
Abstract
Primates living in high-latitude habitats are expected to employ feeding strategies appropriate to harsh environments characterized by strong seasonality in temperature and food resources. We studied diet, feeding behavior, and seasonal changes in food availability in wild Taihangshan macaques (Macaca mulatta tcheliensis) living in a northern montane habitat (800–1700 m) in Mt. Tiantanshan area, Jiyuan, north China. From June 2016 to May 2017, we collected data on the diet and feeding behavior of one group of macaques and measured food availability in twenty 20-m × 20-m plots in their home range. We identified 105 plant species eaten by the macaques, belonging to 44 genera and 30 families. The macaques’ diet comprised nine major categories: seeds (45.9% ± 31.2%), tree leaves (23.3% ± 27.2%), herbaceous leaves (15.3% ± 11.0%), young bark (5.6% ± 10.1%), fruits (2.5% ± 3.4%), buds (2.4% ± 5.5%), flowers (1.9% ± 3.4%), herb roots (1.8% ± 2.2%), and young twigs (1.5% ± 1.9%). We found significant seasonal variation in the dietary proportion of leaves, seeds, herbaceous roots, and young twigs. The proportion of each food type consumed was positively correlated with its availability in the habitat, but preferred food types varied monthly and seasonally. Seeds were the most preferred food, while tree leaves and herbaceous leaves served as fallback foods when seed availability was low. Faced with the scarcity of food resources and strongly seasonal changes in their high-latitude habitat, Taihangshan macaques employed a seed- and leaf-based feeding strategy, reflecting their ecological and evolutionary adaptation to temperate forests.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Primates originated in tropical rainforest areas and radiated into temperate regions (Grueter et al., 2009; Hanya et al., 2011). Their feeding strategies correspond to local environments (Li, 2006; Ma et al., 2017; Owens et al., 2015; Zhang et al., 2022). Tropical forest-living primates mainly feed on fruit. For example, long-tailed macaques (M. fascicularis) and toque macaques (M. sinica) are frugivores, and fruits account for 40–80% of their annual diets (Berenstain, 1986). Fruits account for more than 70% of the diet in Sulawesi crested black macaques (M. nigra) (OBrien & Kinnaird, 1997), 68% in lion-tailed macaques (M. silenus), and 82% in Tonkean macaques (M. tonkeana) (Riley, 2007).
Temperate forests have a significant annual resource-scarce period compared with tropical forests, which can last several months (Lu et al., 2007; Tsuji et al., 2013). In particular, high-latitude montane forests tend to be less productive than low-latitude areas, with decreased plant densities, lower temperatures, and reduced rainfall (Lomolino, 2001), and lower availability of high-quality foods during the resource-scarce period (Cui et al., 2019; Lu et al., 2007). Primates are severely affected by these challenges, especially in winter (Grueter et al., 2009; Xiang et al., 2010). Furthermore, fruiting and flushing seasons, fall and spring, are shorter than in tropical forests (Tsuji et al., 2013). Many temperate primates eat young leaves, flowers, fruits, and seeds when available and rely on mature leaves, buds, lichen, and tree barks in food-scarce seasons (Cui et al., 2019; Grueter et al., 2009; Hanya, 2004; Lu et al., 2007). Primates prioritize preferred foods and adjust their food selection to consume fallback foods (foods with lower nutrients and energy return but which are abundantly distributed in the environment) when the preferred foods are scarce (Hemingway & Bynum, 2005; Lambert & Rothman, 2015).
Being the most widely distributed non-human primate species worldwide, rhesus macaques (M. mulatta) range from tropical to temperate regions (Cui et al., 2019). Taihangshan macaques (M. tcheliensis), an endemic subspecies of rhesus macaque to China, inhabit the southern Mt. Taihangshan area, currently the northernmost zone of the natural distribution of rhesus macaques (Cui et al., 2019, 2020; Guo et al., 2020; Lu et al., 2007). Under the influence of a typical continental monsoon climate, the southern Mt. Taihangshan area experiences notable fluctuations in temperature, rainfall, and other ecological variables (Cui et al., 2020; Lu et al., 2007), influencing the foraging activities of the macaques, especially in winter and early spring with snow and lower food availability (Lu et al., 2007). Taihangshan macaques employ a flexible nutritional strategy according to changes in the availability of food resources, and seeds play an important role in their annual diets (35.1% dry matter intake) (Cui et al., 2018, 2019, 2020). Seeds dominate the diets of Taihangshan macaques in autumn when seeds are most available, whereas leaves and herbaceous plants gradually replace seeds as seed availability decreases from winter and spring to summer (Cui et al., 2019).
In this study, we revealed the foraging adaptations of Taihangshan macaques to the temperate forest environment. We investigated 1) the seasonal dietary composition of Taihangshan macaques in greater detail than previous studies; and 2) the relationship between food types and availability of preferred and fallback food resources.
Method
Ethical Note
This research complied with the protocols approved by the Wildlife Protection Law of China. This research adhered to the requirement of Chinese laws and the principles for the ethical treatment of primates formulated by the American Society of Primatologists.
Data Availability
All data generated or analyzed during this study are included in this article. Further inquiries can be directed to the corresponding author.
Conflicts of Interests
The authors declare that they have no conflict of interest.
Study Sites
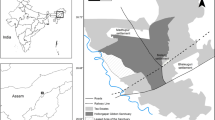
We conducted this study in the Mt. Tiantanshan area of Taihangshan Macaque National Nature Reserve (34°54′—35°42′ N, 112°02′—113°45′ E), Jiyuan, China from June 2016 to May 2017. This site, in the core area of the TMNNR with the highest elevation of 1,715 m (summit of Mt. Tiantanshan), belongs to the low-middle mountain landform type. The vegetation is dominated by natural secondary forests, and the dominant species is Quercus variabilis (Lu et al., 2007). Influenced by the continental monsoon climate, four distinct seasons, spring (from March to May), summer (from June to August), autumn (from September to November), and winter (from December to February of the next year) occur at the study site (Xie et al., 2012). During the study, we monitored temperatures and rainfall monthly by using a thermometer (Deli 9013; − 30–50 ℃; ± 2 ℃) and a rain gauge (SM-1; 205 mm × 650 mm), and consulted the local weather station to obtain the annual snowfall (210.5 mm) and snow depth (5.5 mm). The annual mean temperature was 15.5 °C. The highest mean monthly temperature was 28.0 °C in July, whereas the lowest was 1.5 °C in January. The annual rainfall was 578.8 mm, with rain in summer (327.2 mm) and drought in winter (36.4 mm) (Fig. 1).

Monthly mean temperature and rainfall in the Mt. Tiantanshan area of Taihangshan Macaque National Nature Reserve, Jiyuan, China from June 2016 to May 2017
Study Group
We studied a wild group of Taihangshan macaques inhabiting the Mt. Tiantanshan area (the home range is approximately 24 km2) (Cui et al., 2019). This group, named WW-1, has been identified since 2005 and is accustomed to close observation by researchers (ca 2 m) (Cui et al., 2019). During our study, the group consisted of 100 individuals, including seven adult males, 28 adult females, 43 juveniles, and 22 infants. We focused on individuals who could forage independently to observe and record their feeding behaviors.
Behavioral Observations
We followed and observed group WW-1 for 3–5 days each month and collected behavioral data. We used instantaneous scan sampling to observe and record the behaviors of every visible individual for 5 min in each 15-min cycle (Altmann, 1974). To avoid sampling bias toward certain age-sex groups, we scanned the group, either from left to right when they were resting, or from front to back, according to the direction of the group’s travel, and collected behavioral data for each visible individual. We recorded the food plant species and parts eaten. We also determined that macaques did not digest fruit seeds based on entire seeds observed in the feces. We used focal animal sampling to collect the fresh feces of macaque individuals who swallowed the seeds in fruits and transported the fecal samples to the laboratory for centrifugation after cryopreservation to separate the seeds.
We identified and categorized all items into nine food types: seeds (nuts, fruit seeds, and herbaceous seeds), leaves (leaves of trees and shrubs), herbaceous leaves, buds, fruits, herbaceous roots, young bark, young twigs, and flowers. We excluded insects and their eggs from the analysis, because they were rare in the macaques’ diet.
Diet and Food Availability
We identified the plant species eaten by the macaques based on behavioral records from June 2016 to May 2017 and foraging traces in a 1-km × 2-km transect (8% of the home range) where we randomly established twenty 20-m × 20-m vegetation plots in the core range of group WW-1 (Cui et al., 2019). To identify species eaten, we compared the food species with the plant resource survey records for Taihangshan Macaque National Nature Reserve (Lu et al., 2007). For species that were difficult to identify in situ, we collected fresh samples of the parts eaten and determined the species using the Flora of China online (www.iplant.cn). We recorded 1536 plant species, including 664 trees with a diameter at breast height (DBH) ≥ 5 cm, 342 shrubs (including trees with DBH < 5 cm), 69 vines, and 461 herbaceous plants to estimate the abundance of the nine food types. We calculated the mean basal area of trees using the formula: \(B={(0.5\times \mathrm{DBH})}^{2}\times \pi\). In the middle of each month, we estimated the phenology of the study area based on the proportion of feeding parts in the crown area of all the trees: 0 = absent (no fruit/seeds); 1 = 1–25%; 2 = 26–50%; 3 = 51–75%; 4 = 76–100% of the crown area (Albert et al., 2013). We calculated the food availability index (FAI) of the main food types (leaves, buds, young barks, seeds, young twigs, flowers, and fruits) using the following formula: \(\mathrm{FAI}={\sum }_{i=1}^{n}{D}_{i}{B}_{i}{P}_{i}\), where Di is the density of the species i (number of stem/ha), Bi is the mean basal area of food plant species i (m2/ha), and Pi is the mean phenology score of the available food types provided by food plant species i each month (Albert et al., 2013). To estimate the FAI for herbs, we set up twenty 2-m × 2-m plots randomly along the foraging routes of the group and recorded the numbers of individuals of herbs and their relative cover with the same 5-point score as for phenology (Cui et al., 2019). Characterization of arboreal food resources does not completely describe the food availability for the macaques as they also consumed terrestrial food items, such as fallen Quercus seeds. To quantify seasonal variation in the availability of seeds on the ground, we used the same 5-point scale as above. The abundance of the Quercus seeds reached its maximum in September, so we scored this as 4.
Data Analysis
We used behavioral observation, fecal samples, and feeding traces to identify the plant species and parts eaten. We treated each scan as an independent data point. We first calculated the percentage of scanned individuals feeding on each food type/species out of the total number of individuals recorded in each scan and then divided this value by the percentage of all individuals feeding (Hanya & Bernard, 2012). To avoid bias in the number of food species/types and feeding time budget due to uneven data records in different months, we took the mean data for each hour and then the mean hourly data for each month to determine the monthly percentage of feeding time. We expressed seasonal and annual dietary composition as the mean proportion across the relevant months (Tang et al., 2016).
We used GraphPad Prism (version 9) for data analysis and plotting. We tested the normality of variables with Kolmogorov–Smirnov and Shapiro–Wilk tests. We used the formula \({P}_{i}=\sum {C}_{i}/\sum F\) to calculate the proportion of different food types in the diet, where i is the type of food plants, Pi is the proportion of food type i in the feeding records, \(\sum {C}_{i}\) is the sum of feeding records spent on type i, and \(\sum F\) is the sum of total feeding records. We used the Shannon–Wiener Diversity Index, \({H}^{^{\prime}}=-\sum_{i=1}^{n}{P}_{i}\mathrm{ln}{P}_{i}\), where \({P}_{i}\) is the proportion of type i in the feeding records, to express the dietary breadth of food plants available for macaques (Huang et al., 2015). We divided preferred foods by the preference scores (\({E}_{i}\)) according to the formula \({E}_{i}=({r}_{i}-{n}_{i})/({r}_{i}+{n}_{i})\), where \({r}_{i}\) is the percentage of species i in the diet (based on feeding time), and \({n}_{i}\) is the percentage of species i in the food availability (calculated from 20 plots). \({E}_{i}\) values range from − 1 to 1. When the index is > 0 and seasonal consumption is positively correlated with availability, we regard it as a preferred food. We classified foods whose consumption increases when preferred foods decrease as fallback foods (Grueter et al., 2009; Lambert & Rothman, 2015; Zhang et al., 2022).
We used Kruskal–Wallis tests to examine the differences in dietary composition among four seasons and Mann–Whitney U tests to examine the differences in dietary composition of seeds between two seasons. We used Kruskal–Wallis tests and Mann–Whitney U tests to examine differences in Shannon–Wiener Diversity Index, FAI, and selective index among different seasons (Cui et al., 2019). We tested the relationship between percentages of feeding time and FAI for all food types using Spearman’s rank correlation analysis. We expressed all seasonal data as mean ± SD. All tests were two-tailed with a significance level of 0.05. We used a significance level of 0.0125 for Kruskal–Wallis tests, adjusting the alpha to account for multiple testing.
Results
We identified 185 species of plants in the plots, belonging to 41 families and 88 genera. Taihangshan macaques ate 105 of these species, including 28 species of trees, five species of vines, 34 species of shrubs, and 38 species of herbaceous plants (Supplementary Table S1).
Annual Dietary Composition
We collected 6886 feeding records for Taihangshan macaques during the study. Seeds accounted for 45.9% ± 31.2% of the diet (range: 0–82.5%, mostly in September and October). Tree leaves (young and mature leaves) accounted for 23.3% ± 27.2% (range: 0–63.7%). Herbaceous leaves accounted for 15.3% ± 11.0% (range: 7.6–36.3%, mainly in summer) and were the only food type eaten across the whole year. The other food types were young bark (5.6% ± 10.1%, range: 0–24.5%), fruits (2.5% ± 3.4%, range: 0–8.8%), and buds (2.4% ± 5.5%, range: 0–14.7%), flowers (1.9% ± 3.3%, range: 0–8.1%), herbaceous roots (1.8% ± 2.2%, range: 0–6.0%), and young twigs (1.5% ± 1.9%, range: 0–5.2%) (Fig. 2).

Annual diet composition of Taihangshan macaques in the Mt. Tiantanshan area of Taihangshan Macaque National Nature Reserve, Jiyuan, China, from June 2016 to May 2017
Seasonal Variation in the Diet
There were significant seasonal differences in the proportion of different food types eaten (Table I). Leaves were the highest in summer and declined to zero in winter, seeds were the highest in autumn and decreased to zero in summer, herbaceous roots were only consumed in autumn and winter, and young twigs were only consumed in winter and early spring (Fig. 3). There were no significant seasonal changes in the proportion of other food types: herbaceous leaves, buds, fruits, young bark, and flowers (Table I).

Seasonal variation in the monthly diets of Taihangshan macaques at the Mt. Tiantanshan area of Taihangshan Macaque National Nature Reserve, Jiyuan, China, from June 2016 to May 2017
In summer, tree leaves were macaques’ primary food type, which accounted for more than 55% of their summer diet, when seed availability dropped to zero. Followed by herbaceous leaves (> 30%) (Fig. 3). In autumn, seeds matured and occupied the highest proportion of autumn food composition (Table I), becoming the main food eaten by macaques (> 80%), followed by herbaceous leaves and fruits (both < 10% and > 5%) (Fig. 3). In winter, the proportion of seeds declined compared with autumn but was still higher than in spring and summer (Table S3). The macaques spent most of their feeding time (> 55%) on seeds in winter (Fig. 3). In spring, the proportion of seeds consumed decreased to 35.9%, whereas the proportion of young leaves increased to 48.5%, leaves gradually exceeded seeds, and became the dominant food for macaques in spring with the emergence of young leaves (Fig. 3).
The proportion of seeds consumed was significantly negatively correlated with that of leaves and herbaceous leaves across months, and significantly positively correlated with herbaceous root consumption (Table II). The consumption of leaves was significantly negatively correlated with that of seeds, herbaceous roots, young bark, and twigs (Table II). In addition, there were significantly positive correlations between herbaceous roots and young barks, and between young barks and twigs (Table II).
There were no significant differences in the food diversity index (Kruskal–Wallis test: χ2 = 6.510, degree of freedom (df) = 3, P = 0.070) and food availability index (Kruskal–Wallis test: χ2 = 5.359, df = 3, P = 0.151) among seasons (Tables III and IV).
There was no significant correlation between food diversity and availability in different months of the year (Spearman’s rank correlation analysis: n = 12, r = 0.186, p = 0.561). Consumption of all food types was positively correlated with the proportion of resources available in the habitat (Table V; Fig. 4).

Relationship between percentage of feeding time and food availability index for different food types used by Taihangshan macaques in the Mt. Tiantanshan area of Taihangshan Macaque National Nature Reserve, Jiyuan, China, from June 2016 to May 2017. Circles represent the percentage of feeding time for each food type, and triangles the percentage of FAI for each food type. The background colors represent summer, autumn, winter, and spring from left to right
Food Preferences
Among all food species, 20 genera accounted for more than 1% of the records and contributed 89.4% of annual feeding records (Supplementary Table S2). Preference for food items varied by months and seasons (Fig. 5; Table VI).

Monthly variation in preferences scores for food types fed on by Taihangshan macaques in the Mt. Tiantanshan area of Taihangshan Macaque National Nature Reserve, Jiyuan, China, from June 2016 to May 2017. The background colors of plots represent summer, autumn, winter, and spring from left to right
Taihangshan macaques consumed leaves from March to October, and the preference for leaves increased gradually from avoidance to preference (Fig. 5). There were sufficient young and mature leaves for macaques to feed on in spring and summer, respectively. However, the preference for leaves was higher in autumn than in spring (Fig. 5). Macaques preferred feeding on herbaceous leaves in spring but avoided them in autumn (Fig. 5). They mainly ate buds in March and April with high preference scores and preferred to eat seeds when they were available, especially in autumn (Fig. 5). Fruit was the preferred food type in August, but not in autumn and winter (Fig. 5). The consumption of herbaceous roots was concentrated in autumn and winter with high preference scores (Fig. 5). Macaques ate young bark and twigs in winter although they were not the preferred food (Fig. 5). Flowers were only consumed in spring when most plants bloomed (Fig. 5).
Discussion
Taihangshan macaques ate 146 food items from 105 food species, including nine food categories. Compared with other species of Macaca, the number of food species used is remarkably high. For example, long-tailed macaques, living in the tropical rainforest in Indonesia, feed on 29 species of plants (Nila et al., 2014). Bonnet macaques and lion-tailed macaques in the Western Ghats of India consume 37 and 24 species of plants, respectively (Roy et al., 2011). Rhesus macaques, inhabiting southern China, eat 60 species yearly (Zhou et al., 2009). Macaques in similar latitudes to our study group, in northwestern Pakistan, feed on just 35 plant species (Goldstein & Richard, 1989). Japanese macaques (M. fuscata) in Yakushima consume 93 species of plants in lowland forests in coastal areas (Hill, 1997). Japanese macaques living in coniferous forests eat 59 kinds of plants (Hanya, 2004). In high latitudes, food for macaques is extremely limited (Zhang et al., 2022). The total number of food species eaten by Japanese macaques was 21 in central Japan (Wada & Ichiki, 1980), and female Japanese macaques ate 60 different food items in northern Japan (Tsuji & Takatsuki, 2012). In temperate forests, plant productivity is lower than that in tropical regions, and food availability varies significantly with season (Grueter et al., 2009). The timing of the flushing and fruiting is predictable and shorter than that in tropical forests (Hanya et al., 2013). Therefore, macaques have to rely on more food species and types to survive in high-latitude habitats (Fan et al., 2012; Zhang et al., 2021, 2022).
The main food types chosen by Taihangshan macaques were seeds, leaves, and herbaceous plants including herbaceous leaves and roots. This result differs from the foods chosen by other Macaca species in temperate and alpine forests in Japan, in which the major food types are fruits (48%), foliage (25%), and animal matter (7%) (Tsuji et al., 2013). Taihangshan macaques fed on seeds (mainly the acorns of Quercus trees) that were rich in nutrition and energy, and available in the Mt. Taihangshan area (Cui et al., 2018, 2019). Being the dominant species in local forests, Quercus variabilis produced a large number of seeds in autumn (Cui et al., 2019; Lu et al., 2007). As time passed, seeds gradually became more difficult to find when they were covered by dead leaves or snow, forcing the macaques to consume tree bark, which is available but has lower nutrient content. Snow depth and snow duration were the primary determinants of dietary diversity in Japanese macaques, which had less diverse diets in forests with deep snow (Tsuji et al., 2015). Macaques fed less on foliage, seeds, and fruits but ate more bark and buds with more snow depth and longer snow duration (Tsuji et al., 2015). Taihangshan macaques also spent longer feeding and looking for seeds in winter and early spring than in summer (Cui et al., 2019; this study).
Taihangshan macaques consumed more leaves in spring (young leaves) and summer (mature leaves) than in autumn and winter. Young leaves were available in spring, and contain high protein and low cellulose, which can make up for the energy deficit caused by seed shortage (Zhang et al., 2022). In summer, leaves became the primary food type (> 60%) for Taihanghshan macaques, filling the gap left by the decreased seed availability. Tree leaves and herbaceous leaves appear to be fallback foods for Taihangshan macaques; the selectivity indexes of leaves and herbaceous leaves both significantly negatively correlated with the selectivity index of seeds, and the availability of leaves significantly negatively correlated with the availability of seeds. Other temperate Macaca species also consume large amounts of leaves in summer as fallback foods (Tsuji et al., 2013; Zhang et al., 2021, 2022).
Fruits accounted for 2.48% of the yearly diets of Taihangshan macaques, which was far below the average proportion for Asian Macaca species (48%, range: 8.5–87.0%) (Tsuji et al., 2013) and below that of rhesus macaques at a similar latitude in Pakistan (8.5%) (Goldstein & Richard, 1989). This pattern may derive from differences in vegetation structure between low and high-latitude forests (Cui et al., 2019, 2020; Hanya et al., 2011; Huang et al., 2015; Tsuji et al., 2013; Zhang et al., 2022). Furthermore, during periods of food shortage, particularly in early spring and winter, Taihangshan macaques also fed on herbaceous leaves, roots, as well as the young barks of small shrubs (Cui et al., 2019; Lu et al., 2007).
Conclusions
Taihangshan macaques have significant seasonal variation in their diet. During the seed-rich season (autumn), macaques mainly feed on seeds and reduced the consumption of other food types. In seed-scarce seasons, tree leaves and herb leaves serve as fallback foods. We conclude that Taihangshan macaques exhibit a flexible seed- and leaf-based feeding strategy that makes them an ecological generalist and allows them to inhabit temperate forests.
References
Albert, A., Hambuckers, A., Culot, L., Savini, T., & Huynen, M. C. (2013). Frugivory and seed dispersal by northern pigtailed macaques (Macaca leonina), in Thailand. International Journal of Primatology, 34(1), 170–193. https://doi.org/10.1007/s10764-012-9649-5
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49(3), 227–267.
Berenstain, L. (1986). Primates in nature - Richard,Af. Ethology and Sociobiology, 7(1), 57–62. https://doi.org/10.1016/0162-3095(86)90015-4
Cui, Z. W., Shao, Q., Grueter, C. C., Wang, Z. L., Lu, J. Q., & Raubenheimer, D. (2019). Dietary diversity of an ecological and macronutritional generalist primate in a harsh high-latitude habitat, the Taihangshan macaque (Macaca mulatta tcheliensis). American Journal of Primatology, 81(4). https://doi.org/10.1002/ajp.22965
Cui, Z. W., Wang, Z. L., Shao, Q., Raubenheimer, D., & Lu, J. Q. (2018). Macronutrient signature of dietary generalism in an ecologically diverse primate in the wild. Behavioral Ecology, 29(4), 804–813. https://doi.org/10.1093/beheco/ary003
Cui, Z. W., Wang, Z. L., Zhang, S. Q., Wang, B. S., Lu, J. Q., & Raubenheimer, D. (2020). Living near the limits: Effects of interannual variation in food availability on diet and reproduction in a temperate primate, the Taihangshan macaque (Macaca mulatta tcheliensis). American Journal of Primatology, 82(1). https://doi.org/10.1002/ajp.23080
Fan, P. F., Fei, H. L., & Ma, C. Y. (2012). Behavioral responses of Cao Vit gibbon (Nomascus Nasutus) to variations in food abundance and temperature in Bangliang, Jingxi, China. American Journal of Primatology, 74(7), 632–641. https://doi.org/10.1002/ajp.22016
Goldstein, S. J., & Richard, A. F. (1989). Ecology of rhesus macaques (Macaca mulatta) in northwest Pakistan. International Journal of Primatology, 10(6), 531–567. https://doi.org/10.1007/Bf02739364
Grueter, C. C., Li, D. Y., Ren, B. P., Wei, F. W., Xiang, Z. F., & van Schaik, C. P. (2009). Fallback foods of temperate-living primates: A case study on snub-nosed monkeys. American Journal of Physical Anthropology, 140(4), 700–715. https://doi.org/10.1002/ajpa.21024
Guo, Y. M., Garber, P. A., Tian, J. D., & Lu, J. Q. (2020). Neonatal nipple preference and maternal cradling laterality in wild Taihangshan macaques (Macaca mulatta tcheliensis). American Journal of Primatology, 82(12). https://doi.org/10.1002/ajp.23197
Hanya, G. (2004). Diet of a Japanese macaque troop in the coniferous forest of Yakushima. International Journal of Primatology, 25(1), 55–71. https://doi.org/10.1023/B:Ijop.0000014645.78610.32
Hanya, G., & Bernard, H. (2012). Fallback foods of red leaf monkeys (Presbytis rubicunda) in Danum Valley, Borneo. International Journal of Primatology, 33(2), 322–337. https://doi.org/10.1007/s10764-012-9580-9
Hanya, G., Menard, N., Qarro, M., Ibn Tattou, M., Fuse, M., Vallet, D., Yamada, A., Go, M., Takafumi, H., Tsujino, R., Agetsuma, N., & Wada, K. (2011). Dietary adaptations of temperate primates: Comparisons of Japanese and Barbary macaques. Primates, 52(2), 187–198. https://doi.org/10.1007/s10329-011-0239-5
Hanya, G., Tsuji, Y., & Grueter, C. C. (2013). Fruiting and flushing phenology in Asian tropical and temperate forests: Implications for primate ecology. Primates, 54(2), 101–110. https://doi.org/10.1007/s10329-012-0341-3
Hemingway, C. A., & Bynum, N. (2005). The influence of seassonally on primate diet and ranging. In C. P. van Schaik & D. K. Brockman (Eds.), Seasonality in primates: Studies of living and extinct human and non-human primates (pp. 57–104). Cambridge University Press.
Hill, D. A. (1997). Seasonal variation in the feeding behavior and diet of Japanese macaques (Macaca fuscata yakui) in lowland forest of Yakushima. American Journal of Primatology, 43(4), 305–322.
Huang, Z. H., Huang, C. M., Tang, C. B., Huang, L. B., Tang, H. X., Ma, G. Z., & Zhou, Q. H. (2015). Dietary adaptations of assamese macaques (Macaca assamensis) in limestone forests in southwest China. American Journal of Primatology, 77(2), 171–185. https://doi.org/10.1002/ajp.22320
Lambert, J. E., & Rothman, J. M. (2015). Fallback foods, optimal diets, and nutritional targets: Primate responses to varying food availability and quality. Annual Review of Anthropology, 44(44), 493–512. https://doi.org/10.1146/annurev-anthro-102313-025928
Li, Y. M. (2006). Seasonal variation of diet and food availability in a group of Sichuan snub-nosed monkeys in Shennongjia Nature Reserve, China. American Journal of Primatology, 68(3), 217–233. https://doi.org/10.1002/ajp.20220
Lomolino, M. V. (2001). Elevation gradients of species-density: Historical and prospective views. Global Ecology and Biogeography, 10(1), 3–13. https://doi.org/10.1046/j.1466-822x.2001.00229.x
Lu, J. Q., Hou, J. H., Wang, H. F., & Qu, W. Y. (2007). Current status of Macaca mulatta in Taihangshan mountains area, Jiyuan, Henan, China. International Journal of Primatology, 28(5), 1085–1091. https://doi.org/10.1007/s10764-007-9197-6
Ma, C., Fan, P. F., Zhang, Z. Y., Li, J. H., Shi, X. C., & Xiao, W. (2017). Diet and feeding behavior of a group of 42 Phayre's langurs in a seasonal habitat in Mt. Gaoligong, Yunnan, China. American Journal of Primatology, 79(10). https://doi.org/10.1002/ajp.22695
Nila, S., Suryobroto, B., & Widayati, K. A. (2014). Dietary variation of long tailed macaques (Macaca fascicularis) in Telaga Warna, Bogor, West Java. HAYATI Journal of Biosciences, 21(1), 8–14. https://doi.org/10.4308/hjb.21.1.8
OBrien, T. G., & Kinnaird, M. F. (1997). Behavior, diet, and movements of the Sulawesi crested black macaque (Macaca nigra). International Journal of Primatology, 18(3), 321-351.https://doi.org/10.1023/A:1026330332061
Owens, J. R., Honarvar, S., Nessel, M., & Hearn, G. W. (2015). From frugivore to folivore: Altitudinal variations in the diet and feeding ecology of the Bioko Island drill (Mandrillus leucophaeus poensis). American Journal of Primatology, 77(12), 1263–1275. https://doi.org/10.1002/ajp.22479
Riley, E. P. (2007). Flexibility in diet and activity patterns of Macaca tonkeana in response to anthropogenic habitat alteration. International Journal of Primatology, 28(1), 107–133. https://doi.org/10.1007/s10764-006-9104-6
Roy, K., Singh, M., & Singh, M. (2011). Diet and dietary-niche breadth of diurnal rain forest primates in the Central Western Ghats, India. Folia Primatologica, 82(6), 283–298. https://doi.org/10.1159/000336964
Tang, C. B., Huang, L. B., Huang, Z. H., Krzton, A., Lu, C. H., & Zhou, Q. H. (2016). Forest seasonality shapes diet of limestone-living rhesus macaques at Nonggang, China. Primates, 57(1), 83–92. https://doi.org/10.1007/s10329-015-0498-7
Tsuji, Y., & Takatsuki, S. (2012). Interannual Variation in Nut Abundance Is Related to Agonistic Interactions of Foraging Female Japanese Macaques (Macaca fuscata). International Journal of Primatology, 33, 489–512. https://doi.org/10.1007/s10764-012-9589-0
Tsuji, Y., Hanya, G., & Grueter, C. C. (2013). Feeding strategies of primates in temperate and alpine forests: Comparison of Asian macaques and colobines. Primates, 54(3), 201–215. https://doi.org/10.1007/s10329-013-0359-1
Tsuji, Y., Ito, T. Y., Wada, K., & Watanabe, K. (2015). Spatial patterns in the diet of Japanese macaques. Mammal Review, 45(4), 227–238. https://doi.org/10.1111/mam.12045
Wada, K., & Ichiki, Y. (1980). Seasonal home range use by Japanese monkeys in the snowy Shiga Heights. Primates, 21, 468–483. https://doi.org/10.1007/BF02373835
Xiang, Z. F., Huo, S., & Xiao, W. (2010). Activity budget of Rhinopithecus bieti at Tibet: Effects of day length, temperature and food availability. Current Zoology, 56(6), 650–659. https://doi.org/10.1093/czoolo/56.6.650
Xie, D. M., Lu, J. Q., Sichilima, A. M., & Wang, B. S. (2012). Patterns of habitat selection and use by Macaca mulatta tcheliensis in winter and early spring in temperate forest, Jiyuan, China. Biologia, 67(1), 234–239. https://doi.org/10.2478/s11756-011-0156-5
Zhang, C., Zhou, Q. H., Xu, H. L., & Huang, Z. H. (2021). Diet, food availability, and climatic factors drive ranging behavior in white-headed langurs in the limestone forests of Guangxi, southwest China. Zoological Research, 42(4), 406–411. https://doi.org/10.24272/j.issn.2095-8137.2020.292
Zhang, K. C., Karim, F., Jin, Z. X., Xiao, H. T., Yao, Y. F., Ni, Q. Y., Li, B. J., Pu-Cuo, W. J., Huang, Z. H., & Xu, H. L. (2022). Diet and feeding behavior of a group of high-altitude rhesus macaques: High adaptation to food shortages and seasonal fluctuations. Current Zoology. https://doi.org/10.1093/cz/zoac047
Zhou, Q. H., Tang, H. X., Wei, C. Q., & Huang, C. M. (2009). Diet and seasonal changes in Rhesus macaques (Macaca mulatta) at Seven-star Park, Guilin. Acta Theriologica Sinica, 29(4), 419–426. https://doi.org/10.3969/j.issn.1000-1050.2009.04.011
Acknowledgements
We are grateful to the staff of State-owned Jiyuan Yugong Forest Farm for their logistic support. We are also grateful to Mr. Guoliang Zhao for his assistance in the field works. We thank the Editors and Reviewers for reviewing and improving the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (No. 31472018, 31672302, 32070446).
Author information
Authors and Affiliations
Contributions
JL conceived and designed the experiments. QS, ZC, and CL performed the experiments. QS, ZC, and JT analyzed the data. QS was a major contributor in writing the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Statement of Ethics
This research complied with the protocols approved by the Wildlife Protection Law of China. This research adhered to the requirement of Chinese laws and the principles for the ethical treatment of primates formulated by the American Society of Primatologists (ASP).
Inclusion and Diversity Statement
The author list includes contributors from the location where the research was conducted, who participated in study conception, study design, data collection, and/or interpretation of findings.
Additional information
Handling Editor: Songtao Guo
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Shao, Q., Cui, Z., Liu, C. et al. Diets and Feeding Strategy in Taihangshan Macaques (Macaca mulatta tcheliensis) in a Temperate Forest, North China. Int J Primatol 44, 1074–1090 (2023). https://doi.org/10.1007/s10764-023-00379-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-023-00379-2