
Abstract
The decision-making behavior of individuals is limited by a lack of information and a limited capacity to process information. When seeking locations to construct a hive, the honeybee Apis mellifera collectively succeeds at overcoming these individual limitations and, thus, arrives at (nearly) optimal decisions. This article examines the behaviors and coordination mechanisms that have reinforced this trait in the course of evolution. We consider what lessons might be learned concerning human behavior and how better decisions can be made.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Individuals face a variety of choices every day that vary in their complexity. In trivial everyday decisions (such as selecting consumer goods during the weekly shopping trip to a supermarket), this is a less complex process associated with minimal information gathering and modification costs. In contrast, the choices of one’s career path or life partner are often considered as important decisions with likely long-term consequences. The prerequisites for being able to make the best possible decisions are both having full information and the ability to process it. Due to the ever-growing, barely manageable flood of information available through sources such as the Internet, it has become almost impossible to extract all relevant information. This suggests that individuals can generally only be partially informed about their surroundings.
Information becomes available sequentially (i.e., information, which is relevant to a decision, has the potential, ceteris paribus, to increase over time). The result then becomes a (recurring) problem of weighing the available information: on the one hand, the decision makers can stop collecting information and make a choice. On the other hand, they can seek additional information and then stop collecting information later. The latter may increase the quality of the decision of an individual. However, the marginal utility of additional information decreases over time, while the marginal costs rise disproportionately.
This is particularly evident in medicine. For example, in emergencies where patients are in mortal danger due to some acute state, doctors are unable to take the kind of comprehensive medical history that would be necessary to determine the optimal course of treatment. If, for example, a serious traffic accident makes an immediate operation necessary to ensure the patient’s survival, the attending ER physician immediately initiates the surgery based on the diagnosis relevant to the operation at hand. In such an acute state, the ER physician is hardly going to request the patient’s entire medical record and list of prescriptions from the primary care physician.
Simon (1957) refers to the lack of information about the relevant environment and limited capacities to process the available information as bounded rationality. The human organism will therefore rely on heuristics (i.e., simple rules for making decisions) (Gigerenzer and Selten 2001; Gigerenzer et al. 2011). Contrary to the optimization assumption of rational choice theories, the human does not strive for the optimal choice, but rather the one that is satisfactory.
These limits apply not only to humans, but also to the honeybee Apis mellifera native to Central Europe. A single individual does not have the capacity for comprehensive information gathering and processing. The use of the term “colony” may be misleading, as it might imply the association of individual bees; rather, the colony functions more as a single superorganism despite the large number of individual animals (about 50,000 honeybees). Translated to humankind, one could say that the worker bees represent the body, defense, and digestive organs of this one superorganism. The queen corresponds to the female reproductive organ and the male bees (drones) are the male reproductive organs. The bees’ outstanding ability to act as a single organism requires a high degree of communication abilities. In the genus Apis mellifera, this communication is conducted via pheromones, waggle dances, and trophallaxis, the social exchange of food (Wainselboim and Farina 2000; Seeley et al. 2006; Hölldobler and Wilson 2009).
During the months of May and June, various triggers (such as the strength of the colony, the spaciousness of the hive, the supply of honey plants, and the weather) cause the division of the colony by swarming. Swarming is a vegetative reproduction process of honeybees. Approximately 70 % of the bees (a swarm) leave the colony with the queen who has regained her ability to fly. Those who stay back are worker bees and fertilized queen cells (Zeiler 1989; Tautz and Heilmann 2007; Rangel and Seeley 2012). The swarm leaves the ancestral nesting site and sets out in search of a new suitable cavity. Selecting the appropriate nest site can be regarded as essential for the swarm, because its continued existence depends largely on the quality and suitability of the new nest site.
The nest site must meet various requirements: the bees need adequate protection against rapacious predators and unfavorable weather conditions as well as sufficient space to allow further breeding and store the honey as nutriment needed in winter (Bichtler 1958; Zeiler 1989; Seeley 2014). If the results of the search are less than adequate, the colony is less likely to survive.Footnote 1 The decisions dependent to a successful search for a nest site therefore determine the honeybee colony’s survival. Therefore, one can assume that evolutionary processes (Weibull 1995; mutation mechanisms and selection mechanisms) have led the genus honeybees to develop successful mechanisms and strategies to make such decisions optimally within the collective.
The colony’s common goal consists of selecting a suitable spot for its new abode. For this purpose, the entirety of the individuals must come to a joint decision. Bees provide us with an example of mechanisms in which group decisions are not only successful, but also more successful (superadditive) than individual decisions. The colony, by cooperating with a division of labor and communication together with some competition, manages to increase the resources available. In addition, the bees are capable of correcting errors. This kind of learning is a prerequisite for a successful search for the appropriate nest site and, therefore, a crucial condition for the survival of the Apis mellifera species. Bees perform these tasks sequentially from the individual perspective, but in parallel from the collective perspective.
Against this background, we address the following research questions:
-
(1)
What challenges does the honeybeeApis mellifera face in its search for a nest site, and what mechanisms often allow not only good, but optimal decisions to be made collectively?
-
(2)
How are the decision behaviors of bees similar to and different from bounded rational individuals or the Homo oeconomicus?
-
(3)
What can human beings as individuals or as groups learn from how bees organize?
2 The nest site selection of the honeybee \({\varvec{Apis}\,\, \varvec{mellifera}}\)
2.1 Challenges
After the swarm has left the original parental hive, it sets up temporarily in a suitable place nearby from which scouts start to search for a suitable nest site (Schlegel et al. 2012). A variety of challenges must be overcome in the shortest possible time. The key challenge is not only to identify potential nest sites but also for the collective to find the optimum site. It is also important that the swarm’s decision is unanimous. If the individuals are unable to agree on a common nesting site, the swarm is under the threat of splitting. This would mean a significant loss of resources, which significantly reduces the swarm’s likelihood of survival.
The following determinants are important when deciding on a nest site (Zeiler 1989; Seeley 2014):
-
suitable size (for breeding and the storing of nutriments);
-
suitable entrance;
-
adequate protection from predators (for the bees themselves; and the stored honey); and
-
protection against weather.
The swarm has to decide relatively quickly because the swarm is only capable of carrying enough food for about three days (in the form of honey reserves).
2.2 How honeybees \({\varvec{Apis mellifera}}\) behave in the nest site selection process
The common goal of the colony is to select the most suitable nest site. Although individual differences exist among the bees, the “preferences” of the bees can, for the sake of simplicity, be considered as homogeneous (preferences, such as nest size, height above the ground, location of entrance). Conditioned by their genetic markers, the scouts evaluate the absolute quality of nest sites and do not compare them in relation to other possible nest sites.
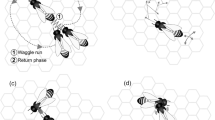
During this search, several hundred scouts fan out over a wide area to identify potentially suitable nest sites. It is noteworthy to say that the individual identity of a single scout bee does not matter because they live in large groups with well-aligned agendas. Thus, the cooperative behavior of the bee colony makes the individual relatively anonymous. If a scout has found a hive she considers appropriate, she flies back to the swarm. She then communicates her discovery to the other neutral scouts via waggle dances (Oeser 1962; Frisch 1965; Seeley 1977, 1995; Degen et al. 2015). She does not only communicate information about the distance and direction, but also her assessment of the quality of the potential nest site. The more intensely she dances, the higher the scout bee evaluates the potential nest site (Srinivasan 2011). Information about potential nest sites in varying qualities arrives at different times (sequential flow of information). By using waggle dances as a form of communication, the scouts lobby for votes in favor of the site they have located, and thus they recruite scouts that have so far remained neutral. These (so far) neutral scouts then fly to the potential nest site and perform their own independent, individual evaluations. They return to the swarm and communicate their valuations through waggle dances. The character and intensity of the dances contain the individual scouts’ evaluations of the potential nest site. However, a scout is not familiar with all of the alternatives and will not be able to follow the whole discussion in the swarm in its entirety. She is only informed of potential nest sites in her direct proximity within the swarm.
Regardless of the quality of the potential site, the intensity of the dance eventually slows down (i.e., this intensity grows weaker each time she returns from her nest site of choice) (Seeley 2014). The gradual withdrawal from the debate means that the lobbying for very suitable places lasts longer than for less good ones. If the approval ratings for a site decline (seen in the decreasing intensity of the tail wagging), this helps the scouts deal with new information. As approval ratings for alternatives drop, the likelihood of the swarm’s making a good decision increases. The evidence for a site must be maintained over a longer period. This is because information gathering is sequential, and a potentially more suitable nest site may be discovered later, perhaps because it was further away and it has taken the scouts longer to return with their findings. Discounting agreement reduces the speed of decision making and helps to avoid errors. To achieve a consensus in the decision-making process, it is essential that the support for a potential site climbs and the support for the other nest sites drops.
Consensus (agreement) among scouts comes about when no scout remains dancing (lobbying) for an alternative site. How is this state reached? The relevant criterion is the quorum (stop signal) of scouts gathering at a potential nesting site. As a rule, no debate occurs among the scouts until a full agreement can be achieved. The scouts have to strike a balance between speed and quality of their decision. Creating a complete consensus among the scouts would take a relatively large amount of time and consequently high opportunity costs (Seeley 2003; Seeley and Visscher 2004; Seeley et al. 2006, 2012). By contrast, a small quorum would allow for a quick decision, but possibly one of insufficient quality. However, when 20–30 bees are lobbying for a nest site, this is apparently enough evidence that an appropriate place has been found and that a good decision can be made. The scouts will fly back to swarm and end the other scouts’ lobbying dances (which is a pretty aggressive way for some bees to silence the others) by sending out special whistling signals. This then triggers the various preparations for the departure of the swarm as a whole (Janson et al. 2005; Greggers et al. 2013; Seeley 2014).
3 \({\varvec{Apis mellifera}}\): more bounded or unbounded rationality?
This section juxtaposes the decision behavior of the bee colony as a superorganism with that of the unbounded (Homo oeconomicus) and the bounded rational decision maker (cf. Table 1).
The honeybee’s goal is to find the best possible nest site. The quality criteria for a nest site can be assumed to be time-invariant. Rational choice theories likewise assume similarly constant criteria. Narrow rational choice approaches assume that agents soley maximize their own profit. Real decisions makers are bounded in their rationality (Simon 1957). They have limited information processing capacities and incomplete information about their relevant environment. They are usually pursuing multiple goals and relying on heuristics.Footnote 2 They strive for satisfactory alternatives and, contrary to the rational choice theories, do not generally make the optimal choice.
Simon (1957) describes the behavior of bounded rational actors as a process of adapting their aspirations. This theory was further developed by Sauermann and Selten (1962) and is briefly outlined below. Unlike neoclassicism, the theory of aspiration adaptation assumes individuals who primarily act in line with their goals. Choosing a good course of action from a set of potential alternatives requires setting goals. These goals allow us to evaluate how well various alternatives will fulfill that aspirational level. Depending on the individual level of aspiration, the consequences will be evaluated as success or failure. Aspiration levels are formed over time and dynamically adapted through successes and failures (Hoppe 1930). This led Simon (1957) to coin the concept of satisficing where the alternative is chosen that, due to external circumstances, will lead to a satisfactory result. A single bee scout acts in a bounded rational manner as an individual person who seeks to find satisfactory options.
For Apis mellifera, the task of assessing the various alternatives is distributed among a larger number of scouts. As an individual, a scout can be compared to bounded rational humans. The assessments of many scouts, who are bounded rational as individuals, allow the superorganism of the bee colony as a collective to select a near-optimal nesting site. Each individual rates the nest site she has deemed as appropriate. By lobbying (advertising in favor of one nest site), neutral scouts are recruited and will also visit the potential nest site.
For bounded rational individuals (in the tradition of Herbert Simon), the order in which choices are made available may be significant. Let us assume that there are two different options namely A and B. The individual experiences each of these two options sequentially. If the individual is first informed of A and has already evaluated this alternative as satisfactory, this option may have already been chosen. If the individual first learns of the B option, this option, too, may be chosen if it is assessed as satisfactory. This helps to explain inconsistencies in individual decisions (Simon 1983).
Each individual in a bee colony is comparable to a human individual. Both species possess only incomplete information about their environments and a limited capacity to process that information. Bees look for potential nesting sites. They aim at the survival of the collective. Once a scout has found one, she lobbies for it with the help of waggle dances (Tautz et al. 1996). However, many scout bees are exploring and evaluating various nest sites, and thus, the information is significantly improved. The decision calculus that brings about an optimal decision includes the trade-off, which also occurs in human groups, between obtaining additional information (quality) and speed of the decision. This decision-making process has been gradually improved in the course of evolutionary natural selection (Passino and Seeley 2006). Thus, the quorum comes close to an optimization.Footnote 3 All that limits the bees’ choice are the distance and speed with which they can identify new nest sites.
The Homo oeconomicus is the rational decision maker who has constant goals (maximizing money) and has full information and unlimited information processing capacities. In contrast, the bounded rational individual loses interest in alternative courses of action if they do not satisfy or they otherwise seem obviously inferior to other alternatives. Under the influence of their genes, the honeybee’s waggle dance becomes continuously weaker, even if the quality of the site has not changed. This mechanism is essential to allow new alternatives to be sufficiently considered to avoid wrong decisions.
The bee colony’s collective information processing is done in a decentralized manner. This democratic procedure provides more data for consideration than any one individual could possibly gather. By contrast, humans are often organized into hierarchies (such as employer/employee relationships). Their environment also includes decentralized elements (such as democratic elections). Likewise, with rational choice theories, initially no clear demarcation exists between centralized and decentralized decision-making processes (such as Downs’ economic theory of democracy, 1957, or Niskanen’s economic theory of bureaucracy, 1971).
The decision-making behavior of the bee colony, as a superorganism, falls somewhere between the bounded rational human individual and the rational Homo oeconomicus. The honeybee superorganism is, however, more like the Homo oeconomicus. One could perhaps even speak of an Apis oeconomicus.
4 What can we learn from the superorganism of the bees?
In many situations, individuals do not soley play against their nature but make decisions within groups. In human homes, for example, we find situations requiring decisions where the entire family exerts a reciprocal influence on each other. Group decisions also take place when making business or political decisions. The nature of the group decisions determines the need to coordinate the various individuals within a group. Appropriate cooperation mechanisms can help to identify potentially shared individual interests to increase the likelihood of a consensus. This works well in some situations, but less well in others. The divergence between individual and collective rationality is an important reason why Pareto-optimal solutions may not be achieved.
We now ask how and to what extent individuals can learn individually and in groups from how bees decide in their search for a nest site. Comparisons between bees and human collectives are not easy. While bees make binding agreements (in other words: they play a cooperative game), people sometimes end up in a prisoner’s dilemma (Ledyard 1995). Evolutionary processes may have resulted in natural selection and elimination of bees not prone to cooperating with the collective.
With their organizational, communications, and coordination mechanisms, bee colonys provide plenty of inspiration for decision making in human collectives. In particular, they offer significant suggestions for breaking of path dependence, mechanisms for correcting errors, and the need for decentralized/centralized decisions.
4.1 Path dependence: discounting
“What used to be okay can’t be all bad now!” is one sentiment associated with path dependence. People are path dependent if they behave very similar to their past decisions without considering new options. Planck (1948) describes the lengthy process of overcoming long-held beliefs as follows:
A new scientific truth does not triumph by convincing its opponents and making them see the light, but rather because its opponents eventually die, and a new generation grows up that is familiar with it.
The honeybees have developed a clever strategy for remaining open to new ways. Regardless of the quality of an alternative, support for this is discounted over time. In other words: The individual’s approval for a nesting site decreases over time. However, the alternative nest sites are not forgotten (until consensus is made), but will continue to be examined by the other scout bees.
Optimal results in the collective are only possible if people are willing to consider new alternatives in their decision-making process. This means complying with basic rules of communication and ways of thinking and tolerating the results, regardless of who had been advocating for them. However, forgetting the old options at all may also be unfavorable to humans since their environment is changing. If heuristics do not work today, maybe they will work tomorrow under different cricumstances.
4.2 Avoiding and correcting mistakes: discounting and independent examination
Bees do not make rash decisions; they check and evaluate the proposals of other scouts objectively. They demonstrate openness to other views (by allowing room for other opinions) and respect a critical review of information by third parties. This makes it possible to process a variety of information so that the collective is able to make better decisions than an individual can make.
The bees eliminate support for lesser-quality nesting sites by the two mechanisms of discounting and independent evaluation by other bees. If a scout makes a mistake and a nest site is mistakenly found to be optimal, an audit will be carried out by other scouts. If they fail to confirm the suitability of this potential site, it will no longer be lobbied for and interest in it will disappear (Seeley 2014). This audit, conducted by scouts that have so far remained neutral, helps the colony to select an optimal nest site. The independent audit of the quality of the various alternatives and the associated decision for the optimal nest site has successfully secured the continued survival of the species Apis mellifera. The overall system of the bee colony can be considered as relatively stable. The vulnerability of bee colonies to make a mistake and choose the wrong nest site is kept low through these mechanisms of information gathering and processing.
People are often limited, however, in conducting unprejudiced and objective reviews of facts. They join the opinions of other individuals, because holding divergent views can result in emotional stress. Therefore, it makes sense in group decisions either to use anonymity as a mechanism (but not as a consequence of the cooperative honeybee behaviorFootnote 4) and allow voting to occur without this emotional baggage, or to call in mediators to settle disputes (Gerardi and Yariv 2008; Freixas and Zwicker 2009). This knowledge can be valuable for successful negotiations.
4.3 Centralized or decentralized decisions
As old as the discussion about the optimal level of government and market for society is, it remains a modern concern (Smith 1776; Eucken 1952; Eichengreen 1995; Taylor 2009). The bee colony makes decentralized decisions about the nest site. What criteria are used to allow the bees to come to very good to optimal results? With homogeneous goals and clear rules for voting, it is possible to make good decisions and cooperate in a decentralized manner. Distributed information processing in a democracy works because the conditions are clearly defined. What have become rules of the game among human beings may be regulated by genetic mechanisms in bees (uniform goals and tools).
As mentioned earlier, humans pursue, in contrast to honeybees, multiple goals and sometimes end up in a prisoner’s dilemma. Humans and honeybees are formed under different levels of selection (Pavlic and Pratt 2013). Hierarchy may serve as a tool to prevent selfish humans to break their association with the group. Otherwise, individuals may choose options in which they individually receive a higher payoff via defecting, but collectively are worser off than all individuals would cooperate (social dilemma; Dawes 2009). This concern does rather less apply to honeybees that are cooperative and thus relatively anonymous.
Orienting to other social norms can greatly simplify the lives of human beings. However, it is not necessarily the case that group decisions are better than individual decisions for humans. The question of whether group decisions are better than individual decisions gains particular importance given the increased social networking via modern communication media (such as Facebook and Twitter). According to Nietzsche (1886), crowds are definitely not superior to individuals: “Madness is rare in individuals—but in groups, political parties, nations, and ages it is the rule.” Other authors are more optimistic or even identify determinants for group intelligence. Surowiecki (2005), for example, proposes four conditions which characterize wise crowds: diversity of opinion, independence of opinion, decentralization, and an aggregation mechanism that transforms private judgements into collective decisions. These conditions are especially well met by the honeybee Apis mellifera. The bees have a clear division of responsibilities, division of labor, and the skills to deal with difficult tasks. Successful communication is also essential, as are mechanisms for problem solving. If these requirements are not met, the entire process can be disrupted and hierarchies may prove better-suited as decision-making structures.
We also learn from the bees that successful cooperation does not have to be free of confrontation or debate (Livnat and Pippenger 2006). Competition and peaceful conflict can increase the quality of decisions. This aspect is related to Hayek’s notion of “competition as a discovery procedure” (Hayek 1969). Given the rules of a game, competition can help to successfully process information.
5 Concluding remarks
The honeybee Apis mellifera and its decision-making process for finding a new nest site provides an almost perfect model of successful collective coordination of individuals. Based on the limited information available to a single bee, many bees manage to organize themselves into a collective so that at the end (almost) optimal decisions on the selection of a nest site can be made. Although people pursue multiple goals, organizational principles can be identified that may also be useful for humans.
We have demonstrated that it is worthwhile to take a closer look at decision-making processes in other species. However, a stronger interdisciplinary cooperation among researchers from different disciplines (especially biologists, economists, and psychologists) is desirable to come to useful insights collectively.
Our article raises, however, more questions than it can answer. The search for answers should include looking at other superorganisms (such as ants) that achieve good coordination of individuals in the collective. However, negative examples in which cooperation functions inadequately in the animal kingdom should not be neglected. These, too, can provide valuable lessons that can lead to better decisions and coordination mechanisms for humans.
Notes
Unlike many insects that overwinter as individual eggs, larvae, or pupae in a state of hibernation, bees spend their winters awake. In the hive, we find an approximately constant temperature even in winter. The bees come together to create a winter cluster, allowing heat to be exchanged from bee to bee. Thus, temperatures of more than \(10^{\circ }\hbox {C}\) can be reached within the cluster, keeping the bee from torpor, which causes death due to hypothermia (Bichtler 1958; Seeley 2014).
Gigerenzer et al. (1999) define the consequence of the fit between heuristic and environment as ecological rational. The individual adapts to the environmental challenges by using simple and frugal heuristics. For example, the recognition heuristic reads as follows: “If one of two objects is recognized and the other is not, then infer that the recognized object has the higher value.” Imagine, an individual has to bet which of the two towns Dortmund or Munich has the higher population. Our individual may never have heard about Dortmund and correctly infers that Munich has the higher population. This heuristic, which only works in cases of limited knowledge, does not necessarily lead to correct inferences. However, it shows that a lack of recognition may be even informative (Goldstein and Gigerenzer 2002).
Many honeybees at any time are not fulfilling tasks in favor of the colony. Moreover, they make many mistakes as individuals. However, the outcome of the honeybees collective is significantly improved by coordinating them as superorganism.
Anonymity is, however, not universal among more basal eusociality (e.g., paper wasps are not anonymous; Sheehan and Tibbetts 2011).
References
Bichtler, E. (1958). Im Bienenland. Berlin: Deutscher Bauernverlag.
Dawes, R. M. (2009). Social dilemmas. Annual Review of Psychology, 31, 169–193.
Degen, J. K., Kirbach, A., Reiter, L., Lehmann, K., Norton, P., Storms, M., Koblofsky, M., Winter, S., Georgieva, P. B., Nguyen, H., Chamkhi, H., Greggers, U., & Menzel, R. (2015). Exploratory behaviour of honeybees during orientation flights. Animal Behaviour, 102, 45–57.
Downs, A. (1957). An economic theory of democracy. New York: Harper and Row.
Eichengreen, B. (1995). Golden fetters. New York: Oxford University Press.
Eucken, W. (1952, 1990). Grundsätze der Wirtschaftspolitik. Tübungen: Mohr Siebeck.
Freixas, J., & Zwicker, W. S. (2009). Anonymous yes–no voting with abstention and multiple levels of approval. Games and Economic Behavior, 67(2), 428–444.
Gerardi, D., & Yariv, L. (2008). Information acquisition in committees. Games and Economic Behavior, 62(2), 436–459.
Gigerenzer, G., Hertwig, R., & Pachur, T. (2011). Heuristics: The foundations of adaptive behavior. Oxford: Oxford University Press.
Gigerenzer, G., & Selten, R. (2001). Bounded rationality: The adaptive toolbox. Cambridge, MA: MIT Press.
Gigerenzer, G., Todd, P. M., & the ABC Research Group. (1999). Simple heuristics that make us smart. New York: Oxford University Press.
Goldstein, D. G., & Gigerenzer, G. (2002). Models of ecological rationality: The recognition heuristic. Psychological Review, 109(1), 75–90.
Greggers, U., Schöning, C., Degen, J., & Menzel, R. (2013). Scouts behave as streakers in honeybee swarms. The Science of Nature (Naturwissenschaften), 100(8), 805–809.
Hayek, F. A. (1969). Der Wettbewerb als Entdeckungsverfahren. In A. F. Hayek (Ed.), Freiburger Studien (pp. 249–265). Tübungen: Mohr Siebeck.
Hölldobler, B., & Wilson, E. O. (2009). The superorganism: The beauty, elegance, and strangeness of insect societies. New York: Norton & Company.
Hoppe, F. (1930). Erfolg und Mißerfolg. Psychologische Forschung, 14(1/2), 1–62.
Janson, S., Middendorf, M., & Beekman, M. (2005). Honeybee swarms: How do scouts guide a swarm of uninformed bees? Animal Behaviour, 70(2), 349–358.
Ledyard, J. O. (1995). Public goods: A survey of experimental research. In J. H. Kagel & A. E. Roth (Eds.), Handbook of experimental economics (pp. 111–194). Princeton, NJ: Princeton University Press.
Livnat, A., & Pippenger, N. (2006). An optimal brain can be composed of conflicting agents. Proceedings of the National Academy of Sciences, 103(9), 3198–3202.
Nietzsche, F. (1886). Jenseits von Gut und Böse. Leipzig: Naumann.
Niskanen, W. A. (1971). Bureaucracy and representative government. Chicago: Aldine-Atherton.
Oeser, G.-A. (1962). Der Bien und Du. Dresden: Deutscher Landwirtschafts Verlag.
Passino, K. M., & Seeley, T. D. (2006). Modeling and analysis of nest-site selection by honeybee swarms: The speed and accuracy trade-off. Behavioral Ecology and Sociobiology, 59(3), 427–442.
Pavlic, T. P., & Pratt, S. C. (2013). Superorganismic behavior via human computation. In P. Michelucci (Ed.), Handbook of human computation (pp. 911–960). New York: Springer.
Planck, M. (1948). Wissenschaftliche selbstbiographie: Mit einem Bildnis und der von Max von Laune gehaltenen Traueransprache. Leipzig: Johann Ambrosius Barth Verlag.
Rangel, J., & Seeley, T. D. (2012). Colony fissioning in honey bees: Size and significance of the swarm fraction. Insectes Sociaux, 59(4), 453–462.
Sauermann, H., & Selten, R. (1962). Anspruchsanpassungstheorie der Unternehmung. Zeitschrift für die gesamte Staatswissenschaft, 118(4), 577–597.
Schlegel, T., Visscher, P. K., & Seeley, T. D. (2012). Beeping and piping: Characterization of two mechano-acoustic signals used by honey bees in swarming. The Science of Nature (Naturwissenschaften), 99(12), 1067–1071.
Seeley, T. D. (1977). Measurement of nest cavity volume by the honey bee (Apis mellifera). Behavioral Ecology and Sociobiology, 2(2), 201–227.
Seeley, T. D. (1995). The wisdom of the hive: The social physiology of honey bee colonies. Cambridge, MA: Harvard University Press.
Seeley, T. D. (2003). Consensus building during nest-site selection in honey bee swarms: The expiration of dissent. Behavioral Ecology and Sociobiology, 53(6), 417–424.
Seeley, T. D. (2014). Bienendemokratie: Wie Bienen kollektiv entscheiden und was wir davon lernen können. Frankfurt am Main: Fischer.
Seeley, T. D., & Visscher, P. K. (2004). Quorum sensing during nest-site selection by honeybee swarms. Behavioral Ecology and Sociobiology, 56(6), 594–601.
Seeley, T. D., Visscher, P. K., & Passino, K. M. (2006). Group decision making in honey bee swarms. American Scientist, 94(3), 220–229.
Seeley, T. D., Visscher, P. K., Schlegel, T., Hogan, P. M., Franks, N. R., & Marshall, J. A. R. (2012). Stop signals provide cross inhibition in collective decision-making by honeybee swarms. Science, 335(6064), 108–111.
Sheehan, M. J., & Tibbetts, E. A. (2011). Specialized face learning is associated with individual recognition in paper wasps. Science, 334(6060), 1272–1275.
Simon, H. A. (1957). Models of man, social and rational: Mathematical essays on rational human behavior in a social setting. New York: Wiley.
Simon, H. A. (1983). Reason in human affairs. Stanford, CA: Stanford University Press.
Smith, A. (1776). An inquiry into the nature and causes of the wealth of nations. London: W. Strahan.
Srinivasan, M. V. (2011). Honeybees as a model for the study of visually guided flight, navigation, and biologically inspired robotics. Physiological Reviews, 91(2), 413–460.
Surowiecki, J. (2005). The wisdom of crowds. New York: Random House.
Tautz, J., & Heilmann, H. R. (2007). Phänomen Honigbiene. München: Spektrum Akademischer Verlag.
Tautz, J., Rohrseitz, K., & Sandeman, D. C. (1996). One-strided waggle dance in bees. Nature, 382(32), 32.
Taylor, J. B. (2009). Getting off track: How government actions and interventions caused, prolonged, and worsened the financial crisis. Stanford, CA: Hoover Institution Press.
von Frisch, K. (1965). Tanzsprache und Orientierung der Bienen. Berlin: Springer.
Wainselboim, A. J., & Farina, W. M. (2000). Trophallaxis in the honeybee Apis mellifera (L.): The interaction between flow of solution and sucrose concentration of the exploited food sources. Animal Behaviour, 59(6), 1177–1185.
Weibull, J. W. (1995). Evolutionary game theory. Cambridge, MA: MIT Press.
Zeiler, C. (1989). 300 Ratschläge für den Freizeitimker. Leipzig: Neumann Verlag.
Acknowledgments
We would like to thank the editors of the special issue Ulrich Witt and Jennifer Fewell as well as the anonymous reviewers for their comments, ideas, and criticism. We would also like to express our sincere thanks to the beekeeper Werner Kaatz for all our fruitful discussions, the good moments we had together and sharing his knowledge with us. In addition, we are very grateful for the financial support provided by ScienceCampus Halle (WCH) and the German Research Foundation (DFG).
Author information
Authors and Affiliations
Corresponding author
Additional information
The title of our paper was inspired by the famous quotation of Muhammad Ali “Float like a butterfly, sting like a bee”.
Rights and permissions
About this article

Cite this article
Grüner, S., Fietz, A. & Jantsch, A. Float like a butterfly, \({\varvec{decide}}\) like a bee. J Bioecon 17, 243–254 (2015). https://doi.org/10.1007/s10818-015-9204-5
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10818-015-9204-5
Keywords
- Honeybee Apis mellifera
- Bounded rationality
- Collective intelligence/group intelligence
- Superorganism
- Apis oeconomicus