
Abstract
Tropical dry forests are among the most diverse and threatened habitats in the world, yet they are rarely protected and remain poorly studied. In Brazil, dry forests are naturally fragmented and embedded within various biomes, thus making it important to assess biotic homogeneity among geographically separated forest fragments. We sampled 7732 individuals belonging to 48 species to quantify the diversity of fruit-feeding butterfly communities at four Brazilian dry forest sites, and found differences in community structure between northern and central sites. Species richness per plot was the same in both areas, but abundance per plot was higher in northern sites. Species composition differed between sites mostly due to species of Satyrinae. Additive partitioning showed that beta diversity corresponded to 70.1% of all diversity. Rather than species loss, beta diversity primarily represented species turnover that was potentially driven by differences in the surrounding habitats. Butterfly community composition and abundance were influenced by vegetation where abundance increased with tree density and basal area, and decreased with the average tree height. Butterfly species richness and abundance were higher in the wet season than in the dry season, and all species sampled in the dry season were present in the wet season. Differences in community structure across relatively short geographic distances in the same type of habitat highlight the importance of conserving tropical dry forest fragments to ensure the maintenance of butterfly diversity and, presumably, other insects.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Neotropical dry forests have a patchy distribution from northern Argentina to northeast Brazil, along the Pacific slope of the Andes, northward along the coast of Central America to northwest Mexico (Pennington et al. 2000; Särkinen et al. 2011; Sánchez-Azofeifa et al. 2014) and are the least known of all tropical forest types (Banda-R et al. 2016). These forests are characterized by a pronounced dry season of 5–6 month duration when > 50% of the trees lose their leaves (Särkinen et al. 2011). Each forest may have a distinctive plant species composition (Neves et al. 2015), and high β diversity (Banda et al. 2016). Tropical dry forests (hereafter TDFs) are among the most threatened of all Neotropical ecosystems because they occur in areas with a long history of continuous human settlement and agriculture (Espírito-Santo et al. 2009; Sánchez-Azofeifa et al. 2014; Banda-R et al. 2016). When no longer used for agriculture or cattle ranching, TDF areas are usually abandoned and a secondary succession commences. Thus, natural regrowth of TDF produces a mosaic of secondary fragments within undisturbed areas (Quesada et al. 2009; Neves et al. 2010; Sánchez-Azoifeifa et al. 2014).
In South America, the Pleistocene Arc Hypothesis (Prado and Gibbs 1993; Pennington et al. 2000) attempts to explain the disjunct distribution of TDFs and also differences in their vegetation. This hypothesis suggests that TDFs had a continuous range in South America before the Pleistocene glaciation, including areas that are now rainforests and cerrado. The end of this cool and dry period led to an expansion of wet forests, and TDFs persisted in small fragments within different biomes (Prado and Gibbs 1993). Therefore, geographical isolation and neighboring vegetation likely played a crucial role in the ecological history of different TDFs.
Although TDFs occur in all Brazilian biomes where local climate, soil and topographic conditions permit (Pennington et al. 2009), they occupy only 3.21% (27,367,815 ha) of Brazil’s land area (Espírito-Santo et al. 2009). In northern Minas Gerais TDFs are found mostly in the cerrado-caatinga transition zone (Espírito-Santo et al. 2009) while in central Minas Gerais they occur as small forest fragments on limestone soils (Coelho et al. 2012, 2014).
Butterflies have played an important role in refining our understanding of ecological and evolutionary patterns and processes (DeVries and Walla 2001; Bonebrake et al. 2010; DeVries et al. 2011; Valtonen et al. 2013; Espeland et al. 2015; Penz et al. 2015; Fordyce and DeVries 2016). Explanations for butterfly distribution patterns generally center on the abiotic conditions that affect vegetation (e.g., Didham et al. 2007; Basset et al. 2012). For example, temperature, humidity and rainfall correlate positively with butterfly diversity (Turner et al. 1987; Luoto et al. 2006; but see; Robinson et al. 2012; Komac et al. 2013), and these climatic factors in turn influence habitat type (Robinson et al. 2012; Carneiro et al. 2014). In temperate regions habitat heterogeneity has been associated with butterfly species diversity (Fartmann et al. 2013), presumably because greater heterogeneity can provide refuges from adverse environmental conditions, thus allowing more species to co-exist (Stein et al. 2014). At broad geographical scales, the surrounding landscape can influence diversity patterns by imposing spatial variation in vegetation structure and ultimately generating landscape heterogeneity (Komac et al. 2013; Marques and Schoereder 2013; Ribeiro et al. 2012). In addition, the variation in larval and adult resources, vegetation structure, and light levels within particular habitats are factors that likely influence local butterfly species diversity and local distributions (DeVries and Walla 2001; Öckinger et al. 2009).
Tropical insect diversity is known to differ between wet and dry seasons (Bigger 1976; Wolda 1978; Ribeiro et al. 2010). Seasonal variation in species abundances is thought to arise from environmental fluctuations such as changes in day length (Wolda 1989; Brito et al. 2014), population dynamics, and migration (Torres et al. 2009). Seasonal fluctuations in species diversity may be mediated by resource availability (e.g., Wardhaugh 2014), rainfall, and temperature, and the influence these factors have on diversity can vary depending on locality (Grøtan et al. 2012, 2014).
Fruit-feeding nymphalids form a guild of adult butterflies that utilize the juices of rotting fruit as their major food resource (DeVries et al. 1997; Freitas et al. 2014), and they comprise a significant proportion of local nymphalid species richness (DeVries 1987; DeVries et al. 1997; Brown Jr. and; Freitas 2000; Horner-Devine et al. 2003; Ribeiro et al. 2010). Fruit-feeding nymphalids are easily sampled with a standardized trap protocol (see DeVries et al. 2016), they respond to environmental changes in space and time (DeVries et al. 1997, 1999; DeVries and Walla 2001; Ribeiro et al. 2010; Grøtan et al. 2012, 2014; Freitas et al. 2014), and can be used to assess variation in species diversity within and among forest types (e.g., DeVries et al. 2011; Fordyce and DeVries 2016).
This study uses species composition, richness and abundance distributions to compare spatial and seasonal patterns of fruit-feeding nymphalids among four TDF sites in the state of Minas Gerais: three northern sites within a cerrado-caatinga transition zone, and one in the central area of the state that is embedded within the cerrado sensu strictu. Given the marked differences in Brazilian TDFs (e.g., Espírito-Santo et al. 2009; Neves et al. 2015), we sought to answer the following questions: (i) Does geographical location influence community diversity? (ii) Given that the northern and central TDF sites are surrounded by different vegetation types, do they differ in β diversity? (iii) Does local vegetation structure affect butterfly diversity? (iv) Does species diversity vary significantly among seasons? We then briefly summarize what is known about the ecology of TDF butterflies, and place our study in the context of what future role butterflies can play in the conservation of Brazilian TDFs.
Materials and methods
Study area
Fieldwork was conducted during March and September 2012 and February and September of 2013 in four conservation areas (sites hereafter, Fig. 1). Three sites were located in northern Minas Gerais in a transitional zone between the cerrado and caatinga (Lagoa do Cajueiro State park, 20,500 ha; Serra Azul Biological Reserve, 3841 ha; Jaíba Biological Reserve, 6358 ha—separated from each other by at least 15 Km) and one central site was located within Cerrado and rocky grassland vegetation (Serra do Cipó, 31,450 ha).

Location of the four dry forest sites in Minas Gerais, Brazil. In the transition area between Caatinga and Cerrado, northern sites Lagoa do Cajueiro State Park, Jaíba Biological Reserve, and Serra Azul Biological Reserve. In the Cerrado, the central site: Serra do Cipó
Annual average rainfall in the northern TDFs is approximately 818 ± 242 mm (Madeira et al. 2009), with less than 60 mm in the marked dry season from May to October (Espírito-Santo et al. 2008). The vegetation is highly deciduous, with ca. 95% leaf loss (Neves et al. 2015), and an annual average temperature of 24 °C (Madeira et al. 2009). The TDFs of Serra do Cipó are patchily distributed and found primarily in isolated enclaves of limestone that support tree-sized vegetation, and the surrounding areas have a distinct floristic composition (Coelho et al. 2012, 2014). The climate is mesothermic with dry winters and rainy summers, and an annual average rainfall of 1500 mm, and annual average temperatures ranging from 17.4 to 19.8 °C (Madeira and Fernandes 1999).
Sampling design
As TDFs potentially have unique communities (e.g., Quesada et al. 2009), the sampling design was planned accordingly. Three sampling areas were selected in the large Cajueiro State Park, and each included three plots. As they are smaller, the Jaíba and Serra Azul Biological Reserves constituted two separate areas with three plots each. Finally, the sampling at Serra do Cipó matched that done in Cajueiro, including three areas with three plots each. An initial analysis showed that the studied areas actually constituted two distinctive communities (not eight; see Supplementary Material F1 in Supplementary Information), and we therefore opted for combining these areas into two geographical units: northern and central sites.
One plot of 50 × 20 m (0.1 ha) was delimited inside each area (24 plots total), and four butterfly traps were deployed in each plot (96 traps total). Samples from the traps in each plot were combined for analyses (study sample unit = plot). In each of the 24 plots, all plants with diameter at breast height ≥ 5 cm were marked for analysis of vegetation structure (Norden et al. 2015). The measures were density, richness, mean height and basal area of all marked trees inside each plot.
Butterfly traps were baited with fermented banana and sugar cane juice. On the first day baited traps were placed with their base about 1 m above the ground, and captured butterflies were removed daily during the following 3 days. Bait was replaced daily. When possible, captured individuals were marked with permanent ink, and the species identification, sampling locality, and date were recorded in a field book, and then the butterfly was released. When field identification was not possible, individuals were sampled, stored in separate envelopes with all sampling information written on the envelope, and subsequently identified in the lab using field guides (DeVries 1987; Uehara-Prado et al. 2004). Taxonomic nomenclature follows Lamas (2004) and Heikkila et al. (2011). Sampled butterflies were deposited at the Federal University of Minas Gerais.
Statistical analysis
Species richness at each site was estimated using extrapolation and rarefaction curves based on the Chao1 estimator, and configured at 40 knots and 300 bootstraps to determine confidence intervals (Chao and Jost 2012; Carneiro et al. 2014). This method compares richness at equal sample coverage to produce a less biased measure of community richness than traditional rarefaction (Chao and Jost 2012). Analyses were done using iNEXT (Hsieh et al. 2013), available from https://chao.shinyapps.io/iNEXTOnline/.
Comparisons of species composition among sites were done using non-metric multidimensional scaling (NMDS) based on a Bray–Curtis dissimilarity matrix. This analysis was followed by a permutational multivariate analysis of variance (PERMANOVA; Anderson 2001). Another PERMANOVA was done to test the effect of vegetation structure (tree density, richness, mean height and basal area—independent variables) on species composition (species identity as dependent variable). The NMDS and PERMANOVA were performed using functions from the MASS (Venables and Ripley 2002) and vegan (Oksanen et al. 2017) packages in R v.3.0.2 (R Core Team 2014).
Generalized linear models (GLMs) were built to test if butterfly abundance and richness per plot (response variables) differed by sites, vegetation structure (tree density, richness, mean height and basal area), and their interaction (explanatory variables). All GLMs were submitted to residual analysis to evaluate adequacy of the error distribution (Crawley 2013). Minimum adequate models were generated by stepwise omission of non-significant terms.
Since our TDFs were fragmented and occurred in different habitats, we used additive partitioning of diversity to help understand species composition patterns at multiple biogeographical scales. This allowed identification of the regional scale that contributes most to the local species pool (Lande 1996), which is valuable when examining variation in community composition in fragmented areas (Si et al. 2015). To assess the contribution of each sampling level to total diversity, additive partitions of the data were made with four degrees of diversity: (α1) diversity within plot, (β1) diversity between plots within sites, (β2) diversity between sites within regions, and (β3) diversity between regions (northern vs. central regions). A comparison between the observed and expected diversity for α and β components (individual based randomization, with 1000 simulations) was considered different when p < 0.05. This analysis was performed using the vegan package (Oksanen et al. 2017).
To test whether dissimilarities among multiple plots (β diversity) were due to nestedness (species gains or losses) or turnover components (species replacement among plots), a model using the Sorensen dissimilarity index was tested with the betapart package (Baselga and Orme 2012). The Sorensen dissimilarity index is widely used due to its dependence on the proportion of species shared between two communities (Baselga 2010), and produces three indices: Simpson dissimilarity (turnover component), Sorensen dissimilarity (the total beta diversity), and the nested-resultant fraction of Sorensen dissimilarity (nestedness component).
A comparison of seasonal composition was done using NMDS ordination based on a Bray–Curtis dissimilarity matrix, on data for each sampling plot per season (wet and dry). To test for differences in species composition between seasons, a PERMANOVA (Anderson 2001) was performed on the Bray–Curtis dissimilarity matrix. For temporal analysis, linear mixed effects models were constructed using richness and abundance of each plot (24 plots—dependent variables) in each season per year (2012 and 2013 independent variable) with the plot as the random factor. The package lme4 was used to perform the analysis (Bates et al. 2015). All statistical analyses were performed in R v.3.0.2 (R Core Team 2014).
Results
Species diversity patterns
We sampled 7,732 individual butterflies comprising 48 species in four subfamilies (Biblidinae, Charaxinae, Nymphalinae, Satyrinae; see Supplementary Material T1 in Supplementary Information). Twenty-eight species were sampled in northern sites (28 in Cajueiro, 18 in Jaíba, and 16 in Serra Azul), and 39 at the central site Serra do Cipó. Of the total sampled species, 20 were found only in the central site, and nine species were found only in the northern sites. Of the total sample abundances, eight were singletons, three were doubletons (see Supplementary Material T1), and 14 species of Biblidinae accounted for 89% (N = 6889) of all individuals. Eunica tatila represented 57% of the total individuals in Biblidinae. This species was sampled only in the northern sites, and was abundant during all sampling periods (66 ± 48 individuals per site per sampling period). Hamadryas februa was also abundant in the northern sites, comprising of 37% of the Biblidinae, but only three individuals were sampled in the central sites. Members of the Satyrinae accounted for 48% of the total species richness, and were sampled mostly in the central sites—20 out of 23 species, including ten found only in the central site.
Rarefaction-extrapolation based on species accumulation curves suggest high coverage for each site (Fig. 2a) with a total richness of 59 species, with 99% sample coverage, and corresponding to 81% complete sampling at the regional scale (Fig. 2b). Based on these curves, the expected number of species were: 44 for Lagoa do Cajueiro (28 sampled), 22 in Jaíba (18 sampled), and 21 in Serra Azul (16 sampled), plus 47 for the central site Serra do Cipó (39 sampled). Although the total richness in Serra do Cipó was higher than in northern sites (39 vs. 28), the GLM indicated that species richness did not differ among sites (14 ± 4 species per plot; p = 0.20, Quasipoisson distribution). Abundance at Serra do Cipó was lower than in the northern sites (56 ± 25, and 482.00 ± 188 individuals per plot, p < 0.01, Gaussian distribution), and species composition differed between Serra do Cipó and the northern sites (F = 11.69; R2 = 0.64; p < 0.001; Fig. 3).

Rarefaction-extrapolation species accumulation curves of butterfly richness between different dry forest sites in Minas Gerais: a total sampling. b Sampling areas. Color code: black = Lagoa do Cajueiro; dark blue = Jaíba; green = Serra Azul; red = Serra do Cipó

NMDS plot showing similarity of butterfly species composition per site. Symbols: circle = Lagoa do Cajueiro; square = Jaíba; cross = Serra Azul; triangle = Serra do Cipó. The polygon marks Serra do Cipó, which differed in diversity from the northern sites in the PERMANOVA analysis
Additive partitioning revealed significant differences between observed and expected species richness at all scales (Fig. 4). Although diversity within plots (α1) represented only 30% of the richness, it was higher than expected (p < 0.001). In contrast, the β-diversity component (β1 between plots; β2 between sites; and β3 between the northern and central regions) accounted for 70% of the species richness. The differences between plots within sites (β1 = 23%) and between sites within regions (β2 = 17%) were lower than expected (p < 0.001 and p = 0.03, respectively), but there was a higher than expected difference between regions (β3 = 30%, p < 0.001; Fig. 4). The β-diversity among multiple plots was due mostly to species turnover (βSIM = 0.84), and this suggests local species replacement rather than species loss within sites.

Observed and expected species richness across multiple sampling scales: α1 (black) = richness per plot; β1 (dark grey) = difference of richness between plots; β2 (grey) = difference of richness between sites; and β3 (light grey) = difference of richness per region (northern and central). All scales showed significant differences between observed and expected
Vegetation structure
The PERMANOVA analysis indicated that differences in butterfly community composition per plot corresponded to tree species richness, mean height and basal area, and individual site (Table 1, see Supplementary Material F1 in Supplementary Information). No vegetation structure variable explained butterfly species richness per plot (Table 2).
Tree density, mean height, basal area, individual site, and the interaction of site and basal area influenced butterfly abundance (Table 2). Abundance was positively correlated with tree density and basal area (Fig. 5a, c), but negatively correlated with tree height (Fig. 5b). Tree richness had no effect on butterfly abundance.

Butterfly abundance per plot corresponding to: a tree density, b mean tree height, and c basal area and the interaction with site. The lines in (a) and (b) represent all sites (there were no differences between them). In (c) solid line = Serra do Cipó, and dashed line = northern sites
Seasonality
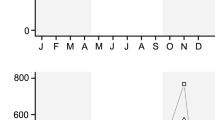
Butterfly species composition (R2 = 0.031, p = 0.013; Fig. 6a), richness (p = 0.001), and abundance (p < 0.01) varied significantly between dry and wet seasons. We found no species that occurred only in the dry season—all dry season species were also sampled in the wet season (Fig. 6b). Finally, in the wet season we sampled more species and individuals than in the dry season (Fig. 7).

NMDS plot illustrating differences in butterfly composition between dry and wet seasons. Symbols: triangles and solid line: wet season, and solid circles and dashed line : dry season. The polygons separate the dry from the wet season, which were statistically different in the PERMANOVA analysis

Significant differences between wet and dry seasons in: a butterfly richness and b abundance. Wet season samples produced 8.23 ± 0.55 species and 93.94 ± 11.75 individuals, whereas dry season samples produced 3.86 ± 0.28 species and 74.95 ± 10.15 individuals. Bars represent the standard deviation
Discussion
Surprisingly, this is the first study to compare spatial and seasonal patterns of fruit-feeding butterfly diversity among tropical dry forests in the Cerrado region. We found significant regional differences in species diversity between northern and central TDFs (Figs. 2, 3). This suggests that northern and central sites represent separate regional communities, and the high β-diversity between them (Fig. 3) reflects potential effects of surrounding habitats and their different ecological history (e.g., Dapporto et al. 2014; Figueiredo-Silva et al. 2017). The β-diversity due to species turnover implies that replacement of some species may be a consequence of environmental sorting or spatial and historical constraints (Baselga 2010). Working with South American TDF plant communities, Neves et al. (2015) found that precipitation and edaphic conditions explained most variation in species composition. Encompassing a greater scale in Latin America, the study of TDFs by Banda-R et al. (2016) found the transition caatinga-cerrado plant community to be different from that of cerrado. As observed here (Table 1), differences in vegetation structure can reflect differences in faunal composition (Didham et al. 2007; Ramos 2000).
Based mainly on taxa of Satyrinae, we found that the fauna of the central site differed from the transitional habitat communities in the northern sites (Fig. 3, see Supplementary Material T1 in Supplementary Information). Representing nearly half of the total species richness, the Satyrinae accounted for the greatest difference in species diversity between northern and central sites (see Supplementary Material T1 in Supplementary Information). Of the 23 Satyrinae species found in this study, only 7 were common to both sites (all four Pharneuptychia species, Paryphthimoides phronius, Paryphthimoides poltys and Opsiphanes invirae), and 13 species occurred only in the central site.
The species composition between the northern sites and Catimbau National Park (separated by ca. 1400 km) was more similar than the northern sites and Serra do Cipó (separated by ca. 500 km). Only 13 fruit-feeding nymphalid species are known from Catimbau (Nobre et al. 2012), and ten of these species were sampled in the northern sites. The high species overlap between Catimbau and our northern sites suggests the Caatinga fauna is derived mostly from communities that occupy the Caatinga-Cerrado transition zone.
Differences in butterfly abundance between the northern and central sites seem associated with their geographical location, and the individual abundance of two species (E. tatila and Hamadryas februa, see below). The northern sites located within a mosaic of dry forest and surrounded by agriculture had a greater abundance than the central site. Inasmuch as the central site TDF fragments are imbedded within Cerrado and limestone outcrops (Coelho et al. 2014), this may explain their lower abundance per plot. This observation is consonant with suggestions that landscape (Ribeiro et al. 2012) and vegetation structure (DeVries et al. 1999; Ramos 2000) adjacent to sampling points can influence butterfly community composition.
Eunica tatila constituted 51% of the total sampled abundance (3,939 individuals) and only occurred in the northern sites, a pattern that corresponds to a study by Gozzi et al. (2008) in a transitional area between cerrado and caatinga. Although E. tatila is known to migrate (Welling 1959), it was abundant in all sampling periods, indicating that northern sites contain larval and adult resources that supported multiple generations per year. In contrast, E. tatila is rare in some Caatinga sites (Nobre et al. 2012) and in Mexican TDFs (Pozo et al. 2008), suggesting that such resources might be limited or seasonal. Hamadryas februa accounted for ca. 33% of all individuals and was the second most abundant species (2578 individuals in the northern sites, three in the central site). The abundance of H. februa suggests a potential ecological release, where a high density of host plant might be driving local population growth (see example in DeVries et al. 1999). As the abundance of both E. tatila and H. februa could have been a response to larval resource density during our sampling period, future work is necessary to understand if there is yearly variation in their local abundance.
Several rainforest studies found vegetation structure to be a good predictor of butterfly diversity (Shahabuddin and Terborgh 1999; Ramos 2000; Barlow et al. 2007) and abundance (Ramos 2000). We found that vegetation parameters influenced butterfly species composition and abundance (Tables 1, 2). This might be due to the fact that different butterfly species occurred on plots with different vegetation structure, reflecting their microhabitat preference (e.g., light vs. shade). Butterfly abundance was lower in areas with taller trees (Fig. 4) and higher in areas with greater tree density and basal area (Table 2). Although not assessed directly, areas with greater tree density might be expected to have an increased availability of shade required by some species (e.g., Satyrinae). Finally, if TDF butterflies show vertical stratification as in rainforests (DeVries and Walla 2001; DeVries et al. 2011; Fordyce and DeVries 2016; Neves et al. 2014), we may have underestimated total butterfly diversity by not sampling potential canopy-inhabiting species in areas with tall trees.
We found no relationship between butterfly richness and tree richness or density, mean height, and basal area (Table 2; see also Barlow et al. 2008). This may be due to our using vegetation structure as a proxy for habitat complexity, and not assessing plant diversity directly. In contrast, previous work showing positive correlations between butterfly and plant diversity addressed resource-based relationships between herbivores and plants (e.g., European grasslands: Steffan-Dewenter and Tscharntke 2000; Checa et al. 2014). Given the absence of comprehensive data on plant diversity and larval host plants at our study sites, the influence of plant richness on butterfly diversity in TDFs remains to be tested.
Availability of ephemeral resources has a significant seasonal effect on tropical insect abundance (Wardhaugh 2014). As expected, TDF species diversity was highest in the wet season (Fig. 6), corroborating studies in other habitats (Shahabuddin and Terborgh 1999; Pozo et al. 2008; Nobre et al. 2012; Checa et al. 2014). Inasmuch as TDF trees may lose > 95% of their leaves during the dry season (Pezzini et al. 2014; Espírito-Santo et al. 2014; Neves et al. 2014), such seasonal change in forest structure can affect the availability of leaves needed for larval development and species that require shade for adult activities (Wardhaugh 2014).
Compared to other habitats, Brazilian TDFs are poorly studied and their entomofauna remains almost completely unknown. Only four studies have considered TDF butterflies; two in Minas Gerais (Gozzi et al. 2008; Neves et al. 2014), and two in Pernambuco (Nobre et al. 2008, 2012). The present investigation showed that surrounding habitats and seasonality have important effects on butterfly diversity, community structure and dynamics. Although all Neotropical TDFs show a similar physiognomy, they are strongly heterogeneous by virtue of their patchy distribution within various biomes, and contain endemic taxa (Banda-R et al. 2016). Our results argue that conservation efforts should not only account for the distinctiveness of TDFs, but also the integrity of surrounding habitats that influence their communities. Fruit-feeding nymphalid butterflies, similarly to ants (Figueiredo-Silva et al. 2017), seem to reflect ecological patterns of TDF plant communities by exhibiting high β-diversity resulting from species turnover.
By analyzing species diversity at multiple sites our study represents the most comprehensive ecological overview of TDF butterflies, and provides a first step toward identifying ecological attributes that drive TDF insect community dynamics. In contrast to other tropical habitats, the most basic natural history information is unknown for most TDF butterflies (larval host plants, species distributions, etc). The geographic location of TDFs and their association with surrounding habitats explain their uniqueness, and urgency for their conservation. We expect that the patterns we found for fruit-feeding butterflies are also applicable to other organisms, and it is clear that once these areas are lost, it is unlikely they could be regenerated. We feel that Brazilian TDF environments constitute exceptional opportunities to conduct comparative studies of species diversity and seasonal dynamics across broad geographical areas, thus making a substantial contribution to conservation biology.
References
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Aust Ecol 26:32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Banda-R K, Delgado-Salinas A, Dexter KG et al (2016) Plant diversity patterns in neotropical dry forests and their conservation implications. Science 353:1383–1387. doi: 10.1126/science.aaf5080
Barlow J, Gardner TA, Araujo IS et al (2007) Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. Proc Natl Acad Sci 104:18555–18560. doi: 10.1073/pnas.0703333104
Barlow J, Araujo IS, Overal WL, Gardner TA, Mendes FS, Lake IR, Peres CA (2008) Diversity and composition of fruit-feeding butterflies in tropical Eucalyptus plantations. Biodivers Conserv 17:1089–1104. doi:10.1007/s10531-007-9240-0
Baselga A (2010) Partioning the turnover and nestedness components of beta diversity. Glob Ecol Biogeogr 19:134–143. doi: 10.1111/j.1466-8238.2009.00490.x
Baselga A, Orme CDL (2012) Betapart: an R package for the study of beta diversity. Methods Ecol Evol 3:808–812. doi: 10.1111/j.2041-210X.2012.00224.x
Basset Y, Eastwood R, Sam L et al (2012) Cross-continental comparison of butterfly assemblages in tropical rainforests: implications for biological monitoring. Insect Conserv Divers 6:223–233. doi: 10.1111/j.1752-4598.2012.00205.x
Bates D, Maechler M, Bolker B, Walker S (2015) lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1-8. Accessed 31 May 2016
Bigger M (1976) Oscillations of tropical insect populations. Nature 259:207–209. doi: 10.1038/259207a0
Bonebrake TC, Ponísio LC, Boggs CL, Ehrlich PR (2010) More than just indicators: a review of tropical butterfly ecology and conservation. Biol Conserv 143:1831–1841. doi: 10.1016/j.biocon.2010.04.044
Brito MM, Ribeiro DB, Raniero M, Hasui E, Ramos FN, Arab A (2014) Functional composition and phenology of fruit-feeding-butterflies in a fragmented landscape: variation of seasonality between habitat specialists. J Insect Conserv 18:547–560. doi: 10.1007/s10841-014-9650-8
Brown KS Jr, Freitas AVL (2000) Atlantic forest butterflies: indicators for landscape conservation. Biotropica 32:934–956. doi: 10.1111/j.1744-7429.2000.tb00631.x
Carneiro E, Mielke OHH, Casagrande MM, Fiedler K (2014) Skipper richness (Hesperiidae) along elevational gradients in Brazilian Atlantic Forest. Neotrop Entomol 43:27–38. doi: 10.1007/s13744-013-0175-8
Chao A, Jost L (2012) Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology 93:2533–2547. doi: 10.1890/11-1952.1
Checa MF, Rodriguez J, Willmott KR, Liger B (2014) Microclimate variability significantly affects the composition abundance and phenology of butterfly communities in a highly threatened neotropical dry forest. Fla Entomol 97:1–13. doi: 10.1653/024.097.0101
Coelho MS, Almada ED, Quintino AV, Fernandes GW, Santos RM, Sanchez-Azofeifa A, Espírito-Santo MM (2012) Floristic composition and structure of a seasonally dry tropical forest at different successional stages in the Espinhaço Mountains, southeastern Brazil. Interciência 37:190–196
Coelho MS, Fernandes GW, Sánchez-Azofeifa A (2014) Brazilian tropical dry forest on basalt and limestone outcrops: status of knowledge and perspectives. In: Sánchez-Azofeifa A, Powers J, Fernandes GW, Quesada M (eds) Tropical dry forests in the Americas: ecology, conservation, and management. CRC Press, London, pp 55–68
Crawley MJ (2013) The R book, 2nd edn. Willey, Chichester
Dapporto L, Fattorini S, Voda R, Dinca V, Vila R (2014) Biogeography of western Mediterranean butterflies: combining turnover and nestedness components of faunal dissimilarity. J Biogeogr 41:1639–1650
DeVries PJ (1987) The butterflies of Costa Rica and their natural history. Volume I: Papilionidae, Pieridae, Nymphalidae. Princeton University Press, New Jersey
DeVries PJ, Walla TR (2001) Species diversity and community structure in Neotropical fruit-feeding butterflies. Biol J Linn Soc 74:1–15. doi: 10.1111/j.1095-8312.2001.tb01372.x
DeVries PJ, Murray D, Lande R (1997) Species diversity in vertical, horizontal and temporal dimensions of a fruit-feeding butterfly community in an Ecuadorian rainforest. Biol J Linn Soc 62:343–364. doi: 10.1111/j.1095-8312.1997.tb01630.x
DeVries PJ, Greeney HF, Walla TR (1999) Species diversity in spatial and temporal dimensions of fruit-feeding butterflies from two Ecuadorian rainforests. Biol J Linn Soc 68:333–353. doi 10.1111/j.1095-8312.1999.tb01175.x
DeVries PJ, Alexander LG, Chacon IA, Fordyce JA (2011) Similarity and difference among rainforest fruit-feeding butterfly communities in Central and South America. J Anim Ecol 81:472–482. doi: 10.1111/j.1365-2656.2011.01922.x
DeVries PJ, Hamm CA, Fordyce JA (2016) A standardized butterfly sampling protocol: fruit-feeding butterflies (Nymphalidae). In: Larsen TH (ed) Core standardized methods for rapid biological field assessment. Conservation International, Arlington, pp 139–148
Didham RK, Tylianakis JM, Gemmell NJ, Rand TA, Ewers RM (2007) Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol Evol 22:489–496. doi: 10.1016/j.tree.2007.07.001
Espeland M, Hall JPW, DeVries PJ, Lees DC, Cornwall M, Hsu Y-F, Wu L-W, Campbell D, Talavera G, Vila R, Salzman S, Ruehr S, Lohman DJ, Pierce NE (2015) Ancient neotropical origin and recent recolonisation: phylogeny, biogeography and diversification of the Riodinidae (Lepidoptera: Papilionoidea). Mol Phylogenet Evol 93:296–306. doi:10.1016/j.ympev.2015.08.006
Espírito-Santo MM, Fagundes M, Sevilha AC, Scariot A, Sánchez-Azofeifa A, Noronha SE, Fernandes GW (2008) Florestas estacionais deciduais brasileiras: distribuição e estado de conservação. MG Biota 1:5–13
Espírito-Santo MM, Sevilha AC, Anaya FC, Barbosa R, Fernandes GW, Sanchez-Azofeifa A, Scariot A, Noronha SE, Sampaio CA (2009) Sustainability of tropical dry forests: two case studies in southeastern and central Brazil. For Ecol Manage 258:922–930. doi: 10.1016/j.foreco.2009.01.022
Espírito-Santo MM, Olívio-Leite L, Neves FS, Ferreira-Nunes YR, Zazá-Borges MA, Dolabela-Falcão LA, Fonseca-Pezzini F, Louro-Berbara R, Maia-Valerio H, Fernandes GW, Reinaldo-Leite M, Santos-Clemente CM, Esdras-Leite M (2014) In: Sánchez-Azofeifa A, Powers J, Fernandes GW, Quesada M (eds) Tropical dry forests in the Americas: ecology, conservation, and management. CRC Press, London, pp 69–82
Fartmann T, Müller C, Poniatowski D (2013) Effects of coppicing on butterfly communities of woodlands. Biol Conserv 159:396–404. doi: 10.1016/j.biocon.2012.11.024
Figueiredo-Silva L, Souza RM, Solar RRC, Neves FS (2017) Ant diversity in Brazilian tropical dry forest across multiple vegetation domains. Environ Res Lett. doi:10.1088/1748-9326/aa5f2a
Fordyce JA, DeVries PJ (2016) A tale of two communities: Neotropical butterfly assemblages show higher beta diversity in the canopy compared to the understory. Oecologia 181:235. doi:10.1007/s00442-016-3562-0
Freitas ALV, Iserhard CA, Santos JP, Carreira JYO, Ribeiro DB, Melo DHA, Rosa AHB, Marini-Filho OJ, Accacio GM, Uehara-Prado M (2014) Studies with butterflies bait traps: an overview. Rev Colomb Entomol 40:209–218
Gozzi MR, Beirão MV, Medeiros LR, Neves FS, Fagundes M (2008) Borboletas frugívoras em uma região de transição entre Cerrado sensu stricto e Caatinga no norte de Minas Gerais, Brasil. MG Biota 4:25–37
Grøtan V, Lande R, Engen S, Saether BE, DeVries PJ (2012) Seasonal cycles of species diversity and similarity in a tropical butterfly community. J Anim Ecol 81:714–723. doi: 10.1111/j.1365-2656.2011.01950.x
Grøtan V, Lande R, Chacon I, DeVries PJ (2014) Seasonal cycles of species diversity and similarity in a Central American butterfly community. Ecography 37:509–516. doi: 10.1111/j.1365-2656.2011.01950.x
Heikkila M, Kaila L, Mutanen M, Peña C, Wahlberg N (2011) Cretaceous origin and repeated tertiary diversification of the redefined butterflies. Proc R Soc B Biol Sci 279:1093–1099. doi: 10.1098/rspb.2011.1430
Horner-Devine MC, Daily GC, Ehrlich PR, Boggs CL (2003) Countryside biogeography of tropical butterflies. Conserv Biol 17:168–177. doi: 10.1046/j.1523-1739.2003.01310.x
Hsieh TC, Ma KH, Chao A (2013) iNEXT online: interpolation and extrapolation (Version 1.0) [Software]. http://chao.stat.nthu.edu.tw/inext/. Accessed 14 Mar 2016
Komac B, Stefanescu C, Caritg R, Domènech M (2013) Forces driving the composition of butterfly assemblage in Andorra. J Insect Conserv 17:897–910. doi:10.1007/s10841-013-9571-y
Lamas G (2004) Checklist: part 4A. Hesperioidea-Papilionoidea. Scientific Publishers, Gainsville
Lande R (1996) Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 76:5–13. doi: 10.2307/3545743
Luoto M, Heikkinen RK, Pöyry J, Saarinen K (2006) Determinants of the biogeographical distribution of butterflies in boreal regions. J Biogeogr 33:1764–1778. doi: 10.1111/j.1365-2699.2005.01395.x
Madeira JA, Fernandes GW (1999) Reproductive phenology of sympatric taxa of Chamaecrista (Leguminosae) in Serra do Cipó, Brazil. J Trop Ecol 15:463–479
Madeira BG, Espírito-Santo MM, D’Ângelo-Neto S, Nunes YRF, Sánchez-Azofeifa A, Fernandes GW, Quesada M (2009) Changes in tree and liana communities along a successional gradient in a tropical dry forest in south-eastern Brazil. Plant Ecol 201:291–304. doi: 10.1007/s11258-009-9580-9
Marques T, Schoereder JH (2013) Ant diversity partitioning across spatial scales: ecological processes and implications for conservation in tropical dry forests. Aust Ecol 39:72–82. doi: 10.1111/aec.12046
Neves FS, Fonseca VH, Espírito-Santo MM, Vaz-de-Mello FZ, Louzada J, Sánchez-Azofeifa A, Fernandes GW (2010) Sucessional and seasonal changes in a community of dung beetles (Coleoptera: Sacarabaeinae) in Brazilian tropical dry forest. Nat Conserv 8:160–164. doi: 10.4322/natcon.00802009
Neves FS, Silva JO, Marques T, Mota-Souza JG, Madeira B, Espírito-Santo MM, Fernandes GW (2014) In: Sánchez-Azofeifa A, Powers J, Fernandes GW, Quesada M (eds) Tropical dry forests in the Americas: ecology, conservation, and management. CRC Press, London, pp 221–236
Neves DM, Dexter KG, Pennington RT, Bueno ML, Oliveira-Filho AT (2015) Environmental and historical controls of floristic compositions across the South America dry diagonal. J Biogeogr 42:1566–1576. doi: 10.1111/jbi.12529
Nobre CEB, Schlindwein C, Mielke OH (2008) The butterflies (Lepidoptera: Papilionoidea and Hesperioidea) of Catimbau National Park. Pernamb Brazil Zootaxa 1751:35–45
Nobre CEB, Ianuzzi L, Schlindwein C (2012) Seasonality of fruit-feeding butterflies (Lepidoptera, Nymphalidae) in Brazilian semiarid area. ISRN Zool. doi:10.5402/2012/268159
Norden N, Angarita HA, Bongers F, Martínez-Ramos M, Cerda IG, Breugel MV, Lebrija-Trejos E, Meave JA, Vandermeer J, Williamson GB, Finegan B, Mesquita R, Chazdon RL (2015) Successional dynamics in Neotropical forests are as uncertain as they are predictable. Proc Natl Acad Sci 112:8013–8018. doi:10.1073/pnas.1500403112
Öckinger E, Dannestam A, Smith HG (2009) The importance of fragmentation and habitat quality of urban grasslands for butterfly diversity. Landsc Urban Plan 93:31–37. doi: 10.1016/j.landurbplan.2009.05.021
Oksanen J, Blanchet FG, Kindt P, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2017) Vegan: community ecology package. R package version 2.4-2. https://CRAN.R-project.org/package=vegan. Accessed 14 Mar 2016
Pennington RT, Prado DE, Pendry CA (2000) Neotropical seasonally dry forests and quaternary vegetation changes. J Biogeogr 27:261–273. doi: 10.1046/j.1365-2699.2000.00397.x
Pennington RT, Lavin M, Oliveira-Filho AT (2009) Woody plant diversity, evolution, and ecology in the tropics: perspectives from seasonally dry tropical forests. Annu Rev Ecol Evol Syst 40:437–457. doi: 10.1146/annurev.ecolsys.110308.120327
Penz CM, Tufto J, DeVries PJ, Lande R (2015) Butterfly dispersal across Amazonia: implications for biogeography. Ecography 38:410–418. doi: 10.1111/ecog.01172
Pezzini FF, Ranieri BD, Brandão DO, Fernandes GW, Quesada M, Espírito-Santo MM, Jacobi CM (2014) Changes in tree phenology along natural regeneration in a seasonally dry tropical forest. Plant Biosyst 148:965–974. doi: 10.1080/11263504.2013.877530
Pozo C, Luis-Martínez A, Llorente-Bousquets J, Salas-Suárez N, Maya-Martínez A, Vargas-Fernández I, Warren AD (2008) Seasonality and phenology of the butterflies (Lepidoptera: Papilionoidea and Hesperioidea) of Mexico’s Calakmul region. Fla Entomol 91:407–422. doi: 10.1653/0015-4040
Prado DE, Gibbs PE (1993) Patterns of species distribution in the dry seasonal forest of South America. Ann Mo Bot Gard 80:902–927. doi: 10.2307/2399937
Quesada M, Sanchez-Azofeifa GA, Alvarez-Añorve M et al (2009) Sucessional and management of tropical dry forests in the Americas: review and new perspectives. For Ecol Manage 258:1014–1024. doi: 10.1016/j.foreco.2009.06.023
R Core Team (2014) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/. Accessed 14 Mar 2016
Ramos FA (2000) Nymphalid butterfly communities in an Amazonian forest fragment. J Res Lepidoptera 35:29–41
Ribeiro DB, Prado PI, Brown KS Jr, Freitas AVL (2010) Temporal diversity patterns and phenology in fruit-feeding butterflies in the Atlantic Forest. Biotropica 42:710–716. doi: 10.1111/j.1744-7429.2010.00648.x
Ribeiro DB, Batista R, Prado PI, Brown KS Jr, Freitas AVL (2012) The importance of small scales to the fruit-feeding butterfly assemblages in a fragmented landscape. Biodivers Conserv 21:811–827. doi:10.1007/s10531-011-0222-x
Robinson N, Armstead S, Bowers MD (2012) Butterfly community ecology: the influence of habitat type, weather patterns, and dominant species in a temperate ecosystem. Entomol Exp Appl 145:50–61. doi: 10.1111/j.1570-7458.2012.01308.x
Sánchez-Azofeifa A, Calvo-Alvarado J, Espírito-Santo MM, Fernandes GW, Powers JS, Quesada M (2014) Tropical dry forest in the Americas: The tropi-dry endeavor. In: Sánchez-Azofeifa A, Powers J, Fernandes GW, Quesada M (eds) Tropical dry forests in the Americas: ecology, conservation, and management. CRC Press, London, pp 1–16
Särkinen T, Iganci JRV, Linares-Palomino R, Simon MF, Prado DE (2011) Forgotten forests—issues and prospects in biome mapping using seasonally dry tropical forests as a case study. BMC Ecol 11:27. doi: 10.1186/1472-6785-11-27
Shahabuddin G, Terborgh JW (1999) Frugivorous butterflies in Venezuelan forest fragments: abundance, diversity and the effects of isolation. J Trop Ecol 15:703–722
Si X, Baselga A, Ding P (2015) Revealing beta-diversity patterns of breeding bird and lizard communities on inundated land-bridge islands by separating the turnover and nestedness components. PLOS ONE 10(5):e0127692. doi: 10.1371/journal.pone.0127692
Steffan-Dewenter I, Tscharntke T (2000) Butterfly community structure in fragmented habitats. Ecol Lett 3:449–456. doi: 10.1111/j.1461-0248.2000.00175.x
Stein A, Gerstner K, Kreft H (2014) Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scale. Ecol Lett 17:866–880. doi: 10.1111/ele.12277
Torres C, Osorio-Beristain M, Mariano NA, Legal L (2009) Sex-dependent seasonal feeding activity variations among two species of Nymphalidae (Lepidoptera) in the Mexican tropical dry forest. Ann Soc Entomol FR 45:265–274. doi:10.1080/00379271.2009.10697610
Turner JRG, Gatehouse CM, Corey CA (1987) Does solar energy control organic diversity? Butterflies, moths and the British climate. Oikos 48:195–205. doi: 10.2307/3565855
Uehara-Prado M, Freitas AVL, Francini RB, Brown KS Jr (2004) Guia das borboletas frugívoras da reserva estadual do Morro Grande e região de Caucaia do Alto. Cotia SP Biota Neotropica 4:1–9. doi: 10.1590/S1676-06032004000100007
Valtonen A, Molleman F, Chapman CA, Carey JR, Ayres MP, Roininen H (2013) Tropical phenology: bi-annual rhythms and interannual variation in an Afrotropical butterfly assemblage. Ecosphere 4:1–28. doi:10.1890/ES12-00338.1
Venables WN, Ripley BD (2002) Modern applied statistics with S. Springer, New York
Wardhaugh CW (2014) The spatial and temporal distributions of arthropods in forest canopies: uniting disparate patterns with hypotheses for specialization. Biol Rev 89:1021–1041. doi: 10.1111/brv.12094
Welling EC (1959) Notes on butterfly migrations in the Peninsula of Yucatan. J Lepidoterol Soc 13:62–64
Wolda H (1978) Fluctuations in abundance of tropical insects. Am Nat 112:1017–1045. doi: 10.1086/283344
Wolda H (1989) Seasonal cues in tropical organisms. Rainfall? Not necessarily! Oecologia 80:437–442. doi: 10.1007/BF00380064
Acknowledgements
The authors thank Isabela Oliveira, Fabiola Ladeira, Ana Carolina Pires, and André Aroeira for help during fieldwork; Luiz Eduardo Reis and Luiz Falcão for helping in the logistic, Geanne Pereira and Vanessa Diniz for helping with the organization of the butterflies; Eric Pereira and Gabriela Duarte for helping with the map; André Victor L Freitas and Eduardo Barbosa for helping the butterfly identification. We gratefully acknowledge the staff of the Instituto Estadual de Florestas (IEF-MG) and Instituto Chico Mendes (ICMBIO), for allowing us to stay and work in the parks, and for logistical support. We thank the grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; Grant No. 563304/2010-3), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG; Grant Nos. CRA-APQ-04738-10, CRA-APQ 00001-11) and the Inter-American Institute for Global Change Research (IAI-CRN II-021). MVB acknowledges a research scholarship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). This study was in partial fulfillment of requirements for a PhD degree to MVB at the Universidade Federal de Minas Gerais.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
10841_2017_24_MOESM1_ESM.docx
Supplementary material 1 T1 Species list and abundance of fruit-feeding butterflies sampled in dry forest habitats in Minas Gerais, Brazil. Northern sites: Lagoa do Cajueiro, Jaíba, Serra Azul. Central site: Serra do Cipó. Subfamily abundances are indicated in boldface. (DOCX 79 KB)
10841_2017_24_MOESM2_ESM.tiff
Supplementary material 2 F1 NMDS plot showing similarity of butterfly species composition per area. The clear separation of the northern and central site communities redirected subsequent analyses to a regional level (instead of a local level). (TIFF 9134 KB)
Rights and permissions
About this article

Cite this article
Beirão, M.V., Neves, F.S., Penz, C.M. et al. High butterfly beta diversity between Brazilian cerrado and cerrado–caatinga transition zones. J Insect Conserv 21, 849–860 (2017). https://doi.org/10.1007/s10841-017-0024-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-017-0024-x