T (2012) Negative regulation of cell motile and invasive activities by lysophosphatidic acid receptor-3 in colon cancer HCT116 cells. Tumor Biol 33:1899–1905
Fukushima K, Takahashi K, Yamasaki E, Onishi Y, Fukushima N, Honoki K, Tsujiuchi T (2017) Lysophosphatidic acid signaling via LPA1 and LPA3 regulates cellular functions during tumor progression in pancreatic cancer cells. Exp Cell Res 352:139–145
Furui T, LaPushin R, Mao M, Khan H, Watt SR, Watt MAV, Lu Y, Fang X, Tsutsui S, Siddik ZH, Bast RC Jr, Mills GB (1999) Overexpression of Edg-2/vzg-1 induces apoptosis and anoikis in ovarian cancer cells in a lysophosphatidic acid-independent manner. Clin Cancer Res 5:4308–4318
Furuta D, Yamane M, Tsujiuchi T, Moriyama R, Fukushima N (2012) Lysophosphatidic acid induces neurite branch formation through LPA3. Mol Cell Neurosci 50:21–34
Geraldo LHM, Spohr TCLS, Amaral RFD, Fonseca ACCD, Garcia C, Mendes FA, Freitas C, dosSantos MH, Lima FRS (2021) Role of lysophosphatidic acid and its receptors in health and disease: novel therapeutic strategies. Signal Trandsuct Target Ther 6:45
Gerrard JM, Kingdom SE, Peterson DA, Peller J, Krantz KE, White JG (1979) Lysophosphatidic acids. Influence on platelet aggregation and intracellular calcium. Am J Pathol 96:423–438
Hamilton G, Rath BA (2014) A short update on cancer chemoresistance. Wine Med Wochenschr 164:456–460
Hayashi M, Okabe K, Kato K, Okumura M, Fukui R, Fukushima N, Tsujiuchi T (2012) Differential function of lysophosphatidic acid receptors in cell proliferation and migration of neuroblastoma cells. Cancer Lett 326:91–96
Hermann PC, Huber SL, Herrler T, Aicher A, Ellwart JW, Guba M, Bruns CJ, Heeschen C (2007) Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer. Cell Stem Cell 1:313–323
Honoki K, Fujii H, Kubo A, Kido A, Mori T, Tanaka Y, Tsujiuchi T (2010) Possible involvement of stem-like populations with elevated ALDH1 in sarcomas for chemotherapeutic drug resistance. Oncol Rep 24:501–505
Hu Y-L, Tee M-K, Goetzl EJ, Auersperg N, Mills GB, Ferrara N, Jaffe RB (2001) Lysophosphatidic acid induction of vascular endothelial growth factor expression in human ovarian cancer cells. J Natl Cancer Inst 93:762–768
Huang RX, Zhou PK (2020) DNA damage response signaling pathways and target for radiotherapy sensitization in cancer. Signal Transduct Target Ther 5:60
Huang W-H, Chang M-C, Tsai K-S, Hung M-C, Chen H-L, Hung S-C (2013) Mesenchymal stem cells promote growth and angiogenesis of tumors in mice. Oncogene 32:4343–4354
Insel PA, Zhang L, Murray F, Yokouchi H, Zambon AC (2012) Cyclic AMP is both a pro-apoptotic and anti-apoptotic second messenger. Acta Physiol 204:277–287
Ishii S, Noguchi K, Yanagida K (2009) Non-edg family lysophsphatidic acid (LPA) receptors. Prostaglandins Other Lipid Mediat 89:57–65
Ishii S, Hirane M, Fukushima K, Tomimatsu A, Fukushima N, Tsujiuchi T (2015) Diverse effects of LPA4, LPA5 and LPA6 on the activation of tumor progression in pancreatic cancer cells. Biochem Biophys Res Commun 461:59–64
Jeon ES, Moon HJ, Lee MJ, Song HY, Kim YM, Cho M, Suh D-S, Yoon M-S, Chang CL, Jung JS, Kim JH (2008) Cancer-derived lysophosphatidic acid stimulates differentiation of human mesenchymal stem cells to myofibroblast-like cells. Stem Cells 26:789–797
Jeon ES, Heo SC, Lee IH, Choi YJ, Park JH, Choi KU, Park DY, Suh DS, Yoon MS, Kim JH (2010) Ovarian cancer-derived lysophosphatidic acid stimulates secretion of VEGF and stromal cell-derived factor-1α from human mesenchymal stem cells. Exp Mol Med 42:280–293
Jinno N, Yoshida M, Hayshi K, Naitho I, Hori Y, Natsume M, Kato A, Kachi K, Asano G, Atsuta N, Sahashi H, Kataoka H (2021) Autotaxin in ascites projotes peritoneal dissemination in pancreatic cancer. Cancer Sci 112:668–678
Kathawala RJ, Gupta P, Ashby CR Jr, Chen Z-S (2015) The modulation of ABC transporter-mediated multidrug resistance in cancer: a review of the past decade. Drug Resist Updat 18:1–17
Kato K, Fukui R, Okabe K, Tanabe E, Kitayoshi M, Fukushima N, Tsujiuchi T (2012) Constitutively active lysophosphatidic acid receptor-1 enhances the induction of matrix metalloptoteinase-2. Biochem Biophys Res Commun 417:790–793
Kessenbrock K, PlaksV, Werb Z (2010) Matrix metalloproteinases: regulators of the tumor microenvironment. Cell 141:52–67
Kitayama J, Shida D, Sako A, Ishikawa M, Hama K, Aoki J, Arai H, Nagawa H (2004) Over-expression of lysophosphatidic acid receptor-2 in human invasive ductal carcinoma. Breast Cancer Res 6:R640–R646
Komachi M, Tomura H, Malchinkhuu E, Tobo M, Mogi C, Yamada T, Kimura T, Kuwabara A, Ohta H, Im D-S, Kurose H, Takeyoshi I, Sato K, Okajima F (2009) LPA1 receptors mediate stimulation, whereas LPA2 receptors mediate inhibition, of migration of pancreatic cancer cells in response to lysophosphatidic acid and malignant ascites. Carcinogenesis 30:457–465
Kurisu R, Takamoto M, Minami K, Ueda N, Yamada M, Shima N, Otani T, Sakai Y, Kondo D, Tsujiuchi T (2022) Effects of lysophosphatidic acid (LPA) signaling via LPA receptors on cellular functions associated with ATP reduction in osteosarcoma cells treated with ethidium bromide. J Bioenerg Biomemr 54:109–117
Leblanc R, Peyruchaud O (2015) New insights into the autotaxin/LPA axis in cancer development and metastasis. Exp Cell Res 333:183–189
Leve F, Peres-Moreira RJ, Binato R, Abdelhey E, Morgado-Díaz JA (2018) LPA induces colon cancer cell proliferation through a cooperation between the ROCK and STAT-3 pathways. PLoS ONE 10:e0139094
Lin M-E, Herr DR, Chun J (2010) Lysophosphatidic acid (LPA) receptors: signaling properties and disease relevance. Prostaglandins Other Lipid Mediat 91:130–138
Minami K, Ueda N, Maeda H, Ishimoto K, Otagaki S, Tsujiuchi T (2019) Modulation of chemoresistance by lysophosphatidic acid (LPA) signaling through LPA5 in melanoma cells treated with anticancer drugs. Biochem Biophys Res Commun 517:359–363
Minami K, Ueda N, Ishimoto K, Tsujiuchi T (2020a) LPA5-mediated signaling induced by endothelial cells and anticancer drug regulates cellular functions of osteosarcoma cells. Exp Cell Res 388:111813
Minami K, Ueda N, Ishimoto K, Kurisu R, Takamoto M, Ikeda H, Tsujiuchi T (2020b) Cooperation of G12/13 and Gi proteins via lysophosphatidic acid receptor-2 (LPA2) signaling enhances cancer cells survival to cisplatin. Biochem Biophys Res Commun 542:427–432
Minami K, Ueda U, Ishimoto K, Kurisu R, Takamoto M, Ikeda H, Tsujiuchi T (2021) Roles of endothelial cells in the regulation of cell motility via lysophosphatidic acid receptor-2 (LPA2) and LPA3 in osteosarcoma cells. Exp Mol Pathol 118:104596
Mokady D, Meiri D (2015) RhoGTPases - A novel link between cytoskeleton organization and cisplatin resistance. Drug Resist Updat 19:22–32
Morrison R, Schleicher SM, Sun Y, Niermann KJ, Kim S, Sparatt DE, Chung CH, Lu B (2011) Targeting the mechanisms of resistance to chemotherapy and radiotherapy with the cancer stem cell hypothesis. J Oncol 2011:941876
Obo Y, Yamada T, Furukawa M, Hotta M, Honoki K, Fukushima N, Tsujiuchi T (2009) Frequent mutations of lysophosphatidic acid receptor-1 gene in rat liver tumors. Mutat Res 660:47–50
Okabe K, Hayashi M, Fujii M, Honoki K, Mori T, Fukushima N, Tsujiuchi T (2010) Mutations of lysophosphatidic acid receptor genes in human osteosarcoma cells. Pathobiology 77:278–282
Okabe K, Hayashi M, Yamawaki Y, Teranishi M, Honoki K, Mori T, Fukushima N, Tsujiuchi T (2011) Possible involvement of lysophosphatidic acid receptor-5 gene in the acquisition of growth advantage of rat tumor cells. Mol Carcinog 50:635–642
Okabe K, Hayashi M, Kato K, Okumura M, Fukui R, Honoki K, Fukushima N, Tsujiuchi T (2013) Lysophosphatidic acid receptor-3 increases tumorigenicity and aggressiveness of rat hepatoma RH7777 cells. Mol Carcinog 52:247–254
Okuda A, Takai M, Kurisu R, Takamoto M, Ikeda H, Tsujiuchi T (2023) Roles of lysophosphatidic acid (LPA) receptor-2 (LPA2) in the regulation of cellular responses induced by X-ray irradiation and hydrogen peroxide in pancreatic cancer cells. Int J Radiat Biol in press
Pasternack SM, von Kügelgen I, Aboud KA, Lee YA, Rüschendorf F, Voss K, Hillmer AM, Molderings GJ, Franz T, Ramirez A, Nürnberg P, Nöthen MM, Betz RC (2008) G protein-coupled receptor P2Y5 and its ligand LPA are involved in maintenance of human hair growth. Nat Genet 40:329–334
Ptaszynska MM, Pendrak ML, Stracke ML, Roberts DD (2010) Autotaxin signaling via lysophosphatidic acid receptors contributes to vascular endothelial growth factor-induced endothelial cell migration. Mol Cancer Res 8:309–321
Samadi N, Bekele R, Capatos D, Venkatraman G, Sariahmetoglu M, Brindley DN (2011) Regulation of lysophosphatidate signaling by autotaxin and lipid phosphate phosphatases with respect to tumor progression, angiogenesis, metastasis and chemo-resistance. Biochimie 93:61–70
Sano T, Baker D, Virag T, Wada A, Yatomi Y, Kobayashi T, Igarashi Y, Tigyi G (2002) Multiple mechanisms linked to platelet activation result in lysophosphatidic acid and sphingosine 1-phosphate generation in blood. J Biol Chem 277:21197–21206
Sau A, Tregno FP, Valentino F, Federici G, Caccuri AM (2010) Glutathione transferases and development of new principles to overcome drug resistance. Arch Biochem Biophys 500:116–122
Schulte KM, Beyer A, Köhrer K, Oberhäuser S, Röher HD (2001) Lysophosphatidic acid, a novel lipid growth factor for human thyroid cells: over-expression of the high-affinity receptor edg4 in differentiated thyroid cancer. Int J Cancer 92:249–256
Seo EJ, Kwon YW, Jang IH, Kim DK, Lee SI, Choi EJ, Kim K-H, Suh D-S, Lee JH, Chou KU, Lee JW, Mok HJ, Kim KP, Matsumoto H, Aoki J, Kim JH (2016) Autotaxin regulates maintenance of ovarian cancer stem cells through lysophosphatidic acid-mediated autocrine mechanism. Stem Cells 34:551–564
Shida D, Kitayama J, Yanaguchi H, Okaji Y, Tsuno NH, Watanabe T, Takuwa Y, Nagawa H (2003) Lysophosphatidic acid (LPA) enhances the metastatic potential of human colon carcinoma DLD1 cells through LPA1. Cancer Res 63:1706–1711
Shida D, Kitayama J, Yamaguchi H, Hama K, Aoki J, Arai H, Yamashita H, Mori K, Sako A, Konishi T, Watanabe T, Sakai T, Suzuki R, Ohta H, Takuwa Y, Nagawa H (2004a) Dual mode regulation of migration by lysophosphatidic acid in human gastric cancer cells. Exp Cell Res 301:168–178
Shida D, Watanabe T, Aoki J, Hama K, Kitayama J, Sonoda H, Kishi Y, Yamaguchi H, Sasaki S, Sako A, Konishi T, Arai H, Nagawa H (2004b) Aberrant expression of lysophosphatidic acid (LPA) receptors in human colorectal cancer. Lab Invest 84:1352–1362
Shimomura Y, Garzon MC, Kristal L, Shapiro L, Christiano AM (2009) Autosomal recessive wooly hair with hypotrichosis caused by a novel homozygous mutation in the P2RY5 gene. Exp Dermatol 18:218–221
Steegborn C (2014) Structure, mechanisms, and regulation of soluble adenylyl cyclases – similarities and differences to transmembrane adenylyl cyclases. Biochim Biophys Acta 1842:2535–2547
Stoddard NC, Chun J (2015) Promising pharmacological directions in the world of lysophosphatidic acid signaling. Biomol Ther 23:1–11
Stracke ML, Krutzsch HC, Unsworth EJ, Arestad A, Cioce V, Schiffmann E, Liotta LA (1992) Identification, purification, and partial sequence analysis of autotaxin, a novel motility-stimulating protein. J Biol Chem 267:2524–2529
Takagi S, Sasaki Y, Koike S, Takemoto A, Seto Y, Haraguchi M, Ukaji T, Kawaguchi T, Sugawara M, Saito M, Funauchi Y, Ae K, Matcumoto S, Fujita N, Katayama R (2021) Platelet-derived lysophosphatidic acid mediated LPAR1 activation as a therapeutic target for osteosarcoma metastasis. Oncogene 40:5548–5558
Takahashi K, Fukushima K, Onishi Y, Fukushima N, Honoki K, Tsujiuchi T (2017a) Lysophosphatidic acid (LPA) signaling via LPA4 and LPA6 negatively regulates cell motile activities of colon cancer cells. Biochem Biophys Res Commun 483:652–657
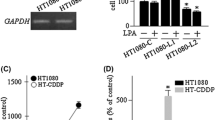
Takahashi K, Fukushima K, Fukushima N, Honoki K, Tsujiuchi T (2017b) Enhanced cellular functions through induction of LPA2 by cisplatin in fibrosarcoma HT1080 cells. Mol Cell Biochem 431:29–35
Takahashi K, Fukushima K, Otagaki S, Ishimoto K, Minami K, Fukushima N, Honoki K, Tsujiuchi T (2018a) Effects of LPA1 and LPA6 on the regulation of colony formation activity in colon cancer cells treated with anticancer drugs. J Recept Signal Transduct Res 38:71–75
Takahashi K, Fukushima K, Tanaka K, Minami K, Ishimoto K, Otagaki S, Fukushima N, Honoki K, Tsujiuchi T (2018b) Involvement of LPA signaling via LPA receptor-2 in the promotion of malignant properties in osteosarcoma cells. Exp Cell Res 369:316–324
Takai M, Takauchi M, Kuribayashi M, Tsujiuchi T (2023) LPA receptor-mediated signaling regulates cell motility and survival to anticancer drug of pancreatic cancer cells under glucose-deprived and hypoxic conditions. Biochem Biophys Res Commun 661:21–27
Tanabe E, Kitayoshi M, Hirane M, Araki M, Dong Y, Fukushima N, Tsujiuchi T (2013) Downregulation of activation factors of endothelia and fibroblasts via lysophosphatidic acid signaling in a mouse lung cancer LL/2 cell line. J Recept Signal Transduct Res 33:286–290
Tokumura A, Fukuzawa K, Akamatsu Y, Yamada S, Suzuki T, Tsukatani H (1978) Identification of vasopressor phospholipid in crude soybean lecithin. Lipids 13:468–472
Tsujino M, Fujii M, Okabe K, Mori T, Fukushima N, Tsujiuchi T (2010) Differential expressions and DNA methylation patterns of lysophosphatidic acid receptor genes in human colon cancer cells. Virchows Arch 457:669–676
Tsujiuchi T, Araki M, Hirane M, Dong Y, Fukushima N (2014) Lysophosphatidic acid receptors in cancer pathobiology. Histol Histopathol 29:313–321
Tsukamoto S, Honoki K, Fujii H, Tohma Y, Kido A, Mori T, Tsujiuchi T, Tanaka Y (2012) Mesenchymal stem cells promote tumor engraftment and metastatic colonization in rat osteosarcoma model. Int J Oncol 40:163–169
Ueda N, Minami K, Ishimoto K, Tsujiuchi T (2020) Effects of lysophosphatidic acid (LPA) receptor-2 (LPA2) and LPA3 on the regulation of chemoresistance to anticancer drug in lung cancer cells. Cell Signal 69:109551
van Meeteren LA, Moolenaar WH (2007) Regulation and biological activities of the autotaxin-LPA axis. Prog Lipid Res 46:145–160
Visvader JE, Lindeman GJ (2008) Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat Rev Cancer 8:755–768
Xu Y, Gaudette DC, Boynton JD, Frankel A, Fang X-J, Sharme A, Hurteau J, Casey G, Goodbody A, Mellors A, Holub BJ, Mills B (1995) Characterization of an ovarian cancer activating factor in ascites from ovarian cancer patients. Clin Cancer Res 1:1223–1232
Yamada T, Furukawa M, Hotta M, Yamasaki A, Honoki K, Fukushima N, Tsujiuchi T (2009) Mutations of lysophosphatidic acid receptor-1 gene during progression of lung tumors in rats. Biochem Biophys Res Commun 378:424–427
Yu S, Murph MM, Lu Y, Liu S, Hall HS, Liu J, Stephens C, Fang X, Mills GB (2008) Lysophosphatidic acid receptors determine tumorigenicity and aggressiveness of ovarian cancer cells. J Natl Cancer Inst 100:1630–1642
Yung YC, Stoddard N, Chun J (2014) LPA receptor signaling: pharmacology, physiology, and pathophysiology. J Lipid Res 55:1192–1214
Zhang H, Kong Q, Wang J, Jiang Y, Hua H (2020) Complex roles of cAMP-PKA-CREB signaling in cancer. Exp Hematol Oncol 9:32
Acknowledgements
This work was supported by JSPS KAKENHI Grant Numbers JP18K07249, 21K07109 (to T.T.) and 20591765, 15K10455 (to K.H.).
Author information
Authors and Affiliations
Contributions
All authors contributed to writing and editing the manuscript. All authors have approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Compliance with ethical standards
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Takai, M., Mori, S., Honoki, K. et al. Roles of lysophosphatidic acid (LPA) receptor-mediated signaling in cancer cell biology. J Bioenerg Biomembr 56, 475–482 (2024). https://doi.org/10.1007/s10863-024-10028-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10863-024-10028-9