
Abstract
Chronic inflammation is a common underlying factor in many major diseases, including heart disease, diabetes, cancer, and autoimmune disorders, and is responsible for up to 60% of all deaths worldwide. Metformin, statins, and corticosteroids, and NSAIDs (non-steroidal anti-inflammatory drugs) are often given as anti-inflammatory pharmaceuticals, however, often have even more debilitating side effects than the illness itself. The natural product-based therapy of inflammation-related diseases has no adverse effects and good beneficial results compared to substitute conventional anti-inflammatory medications. In this review article, we provide a concise overview of present pharmacological treatments, the pathophysiology of inflammation, and the signaling pathways that underlie it. In addition, we focus on the most promising natural products identified as potential anti-inflammatory therapeutic agents. Moreover, preclinical studies and clinical trials evaluating the efficacy of natural products as anti-inflammatory therapeutic agents and their pragmatic applications with promising outcomes are reviewed. In addition, the safety, side effects and technical barriers of natural products are discussed. Furthermore, we also summarized the latest technological advances in the discovery and scientific development of natural products-based medicine.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Tissue dysfunction or homeostatic imbalance triggers the immune system’s adaptive response, known as inflammation. Inflammatory responses are caused as part of the host’s initial line of defense. However, chronic inflammation can result from the recurrent stimulation by dangerous substances or inadequate management of acute inflammation (Ma et al. 2021). Diseases such as cardiovascular disease (CVDs), neurodegeneration, and inflammatory bowel disease (IBD) are linked to chronic inflammation (Hussain et al. 2020). Chronic inflammation and the diseases that arise from it are the major risk to public health. Diseases that promote chronic inflammation, such as diabetes, CVDs, cancer, stroke, and obesity, kill three out of every five people worldwide. Lifelong disability, expensive treatment, and wage loss all contribute to a substantial economic burden (Bleich et al. 2015; Nguyen et al. 2018).
Metformin, statins, and corticosteroids, and NSAIDs (non-steroidal anti-inflammatory drugs) are often given as anti-inflammatory pharmaceuticals that reduce inflammation through various mechanisms. Existing treatments for most chronic diseases, however, often have even more debilitating side effects than the illness itself. The natural product-based therapy of inflammation-related diseases has no hazardous side effects and great curative results to substitute conventional anti-inflammatory medications. Natural products, which are substances obtained from natural resources such as plants, animals, and micro-organisms, had been utilized to cure a wide range of human problems for millennia (Bernardini et al. 2018; Dias et al. 2012; El-Hadary and Ramadan 2019). Recent investigations have observed that natural products possessing anti-inflammatory action can act as possible sources for the advancement of natural anti-inflammatory drugs due to their high pharmacological activity and low toxicity (Wu et al. 2021; Alharbi et al. 2022). Natural products are more successful at alleviating subtle pain in the early phase of many diseases, but they are not always effective in treating the more severe symptoms of chronic disorders. The understanding of chronic inflammation and the use of natural products for its treatment have advanced significantly in recent years. Scientists aim to develop natural products with potent anti-inflammatory activity and no negative effects to replace traditional anti-inflammatory drugs. This review aims to provide insight into the significance of natural products in treating inflammation-related disorders and their mechanisms of action. It discusses various commonly used natural products, their effects on different inflammatory cellular signaling pathways, and summarizes the positive results from clinical and preclinical studies. The safety and potential side effects of these natural products are also addressed, as well as the latest advances in their discovery and development.
Chronic inflammation-related diseases
Both acute and chronic inflammations are protective responses to harmful pathogens including viruses, bacteria, and fungi. In most cases, inflammation is beneficial to the body (Deng et al. 2022). Pain, redness, bloating, heat, and failure to function are the hallmarks of inflammation (Scott et al. 2004). This physiological process, which tries to eliminate the stimulus and return the body to a state of balance, includes vasodilation as well as the recruitment of leukocytes and granulocytes into inflamed tissues. If the acute inflammatory reaction is not successful in halting the stimulus, it will transform into a chronic inflammatory cascade. Disorders such as obesity (Villarroya et al. 2018), atherosclerosis (Kasikara et al. 2018), rheumatoid arthritis (RA) (Chimenti et al. 2015), gout (So and Martinon 2017), Alzheimer’s disease (AD) (Heneka et al. 2015), depression (Koopman and Aidy 2017), Parkinson’s disease (PD) (Joshi and Singh 2018), and cancer (Coussens and Werb 2002) are all linked to chronic inflammation and its physiological and pathological processes (Fig. 1).

Chronic inflammation contributes to several diseases
RA is a systemic autoimmune syndrome that disturbs the joints and can cause distress, swelling, and rigidity. Synovial lining hyperplasia and pannus formation are caused by the permeation of inflammatory cells, pro-inflammatory cytokines, and the activated fibroblast-like synoviocytes (FLS). These factors ultimately lead to bone and cartilage damage through the stimulation of osteoclasts (Croia et al. 2019).
Atherosclerosis is a disease that occurs within the artery wall and is described as a persistent, low-grade inflammatory condition that is driven by lipids. It is the cause of several CVDs, involving peripheral artery disease, myocardial infarction (MI), coronary artery disease, and stroke. Endothelial dysfunction is responsible for the deposition of low-density lipoprotein (LDL) and the recruitment of monocytes. These monocytes eventually transform into macrophages, after which they take up oxidized LDL (oxLDL) and undergo differentiation into foam cells. A lipid core that may contain foam cells and necrotic debris is covered by smooth muscle cells that multiply and drift from the media to the intima. This process is one of the factors that leads to an atherosclerotic plaque. In conclusion, thrombosis can be caused by the rupture of an unstable plaque (Nguyen et al. 2019).
Asthma is characterized by bronchial hyper-reactivity, airway inflammation, and structural changes in the airways. Dendritic cells migrate and mature when the airway epithelium produces inflammatory mediators in response to environmental stimuli. Dendritic cells that have been stimulated can cause naive T cells to specialize into subsets. Overproduction of mucus and changes to the structure of the airways result from subsequent inflammatory processes such as cytokine release and recruitment of inflammatory cells (Mims 2015).
Numerous studies have linked neuroinflammation to the etiology of AD, finding that microglia (brain-resident macrophages) clusters all around Aβ plaques and the related inflammatory cytokines are elevated in AD patient. Once engaged, microglia move to the Aβ deposits, where they remove the Aβ through phagocytosis and exhibit neuroprotective action. However, microglia are harmful when activated for too long. Their ability to clear Aβ decreases, and they start releasing inflammatory intermediaries such as cytokines, chemokines, nitric oxide (NO), reactive oxygen species (ROS), and proteolytic enzymes. Increased synthesis and deposition of Aβ, hyper-phosphorylation of tau, migration of peripheral monocytes and lymphocytes, and neuronal injury all result from these mediators, which engage in many roles in the pathogenesis of AD. In addition, Aβ can trigger astrocytes to secrete neurotoxic substances, worsening neuroinflammation and neurodegeneration (Baik et al. 2016; Bolmont et al. 2008; Wang et al. 2015; Lee et al. 2010).
Through its effects on the tumor microenvironment, inflammation has been also related to numerous stages in carcinogenesis, involving tumor development, progression, and metastasis (Fig. 2) (Greten and Grivennikov 2019). Inflammation may begin as a result of exposure to a pathogen, an immune-mediated disease (such as IBD), subclinical inflammation (such as that caused by obesity), exposure to environmental carcinogens (such as tobacco smoke and pollutants), or the activation of a proto-oncogene (Mantovani et al. 2008). DNA damage and genomic instability induced by ROS, NOS, and other mediators promote neoplastic transformation of the inflamed tissues (Colotta et al. 2009). The crosstalk among the immune cells, including tumor-coupled immune cells; macrophages, neutrophils, dendritic cells, mast cells, and T cells results in the formation of an inflammatory microenvironment influences the tumor growth (Molinaro et al. 2018; Diakos et al. 2014).

Chronic inflammation accelerates the growth of tumors
Cellular signaling mediates inflammatory processes
The immune response of inflammation triggers many signaling pathways, and the transcription of mRNAs encoding pro-inflammatory cytokines and chemokines such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), and IL-6 is up-regulated (Tanaka et al. 2014). During inflammation, numerous mechanisms, including the nuclear factor kappa-B (NF-κB) pathway influence the transcription level of pro-inflammatory cytokines (Wajant et al. 2003). Furthermore, upon activation of toll-like receptor-4 (TLR4), it subsequently stimulates mitogen-activated protein kinase (MAPK)pathways through a substantial activation of the stress-related Jun n-terminal kinase (JNK) group, a modest response from the p38-MAPK, and a limited initiation of the established extracellular signal-regulated kinases (ERKs) that participate in activator protein-1 (AP-1) regulation (Kant et al. 2011). The expression of inflammatory cytokines and mediators, cell proliferation and survival are all regulated by additional inflammatory pathways, such as Janus kinase/signal transducers and activators of transcription (JAK-STAT). These pathways help to orchestrate the inflammatory responses (Fig. 3) (Hu et al. 2021). Cytokines and chemokines attract extra immune cells to the location of infection, such as circulating neutrophils, which promote microbial death by producing proteases (Netea et al. 2017), interferon (IFN)-γ, as well as ROS. Cytokines likewise stimulate the synthesis of cyclooxygenase-2 (COX-2) which is responsible for the production of prostaglandins (PG) (Gomez et al. 2015; Gandhi et al. 2017). Furthermore, the dendritic cells enhance the immune reaction via naïve T- and B-cell stimulation. The removal of the extraneous or endogenous substance and the reprogramming of effector cells to efficiently stop the manufacture of inflammatory mediators result in the determination of inflammation and the restoration of homeostasis. Failure to do so, on the other hand, resulted in sustained periods of unresolved inflammation, which contributes to practically all chronic or degenerative diseases. In order to make therapeutic advances against most of these degenerative diseases for which there is now no treatment, it is necessary to produce medications that target underlying inflammation (Netea et al. 2017; Ma et al. 2018; Afonina et al. 2017).

The cellular signaling involves in the body’s inflammatory response. MyD88-dependent pathways, when TLRs are activated, they initiate a sequence of intracellular signaling events that ultimately result in the movement of AP-1 and NF-κB to the nucleus, thereby controlling the pro-inflammatory cytokine production and inflammatory cell recruitment, which eventually contribute to the inflammatory response. (Toll-like receptors (TLRs), myeloid differentiation factor-88 (MyD88), the mitogen-activated protein kinase (MAPK), nuclear factor kappa-B (NF-κB), and Janus kinase (JAK)-signal transducer and activator of transcription (STAT), activator protein-1 (AP-1)
Current treatments of inflammation and their side effects
The currently available treatments alleviate the signs and symptoms of inflammation, such as heat, redness, swelling, and pain via suppressing the pro-inflammatory chemical mediators, but they can also lead to severe immune suppression, which can increase the risk of opportunistic infections. It is known that the temporal events of self-limited acute inflammatory responses resolve at the histologic level with the removal of inflammatory cells from the tissue and the restoration of function (Serhan 2017; Dyall et al. 2022). However, if the inflammatory response is not resolved, it can become chronic inflammation, which can lead to serious health problems. Thus, it is important to identify and address the underlying cause, reduce exposure to toxins, and adopt a healthy lifestyle including regular exercise, a balanced diet, stress reduction, and smoking cessation can also help reduce inflammation.
Steroidal and NSAIDs are the most popular agents to treat inflammation. Long-term use of these treatments, however, can have negative and occasionally life-threatening side effects, such as gastrointestinal damage and failure of liver and kidney. They are rarely curative and have low success rates; therefore, patients need safer, less toxic, and cheaper treatments (Li et al. 2017). The use of more selective drugs and innovative treatments affecting certain cells such as T-cell targeted medicines (tacrolimus, laquinimod, and edratide), cytokine inhibitors (adalimumab and infliximab) (Ginwala et al. 2019), and anti-B-cell agents (BLyS and rituximab) has been implemented in inflammation treatment with variable degrees of success. Nevertheless, research into more benign and economically viable therapy options remains urgently needed (Li et al. 2017; Rengasamy et al. 2019). Steroids, NSAIDs, COX inhibitors, and anti-TNF therapies can cause side effects such as stomach ulcers, bleeding, high blood pressure, diabetes, liver damage, increased risk of infections, allergic reactions, increased risk of cancer and suppression of the immune system if used for an extended period. As a result, new tactics are required to reduce these side effects. In this context, Natural products are good candidates to treat inflammation because they often contain compounds that have anti-inflammatory properties and can target multiple pathways involved in the inflammatory process. They are often less toxic, have fewer side effects, can have synergistic effects, and may have other health benefits. Natural products-based medication may help give fresh ideas and a renewed sense of optimism for treating inflammation (Dinarello 2010; Roy et al. 2022).
Anti-inflammatory promise of curative herbal products and their active constituents
Polyphenols
Curcumin
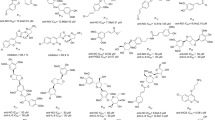
Fruits, vegetables, and other foods contain polyphenols, a group of natural compounds. Polyphenol curcumin (Fig. 4) is derived from Curcuma longa (turmeric). It has antioxidant, anti-inflammatory, anticancer, and antiviral activities across a spectrum of disease types, thanks to its pharmacological versatility and broad molecular target specificity (Wang et al. 2021a; Edwards et al. 2017; Qadir et al. 2016). At the molecular level, it has been demonstrated that this multitargeted drug can act through a variety of different cell signaling pathways: the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR), the glycogen synthase kinase-3 beta (GSK-3β), the Ras/Raf/MEK/ERK, the p53, and through Nrf2/ARE, Akt, and NF-κB pathways (Kunnumakkara et al. 2017; Paulraj et al. 2019). Recent research has shown that curcumin can raise the levels of nuclear translocation and stimulate the biological outcomes of Nrf2. Therefore, it can be a potential therapeutic candidate for treating numerous disorders that are produced by oxidative stress (Rahban et al. 2020). In addition, curcumin can inhibit the activities of xanthine oxygenase, lipo-oxygenase (LOX), COX, NO synthesis, ROS formation, and the production of pro-inflammatory mediators including NF-κB (Mathy-Hartert et al. 2009; Bezáková et al. 2014; Goel et al. 2001; Chan et al. 2005; Sivani et al. 2022; Buhrmann et al. 2011). Curcumin blocked the joint histopathological alterations in collagen-induced arthritis (CIA) rats, involving edema, cartilage/bone damage, synovial hyperplasia, coupled with pannus development, by diminishing the release of pro-inflammatory cytokines (Wang et al. 2019a). A meta-analysis found that treatment with standardized turmeric extracts for 8–12 weeks (typically consisting of one thousand milligrams of curcumin per day) can reduce arthritis symptoms (primarily those related to pain and inflammation) and leads to progresses in relieving symptoms that are comparable to those produced by ibuprofen and diclofenac sodium (Yuan et al. 2019). Consequently, curcumin and turmeric extracts was prescribed for the treatment of arthritis. Additional evidence comes from a study performed by Delecroix et al. It was demonstrated that professional rugby players who took supplements containing 2 g of curcumin and 20 g of piperine experienced less muscular soreness 24 h following their workout (Delecroix et al. 2017).

Chemical structures of curcumin, EGCG, resveratrol, and lentinan
Curcumin was noticed to have an atheroprotective effect by modifying macrophage polarization from the pro-inflammatory M1 to the anti-inflammatory M2 phenotype via the TLR4/MAPK/NF-κB pathways (Zhou et al. 2015). Recently, curcumin was found to alleviate airway irritation in ovalbumin (OVA)-stimulated asthma model, causing reduction of cytokine quantities and inflammatory cell number via its activity on MAPK/NF-κB pathways (Zhu et al. 2019). It correspondingly abridged the infiltration of inflammatory cell as well as IL-6 and transforming growth factor beta (TGF-β) levels owing to its repressing properties on NF-κB and COX-2 release, which led to attenuation of airway remolding that occurred in a model of chronic obstructive pulmonary disease (COPD) that was persuaded by lipopolysaccharide (LPS) and cigarette burn (Yuan et al. 2018). Curcumin hindered inflammation by targeting NF-κB transcription factors, TNF-α, ILs, enzymes of COX-2, iNOS, and 5-LOX, and malignancies proteins (Giordano and Tommonaro 2019). The chemical inhibitory action of curcumin is one of the various methods that can be used to limit the proliferation of tumors. Because of this, using curcumin topically significantly reduces inflammation brought on by tetradecanoylphorbol-13-acetate 12-O (Rakariyatham et al. 2019), hyperplasia, ornithine decarboxylase activity, formation of active oxygen species, DNA oxidative damage and papillomavirus formation (Hour et al. 2002; Zoi et al. 2021; Shishodia et al. 2007). Curcumin’s several interactions with the cells of the human gastrointestinal tract have been shown to have the following effects: inhibition of lipid peroxidation, suppression of COX-2 expression and PGE2 generation, and elevation of glutathione-s-transferase enzyme levels (Pulido-Moran et al. 2016; Mbese et al. 2019).
Epigallocatechin-3-gallate (EGCG)
One of the utmost prevalent catechins in green tea (Camellia sinensis) is called epigallocatechin-3-gallate (EGCG, Fig. 4). Reduced ILs production, COX-2, and matrix metalloproteinases-2 (MMP-2) expression in human being rheumatoid arthritis synovial fibroblasts (RASFs) was attributed to EGCG’s potential interference with the IL-1 signaling cascade via reversible binding and inhibition of an upriver kinase TGF-activated kinase 1 (TAK1) (Fechtner et al. 2017). ROS production as well as TNF-α, IL-5, Th2 cytokine, and MMP-9 expression was reduced by EGCG, in an asthma model that was induced by toluene diisocyanate. This, in turn, prevents the inflammatory cells from recruitment and extravasation to the airway lumen, which in turn reduces airway hyperresponsiveness (Kim et al. 2006). EGCG inhibits the expansion of psoriasis-like skin inflammation caused by imiquimod by lowering the quantity of T cells that infiltrate the skin and lowering the quantities of cytokines associated with Th17 (Zhang et al. 2016). In addition, there is evidence that EGCG may have a neuroprotective effect via hindering NF-κB and MAPK activation, which in turn prevented IL-1β + Aβ-induced the enzyme of COX-2, IL-6, and -8 in human astrocytoma cells. This allowed it to exert anti-neuroinflammatory effects in AD (Rubio-Perez and Morillas-Ruiz 2012). In LPS-induced PD rats, EGCG not only reduces the levels of TNF-α and NO, but it also enhances the quantity of dopaminergic neurons in the midbrain (Al-Amri et al. 2013). In addition, EGCG protects prostate cancer cells from inflammation by preventing the gene expression of inflammatory chemokines and cytokines, the activities of MMP-9 and -2, and cell migration by disrupting NF-κB (Sen et al. 2009). This is achieved by EGCG’s interference with these proteins (Wang et al. 2021a).
Since EGCG is rapidly destroyed in both acidic and neutral conditions, it is unsteady in the GI tract and is not instantly absorbed. Consequently, chitosan encapsulated EGCG nanoparticles (NPs), considerably slowed its breakdown in digestive juices. Furthermore. EGCG targeted therapy is required to protect healthy cells while targeting cancerous ones (Safer et al. 2019). When EGCG was encapsulated in polylactic acid-polyethylene glycol NPs (nano-EGCG), its medicinal efficiency was increased by a factor of more than 10 in terms of employing proapoptotic and angiogenic repressing properties; these are the most important determining factors of EGCG’s chemopreventive properties in both in vitro and in vivo schemes (Siddiqui et al. 2009). There is evidence that EGCG can help fight obesity. Treatment of diabetic mice with black tea extract for an extended period of time decreased their body weight and prevented a rise in blood sugar after eating (Grove and Lambert 2010). Six overweight males were given EGCG 300 mg twice daily for 2 weeks, and their resting energy expenditure did not change significantly compared to the people given placebo. Interestingly, EGCG administration resulted in considerably reduced respiratory quotient values compared to the placebo during the first postprandial monitoring phase (Boschmann and Thielecke 2007). These ground-breaking results point to the anti-obesity effects of EGCG on its own through increasing fat oxidation.
Resveratrol
Grapes, peanuts, blueberries, and mulberries are just a few examples of the many plant species that contain the polyphenolic stilbene resveratrol (trans-3,5,4ʹ-trihydroxystilbene, Fig. 4). Both its ability to scavenge ROS and its ability to increase cellular antioxidant defense have been credited with resveratrol’s antioxidant potential (Lastra and Villegas 2007; Pervaiz and Holme 2009). Through the lowering of COX-2, PGE2, ROS, cytokines, and Th17 cell polarization, it has been demonstrated that resveratrol is a promise in the treatment of RA. Resveratrol may reduce atherosclerosis by decreasing the TLR4-mediated inflammatory response, restricting monocyte adherence by downregulating intercellular adhesion molecule-1 (ICAM-1) expression, and preventing the development of monocytes into macrophages (Martins et al. 2014; Chen et al. 2018; Malaguarnera 2019). Resveratrol reduced the inflammation and remodeling of airways in asthma-induced animal models, and the underlying machinery is owing to its ability to control the mobility group box 1/TLR4/NF-κB pathway and spleen associated tyrosine kinase (Syk) protein production (Jiang et al. 2019). It can serve as a stimulator of sirtuin 1 (SIRT1), lowered the lung inflammatory response and oxidative stress in COPD mice, and inhibited the release of airway smooth muscle cytokines and chemokines (Wang et al. 2017a). It was discovered that resveratrol was helpful in improving inflammatory IBD, and its therapeutic efficacy on ulcerative colitis (UC) and Crohn’s disease (CD) is, respectively, related to the reduction of pro-inflammatory cytokines and the rebalancing of Treg/Th17 cells (Shi et al. 2017; Samsami-Kor et al. 2015; Zhu et al. 2011). Both in vitro and in vivo testing on animal models of neurodegenerative disorders revealed that resveratrol possessed neuroprotective properties. For example, treatment of resveratrol reduced Aβ-caused inflammation in astrocytes and cultured microglia and decreased microglial activation in an animal model of brain amyloid sedimentation. This was accomplished by suppressing NF-κB signaling (Zhao et al. 2018a). In addition, resveratrol protected SNpc dopaminergic neurons in an animal model of PD by controlling inflammatory responses via activation of suppressor of cytokine signal 1 (SOCS1) (Lofrumento et al. 2014).
The anti-inflammatory properties of resveratrol have been demonstrated to have a chemopreventive effect on liver cancer (Zhang et al. 2018a). The anticancer effects of resveratrol-loaded nanoparticles have also been shown in cancer cell lines. Blocking IL-18 release and IL-18-induced vascular cell adhesion molecule-1 (VCAM-1) expression inhibited hepatic melanoma spread and microvascular adhesion. While it blocked COX-2 and NF-κB signaling, it reduced inflammation and liver carcinogenesis (Shen et al. 2017). Resveratrol inhibits iNOS production in colon cancer cells, blocks the Insulin-like growth factor 1 receptor (IGF-1R)/Akt/Wnt pathways, and triggers p53, all of which work together to slow the growth of cancer cells and tumors (Honari et al. 2019; Sá et al. 2018; Clark et al. 2017).
Polysaccharides (PSA)
The polysaccharide-based dietary fibers have collected a lot of interest in the latest years. The health benefits of dietary fiber are numerous and include the reduction of total and LDL cholesterol, the maintenance of a healthy blood pressure, the prevention of constipation, and the promotion of healthy gut microflora (Dhingra et al. 2012). The human gut microbiome includes the anaerobe Bacteroides fragilis (B. fragilis). Capsular PSA is a natural substance that helps keep the peripheral nervous system and/or central nervous system free of inflammation (Mazmanian et al. 2008; Ramakrishna et al. 2019). PSA is a zwitterionic polysaccharide that is also a component of the immunogenic capsule produced by the non-toxin-producing B. fragilis (NTBF) strain NCTC9343; an exceptional T-cell-dependent antigen which is responsible for mediating the rise of CD4+ T cells and inducing the release of the immunosuppressive cytokine IL-10 in regulatory T cells (Johnson et al. 2015). A recent study on biodiversity revealed that there is a direct connection between NTBF and the immune system. Because of this connection, NTBF may have potential application as a probiotic (Sun et al. 2019). By suppressing TLR4-intermediated NF-κB transcription and modulating drug-absorbing enzyme and transporter expression, PSA derived from the probiotic B. fragilis improves aberrant voriconazole metabolism. As a result, PSA has the potential to act as a therapeutic adjuvant in clinical medication (Wang et al. 2021b). Most fungal PSA are derived from the Basidiomycetes family (and some of which are derived from the Ascomycetes). They have become a component of traditional food and medicine that is well known and utilized by many people. They display a broad variety of pharmacological actions such as antibacterial, anticancer, immunoregulatory, and antidiabetic (Giavasis 2014).
Lentinan
Lentinan (Fig. 4), which is often referred to as Lentinus edodes, is a medicinal fungal polysaccharide that is extensively used and has been the subject of massive research. It is a recently commercialized and therapeutic medicine in the treatment of diabetes, and it possesses potent anticancer activity. Lentinan has a structure that is characterized by a 1,3-β-glucopyranosidic backbone with two 1,6-β-d-glucopyranosidic branches interspersed each five glucose units along the main chain (Jiao et al. 2011). The highly ordered structure of lentinan is what is believed to be responsible for its potent antitumor activity. This structure can stimulate IL-12 which is responsible for stimulating the production of natural killer cells and T lymphocytes (Wiederschain 2007). It was also shown to be an efficient adjuvant for T cells and to boost the growth of blood mononuclear cells such as lymphocytes, monocytes, and macrophages, which increases the body’s resistance to the transition of cancerous cells into other types of cancer (Lemieszek and Rzeski 2012).
Schizophyllan
Schizophyllan (SPG, Fig. 5), which is a form of fungal polysaccharide, was also one of the earliest natural products separated from the fungus S. commune. SPG possessed significant biological properties and was subsequently used as an immunomodulator, an anticancer medication, and a component in bioactive cosmetics in various pharmaceuticals and functional foods. These anticancer PSA share a structural property that is analogous to that of 1,3-β-glucan with 1,6-β-glucose branches attached to the skeleton. The existence of a broad range of anticancer PSA in various species of fungi is explained by the fact that biologically dynamic compounds could be acquired by the enzymatic degradation of the hemicellulose skeleton in the fungal mycelial cell wall (Zhang et al. 2013). Because of its impressive clinical efficacy in treating lung, cervical, and stomach malignancies, it was approved for use in Japan (Lemieszek and Rzeski 2012). The large molecular weight and viscosity of SPG have, however, limited its use in the pharmaceutical and cosmetics industries. Three ultrasonic-treated SGP (USPG) sections with low viscosity and small molecular weight were developed in a recent study that used ultrasonic technology to handle SPG. USPG sections increased NO generation in macrophages RAW264.7, lymphocyte growth, spleen lymphocyte IL-2 and TNF-α, and T-47D cell inhibition compared to SPG (Zhong et al. 2015a). Consistent with previous reports, these findings showed that ultrasonic therapy improved the immunological and anticancer activity of native SPG. The ultrasonic treatment technique for SPG production was found as an effective method to provide PSA with higher biological activity, and USPG60 may be a promising operational element for immunoregulatory and cancer therapy applications (Zhong et al. 2015b).

Chemical structures of schizophyllan, apigenin, and luteolin
Sulfated PSA
Most research into bioactive PSA has focused on sulfated PSA. Abundant sulfated PSA derived from algae showed immuno-inflammatory, antioxidant, antibacterial, antiproliferative, and antilipidemic properties in addition to their virus-fighting powers (Jiao et al. 2011; Wijesinghe and Jeon 2012). Research has shown that a sulfated polysaccharide isolated from the marine red alga Schizymenia pacifica can inhibit HIV replication. This sulfated polysaccharide preferentially inhibits the activity of HIV reverse transcriptase (RT) and HIV replication in vitro (Witvrouw and Clercq 1997). Sulfated fucoidan, isolated from commercially grown Cladosiphon okamuranus TOKIDA, has been indicated to reduce the proliferation of human leukemia U937 cells through the induction of apoptosis linked with the degradation of poly [ADP-ribose] polymerase (PARP), caspase-3 and -7 activation. C. okamuranus TOKIDA, the sulfated fucoidan producer, is now a commercially grown organism. In Japan, a fucoidan extracted from C. okamuranus TOKIDA is included in a diversity of healthful foods, beverages, and personal care goods (Teruya et al. 2007).
Flavonoids
Flavonoids, also known as phytonutrients, are a set of compounds that serve multiple purposes and can be found in many different types of food and drink. They provide an essential library of compounds that can be utilized as medicinal entities due to their broad array of biological action involving antioxidant, anti-mutagenic, anti-inflammatory, and antiviral activities in addition to their appealing features (Tungmunnithum et al. 2018; Panche et al. 2016; Patel et al. 2018).
Apigenin and luteolin
It has been established that chamomile products taken orally are effective in reducing inflammation caused by gastrointestinal illnesses. The positive impacts of chamomile formulations are linked to the existence of a wide variety of plant metabolites. These metabolites include bioflavonoids, coumarins, proazulenes, their breakdown product chamazulene, and the essential oil. Apigenin and luteolin (Fig. 5) end products, the utmost abundant flavonoids, have strong in vivo and in vitro anti-inflammatory capacities via regulation of numerous related pathways (Flemming et al. 2015; Gerritsen et al. 1995; Alghamdi et al. 2022).
Treatment with luteolin improves colonic damage and inflammation in rats with UC by reducing TNF-α-induced inflammatory injury and altering the gut flora’s composition and structure. Inhibiting the IL-6/STAT3 signaling pathway, as luteolin does, slows the progression of colon cancer (Li et al. 2021).
By employing valuable participants of the flavonoid family, such as apigenin and lutein, the PI3K/Akt pathway can present a promising medicinal option for conquering the clinical problems of lung cancer malignant tumor heterogeneity and developed resistance (Zhou et al. 2017). Lung cancer cells exposed to lutein showed proliferation suppression and activated apoptosis by blocking the PI3K/Akt signal (Zhang et al. 2018b). Apigenin is a recent AKT suppressor in lung cancer, and it was shown to limit the phosphorylation of Akt. In addition, it was shown to inhibit HEF1 (human enhancer of filamentation 1), MMP-9, and GSK-3 genes expression (Zhou et al. 2017; Tong and Pelling 2013).
Hispidulin and diosmetin
As a traditional medicine, the plant Clerodendrum inerme (L.) Gaertn., which is a member of the Verbenaceae family. Through the inhibition of a variety of inflammatory indicators in LPS-stimulated macrophages, the ethyl acetate portion of the extract has the most effective anti-inflammatory activity compared to other portions. In addition, the inhibition of inflammatory markers led to the isolation of three well-known flavones: acacetin, diosmetin, and hispidulin. Hispidulin (Fig. 6) is the most effective anti-inflammatory medication out of the three flavones because it inhibits the DNA-binding action of NF-κB/JNK signaling pathway. This, in turn, leads to a decline in the levels of critical inflammatory targets such as iNOS and COX-2 (Srisook et al. 2015). Hispidulin reduces the synthesis of NO along with the release of inflammatory cytokines in Raw264.7 macrophage cells that have been treated with LPS (Park et al. 2017).

Chemical structures of hispidulin, diosmetin, and quercetin
Diosmetin (Fig. 6) promotes apoptosis and suppresses the spread of cancer cells while possessing no effect on healthful cells (Androutsopoulos et al. 2009). Several signal transduction pathways of interest in cancer have been linked to diosmetin’s growth-inhibiting properties (Liu et al. 2016; Barrajón-Catalán et al. 2015; Ciolino et al. 2002). Diosmetin induces apoptosis in cancer cells, which may have anti-carcinogenic effects. Diosmetin restrains the expansion of human prostatic malignant cells through triggering apoptosis and cell cycle arrest (Oak et al. 2018). The combination therapy of diosmetin and paclitaxel synergistically decelerated the progress of lung cancer cells by causing an accumulation of ROS triggered by the disturbance of the PI3K/Akt/GSK-3β/Nrf2 pathway (Chen et al. 2019).
Quercetin
Quercetin (Fig. 6) is a polyphenolic bioflavonoid found in a broad range of fruits and vegetables. In CIA rats, quercetin was found to have anti-arthritic properties via altering the equilibrium between Th17 and regulatory T cells and triggering the heme oxygenase 1 (HO-1)-arbitrated anti-inflammatory reaction (Yang et al. 2018). Through the attenuation of the TLR/NF-κB pathway, quercetin could inhibit the oxLDL-stimulated atherosclerotic inflammatory process. This included the adherence of endothelial leukocytes as well as the production of inflammatory mediators such as TNF-α, IL-6, COX, 5-LOX, and NOS. Quercetin was also able to reduce the amount of cerebral edema in the brain tissues and protect against the damage that high altitude can cause to the brain (Mehany et al. 2022). Quercetin decreased allergic airway inflammation and airway hyperresponsiveness in mice with OVA-induced asthma (Park et al. 2009). Furthermore, it reduced COPD-related lung inflammation and emphysema in mouse models. A 7-day quercetin administration reduced the post-exercise glucose-caused insulin response, improved total antioxidant capacity (TAC) and superoxide dismutase (SOD) levels and decreased malondialdehyde (MDA) levels in the recovery period, according to results from a pilot study. However, it was observed that no significant differences existed between quercetin and placebo trials with regards to glucose, respiratory exchange rate, TNF-α, myoglobin, and high sensitivity C-reactive protein (Tsao et al. 2022). Applying LPS-stimulated rat intestinal microvascular endothelial cells, the anti-inflammatory effect of quercetin on the intestine was examined. Quercetin suppressed the levels of the adhesion molecules in cellular model of IBD via inhibiting TLR4/NF-κB and JNK/STAT signaling pathways (Bian et al. 2018). In addition, quercetin effectively suppressed inflammatory mediators’ levels in an imiquimod-stimulated animal model of psoriasis via inhibiting NF-κB signaling (Chen et al. 2017). The anti-inflammatory effects of quercetin can have a part in preventing against cancer. As a suppressor of the IL-6-stimulated STAT3 signaling pathway, it counteracted the glioblastoma cells’ ability to proliferate and migrate by lowering the production of downriver genes such Cyclin D1 and MMP-2 (Michaud-Levesque et al. 2012). In addition, Quercetin causes apoptosis in tongue squamous cell carcinoma cells through the JNK activation regulated ERK/GSK-3/mediated mitochondria-dependent apoptotic signaling pathway (Huang et al. 2022). It also showed promise against colon cancer by diminishing TNF-α, COX-2, and IL-6 levels and by inhibiting the TLR4/NF-κB pathway, which controls the production of migration and invasion proteins such as E-cadherin and MMPs (Li et al. 2016).
Alkaloids
Berberine
Many medicinal plants contain berberine (Fig. 7), which is an isoquinoline alkaloid. It has anti-arthritic properties by controlling the equilibrium among Th17 and Treg cells via lowering IL-6 and -17 levels but simultaneously raising IL-10 and TGF-β levels (Wang et al. 2017b). In addition, recent meta-analyses demonstrated that berberine was effective in reducing total cholesterol (TC) as well as LDL cholesterol. The combination of berberine and silymarin was found to considerably reduce cholesterol levels, according to the findings of a meta-analysis that comprised 19 separate controlled and cross-sectional assessments (Bertuccioli et al. 2020). Berberine was able to treat asthma (Li et al. 2016) and COPD by inhibiting the NF-κB pathway, which may result in decreased thymic stromal lymphopoietin (TSLP) and pro-inflammatory cytokines levels. This is a suggested molecular way by which berberine works in the treatment of these conditions (Wang et al. 2019b; Moon et al. 2011). The therapeutic potential of berberine for IBD was further supported by the fact that it inhibited the differentiation of Th1 and Th17 cells and their correlated cytokines and modified the population of M1 macrophages (Cui et al. 2009; Li et al. 2015). Berberine has the potential to defend microglia cells towards the Aβ-induced inflammation that is associated with AD by downregulating inflammatory mediators through NF-κB and MAPK signal pathway (Jia et al. 2012). Diabetes-related nephropathy, renal ischemia, renal fibrosis, renal damage, and renal stones are only few of the renal diseases that have been shown by several researchers to be susceptible to berberine’s renoprotective properties. In addition, it protects the kidneys against the toxicity that many drugs and substances, such as chemotherapy, heavy metals, aminoglycosides, and NSAIDs, might cause (Hassanein et al. 2022).

Chemical structures of berberine, piperine, and sinomenine
Piperine
Piperine (Fig. 7) is the primary active chemical found in both black pepper (Piper nigrum) and long pepper (Piper longum). In IL-1-induced FLS from RA subjects, piperine suppressed IL-6, COX-2, and MMPs levels. In addition, it reduced the affected region in the ankle joints of animal arthritis models (Bang et al. 2009). Piperine helps to reduce inflammation triggered by gouty arthritis by inhibiting the activity of the NLRP3 inflammasome. A recent pharmacodynamic paradigm of piperine as a competitive suppressor of JNK-1 (MAPK family) and IKK-1β (NF-κB activator) was demonstrated by a molecular docking study. This model examined the starring role that piperine plays in the immunosuppression of gout inflammation by focusing on how it regulates the NLRP3 inflammasome. These findings pointed to the viability of piperine both as a preventative measure and as a potential treatment for gouty arthritis (Jati et al. 2022). In animal models of UC, piperine served as therapeutic candidate for the treatment of IBD because it reduced oxidative stress, TNF-α, IL-1β, IL-6, NF-κB and IFN-γ levels, inflammatory mediators (iNOS and COX-2), while increasing anti-inflammatory cytokine (IL-10) (Guo et al. 2020; Gupta et al. 2015). When it was given orally, it decreased parasitaemia in Plasmodium berghei ANKA-infected mice (Khairani et al. 2022). Moreover, reduced Rac1 gene and protein expression is a mechanism by which piperine was noticed to diminish the proliferation and migration of human breast cancer cells (Buranrat and Junking 2022). Indeed, piperine’s neuroprotective effect in PD was not limited to its antioxidant and anti-apoptotic properties; the compound as well demonstrated anti-inflammatory properties through reducing levels of cytokines and chemokines in the striatum, which in turn reduced microglia triggering (Bang et al. 2009; Guo et al. 2020; Gupta et al. 2015).
Sinomenine
Sinomenine (Fig. 7) is an alkaloid that has been taken from the root of Sinomenium acutum for ages to treat RA. It stops RA from getting worse because it has anti-inflammatory effects, such as dropping the release of VCAM-1 and inflammatory cytokines and chemokines by blocking NF-κB, variable monocyte/macrophage populations, adjusting the number of Treg and Th17 cells in gut-allied lymphoid tissues, lowering PGE2 by blocking the expression of mPGES-1, and preventing FLS propagation through modulation of α7 nicotinic acetylcholine receptor (α7nAChR) (Liu et al. 2018; Zhi et al. 2022; Yi et al. 2018). Although sinomenine has been analyzed primarily for its anti-arthritic properties, recent investigations have shown that it may also have therapeutic value for treating other inflammatory illnesses. By inhibiting TLR/NF-κB signaling, it reduced pro-inflammatory cytokine release, reduced mucosal inflammation, and reduced clinical indications in investigational colitis, suggesting that it may be a good nominee for treating IBD (Xiong et al. 2017). In addition, sinomenine therapy has been established to boost dopamine receptor D2 (DRD2)-mediated nuclear expression of B-crystallin (CRYAB) in astrocytes, by this means inhibiting STAT3 and reducing neuroinflammation (Qiu et al. 2016). In addition, it has a role in alleviating morphine dependence and sleep disturbance disorders since it inhibits NMDA receptor-mediated Ca2+ influx and to promote GABAA receptor-mediated Cl− influx (Hong et al. 2022). Sinomenine was also able to restrain Aβ-induced microglial stimulation in AD by decreasing the release of ROS, NOS, inflammatory cytokines, and chemokines, and it also protected neurons from indirect toxicity (Shukla and Sharma 2011). It also has a neuroprotective impact on PD dopaminergic neurotoxicity by lowering the production of TNF-α, PGE2 and ROS by triggered microglia. This neuro-preservation was linked to the fact that it inhibited microglial nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (Bao et al. 2022; Qian et al. 2007). Recently, sinomenine was found to possess antiarrhythmic, antioxidant, and anti-inflammatory properties in myocardial ischemia–reperfusion injury (MIRI) mouse model since it was able to diminish MIRI-caused cardiac damage in vivo by inhibiting oxidative stress, inflammation, and apoptosis (Xia et al. 2022).
Terpenoids
Boswellic acids
The main bioactive components of B. serrata extracts are known as boswellic acids, which belong to the class of compounds known as triterpenes. More than a dozen distinct types of boswellic acids were recognized inside the resin. Their anti-inflammatory effects are produced by a variety of distinct modes of action. They involve the suppression of leukotriene and, to a lesser level, PG production, the decreased production of pro-inflammatory cytokines, and the inhibition of ROS and NOS formation (Ammon 2016). One of the most significant pharmaceutically active boswellic acids is acetyl-11-keto-β-boswellic acid (AKBA, Fig. 8). AKBA decreases the atherosclerotic lesion dimension and inflammation in atherogenesis by suppressing NF-κB signaling and plasma concentrations of NF-κB-dependent genes in LPS-treated inflammation (Shang et al. 2016; Taherzadeh et al. 2022; Takada et al. 1950). It inhibited the inflammatory reactions and mucosal damage by reducing leukocyte-endothelial cell adhesive interactions in a rat model of experimental IBD (Krieglstein et al. 2001). AKBA was able to alleviate the signs of psoriasis in an animal model by modulating a pathway that is related to TLR7/8. This was accomplished by inhibiting dendritic cells activation and T-cell differentiation. In addition, AKBA was able to downregulate the release of IL-12/-23 in dendritic cells (Wang et al. 2018a). AKBA relieved neuroinflammation in subjects with standard physical characteristics of AD by lowering the amounts of inflammatory cytokines (IL-1β, IL-6 and TNF-α) in cortex and hippocampus, most likely through the NF-κB pathway. This probably influenced in part to its neuroprotective activities (Gunasekaran et al. 2021; Wei et al. 2020). By downregulating NF-κB and its target genes, which are implicated in inflammation, AKBA was able to inhibit the development of human colorectal cancer in a xenograft animal model, which led to a reduction in tumor size (Yadav et al. 2012). Curcumin and boswellic acids were recently ranked among the most successful herbs for pain killing in osteoarthritis in the short duration of action by a new meta-analysis that evaluated a significant number of dietary supplements (Bannuru et al. 2018; Haroyan et al. 2018). The synergistic effects of curcumin and boswellic acid suggest that this combination may be more effective than either component used alone in treating osteoarthritis; nonetheless, it is not yet routinely utilized in practice.

Chemical structures of AKBA, celastrol, and triptolide
Celastrol
Celastrol (Fig. 8) is a pentacyclic triterpenoid obtained from Tripterygium wilfordii. Celastrol inhibited the invasion and migration of FLS from RA subjects through decreasing TLR4/NF-B-mediated MMP-9 expression and altering the synthesis of chemokines and chemokine receptors (Fang et al. 2017). Celastrol enhanced the neurological performance and decreased the cortical cell mortality and cerebral infarct volume in transient middle cerebral artery occlusion mice. It also prevented I/R-induced increases in lactate dehydrogenase A (LDHA), glucose transporter1 (GLUT1), and hexokinase2 (HK2) and lactate generation and decreases in ATP and glucose levels. It also suppressed the I/R-induced hypoxia-inducible factor-1 (HIF-1α) and pyruvate dehydrogenasekinase1 (PDK1) overexpression (Chen et al. 2022a, b; Zhang et al. 2020). In a mouse model of asthma, oral administration of celastrol reduced airway inflammation and tissue remodeling by correcting the inequity between MMP-2/-9 and tissue suppressors of metalloproteinases (TIMP-1/-2) caused by pro-inflammatory cytokines through MAPKs and NF-κB (Kim et al. 2009). In COPD animals produced by cigarette smoke, celastrol decreased cytokines and oxidative stress indicators through endothelin receptor type B (EDNRB)/kininogen 1 (KNG1) signaling (Shi et al. 2018). Inhibiting NLRP3 inflammasome stimulation, decreasing rise of the IL-23/IL-17 pathway, and reestablishing anti-inflammatory response in a mouse model of dextran sulfate sodium (DSS)-induced colitis suggests celastrol as a feasible nominee for UC treatment (Shaker et al. 2014). Reduced oxidative stress and inflammation in the kidney and adipose tissue, as well as improved insulin sensitivity in T2D mice, were all effects of celastrol’s role as an NF-κB inhibitor (Kim et al. 2013). Albumin nanoparticles loaded with the biodegradable steroid celastrol reduce fat accumulation, boost insulin sensitivity, and lessen inflammation in mice with a diet-induced obesity. Enhanced bioavailability and in vivo effectiveness of celastrol-BSA-NPs for the management of diet-induced obesity were observed when compared with free celastrol (Fan et al. 2022). In addition, cancer treatment can benefit from its ability to reduce inflammation. In gastric cancer, celastrol blocked the action of biglycan (BGN), which resulted in the stimulation of necroptosis and the suppression of pro-inflammatory cytokine release (Guo et al. 2019). In addition, it was found to decrease inflammatory responses in UC-associated colorectal cancer by switching the overrun of cytokines production, upregulation of COX-2 and iNOS, and stimulation of NF-κB (Lin et al. 2015). Celastrol was found to be a histone deacetylase inhibitor reducing the growth lung and colorectal cancer cells via shifting macrophage polarization from M2 to M1 and influencing the microenvironment of the colorectal tumor (Chen et al. 2022b; Wang et al. 2022).
Triptolide
T. wilfordii is the source of the diterpenoid epoxide known as triptolide (Fig. 8). In CIA rats, triptolide was found to modify the JAK2/STAT3 signal pathway, which followed by a decline in the release of pro-inflammatory cytokines. Consequently, the severity of arthritis was reduced (Fan et al. 2016). Triptolide could also slow the expansion of atherosclerosis by blocking the production of inflammatory cytokines in macrophages (Luo and Yang 2016).
By way of IL-6/STAT3 signaling, triptolide was able to downregulate Th1 cytokines which resulted in an improvement in the experimental colitis (Li et al. 2010). In keratinocytes, triptolide caused interference with IFN-γ signaling, which led to a reduction in IFN-γ-caused ICAM-1 expression and, as a result, an inhibition of IFN-γ-induced skin inflammation, which is implicated in psoriasis (Hongqin et al. 2011). Using an Aβ-activated cultured microglia and an animal model of AD, researchers found that triptolide suppressed TNF-α and IL-1β production, providing evidence for its neuroprotective effect in AD (Jiao et al. 2008). In addition, it showed neuroprotective effects on PD by defending dopaminergic neurons from inflammation-mediated injure caused by microglial activation, cytokine overproduction, COX-2 overexpression, and PGE2 release (Huang et al. 2018). In addition, because it inhibited the development of family receptors for chemokines, IL, TNF-α and TGF-β, triptolide was thought to be an active agent against the spread of colon cancer (Johnson et al. 2011). Although triptolide has been shown to have therapeutic potential for a range of disorders through in vitro and in vivo research, there are still a lot of obstacles to go over before it can be used in clinical practice. Poor water solubility, a small therapeutic window, multiple organ toxicity, and substantial side effects after prolonged use are the four most significant problems (Tong et al. 2021; Hou et al. 2019). Intravenous administration of nanoparticle-encapsulated triptolide (Nf-trip) has revealed an improved aggregation of the drug in the cancer tissues. The Nf-trip reaction to the acidic tumor microenvironment is permitted by the pH-vulnerable peptide. This, in turn, boosts the site-exact triptolide release and cell-specific drug absorption, which, in the end, increases the efficiency while simultaneously decreasing the toxicity (Ling et al. 2014). Furthermore, Wang et al. used the solid dispersion approach to produce a triptolide AS-PPT that was loaded with a HOOC-PEG-PDLLA aptamer (AS1411) and evaluated in chemo-resistant pancreatic cancer (CPC) cell lines. The in vitro data revealed that anticancer activity of AS-PPT was greater than that of the parent triptolide. In addition, AS-PPT had a similar certain binding ability with gemcitabine-resistant human pancreatic cancer cells (MIA PaCa-2). Applying biophotonic imaging, AS-PPT selectively targeted the tumor tissues (Wang et al. 2016). Another water-soluble derivative of triptolide F6008 called PG490-88 has also shown promising outcomes in preclinical mouse xenograft models of lung and colon cancer. PG490-88 was also found as a preventive drug against I/R-induced lung injury (Pao et al. 2019). Another triptolide derivative known as MRx102 exhibited a cytotoxic activity in human leukemia cells when tested in vitro (Reno et al. 2016). These results show that triptolide has a therapeutic potential against a wide range of disorders, and it could do so by transitioning from the laboratory to clinical settings.
Betulin and betulinic acid
Betulin and betulinic acid (Fig. 9) are pentacyclic triterpene secondary metabolites found in over 200 plant species. They display many pharmacological effects, involving anti-inflammatory, d anti-HIV, anticancer, and anti-parasitic actions (Jonnalagadda et al. 2017; Oliveira-Costa et al. 2022; Cichewicz and Kouzi 2004; Zhao et al. 2018b; Meira et al. 2016). Betulin considerably attenuated mucin glycoprotein (MUC5AC) synthesis in NCI-H292 cells and down-regulated MUC5AC mRNA expression in response to phorbol 12-myristate 13-acetate (PMA). It also blocked PMA-induced NF-κB stimulation. Owing to the betulin’s inhibitory effect of kappa-B kinase (IKK), NF-κB p65 could be translocated into the nucleus and degrade inhibitory kappa-B alpha (IκBα). Consequently, this resulted in a reduction in MUC5AC glycoprotein synthesis in NCI-H292 cells. Based on these findings, it seemed that betulin suppresses mucin gene expression in human airway epithelial cells by modifying the NF-κB signaling pathway (Hossain et al. 2022). Using an OVA-induced asthma model, betulin was found to reduce the allergic airway inflammation through regulating inflammatory cytokines, transglutaminase, and antioxidants (Kamaraj et al. 2021). By way of the TLR4/NF-κB and SIRT1/AMPK intracellular signaling pathways, betulin prevented renal injury and ethanol-induced liver impairment (Bai et al. 2016; Zhao et al. 2016). Using a rat model of lung inflammation produced by LPS, researchers found that a betulin derivative suppressed neutrophil recruitment and inflammatory mediator expression (Nader and Baraka 2012). Betulin has been also shown to reduce lung damage in sepsis and mammary gland inflammation injury (Zhao et al. 2016; Guo et al. 2015), and decrease the release of inflammatory cytokines in human cardiac cells (Zhang et al. 2015). Betulin was also reported to be candidate of palliative therapy for COPD by targeting the P2X7 receptor in human bronchial epithelial cells (Jiao et al. 2022). Bevirimat, a well-known analog of betulinic acid, was discovered to inhibit HIV through virus maturation inhibition (Martin et al. 2008). Attempts to use this compound to treat HIV have progressed to the Phase II clinical trial stage (Smith et al. 2007). As a result of Bevirimat’s success, several studies were conducted to determine the usefulness of betulin analogs as various classes of therapeutic drugs, and several lead targets with multiple mechanisms of action have been discovered for further development. Extensive anticancer research has shown that betulinic acid induces apoptosis by direct control of mitochondrial pathways and elevated synthesis of caspase-3 (Kim et al. 2021). In addition to obstructing the cell cycle in the G2/M phase (Zhan et al. 2018), betulinic acid has been shown to influence immunoregulation in vivo (Wang et al. 2012). It has also been observed that betulinic acid protects thymocytes from the apoptosis caused by the steroid dexamethasone in an in vivo model (Yi et al. 2016). Through the AKT/Nrf2/HO-1/NF-κB signal axis, betulin also attenuated IL-1β-induced OA (Ren et al. 2021).

Chemical structures of betulin, betulinic acid, bilobalide, and picrotoxinin
Bilobalide
Bilobalide (Fig. 9), a sesquiterpenoid lactone derived from Ginkgo biloba, is an antagonist of GABAA and GABAC receptors. It shares structural similarities with the GABA antagonist picrotoxinin (Fig. 9), involving a lipophilic side chain and a hydrophilic cage. Bilobalide was found to give its action via interacting within the chloride channels of ionotropic GABA receptors. As a result, it is categorized as a negative allosteric modulator. However, there is a significant variation in the in vivo actions of bilobalide and picrotoxinin. In contrast to picrotoxinin, bilobalide is an anticonvulsant. The absence of convulsant activity in a drug that inhibits GABA function may be crucial for cognitive improvement. Bilobalide’s lack of convulsant activity may be due to suppression of glutamate release, which reduces synaptic excitement and outweighs reductions in synaptic inhibition (Johnston et al. 2009). There is evidence that bilobalide can reduce inflammation, boost mitochondrial function, and protect against cerebral ischemia-induced damage (Jiang et al. 2014). Recent research shows that bilobalide has anti-inflammatory impacts on chondrocytes via reducing IL-17-induced inflammatory damage to ATDC5 cells by down-regulation of micro-RNA-125a through the JNK and NF-κB signaling pathways. Chondrocyte inflammation was found to be reduced by bilobalide via the AMPK/SIRT1/mTOR pathway (Mao et al. 2019).
Clinical trials and studies
Between 1981 and 2023, over 50% of U.S. Food and Drug Administration (FDA)-approved pharmaceuticals were natural derivatives of herbal products, involving anticancer treatments and immunosuppressants. As evidenced by their significance in the in vivo and in vitro treatment of multiple diseases, numerous clinical trials are carried out to assess the safety and efficiency of natural derivatives for inflammation-related diseases (Table 1).
In preclinical research, curcumin was found to possess anticancer properties. Curcumin’s anticancer properties include suppressing carcinogenesis, angiogenesis, and tumor growth, enhancing the efficacy of chemotherapy, and protecting healthy cells from radiation therapy damage. Clinical trials evaluating the effectiveness of curcumin in the prevention of breast cancer (NCT01740323, phase II), prostate cancer (NCT01917890 and NCT03211104), pancreatic cancer (NCT00094445, phase II), and colorectal cancer (NCT00027495, phase I) have already been achieved (Table 2). All concluded that curcumin is safe, and no specific adverse effects have been reported. Curcumin has completed phase I clinical trials for the therapy of breast cancer metastasis in combination with docetaxel. In addition, curcumin and paclitaxel have showed safety and efficacy in phase II clinical trials including subjects with advanced metastatic breast cancer. This phenolic compound is now being assessed in phase III clinical studies for its potential to inhibit the progression of cancer in individuals with low-risk prostate cancer who are undergoing active monitoring. Curcumin has not been authorized by the FDA as a therapy for cancer or any other medical condition. Curcumin’s anti-arthritic qualities are another therapeutic possibility; 29 randomized controlled trials with 2,396 applicants and 5 forms of arthritis were incorporated. Curcumin was provided at dosages extending from 120 to 1500 mg for 4–36 weeks. In all investigations, curcumin was found to be safe and relieved inflammation and pain in subjects with arthritis. Because of the minimal quality and small number of randomized controlled trials, the decisions must be cautiously evaluated. Piperine, in conjunction with curcumin, is now being evaluated in a phase I clinical trial for the management of bladder cancer (Chinta et al. 2015), early-stage prostate cancer or patients with monoclonal gammopathy of unknown significance (MGUS), smoldering multiple myeloma (NCT04731844).
The traditional Chinese drug berberine altered the cell cycle, apoptosis, autophagy, and microenvironment of tumors. Moreover, berberine was demonstrated to possess anti-inflammatory and antioxidant properties. In latest years, immune-suppressants as PD-1/PD-L1 suppressants have become apparent one next to the other in the field of cancer immunotherapy. Due to their high cost, however, it is difficult for them to be widely used in clinical settings. Being as an excellent immunomodulator and a type of inexpensive Chinese traditional medicine, berberine is anticipated to be commonly employed in clinical routine as an appropriate immunotherapy treatment. In a phase II/III clinical trial (NCT02226185), berberine prevented the recurrence of colorectal adenoma in a safe and effective manner. The water-soluble prodrug of triptolide known as minnelide has an anticancer efficacy in a variety of preclinical cancer models (Chugh et al. 2012). This prodrug is now being tested in phase II clinical studies (NCT04896073) for the treatment of advanced refractory adenosquamous carcinoma of the pancreas (ASCP) (Skorupan et al. 2022) and in phase I for advanced non-small cell lung cancer with mutated EGFR (NCT05166616). Dose-escalation, safety, pharmacokinetic, and pharmacodynamic of minnelide™ capsules given alone or in order with protein-bound paclitaxel in patients with advanced solid tumors is currently undergoing in phase I clinical trial (NCT03129139).
An ongoing clinical trial involving 37 people at stage III of lung cancer has been conducted. In this investigation, EGCG at a concentration of 440 mol/L was administered both during and 2 weeks after radiotherapy. The radiation therapy oncology group (RTOG) score and the weekly pain score were both found to be significantly lower than pre-treatment levels after treatment (NCT02577393) (Zhu et al. 2021; Niu et al. 2020). Recently, NCT02891538 a randomized, ongoing pilot trial studied the chemopreventive effects of EGCG in patients with histological documentation of primary colon or rectal adenocarcinoma, who have not received any treatments for cancer. Meanwhile, NCT00609310 had proved the hypothesis that nutritional supplement with flavonoids mixture of 20 mg apigenin and 20 mg EGCG will diminish the recurrence rate of colonic neoplasia, compared to placebo. Several clinical investigations have analyzed the acute effects of EGCG on cognitive function (such as attention) or mood status. The acquired data demonstrated that tea intake has considerable acute effects on mood status, work performing, and creativity. In second clinical research, EGCG could regulate cerebral blood flow parameters devoid of compromising cognitive function or mood in 27 healthy people. EGCG was delivered orally in a single dose of 135 mg. Similarly, Scholey et al. demonstrated that 300 mg of EGCG was linked with decreased stress, greater tranquility, and increased electroencephalographic activity in the midline frontal and central brain areas (Scholey et al. 2012).
Notably, the clinical indications of AD do not manifest instantly. Because of this, the acute effects of EGCG or other natural substances on neurocognitive skills were not used to predict usefulness in neurodegenerative disorders such as AD. Currently, one clinical investigation is researching the benefits of EGEG in early-stage AD subjects taking acetylcholine esterase inhibitors (NCT00951834). However, clinical research investigating the long-term outcomes of EGCG on cognitive functioning and large-scale epidemiological surveys examining the relationship between EGCG use and the advancement of AD are required.
Recently, 119 subjects with mild-to-moderate AD were enrolled in a phase II randomized, double-blind, placebo-controlled trial of resveratrol due to the proven safety and promising preclinical data of resveratrol. Furthermore, resveratrol can improve cognitive function in AD through a managed immune response that leads to neuronal survival (with dose up to 2 g, synthetic resveratrol by mouth daily, for 12 months). Resveratrol (NCT01504854) significantly decreased MMP-9 levels in cerebrospinal fluid (CSF) of AD patients to preserve BBB integrity and minimize leukocyte infiltration. A prior report indicate that resveratrol modifies Wnt signaling, a signaling pathway which is stimulated in over 85% of colon cancers. A clinical study (NCT00256334) was presented to describe the actions of resveratrol on the Wnt signaling pathway in which subjects with colon cancer obtained treatment with resveratrol, and its impacts directly on colon cancer and normal colonic mucosa was examined. The safety, pharmacokinetics, and pharmacodynamics of resveratrol in patients with colorectal cancer and hepatic metastases (NCT00920803) had been studied in phase II clinical trial.
Resveratrol has been demonstrated to trigger a protein called Notch-1. Signaling of Notch-1 is indicated to inhibit tumor cell growth. The effects of resveratrol on Notch-1 had been examined in neuroendocrine tumor patients in completed pilot trial (NCT01476592).
Currently, a phase I trial (NCT01912820) is evaluating the effect of quercetin on the absorption of green tea polyphenols in the prostate tissue of prostate cancer patients. In addition, quercetin is currently being assessed in a phase II clinical research (NCT03476330) for its chemopreventive potential against the growth of squamous cell carcinoma in subjects with Fanconi anemia. In a mouse model of COPD, oral intake of a low dose of quercetin reduces inflammation, oxidative stress, and MMP production. A recent phase I clinical research was done to establish the safety of quercetin supplementation in COPD patients in order to further investigate its therapeutic potential (NCT01708278). The purpose of another short trial with 8 participants receiving quercetin 2000 mg/day and 4 participants receiving placebo was to determine whether quercetin supplementation can lower inflammation and oxidative stress indicators in COPD patients (NCT03989271).
Conclusion and future perspectives
Prior to the dawn of combinatorial chemistry and advanced techniques such as genomics and proteomics, pharmaceutical companies relied heavily on natural compounds to develop synthetic drugs, which were originally produced in the formula of crude compilations and, more lately, consist of purified compounds due to the dawn of combinatorial chemistry and such techniques. In addition, synthetic derivatives of natural substances with increased properties can be produced.
Now, the dietary supplement market has expanded dramatically in the 25 years, dietary supplements market is expected to present the potential growth opportunities in coming years with the revenue of $220.8 billion by 2027. Although customers frequently mistake “natural” for “safe,” scientists understand the constituents in these natural products can cause toxicity. In addition, when they are ingested alongside pharmaceuticals, the precipitant natural products can change how drugs are delivered and disposed of, enhancing, or decreasing the therapeutic effect of the target drug(s) (Gaston et al. 2020). One of the major concerns regarding consuming herbal supplies is safety. Several defenders of herbal supplies argued that goods with a sustained record of widespread consumption are typically safe when applied accurately at conventional therapeutic dosages. A critical question is the degree to which the lack of proof of toxicity could be testimony of safety of herbal plants or the non-existence of toxicity. Acute symptoms and short-term toxic effects are likely to be identified and linked with medicinal plants. Consequently, the deficiency of evidence of these side effects in the context of conventional usage of medicinal plants is not evidence of the absence of the possibility to produce them. Additional existing difficulties with herbal supplies that need to be resolved include the difficulty of cultivating specific species, unintentional overuse of medicinal and aromatic plants, purity of herbal drugs, extinction of endangered plant species, deforestation and urbanization, quality, safety, effectiveness, dosage, and standardization.
Combinatorial chemistry has led to the relative comfort with which synthetic records of small molecule medications can be created, reducing the effectiveness of natural product-based drug discovery. Nonetheless, significant discoveries such as that of the first organically derived medicine, morphine, as well as those of penicillin and streptomycin and the newer anti-parasitic drugs avermectins and artemisinin demonstrate that natural products are unquestionably the best supplier of drugs. With modern methodologies comes a greater understanding of the structures of many natural chemicals, which in turn permits the comprehension of their specific mechanism of action in health and disease. The resurgence of interest in plant-derived drugs coincides with significant advances in science and technology in other important research areas, including a better understanding of diseases and their fundamental mechanisms, improvements in analytical tools and screening techniques, an increase in the number of testable targets, and improved opportunities for natural leads to be optimized using synthetic modification techniques.
Natural products’ poor stability, aqueous solubility, and bioavailability have frequently impeded the therapeutic translation of these compounds despite the encouraging preclinical findings. There have been attempts to get around these restrictions, especially using nano-based drug delivery systems(s) due to their favorable properties such as being biodegradable, biocompatible, available, renewable, and exhibiting low toxicity (Balaji et al. 2018; Patra et al. 2018). The commonly used nanocarriers which are formulated for natural product-based drug delivery are crystal nanoparticles, liposomes, and polymeric nanoparticles. For instance, Gupta et al. designed chitosan-based nanoparticles loaded with Taxus brevifolia-derived Paclitaxel (Taxol) which demonstrated superior activity with sustained release, high cell uptake, and lower toxicity compared to the pure paclitaxel (Gupta et al. 2017). Another example is novel nanoparticles, developed by Chang et al., shelled with heparin for berberine delivery to treat Helicobacter pylori (Chang et al. 2011). In a study by Testa et al., loading quercetin into nanoparticles improves its efficacy in preventing neuroinflammation caused by oxysterols (Testa et al. 2014). In addition, when polymeric micelles were used to deliver quercetin, it showed continuous in vitro release and boosted accessibility of the drug under in vivo conditions (Dian et al. 2014). Takahashi et al. developed liposome-encapsulated curcumin (LEC) nanoparticles possessing rat plasma levels of 4.96 higher than pure curcumin (Takahashi et al. 2009).
Various methods such as particle-size reduction, salt formation, solubilization, and complexation with β cyclodextrins are used to improve the solubility of drugs with low aqueous solubility. Solid dispersion can improve the solubility and bioavailability of natural products such as plant extracts and essential oils by dispersing them in a hydrophilic carrier at the molecular level, resulting in an amorphous state that increases surface area and dissolution rate. The process needs to be carefully optimized to ensure that the natural product is not degraded or modified, and additional safety evaluations may be required (Bhalani et al. 2022). As mentioned earlier, Wang and colleagues utilized the solid dispersion method to create a triptolide AS-PPT, which was loaded with an aptamer called HOOC-PEG-PDLLA (AS1411) and tested its efficacy in chemo-resistant pancreatic cancer (CPC) cell lines. Another technique used to improve molecular weight, viscosity, and filtration of SPG, as cited former, is ultrasonic treatment technique which was successfully used to reduce the molecular weight of SPG that subsequently provide PSA with higher biological activity.
The use of genomic, proteomic, transcriptomic, and metabolomic technologies has significantly impacted the development of natural product-based therapies and unlock the potential of natural products for drug discovery. These technologies have enabled the identification of biosynthetic gene clusters, specific proteins that interact with natural products, and the metabolic pathways involved in drug delivery (Thomford et al. 2018; Tocmo et al. 2021). Transcriptomic analysis provides insight into cellular processes involved in drug delivery, leading to the identification of potential drug targets, while metabolomic analysis identifies biomarkers that indicate disease progression and response to treatment. Moreover, genetic mutations that influence drug response can be identified using genomic analysis, leading to the development of personalized therapies (Shi et al. 2022). Proteomic analysis helps optimize drug delivery by identifying proteins involved in uptake, transport, and metabolism. Natural product-based therapies that target cancer cells, inflammation, and metabolic pathways have been developed, and potential drug candidates have been identified (Krushkal et al. 2021; Liu et al. 2021). The study of natural products, particularly in terms of their metabolism, toxicity, and activity, may be significantly impacted by LC–MS-based metabolomics. For instance, a recent study by Wu et al. related to the serum metabolomics analysis of the anti-Inflammatory effects of gallic acid on rats elucidated the protective effect of gallic acid against acute inflammation (Wu et al. 2022). Regarding bioinformatics, BATMAN-TCM, a bioinformatics tool of network pharmacology, has also been made available online for forecasting the targets and pharmacologic pathways of natural products (Wang et al. 2018b). Genomic, proteomic, transcriptomic, and metabolomic technologies have advanced and developed tremendously, which could remove many obstacles and considerably may enhance our knowledge of the pharmacology of natural products for the treatment of inflammation.
To sum up, natural products still offer a lot of potential for discovering a range of scaffolds with diverse structures and bioactivities that can be developed directly or optimized into new drugs. As illustrated in Fig. 10, the distinct molecular pathways modulated by each of the previously mentioned natural products in the context of inflammation were summarized.

The distinct molecular pathways modulated by each of the previously mentioned natural products in the context of inflammation. (Toll-like receptors (TLRs), myeloid differentiation factor-88 (MyD88), the mitogen-activated protein kinase (MAPK), nuclear factor kappa-B (NF-κB), and Janus kinase (JAK)-signal transducer and activator of transcription (STAT), Phosphoinositide 3-kinase (PI3K), Protein kinase B (Akt), activator protein-1 (AP-1). Blunt arrows (┴) indicate inhibition, while sharp arrows (→) indicate stimulation
Although drug development faces high failure rates, natural products face additional obstacles such as limited accessibility, sustainable supply, and intellectual property constraints. Despite these challenges, the advancements in science and technology provide a solid foundation for natural products-based drug discovery to make significant contributions to improving human health and extending lifespans.
Data availability
Enquiries about data availability should be directed to the authors.
References
Afonina IS, Zhong Z, Karin M, Beyaert R (2017) Limiting inflammation-the negative regulation of NF-κB and the NLRP3 inflammasome. Nat Immunol 18(8):861–869
Al-Amri JS, Hagras MM, Mohamed IM (2013) Effect of epigallocatechin-3-gallate on inflammatory mediators release in LPS-induced Parkinson’s disease in rats. Indian J Exp Biol 51(5):357–362
Alghamdi A, Almuqbil M, Alrofaidi MA, Burzangi AS, Alshamrani AA, Alzahrani AR et al (2022) Potential antioxidant activity of apigenin in the obviating stress-mediated depressive symptoms of experimental mice. Molecules (basel, Switzerland) 27(24):9055
Alharbi KS, Afzal O, Altamimi ASA, Almalki WH, Kazmi I, Al-Abbasi FA et al (2022) Potential role of nutraceuticals via targeting a Wnt/β-catenin and NF-κB pathway in treatment of osteoarthritis. J Food Biochem 46(12):e14427
Ammon HP (2016) Boswellic acids and their role in chronic inflammatory diseases. Adv Exp Med Biol 928:291–327
Androutsopoulos VP, Mahale S, Arroo RR, Potter G (2009) Anticancer effects of the flavonoid diosmetin on cell cycle progression and proliferation of MDA-MB 468 breast cancer cells due to CYP1 activation. Oncol Rep 21(6):1525–1528
Bai T, Yang Y, Yao YL, Sun P, Lian LH, Wu YL et al (2016) Betulin alleviated ethanol-induced alcoholic liver injury via SIRT1/AMPK signaling pathway. Pharmacol Res 105:1–12
Baik SH, Kang S, Son SM, Mook-Jung I (2016) Microglia contributes to plaque growth by cell death due to uptake of amyloid β in the brain of Alzheimer’s disease mouse model. Glia 64(12):2274–2290
Balaji AB, Pakalapati H, Khalid M, Walvekar R, Siddiqui H (2018) Natural and synthetic biocompatible and biodegradable polymers. Biodegrad Biocompat Polymer Compos 286:3–32
Bang JS, Oh DH, Choi HM, Sur BJ, Lim SJ, Kim JY et al (2009) Anti-inflammatory and antiarthritic effects of piperine in human interleukin 1beta-stimulated fibroblast-like synoviocytes and in rat arthritis models. Arthritis Res Ther 11(2):R49
Bannuru RR, Osani MC, Al-Eid F, Wang C (2018) Efficacy of curcumin and Boswellia for knee osteoarthritis: systematic review and meta-analysis. Semin Arthritis Rheum 48(3):416–429
Bao X, He Y, Huang L, Li H, Li Q, Huang Y (2022) Sinomenine exerts a neuroprotective effect on PD mouse model through inhibiting PI3K/AKT/mTOR pathway to enhance autophagy. Int J Neurosci. https://doi.org/10.1080/00207454.2022.2100780
Barrajón-Catalán E, Taamalli A, Quirantes-Piné R, Roldan-Segura C, Arráez-Román D, Segura-Carretero A et al (2015) Differential metabolomic analysis of the potential antiproliferative mechanism of olive leaf extract on the JIMT-1 breast cancer cell line. J Pharm Biomed Anal 105:156–162
Bernardini S, Tiezzi A, Laghezza Masci V, Ovidi E (2018) Natural products for human health: an historical overview of the drug discovery approaches. Nat Prod Res 32(16):1926–1950
Bertuccioli A, Moricoli S, Amatori S, Rocchi MBL, Vici G, Sisti D (2020) Berberine and dyslipidemia: different applications and biopharmaceutical formulations without statin-like molecules-a meta-analysis. J Med Food 23(2):101–113
Bezáková L, Košťálová D, Obložinský M, Hoffman P, Pekárová M, Kollárová R et al (2014) Inhibition of 12/15 lipoxygenase by curcumin and an extract from Curcuma longa L. Ceska Slov Farm 63(1):26–31
Bhalani DV, Nutan B, Kumar A, Singh Chandel AK (2022) Bioavailability enhancement techniques for poorly aqueous soluble drugs and therapeutics. Biomedicines 10(9):2055
Bian Y, Liu P, Zhong J, Hu Y, Zhuang S, Fan K et al (2018) Quercetin Attenuates Adhesion Molecule Expression in Intestinal Microvascular Endothelial Cells by Modulating Multiple Pathways. Dig Dis Sci 63(12):3297–3304
Bleich SN, Sherrod C, Chiang A, Boyd C, Wolff J, DuGoff E et al (2015) Systematic review of programs treating high-need and high-cost people with multiple chronic diseases or disabilities in the United States, 2008–2014. Prev Chronic Dis 12:E197
Bolmont T, Haiss F, Eicke D, Radde R, Mathis CA, Klunk WE et al (2008) Dynamics of the microglial/amyloid interaction indicate a role in plaque maintenance. J Neurosci 28(16):4283–4292
Boschmann M, Thielecke F (2007) The effects of epigallocatechin-3-gallate on thermogenesis and fat oxidation in obese men: a pilot study. J Am Coll Nutr 26(4):389s-s395
Buhrmann C, Mobasheri A, Busch F, Aldinger C, Stahlmann R, Montaseri A et al (2011) Curcumin modulates nuclear factor kappaB (NF-kappaB)-mediated inflammation in human tenocytes in vitro: role of the phosphatidylinositol 3-kinase/Akt pathway. J Biol Chem 286(32):28556–28566
Buranrat B, Junking M (2022) Piperine suppresses growth and migration of human breast cancer cells through attenuation of Rac1 expression. Asian Pac J Trop Biomed 12(1):39–46
Chan WH, Wu HJ, Hsuuw YD (2005) Curcumin inhibits ROS formation and apoptosis in methylglyoxal-treated human hepatoma G2 cells. Ann N Y Acad Sci 1042:372–378
Chang CH, Huang WY, Lai CH, Hsu YM, Yao YH, Chen TY et al (2011) Development of novel nanoparticles shelled with heparin for berberine delivery to treat Helicobacter pylori. Acta Biomater 7(2):593–603
Chen H, Lu C, Liu H, Wang M, Zhao H, Yan Y et al (2017) Quercetin ameliorates imiquimod-induced psoriasis-like skin inflammation in mice via the NF-κB pathway. Int Immunopharmacol 48:110–117
Chen CY, Kao CL, Liu CM (2018) The cancer prevention, anti-inflammatory and anti-oxidation of bioactive phytochemicals targeting the TLR4 signaling pathway. Int J Mol Sci 19(9):2729
Chen X, Wu Q, Chen Y, Zhang J, Li H, Yang Z et al (2019) Diosmetin induces apoptosis and enhances the chemotherapeutic efficacy of paclitaxel in non-small cell lung cancer cells via Nrf2 inhibition. Br J Pharmacol 176(12):2079–2094
Chen M, Liu M, Luo Y, Cao J, Zeng F, Yang L et al (2022a) Celastrol protects against cerebral ischemia/reperfusion injury in mice by inhibiting glycolysis through targeting HIF-1α/PDK1 axis. Oxid Med Cell Longev 2022:7420507
Chen G, Zhu X, Li J, Zhang Y, Wang X, Zhang R et al (2022b) Celastrol inhibits lung cancer growth by triggering histone acetylation and acting synergically with HDAC inhibitors. Pharmacol Res 185:106487
Chimenti MS, Triggianese P, Conigliaro P, Candi E, Melino G, Perricone R (2015) The interplay between inflammation and metabolism in rheumatoid arthritis. Cell Death Dis 6(9):e1887
Chinta GB, Syed S, Coumar MS, Periyasamy L (2015) Piperine: a comprehensive review of pre-clinical and clinical investigations. Curr Bioactive Compd 11(3):156–69
Chugh R, Sangwan V, Patil SP, Dudeja V, Dawra RK, Banerjee S et al (2012) A preclinical evaluation of Minnelide as a therapeutic agent against pancreatic cancer. Sci Transl Med 4(156):156ra39
Cichewicz RH, Kouzi SA (2004) Chemistry, biological activity, and chemotherapeutic potential of betulinic acid for the prevention and treatment of cancer and HIV infection. Med Res Rev 24(1):90–114
Ciolino HP, Dankwah M, Yeh GC (2002) Resistance of MCF-7 cells to dimethylbenz(a)anthracene-induced apoptosis is due to reduced CYP1A1 expression. Int J Oncol 21(2):385–391
Clark PA, Bhattacharya S, Elmayan A, Darjatmoko SR, Thuro BA, Yan MB et al (2017) Resveratrol targeting of AKT and p53 in glioblastoma and glioblastoma stem-like cells to suppress growth and infiltration. J Neurosurg 126(5):1448–1460
Colotta F, Allavena P, Sica A, Garlanda C, Mantovani A (2009) Cancer-related inflammation, the seventh hallmark of cancer: links to genetic instability. Carcinogenesis 30(7):1073–1081
Coussens LM, Werb Z (2002) Inflammation and cancer. Nature 420(6917):860–867
Croia C, Bursi R, Sutera D, Petrelli F, Alunno A, Puxeddu I (2019) One year in review 2019: pathogenesis of rheumatoid arthritis. Clin Exp Rheumatol 37(3):347–357
Cui G, Qin X, Zhang Y, Gong Z, Ge B, Zang YQ (2009) Berberine differentially modulates the activities of ERK, p38 MAPK, and JNK to suppress Th17 and Th1 T cell differentiation in type 1 diabetic mice. J Biol Chem 284(41):28420–28429
de la Lastra CA, Villegas I (2007) Resveratrol as an antioxidant and pro-oxidant agent: mechanisms and clinical implications. Biochem Soc Trans 35(Pt 5):1156–1160
de Sá CD, Pacheco MT, Frozza RL, Bernardi A (2018) Anti-inflammatory effects of resveratrol: mechanistic insights. Int J Mol Sci 19(6):1812
Delecroix B, Abaïdia AE, Leduc C, Dawson B, Dupont G (2017) Curcumin and piperine supplementation and recovery following exercise induced muscle damage: a randomized controlled trial. J Sports Sci Med 16(1):147–153
Deng W, Du H, Liu D, Ma Z (2022) Editorial: the role of natural products in chronic inflammation. Front Pharmacol 13:901538
Dhingra D, Michael M, Rajput H, Patil RT (2012) Dietary fibre in foods: a review. J Food Sci Technol 49(3):255–266
Diakos CI, Charles KA, McMillan DC, Clarke SJ (2014) Cancer-related inflammation and treatment effectiveness. Lancet Oncol 15(11):e493-503
Dian L, Yu E, Chen X, Wen X, Zhang Z, Qin L et al (2014) Enhancing oral bioavailability of quercetin using novel soluplus polymeric micelles. Nanoscale Res Lett 9(1):2406
Dias DA, Urban S, Roessner U (2012) A historical overview of natural products in drug discovery. Metabolites 2(2):303–336
Dinarello CA (2010) Anti-inflammatory agents: present and future. Cell 140(6):935–950
Dyall SC, Balas L, Bazan NG, Brenna JT, Chiang N, da Costa SF et al (2022) Polyunsaturated fatty acids and fatty acid-derived lipid mediators: recent advances in the understanding of their biosynthesis, structures, and functions. Prog Lipid Res 86:101165
Edwards RL, Luis PB, Varuzza PV, Joseph AI, Presley SH, Chaturvedi R et al (2017) The anti-inflammatory activity of curcumin is mediated by its oxidative metabolites. J Biol Chem 292(52):21243–21252
El-Hadary AE, Ramadan MF (2019) Phenolic profiles, antihyperglycemic, antihyperlipidemic, and antioxidant properties of pomegranate (Punica granatum) peel extract. J Food Biochem 43(4):e12803
Fan D, He X, Bian Y, Guo Q, Zheng K, Zhao Y et al (2016) Triptolide modulates TREM-1 signal pathway to inhibit the inflammatory response in rheumatoid arthritis. Int J Mol Sci 17(4):498
Fan N, Zhao J, Zhao W, Shen Y, Song Q, Shum HC et al (2022) Biodegradable celastrol-loaded albumin nanoparticles ameliorate inflammation and lipid accumulation in diet-induced obese mice. Biomater Sci 10(4):984–996
Fang Z, He D, Yu B, Liu F, Zuo J, Li Y et al (2017) High-throughput study of the effects of celastrol on activated fibroblast-like synoviocytes from patients with rheumatoid arthritis. Genes 8(9):221
Fechtner S, Singh A, Chourasia M, Ahmed S (2017) Molecular insights into the differences in anti-inflammatory activities of green tea catechins on IL-1β signaling in rheumatoid arthritis synovial fibroblasts. Toxicol Appl Pharmacol 329:112–120
Flemming M, Kraus B, Rascle A, Jürgenliemk G, Fuchs S, Fürst R et al (2015) Revisited anti-inflammatory activity of matricine in vitro: comparison with chamazulene. Fitoterapia 106:122–128
Gandhi J, Khera L, Gaur N, Paul C, Kaul R (2017) Role of modulator of inflammation cyclooxygenase-2 in gammaherpesvirus mediated tumorigenesis. Front Microbiol 8:538
Gaston TE, Mendrick DL, Paine MF, Roe AL, Yeung CK (2020) “Natural” is not synonymous with “Safe”: toxicity of natural products alone and in combination with pharmaceutical agents. Regul Toxicol Pharmacol 113:104642
Gerritsen ME, Carley WW, Ranges GE, Shen CP, Phan SA, Ligon GF et al (1995) Flavonoids inhibit cytokine-induced endothelial cell adhesion protein gene expression. Am J Pathol 147(2):278–292
Giavasis I (2014) Bioactive fungal polysaccharides as potential functional ingredients in food and nutraceuticals. Curr Opin Biotechnol 26:162–173
Ginwala R, Bhavsar R, Chigbu DI, Jain P, Khan ZK (2019) Potential role of flavonoids in treating chronic inflammatory diseases with a special focus on the anti-inflammatory activity of apigenin. Antioxidants (basel, Switzerland) 8(2):35
Giordano A, Tommonaro G (2019) Curcumin and cancer. Nutrients 11(10):2376
Goel A, Boland CR, Chauhan DP (2001) Specific inhibition of cyclooxygenase-2 (COX-2) expression by dietary curcumin in HT-29 human colon cancer cells. Cancer Lett 172(2):111–118
Gomez JC, Yamada M, Martin JR, Dang H, Brickey WJ, Bergmeier W et al (2015) Mechanisms of interferon-γ production by neutrophils and its function during Streptococcus pneumoniae pneumonia. Am J Respir Cell Mol Biol 52(3):349–364
Greten FR, Grivennikov SI (2019) Inflammation and cancer: triggers, mechanisms, and consequences. Immunity 51(1):27–41
Grove KA, Lambert JD (2010) Laboratory, epidemiological, and human intervention studies show that tea (Camellia sinensis) may be useful in the prevention of obesity. J Nutr 140(3):446–453
Gunasekaran V, Avarachan J, Augustine A, Khayum A (2021) 3-O-Acetyl-11-keto-β-boswellic acid ameliorates acquired, consolidated and recognitive memory deficits through the regulation of hippocampal PPAR γ, MMP9 and MMP2 genes in dementia model. Heliyon 7(12):e08523
Guo MY, Li WY, Zhang Z, Qiu C, Li C, Deng G (2015) Betulin suppresses S. aureus-induced mammary gland inflammatory injury by regulating PPAR-γ in mice. Int Immunopharmacol 29(2):824–31
Guo D, Zhang W, Yang H, Bi J, Xie Y, Cheng B et al (2019) Celastrol induces necroptosis and ameliorates inflammation via targeting biglycan in human gastric carcinoma. Int J Mol Sci 20(22):5716
Guo G, Shi F, Zhu J, Shao Y, Gong W, Zhou G et al (2020) Piperine, a functional food alkaloid, exhibits inhibitory potential against TNBS-induced colitis via the inhibition of IκB-α/NF-κB and induces tight junction protein (claudin-1, occludin, and ZO-1) signaling pathway in experimental mice. Hum Exp Toxicol 39(4):477–491
Gupta RA, Motiwala MN, Dumore NG, Danao KR, Ganjare AB (2015) Effect of piperine on inhibition of FFA induced TLR4 mediated inflammation and amelioration of acetic acid induced ulcerative colitis in mice. J Ethnopharmacol 164:239–246
Gupta U, Sharma S, Khan I, Gothwal A, Sharma AK, Singh Y et al (2017) Enhanced apoptotic and anticancer potential of paclitaxel loaded biodegradable nanoparticles based on chitosan. Int J Biol Macromol 98:810–819
Haroyan A, Mukuchyan V, Mkrtchyan N, Minasyan N, Gasparyan S, Sargsyan A et al (2018) Efficacy and safety of curcumin and its combination with boswellic acid in osteoarthritis: a comparative, randomized, double-blind, placebo-controlled study. BMC Complement Altern Med 18(1):7
Hassanein EHM, Ibrahim IM, Abd-Alhameed EK, Mohamed NM, Ross SA (2022) Protective effects of berberine on various kidney diseases: emphasis on the promising effects and the underlined molecular mechanisms. Life Sci 306:120697
Heneka MT, Golenbock DT, Latz E (2015) Innate immunity in Alzheimer’s disease. Nat Immunol 16(3):229–236
Honari M, Shafabakhsh R, Reiter RJ, Mirzaei H, Asemi Z (2019) Resveratrol is a promising agent for colorectal cancer prevention and treatment: focus on molecular mechanisms. Cancer Cell Int 19:180
Hong H, Lu X, Lu Q, Huang C, Cui Z (2022) Potential therapeutic effects and pharmacological evidence of sinomenine in central nervous system disorders. Front Pharmacol 13:1015035
Hongqin T, Xinyu L, Heng G, Lanfang X, Yongfang W, Shasha S (2011) Triptolide inhibits IFN-γ signaling via the Jak/STAT pathway in HaCaT keratinocytes. Phytother Res 25(11):1678–1685
Hossain R, Kim KI, Jin F, Lee HJ, Lee CJ (2022) Betulin, an Anti-inflammatory triterpenoid compound, regulates MUC5AC mucin gene expression through NF-kB signaling in human airway epithelial cells. Biomol Ther 30(6):540–545
Hou W, Liu B, Xu H (2019) Triptolide: medicinal chemistry, chemical biology and clinical progress. Eur J Med Chem 176:378–392
Hour TC, Chen J, Huang CY, Guan JY, Lu SH, Pu YS (2002) Curcumin enhances cytotoxicity of chemotherapeutic agents in prostate cancer cells by inducing p21(WAF1/CIP1) and C/EBPbeta expressions and suppressing NF-kappaB activation. Prostate 51(3):211–218
Hu X, Li J, Fu M, Zhao X, Wang W (2021) The JAK/STAT signaling pathway: from bench to clinic. Signal Transduct Target Ther 6(1):402
Huang YY, Zhang Q, Zhang JN, Zhang YN, Gu L, Yang HM et al (2018) Triptolide up-regulates metabotropic glutamate receptor 5 to inhibit microglia activation in the lipopolysaccharide-induced model of Parkinson’s disease. Brain Behav Immun 71:93–107
Huang CF, Liu SH, Ho TJ, Lee KI, Fang KM, Lo WC et al (2022) Quercetin induces tongue squamous cell carcinoma cell apoptosis via the JNK activation-regulated ERK/GSK-3α/β-mediated mitochondria-dependent apoptotic signaling pathway. Oncol Lett 23(3):78
Hussain T, Murtaza G, Yang H, Kalhoro MS, Kalhoro DH (2020) Exploiting anti-inflammation effects of flavonoids in chronic inflammatory diseases. Curr Pharm Des 26(22):2610–2619
Jati GAK, Assihhah N, Wati AA, Salasia SIO (2022) Immunosuppression by piperine as a regulator of the NLRP3 inflammasome through MAPK/NF-κB in monosodium urate-induced rat gouty arthritis. Vet World 15(2):288–298
Jia L, Liu J, Song Z, Pan X, Chen L, Cui X et al (2012) Berberine suppresses amyloid-beta-induced inflammatory response in microglia by inhibiting nuclear factor-kappaB and mitogen-activated protein kinase signalling pathways. J Pharm Pharmacol 64(10):1510–1521
Jiang M, Li J, Peng Q, Liu Y, Liu W, Luo C et al (2014) Neuroprotective effects of bilobalide on cerebral ischemia and reperfusion injury are associated with inhibition of pro-inflammatory mediator production and down-regulation of JNK1/2 and p38 MAPK activation. J Neuroinflamm 11:167
Jiang H, Duan J, Xu K, Zhang W (2019) Resveratrol protects against asthma-induced airway inflammation and remodeling by inhibiting the HMGB1/TLR4/NF-κB pathway. Exp Ther Med 18(1):459–466
Jiao J, Xue B, Zhang L, Gong Y, Li K, Wang H et al (2008) Triptolide inhibits amyloid-β1-42-induced TNF-α and IL-1β production in cultured rat microglia. J Neuroimmunol 205(1):32–36
Jiao G, Yu G, Zhang J, Ewart HS (2011) Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar Drugs 9(2):196–223
Jiao P, Wang Y, Sang T, Jiao J, Li Y (2022) Molecular mechanism of betulin palliative therapy for chronic obstructive pulmonary disease (COPD) based on P2X7 receptor target of gated ion channel. Ann Transl Med 10(12):707
Johnson SM, Wang X, Evers BM (2011) Triptolide inhibits proliferation and migration of colon cancer cells by inhibition of cell cycle regulators and cytokine receptors. J Surg Res 168(2):197–205
Johnson JL, Jones MB, Cobb BA (2015) Polysaccharide A from the capsule of Bacteroides fragilis induces clonal CD4+ T cell expansion. J Biol Chem 290(8):5007–5014
Johnston GAR, Chebib M, Duke RK, Fernandez SP, Hanrahan JR, Hinton T et al (2009) Herbal products and GABA receptors. In: Squire LR (ed) Encyclopedia of neuroscience. Academic Press, Oxford, pp 1095–1101
Jonnalagadda SC, Suman P, Morgan DC, Seay JN (2017) Chapter 2: recent developments on the synthesis and applications of betulin and betulinic acid derivatives as therapeutic agents. In: Attaur R (ed) Studies in natural products chemistry, 53rd edn. Elsevier, pp 45–84
Joshi N, Singh S (2018) Updates on immunity and inflammation in Parkinson disease pathology. J Neurosci Res 96(3):379–390
Kamaraj Y, Dhayalan S, Chinnaiyan U, Kumaresan V, Subramaniyan S, Kumar D et al (2021) Triterpenoid compound betulin attenuates allergic airway inflammation by modulating antioxidants, inflammatory cytokines and tissue transglutaminase in ovalbumin-induced asthma mice model. J Pharm Pharmacol 73(7):968–978
Kant S, Swat W, Zhang S, Zhang ZY, Neel BG, Flavell RA et al (2011) TNF-stimulated MAP kinase activation mediated by a Rho family GTPase signaling pathway. Genes Dev 25(19):2069–2078
Kasikara C, Doran AC, Cai B, Tabas I (2018) The role of non-resolving inflammation in atherosclerosis. J Clin Investig 128(7):2713–2723
Khairani S, Fauziah N, Wiraswati HL, Panigoro R, Setyowati EY, Berbudi A (2022) Oral administration of piperine as curative and prophylaxis reduces parasitaemia in plasmodium berghei ANKA-infected mice. J Trop Med 2022:5721449
Kim SH, Park HJ, Lee CM, Choi IW, Moon DO, Roh HJ et al (2006) Epigallocatechin-3-gallate protects toluene diisocyanate-induced airway inflammation in a murine model of asthma. FEBS Lett 580(7):1883–1890
Kim DY, Park JW, Jeoung D, Ro JY (2009) Celastrol suppresses allergen-induced airway inflammation in a mouse allergic asthma model. Eur J Pharmacol 612(1–3):98–105
Kim JE, Lee MH, Nam DH, Song HK, Kang YS, Lee JE et al (2013) Celastrol, an NF-κB inhibitor, improves insulin resistance and attenuates renal injury in db/db mice. PLoS ONE 8(4):e62068
Kim SY, Hwangbo H, Kim MY, Ji SY, Kim DH, Lee H et al (2021) Betulinic acid restricts human bladder cancer cell proliferation in vitro by inducing caspase-dependent cell death and cell cycle arrest, and decreasing metastatic potential. Molecules (basel, Switzerland) 26(5):1381
Koopman M, El Aidy S (2017) Depressed gut? The microbiota-diet-inflammation trialogue in depression. Curr Opin Psychiatry 30(5):369–377
Krieglstein CF, Anthoni C, Rijcken EJ, Laukötter M, Spiegel HU, Boden SE et al (2001) Acetyl-11-keto-beta-boswellic acid, a constituent of a herbal medicine from Boswellia serrata resin, attenuates experimental ileitis. Int J Colorectal Dis 16(2):88–95
Krushkal J, Negi S, Yee LM, Evans JR, Grkovic T, Palmisano A et al (2021) Molecular genomic features associated with in vitro response of the NCI-60 cancer cell line panel to natural products. Mol Oncol 15(2):381–406
Kunnumakkara AB, Bordoloi D, Padmavathi G, Monisha J, Roy NK, Prasad S et al (2017) Curcumin, the golden nutraceutical: multitargeting for multiple chronic diseases. Br J Pharmacol 174(11):1325–1348
Lee YJ, Han SB, Nam SY, Oh KW, Hong JT (2010) Inflammation and Alzheimer’s disease. Arch Pharmacal Res 33(10):1539–1556
Lemieszek M, Rzeski W (2012) Anticancer properties of polysaccharides isolated from fungi of the Basidiomycetes class. Contemp Oncol (poznan, Poland) 16(4):285–289
Li Y, Yu C, Zhu WM, Xie Y, Qi X, Li N et al (2010) Triptolide ameliorates IL-10-deficient mice colitis by mechanisms involving suppression of IL-6/STAT3 signaling pathway and down-regulation of IL-17. Mol Immunol 47(15):2467–2474
Li C, Xi Y, Li S, Zhao Q, Cheng W, Wang Z et al (2015) Berberine ameliorates TNBS induced colitis by inhibiting inflammatory responses and Th1/Th17 differentiation. Mol Immunol 67(2 Pt B):444–54
Li Z, Zheng J, Zhang N, Li C (2016) Berberine improves airway inflammation and inhibits NF-κB signaling pathway in an ovalbumin-induced rat model of asthma. J Asthma 53(10):999–1005
Li P, Zheng Y, Chen X (2017) Drugs for autoimmune inflammatory diseases: from small molecule compounds to anti-TNF biologics. Front Pharmacol 8:460
Li B, Du P, Du Y, Zhao D, Cai Y, Yang Q et al (2021) Luteolin alleviates inflammation and modulates gut microbiota in ulcerative colitis rats. Life Sci 269:119008
Lin L, Sun Y, Wang D, Zheng S, Zhang J, Zheng C (2015) Celastrol ameliorates ulcerative colitis-related colorectal cancer in mice via suppressing inflammatory responses and epithelial-mesenchymal transition. Front Pharmacol 6:320
Ling D, Xia H, Park W, Hackett MJ, Song C, Na K et al (2014) pH-sensitive nanoformulated triptolide as a targeted therapeutic strategy for hepatocellular carcinoma. ACS Nano 8(8):8027–8039
Liu J, Ren H, Liu B, Zhang Q, Li M, Zhu R (2016) Diosmetin inhibits cell proliferation and induces apoptosis by regulating autophagy via the mammalian target of rapamycin pathway in hepatocellular carcinoma HepG2 cells. Oncol Lett 12(6):4385–4392
Liu W, Zhang Y, Zhu W, Ma C, Ruan J, Long H et al (2018) Sinomenine inhibits the progression of rheumatoid arthritis by regulating the secretion of inflammatory cytokines and monocyte/macrophage subsets. Front Immunol 9:2228
Liu J, Zhou H, Yang Z, Wang X, Chen H, Zhong L et al (2021) Rational construction of genome-reduced Burkholderiales chassis facilitates efficient heterologous production of natural products from proteobacteria. Nat Commun 12(1):4347
Lofrumento DD, Nicolardi G, Cianciulli A, De Nuccio F, La Pesa V, Carofiglio V et al (2014) Neuroprotective effects of resveratrol in an MPTP mouse model of Parkinson’s-like disease: possible role of SOCS-1 in reducing pro-inflammatory responses. Innate Immun 20(3):249–260
Luo L, Yang T (2016) Triptolide inhibits the progression of atherosclerosis in apolipoprotein E(-)/(-) mice. Exp Ther Med 12(4):2307–2313
Ma KC, Schenck EJ, Pabon MA, Choi AMK (2018) The role of danger signals in the pathogenesis and perpetuation of critical illness. Am J Respir Crit Care Med 197(3):300–309
Ma Z, Du B, Li J, Yang Y, Zhu F (2021) An insight into anti-inflammatory activities and inflammation related diseases of anthocyanins: a review of both in vivo and in vitro investigations. Int J Mol Sci 22(20):11076
Malaguarnera L (2019) Influence of resveratrol on the immune response. Nutrients 11(5):946
Mantovani A, Allavena P, Sica A, Balkwill F (2008) Cancer-related inflammation. Nature 454(7203):436–444
Mao D, Li H, Zhang L, Xu J, Yu C, Zhang Q (2019) Bilobalide alleviates IL-17-induced inflammatory injury in ATDC5 cells by downregulation of microRNA-125a. J Biochem Mol Toxicol 33(12):e22405
Martin DE, Salzwedel K, Allaway GP (2008) Bevirimat: a novel maturation inhibitor for the treatment of HIV-1 infection. Antiviral Chem Chemother 19(3):107–113
Martins LA, Coelho BP, Behr G, Pettenuzzo LF, Souza IC, Moreira JC et al (2014) Resveratrol induces pro-oxidant effects and time-dependent resistance to cytotoxicity in activated hepatic stellate cells. Cell Biochem Biophys 68(2):247–257
Mathy-Hartert M, Jacquemond-Collet I, Priem F, Sanchez C, Lambert C, Henrotin Y (2009) Curcumin inhibits pro-inflammatory mediators and metalloproteinase-3 production by chondrocytes. Inflamm Res 58(12):899–908
Mazmanian SK, Round JL, Kasper DL (2008) A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 453(7195):620–625
Mbese Z, Khwaza V, Aderibigbe BA (2019) Curcumin and its derivatives as potential therapeutic agents in prostate, colon and breast cancers. Molecules (basel, Switzerland) 24(23):4386
Mehany ABM, Belal A, Santali EY, Shaaban S, Abourehab MAS, El-Feky OA et al (2022) Biological effect of quercetin in repairing brain damage and cerebral changes in rats: molecular docking and in vivo studies. Biomed Res Int 2022:8962149
Meira CS, Barbosa-Filho JM, Lanfredi-Rangel A, Guimarães ET, Moreira DR, Soares MB (2016) Antiparasitic evaluation of betulinic acid derivatives reveals effective and selective anti-Trypanosoma cruzi inhibitors. Exp Parasitol 166:108–115
Michaud-Levesque J, Bousquet-Gagnon N, Béliveau R (2012) Quercetin abrogates IL-6/STAT3 signaling and inhibits glioblastoma cell line growth and migration. Exp Cell Res 318(8):925–935
Mims JW (2015) Asthma: definitions and pathophysiology. Int Forum Allergy Rhinol 5(Suppl 1):S2-6
Molinaro R, Corbo C, Livingston M, Evangelopoulos M, Parodi A, Boada C et al (2018) Inflammation and cancer: in medio stat nano. Curr Med Chem 25(34):4208–23
Moon PD, Choi IH, Kim HM (2011) Berberine inhibits the production of thymic stromal lymphopoietin by the blockade of caspase-1/NF-κB pathway in mast cells. Int Immunopharmacol 11(11):1954–1959
Nader MA, Baraka HN (2012) Effect of betulinic acid on neutrophil recruitment and inflammatory mediator expression in lipopolysaccharide-induced lung inflammation in rats. Eur J Pharm Sci 46(1–2):106–113
Netea MG, Balkwill F, Chonchol M, Cominelli F, Donath MY, Giamarellos-Bourboulis EJ et al (2017) A guiding map for inflammation. Nat Immunol 18(8):826–831
Nguyen NH, Khera R, Ohno-Machado L, Sandborn WJ, Singh S (2018) Annual burden and costs of hospitalization for high-need, high-cost patients with chronic gastrointestinal and liver diseases. Clin Gastroenterol Hepatol 16(8):1284–92.e30
Nguyen MT, Fernando S, Schwarz N, Tan JT, Bursill CA, Psaltis PJ (2019) Inflammation as a therapeutic target in atherosclerosis. J Clin Med 8(8):1109
Niu X, Zhao H, Zhu W, Zhao Y, Cao X, Xing L (2020) Efficacy of oral epigallocatechin-3-gallate solution administration during radiotherapy for non-small-cell lung cancer patients: a long-term observational study. Radiat Med Prot 01(02):94–8
Oak C, Khalifa AO, Isali I, Bhaskaran N, Walker E, Shukla S (2018) Diosmetin suppresses human prostate cancer cell proliferation through the induction of apoptosis and cell cycle arrest. Int J Oncol 53(2):835–843
Oliveira-Costa JF, Meira CS, Neves M, Dos Reis B, Soares MBP (2022) Anti-inflammatory activities of betulinic acid: a review. Front Pharmacol 13:883857
Panche AN, Diwan AD, Chandra SR (2016) Flavonoids: an overview. J Nutr Sci 5:e47
Pao HP, Liao WI, Wu SY, Hung KY, Huang KL, Chu SJ (2019) PG490-88, a derivative of triptolide, suppresses ischemia/reperfusion-induced lung damage by maintaining tight junction barriers and targeting multiple signaling pathways. Int Immunopharmacol 68:17–29
Park HJ, Lee CM, Jung ID, Lee JS, Jeong YI, Chang JH et al (2009) Quercetin regulates Th1/Th2 balance in a murine model of asthma. Int Immunopharmacol 9(3):261–267
Park JC, Yoo H, Kim CE, Shim SY, Lee M (2017) Hispidulin-7-O-neohesperidoside from Cirsium japonicum var. ussuriense attenuates the production of inflammatory mediators in LPS-induced raw 264.7 cells and HT-29 cells. Pharmacogn Mag 13(52):707–11
Patel K, Singh GK, Patel DK (2018) A review on pharmacological and analytical aspects of naringenin. Chin J Integr Med 24(7):551–560
Patra JK, Das G, Fraceto LF, Campos EVR, Rodriguez-Torres MDP, Acosta-Torres LS et al (2018) Nano based drug delivery systems: recent developments and future prospects. J Nanobiotechnol 16(1):71
Paulraj F, Abas F, Nordin HL, Othman I, Naidu R (2019) Molecular pathways modulated by curcumin analogue, diarylpentanoids in cancer. Biomolecules 9(7):270
Pervaiz S, Holme AL (2009) Resveratrol: its biologic targets and functional activity. Antioxid Redox Signal 11(11):2851–2897
Pulido-Moran M, Moreno-Fernandez J, Ramirez-Tortosa C, Ramirez-Tortosa M (2016) Curcumin and health. Molecules (basel, Switzerland) 21(3):264
Qadir MI, Naqvi ST, Muhammad SA (2016) Curcumin: a polyphenol with molecular targets for cancer control. Asian Pac J Cancer Prev 17(6):2735–2739
Qian L, Xu Z, Zhang W, Wilson B, Hong JS, Flood PM (2007) Sinomenine, a natural dextrorotatory morphinan analog, is anti-inflammatory and neuroprotective through inhibition of microglial NADPH oxidase. J Neuroinflamm 4:23
Qiu J, Yan Z, Tao K, Li Y, Li Y, Li J et al (2016) Sinomenine activates astrocytic dopamine D2 receptors and alleviates neuroinflammatory injury via the CRYAB/STAT3 pathway after ischemic stroke in mice. J Neuroinflamm 13(1):263
Rahban M, Habibi-Rezaei M, Mazaheri M, Saso L, Moosavi-Movahedi AA (2020) Anti-viral potential and modulation of Nrf2 by curcumin: pharmacological implications. Antioxidants (basel, Switzerland) 9(12):1228
Rakariyatham K, Du Z, Yuan B, Gao Z, Song M, Pan C et al (2019) Inhibitory effects of 7,7’-bromo-curcumin on 12-O-tetradecanoylphorbol-13-acetate-induced skin inflammation. Eur J Pharmacol 858:172479
Ramakrishna C, Kujawski M, Chu H, Li L, Mazmanian SK, Cantin EM (2019) Bacteroides fragilis polysaccharide A induces IL-10 secreting B and T cells that prevent viral encephalitis. Nat Commun 10(1):2153
Ren C, Jin J, Hu W, Chen Q, Yang J, Wu Y et al (2021) Betulin alleviates the inflammatory response in mouse chondrocytes and ameliorates osteoarthritis via AKT/Nrf2/HO-1/NF-κB Axis. Front Pharmacol 12:754038
Rengasamy KRR, Khan H, Gowrishankar S, Lagoa RJL, Mahomoodally FM, Khan Z et al (2019) The role of flavonoids in autoimmune diseases: therapeutic updates. Pharmacol Ther 194:107–131
Reno TA, Tong SW, Wu J, Fidler JM, Nelson R, Kim JY et al (2016) The triptolide derivative MRx102 inhibits Wnt pathway activation and has potent anti-tumor effects in lung cancer. BMC Cancer 16:439
Roy A, Roy M, Gacem A, Datta S, Zeyaullah M, Muzammil K et al (2022) Role of bioactive compounds in the treatment of hepatitis: a review. Front Pharmacol 13:1051751
Rubio-Perez JM, Morillas-Ruiz JM (2012) A review: inflammatory process in Alzheimer’s disease, role of cytokines. Sci World J 2012:756357
Safer AM, Leporatti S, Jose J, Soliman MS (2019) Conjugation of EGCG and chitosan NPs as a novel nano-drug delivery system. Int J Nanomed 14:8033–8046
Samsami-Kor M, Daryani NE, Asl PR, Hekmatdoost A (2015) Anti-Inflammatory effects of resveratrol in patients with ulcerative colitis: a randomized, double-blind, placebo-controlled pilot study. Arch Med Res 46(4):280–285
Scholey A, Downey LA, Ciorciari J, Pipingas A, Nolidin K, Finn M et al (2012) Acute neurocognitive effects of epigallocatechin gallate (EGCG). Appetite 58(2):767–770
Scott A, Khan KM, Cook JL, Duronio V (2004) What is “inflammation”? Are we ready to move beyond Celsus? Br J Sports Med 38(3):248–249
Sen T, Moulik S, Dutta A, Choudhury PR, Banerji A, Das S et al (2009) Multifunctional effect of epigallocatechin-3-gallate (EGCG) in downregulation of gelatinase-A (MMP-2) in human breast cancer cell line MCF-7. Life Sci 84(7–8):194–204
Serhan CN (2017) Treating inflammation and infection in the 21st century: new hints from decoding resolution mediators and mechanisms. FASEB J 31(4):1273–1288
Shaker ME, Ashamallah SA, Houssen ME (2014) Celastrol ameliorates murine colitis via modulating oxidative stress, inflammatory cytokines and intestinal homeostasis. Chem Biol Interact 210:26–33
Shang P, Liu W, Liu T, Zhang Y, Mu F, Zhu Z et al (2016) Acetyl-11-keto-β-boswellic acid attenuates prooxidant and profibrotic mechanisms involving transforming growth factor-β1, and improves vascular remodeling in spontaneously hypertensive rats. Sci Rep 6:39809
Shen J, Zhou Q, Li P, Wang Z, Liu S, He C et al (2017) Update on phytochemistry and pharmacology of naturally occurring resveratrol oligomers. Molecules (basel, Switzerland) 22(12):2050
Shi Y, Zhou J, Jiang B, Miao M (2017) Resveratrol and inflammatory bowel disease. Ann N Y Acad Sci 1403(1):38–47
Shi K, Chen X, Xie B, Yang SS, Liu D, Dai G et al (2018) Celastrol alleviates chronic obstructive pulmonary disease by inhibiting cellular inflammation induced by cigarette smoke via the Ednrb/Kng1 signaling pathway. Front Pharmacol 9:1276
Shi YM, Hirschmann M, Shi YN, Ahmed S, Abebew D, Tobias NJ et al (2022) Global analysis of biosynthetic gene clusters reveals conserved and unique natural products in entomopathogenic nematode-symbiotic bacteria. Nat Chem 14(6):701–712
Shishodia S, Chaturvedi MM, Aggarwal BB (2007) Role of curcumin in cancer therapy. Curr Probl Cancer 31(4):243–305
Shukla SM, Sharma SK (2011) Sinomenine inhibits microglial activation by Aβ and confers neuroprotection. J Neuroinflamm 8(1):117
Siddiqui IA, Adhami VM, Bharali DJ, Hafeez BB, Asim M, Khwaja SI et al (2009) Introducing nanochemoprevention as a novel approach for cancer control: proof of principle with green tea polyphenol epigallocatechin-3-gallate. Can Res 69(5):1712–1716
Sivani BM, Azzeh M, Patnaik R, Pantea Stoian A, Rizzo M, Banerjee Y (2022) Reconnoitering the therapeutic role of curcumin in disease prevention and treatment: lessons learnt and future directions. Metabolites 12(7):639
Skorupan N, Ahmad MI, Steinberg SM, Trepel JB, Cridebring D, Han H et al (2022) A phase II trial of the super-enhancer inhibitor Minnelide™ in advanced refractory adenosquamous carcinoma of the pancreas. Future Oncol (lond, Engl) 18(20):2475–2481
Smith PF, Ogundele A, Forrest A, Wilton J, Salzwedel K, Doto J et al (2007) Phase I and II study of the safety, virologic effect, and pharmacokinetics/pharmacodynamics of single-dose 3-o-(3’,3’-dimethylsuccinyl)betulinic acid (bevirimat) against human immunodeficiency virus infection. Antimicrob Agents Chemother 51(10):3574–3581
So AK, Martinon F (2017) Inflammation in gout: mechanisms and therapeutic targets. Nat Rev Rheumatol 13(11):639–647
Srisook K, Srisook E, Nachaiyo W, Chan-In M, Thongbai J, Wongyoo K et al (2015) Bioassay-guided isolation and mechanistic action of anti-inflammatory agents from Clerodendrum inerme leaves. J Ethnopharmacol 165:94–102
Sun F, Zhang Q, Zhao J, Zhang H, Zhai Q, Chen W (2019) A potential species of next-generation probiotics? The dark and light sides of Bacteroides fragilis in health. Food Res Int (ottawa, Ont) 126:108590
Taherzadeh D, Baradaran Rahimi V, Amiri H, Ehtiati S, Yahyazadeh R, Hashemy SI et al (2022) Acetyl-11-keto-β-boswellic acid (AKBA) prevents lipopolysaccharide-induced inflammation and cytotoxicity on H9C2 Cells. Evid-Based Complement Altern Med 2022:2620710
Takada Y, Ichikawa H, Badmaev V, Aggarwal BB (2006) Acetyl-11-keto-beta-boswellic acid potentiates apoptosis, inhibits invasion, and abolishes osteoclastogenesis by suppressing NF-kappa B and NF-kappa B-regulated gene expression. J Immunol (baltimore, Md 1950) 176(5):3127–40
Takahashi M, Uechi S, Takara K, Asikin Y, Wada K (2009) Evaluation of an oral carrier system in rats: bioavailability and antioxidant properties of liposome-encapsulated curcumin. J Agric Food Chem 57(19):9141–9146
Tanaka T, Narazaki M, Kishimoto T (2014) IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol 6(10):a016295
Teruya T, Konishi T, Uechi S, Tamaki H, Tako M (2007) Anti-proliferative activity of oversulfated fucoidan from commercially cultured Cladosiphon okamuranus TOKIDA in U937 cells. Int J Biol Macromol 41(3):221–226
Testa G, Gamba P, Badilli U, Gargiulo S, Maina M, Guina T et al (2014) Loading into nanoparticles improves quercetin’s efficacy in preventing neuroinflammation induced by oxysterols. PLoS ONE 9(5):e96795
Thomford NE, Senthebane DA, Rowe A, Munro D, Seele P, Maroyi A et al (2018) Natural products for drug discovery in the 21st century: innovations for novel drug discovery. Int J Mol Sci 19(6):1578
Tocmo R, Veenstra JP, Huang Y, Johnson JJ (2021) Covalent modification of proteins by plant-derived natural products: proteomic approaches and biological impacts. Proteomics 21(3–4):e1900386
Tong X, Pelling JC (2013) Targeting the PI3K/Akt/mTOR axis by apigenin for cancer prevention. Anticancer Agents Med Chem 13(7):971–978
Tong L, Zhao Q, Datan E, Lin GQ, Minn I, Pomper MG et al (2021) Triptolide: reflections on two decades of research and prospects for the future. Nat Prod Rep 38(4):843–860
Tsao JP, Bernard JR, Hsu HC, Hsu CL, Liao SF, Cheng IS (2022) Short-term oral quercetin supplementation improves post-exercise insulin sensitivity, antioxidant capacity and enhances subsequent cycling time to exhaustion in healthy adults: a pilot study. Front Nutr 9:875319
Tungmunnithum D, Thongboonyou A, Pholboon A, Yangsabai A (2018) Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: an overview. Medicines (basel, Switzerland) 5(3):492
Villarroya F, Cereijo R, Gavaldà-Navarro A, Villarroya J, Giralt M (2018) Inflammation of brown/beige adipose tissues in obesity and metabolic disease. J Intern Med 284(5):492–504
Wajant H, Pfizenmaier K, Scheurich P (2003) Tumor necrosis factor signaling. Cell Death Differ 10(1):45–65
Wang P, Li Q, Li K, Zhang X, Han Z, Wang J et al (2012) Betulinic acid exerts immunoregulation and anti-tumor effect on cervical carcinoma (U14) tumor-bearing mice. Pharmazie 67(8):733–739
Wang WY, Tan MS, Yu JT, Tan L (2015) Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann Transl Med 3(10):136
Wang C, Liu B, Xu X, Zhuang B, Li H, Yin J et al (2016) Toward targeted therapy in chemotherapy-resistant pancreatic cancer with a smart triptolide nanomedicine. Oncotarget 7(7):8360–8372
Wang XL, Li T, Li JH, Miao SY, Xiao XZ (2017a) The effects of resveratrol on inflammation and oxidative stress in a rat model of chronic obstructive pulmonary disease. Molecules (basel, Switzerland) 22(9):1529
Wang X, He X, Zhang CF, Guo CR, Wang CZ, Yuan CS (2017b) Anti-arthritic effect of berberine on adjuvant-induced rheumatoid arthritis in rats. Biomed Pharmacother 89:887–93
Wang MX, Zhao JX, Meng YJ, Di TT, Xu XL, Xie XJ et al (2018a) Acetyl-11-keto-β-boswellic acid inhibits the secretion of cytokines by dendritic cells via the TLR7/8 pathway in an imiquimod-induced psoriasis mouse model and in vitro. Life Sci 207:90–104
Wang Y, Feng Q, He P, Zhu L, Chen G (2018b) Genomics approach of the natural product pharmacology for high impact diseases. Int J Genom 2018:9468912
Wang Q, Ye C, Sun S, Li R, Shi X, Wang S et al (2019a) Curcumin attenuates collagen-induced rat arthritis via anti-inflammatory and apoptotic effects. Int Immunopharmacol 72:292–300
Wang W, Zha G, Zou JJ, Wang X, Li CN, Wu XJ (2019b) Berberine attenuates cigarette smoke extract-induced airway inflammation in mice: involvement of TGF-β1/Smads signaling pathway. Curr Med Sci 39(5):748–753
Wang RX, Zhou M, Ma HL, Qiao YB, Li QS (2021a) The role of chronic inflammation in various diseases and anti-inflammatory therapies containing natural products. ChemMedChem 16(10):1576–1592
Wang X, Ye C, Xun T, Mo L, Tong Y, Ni W et al (2021b) Bacteroides fragilis polysaccharide a ameliorates abnormal voriconazole metabolism accompanied with the inhibition of TLR4/NF-κB pathway. Front Pharmacol 12:663325
Wang S, Hu G, Chen L, Ma K, Hu C, Zhu H et al (2022) Celastrol acts as a new histone deacetylase inhibitor to inhibit colorectal cancer cell growth via regulating macrophage polarity. Cell Biol Int. https://doi.org/10.1002/cbin.11952
Wei C, Fan J, Sun X, Yao J, Guo Y, Zhou B et al (2020) Acetyl-11-keto-β-boswellic acid ameliorates cognitive deficits and reduces amyloid-β levels in APPswe/PS1dE9 mice through antioxidant and anti-inflammatory pathways. Free Radical Biol Med 150:96–108
Wiederschain GY (2007) Polysaccharides. Structural diversity and functional versatility. Biochem Mosc 72(6):675
Wijesinghe WAJP, Jeon Y-J (2012) Biological activities and potential industrial applications of fucose rich sulfated polysaccharides and fucoidans isolated from brown seaweeds: a review. Carbohydr Polym 88(1):13–20
Witvrouw M, De Clercq E (1997) Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. Gen Pharmacol 29(4):497–511
Wu S, Pang Y, He Y, Zhang X, Peng L, Guo J et al (2021) A comprehensive review of natural products against atopic dermatitis: flavonoids, alkaloids, terpenes, glycosides and other compounds. Biomed Pharmacother 140:111741
Wu Y, Li K, Zeng M, Qiao B, Zhou B (2022) Serum metabolomics analysis of the anti-inflammatory effects of gallic acid on rats with acute inflammation. Front Pharmacol 13:830439
Xia B, Li Q, Wu J, Yuan X, Wang F, Lu X et al (2022) Sinomenine confers protection against myocardial ischemia reperfusion injury by preventing oxidative stress, cellular apoptosis, and inflammation. Front Pharmacol 13:922484
Xiong H, Tian L, Zhao Z, Chen S, Zhao Q, Hong J et al (2017) The sinomenine enteric-coated microspheres suppressed the TLR/NF-κB signaling in DSS-induced experimental colitis. Int Immunopharmacol 50:251–262
Yadav VR, Prasad S, Sung B, Gelovani JG, Guha S, Krishnan S et al (2012) Boswellic acid inhibits growth and metastasis of human colorectal cancer in orthotopic mouse model by downregulating inflammatory, proliferative, invasive and angiogenic biomarkers. Int J Cancer 130(9):2176–2184
Yang Y, Zhang X, Xu M, Wu X, Zhao F, Zhao C (2018) Quercetin attenuates collagen-induced arthritis by restoration of Th17/Treg balance and activation of Heme Oxygenase 1-mediated anti-inflammatory effect. Int Immunopharmacol 54:153–162
Yi J, Zhu R, Wu J, Wu J, Xia W, Zhu L et al (2016) In vivo protective effect of betulinic acid on dexamethasone induced thymocyte apoptosis by reducing oxidative stress. Pharmacol Rep 68(1):95–100
Yi L, Lyn YJ, Peng C, Zhu RL, Bai SS, Liu L et al (2018) Sinomenine inhibits fibroblast-like synoviocyte proliferation by regulating α7nAChR expression via ERK/Egr-1 pathway. Int Immunopharmacol 56:65–70
Yuan J, Liu R, Ma Y, Zhang Z, Xie Z (2018) Curcumin attenuates airway inflammation and airway remolding by inhibiting NF-κB signaling and COX-2 in cigarette smoke-induced COPD mice. Inflammation 41(5):1804–1814
Yuan F, Dong H, Gong J, Wang D, Hu M, Huang W et al (2019) A systematic review and meta-analysis of randomized controlled trials on the effects of turmeric and curcuminoids on blood lipids in adults with metabolic diseases. Adv Nutr (bethesda, Md) 10(5):791–802
Zhan XK, Li JL, Zhang S, Xing PY, Xia MF (2018) Betulinic acid exerts potent antitumor effects on paclitaxel-resistant human lung carcinoma cells (H460) via G2/M phase cell cycle arrest and induction of mitochondrial apoptosis. Oncol Lett 16(3):3628–3634
Zhang Y, Kong H, Fang Y, Nishinari K, Phillips GO (2013) Schizophyllan: a review on its structure, properties, bioactivities and recent developments. Bioactive Carbohydr Diet Fibre 1(1):53–71
Zhang SY, Zhao QF, Fang NN, Yu JG (2015) Betulin inhibits pro-inflammatory cytokines expression through activation STAT3 signaling pathway in human cardiac cells. Eur Rev Med Pharmacol Sci 19(3):455–460
Zhang S, Liu X, Mei L, Wang H, Fang F (2016) Epigallocatechin-3-gallate (EGCG) inhibits imiquimod-induced psoriasis-like inflammation of BALB/c mice. BMC Complement Altern Med 16(1):334
Zhang B, Yin X, Sui S (2018a) Resveratrol inhibited the progression of human hepatocellular carcinoma by inducing autophagy via regulating p53 and the phosphoinositide 3-kinase/protein kinase B pathway. Oncol Rep 40(5):2758–2765
Zhang WL, Zhao YN, Shi ZZ, Cong D, Bai YS (2018b) Lutein inhibits cell growth and activates apoptosis via the PI3K/AKT/mTOR signaling pathway in a549 human non-small-cell lung cancer cells. J Environ Pathol Toxicol Oncol 37(4):341–350
Zhang B, Zhong Q, Chen X, Wu X, Sha R, Song G et al (2020) Neuroprotective effects of celastrol on transient global cerebral ischemia rats via regulating HMGB1/NF-κB signaling pathway. Front Neurosci 14:847
Zhao H, Liu Z, Liu W, Han X, Zhao M (2016) Betulin attenuates lung and liver injuries in sepsis. Int Immunopharmacol 30:50–56
Zhao H, Wang Q, Cheng X, Li X, Li N, Liu T et al (2018a) Inhibitive Effect of Resveratrol on the Inflammation in Cultured Astrocytes and Microglia Induced by Aβ(1–42). Neuroscience 379:390–404
Zhao J, Li R, Pawlak A, Henklewska M, Sysak A, Wen L et al (2018b) Antitumor activity of betulinic acid and betulin in canine cancer cell lines. In Vivo (athens, Greece) 32(5):1081–1088
Zhi YK, Li J, Yi L, Zhu RL, Luo JF, Shi QP et al (2022) Sinomenine inhibits macrophage M1 polarization by downregulating α7nAChR via a feedback pathway of α7nAChR/ERK/Egr-1. Phytomedicine 100:154050
Zhong K, Zhang Q, Tong L, Liu L, Zhou X, Zhou S (2015a) Molecular weight degradation and rheological properties of schizophyllan under ultrasonic treatment. Ultrason Sonochem 23:75–80
Zhong K, Tong L, Liu L, Zhou X, Liu X, Zhang Q et al (2015b) Immunoregulatory and antitumor activity of schizophyllan under ultrasonic treatment. Int J Biol Macromol 80:302–308
Zhou Y, Zhang T, Wang X, Wei X, Chen Y, Guo L et al (2015) Curcumin modulates macrophage polarization through the inhibition of the toll-like receptor 4 expression and its signaling pathways. Cell Physiol Biochem 36(2):631–641
Zhou Z, Tang M, Liu Y, Zhang Z, Lu R, Lu J (2017) Apigenin inhibits cell proliferation, migration, and invasion by targeting Akt in the A549 human lung cancer cell line. Anticancer Drugs 28(4):446–456
Zhu X, Liu Q, Wang M, Liang M, Yang X, Xu X et al (2011) Activation of Sirt1 by resveratrol inhibits TNF-α induced inflammation in fibroblasts. PLoS ONE 6(11):e27081
Zhu T, Chen Z, Chen G, Wang D, Tang S, Deng H et al (2019) Curcumin attenuates asthmatic airway inflammation and mucus hypersecretion involving a PPARγ-dependent NF-κB signaling pathway in vivo and in vitro. Mediat Inflamm 2019:4927430
Zhu W, Zhao Y, Zhang S, Li X, Xing L, Zhao H et al (2021) Evaluation of epigallocatechin-3-gallate as a radioprotective agent during radiotherapy of lung cancer patients: a 5-year survival analysis of a phase 2 study. Front Oncol 11:686950
Zoi V, Galani V, Lianos GD, Voulgaris S, Kyritsis AP, Alexiou GA (2021) The role of curcumin in cancer treatment. Biomedicines 9(9):1086
Funding
The authors would like to thank the Deanship of Scientific Research at Umm Al-Qura University for supporting this work by Grant Code: (24UQU4290565DSR007).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gouda, N.A., Alshammari, S.O., Abourehab, M.A.S. et al. Therapeutic potential of natural products in inflammation: underlying molecular mechanisms, clinical outcomes, technological advances, and future perspectives. Inflammopharmacol 31, 2857–2883 (2023). https://doi.org/10.1007/s10787-023-01366-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10787-023-01366-y