
Abstract
Fruit pulp is an easily handled energy source for many frugivorous species but generally has little protein. Accordingly, ripe-fruit specialist primate species with diets dominated by fruit pulp risk protein deficiency. While some species use leaf and flower buds, young leaves, and arthropods as an alternative protein supplement, highly frugivorous spider monkeys (Ateles spp.) use protein-rich young leaves and/or fig fruits. However, not all spider monkey populations have access to abundantly available figs. Comparing infestation frequencies of fruits on trees with those eaten by spider monkeys, we tested the hypothesis that, under such circumstances, spider monkeys preferentially choose those nonfig fruits with pulp infested by insect larvae (a highly protein-rich resource). We predicted that: (i) a large proportion of plant species eaten by Ateles would have insect larvae-infested fruits; and (ii) Ateles would actively select infested fruits. We tested these predictions with Ateles chamek and Ateles marginatus on the banks of the Tapajós River, Brazil. Across a 13-month sampling period, we recorded 27 plant species in the diet of the 2 Ateles species. Of these, 23 (85%) had larvae-infested fruits when sampled; 11 species (40%) had high levels of individual fruits infested (35-78%). We used Ivlev Values to quantify selectivity for infested/uninfested fruits in 20 plant species. Infested fruits were positively selected in 12 species (60%), while aversion to infested fruits occurred in 4 species (20%). This covert carnivory/faunivory in spider monkeys is a largely overlooked aspect of their feeding ecology. This situation would be nearly impossible to ascertain from behavioral observations alone, showing the value of integrated, multimethod approaches. The strategy used by Ateles spp. on the banks of the Tapajós highlights the flexibility of primate foraging choices and the importance of indirect source of protein to ripe-fruit specialist primates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Zoochory provides ecological benefits from individual to ecosystem levels (Howe, 1986). It is the most common seed dispersal syndrome for many tropical forests (Moermond & Denslow, 1983), and notably so in the tropical Americas (Fleming & Kress, 2011). Tree species with fruit adapted for zoochory often are large and abundant (Fricke et al., 2013), especially in lowland rainforests (Bello et al., 2015). Large-sized fruits with succulent pulp characterize such trees, and the pulp represents a large proportion of the fruit’s total mass (Uriarte et al., 2011). The pulp is generally rich in sugars but poor in protein and free amino acids (Table I). Apart from those of such families as the Lauraceae (Jordano, 1995) and Myristicaceae (Kato, 1995), such fruits are typically low in lipids (Simmen & Sabatier, 1996) (Table I). These nutritional limitations pose a problem for primary consumers trying to achieve a diet that is both energetically adequate and nutritionally balanced (Felton, Felton, Lindenmayer, & Foley, 2009a; Oftedal, 1991; Raubenheimer & Simpson, 1997). For example, a general estimate for primates in the tropical Americas is that they require at least 15% of crude protein in their diet (Ange-van Heugten, 2008), while known diets of large-bodied species, for example, yield at most ~8% protein (e.g., Ateles geoffroyi (Kuhl, 1820) and Ateles paniscus (Linnaeus, 1858): Hladik et al., 1971; Simmen & Sabatier, 1996).
Within the atelids (Platyrrhini: Atelidae), members of the genus Ateles are the most specialized for eating sweet, lipid-rich ripe fruit (Di Fiore et al., 2008). In comparison, the closely related woolly monkeys (Lagothrix) eat more sugary fruits, but include a higher proportion of ripe seeds and, especially, arthropods in their diets (Gonzalez et al., 2016; Stevenson et al., 1994). In fact, arthropod consumption may constitute up to 41% of feeding time in some populations (e.g., Lagothrix lagothricha lugens; Cifuentes et al., 2013). The other two remaining large atelids, muriquis (Brachyteles) and howler monkeys (Alouatta), include large proportions of leaves in their diet; some howler monkey species have fruits as a secondary (Julliot & Sabatier, 1993; Simmen & Sabatier, 1996), or even primary (Dunn et al., 2009; Garber et al., 2015; Martins, 2008), diet component. Although atelids may switch seasonally to such items as arthropods, buds, flowers, and young leaves, species of spider monkeys generally maintain diets of up to 90% ripe fruit, whereas only 5%, on average, is devoted to seeds, bark, fungi, termitaria soil, ground soil, and other undetermined food items (Di Fiore et al., 2008). Thus, while spider monkey species may be key dispersers for a number of zoochorous trees in the tropical Americas (Arroyo-Rodríguez et al., 2017; Link & Di Fiore, 2006; Ponce-Santizo et al., 2006), the mainstay of their diet may not allow it to attain a nutritionally balanced intake (sensu, Raubenheimer et al., 2009).
It has been suggested that Ateles might address protein limitations in their diets by consuming protein-rich young leaves (Simmen & Sabatier, 1996) and protein-rich fruits (Russo et al., 2005), as well as by adjusting time allocation patterns (Di Fiore & Rodman, 2001), movement dynamics, and foraging adaptations (Norconk & Kinzey, 1994). However, it has been proposed that such mechanisms cannot explain the patterns of nutrient intake observed, at least for the black-faced black spider monkey, Ateles chamek (Humboldt, 1812; Felton, Felton, Raubenheimer, et al., 2009b). Individuals of this species employ a protein leverage system that maintains a stable daily protein intake while permitting total energy intake to vary as a function of the composition of available food items (Felton, Felton, Raubenheimer, et al., 2009b). At the sites studied by Felton et al. (2009a, b) consumption of figs was key to this system, since their pollination ecology means that ripe fig fruit obligatorily contain not only the developing larvae of agaonid Hymenoptera (fig-wasps), but also the wingless males (Weiblen, 2002) and the parasitoids of both life-stages (Dsouza & Ravishankar, 2014).
Although protein content can influence food-item choice for many animals, comparisons of such content are generally made between plant species (Dasilva, 1994; Gautier-Hion et al., 1984; Hemingway, 1998; McConkey et al., 2002; Stevenson, 2004). Within-species comparisons are rarely made, except for fruit size (Dias da Silva et al., 2020) or ontogeny related-changes (e.g., lipid and sugar content of ripe vs. unripe fruit: Masette et al., 2015; Worman & Chapman, 2005; Chapman, 2005), although there are exceptions (e.g., inter-tree variation in crop size: Houle et al., 2007; age-related variation in fruit nutrient content: Carlson et al., 2013; 2014; Ryan et al., 2013). However, insect infestation of fruits, and its capacity to raise protein levels, is rarely considered as a variable, even though it occurs quite commonly in tropical tree species, where almost all succulent fruit have some form of infesting fruit fly (Ajuja & Liedo, 1993). As many Coleoptera, Lepidoptera, and symphytic Hymenoptera (sawflies) also have larval stages that develop in fruit, a given fruit may have several invertebrate species present at one time (Devescovi et al., 2015). Unfortunately, due to the differential palatability of invertebrate taxa (e.g., the capacity of some, but not others, to secrete noxious chemicals: Laurent et al., 2005), the presence of multiple invertebrate species within the fruit may serve as a complicating factor during dietary analyses (Bravo, 2012; Jordano, 1987). This may complicate efforts to improve our understanding of protein intake via ingestion of insects within fruit. This particular behavior was termed "covert carnivory," and it was predicted that it would be found in other diet-specialist primate species (Barnett et al., 2017). The hypothesis that primates might gain protein in this way was originally proposed by Redford et al. (1984), but this insight received little attention at the time. Although McGrew (2014) introduced the term “faunivory” for the ingestion of species substantially smaller than the consumer, we feel our phrasing of “covert carnivory” is both accurate and euphonious.
Protein-restricted diets may be features of primates with diets whose main elements are low in protein. Examples of these are unripe seeds (Barnett et al., 2017) and fruit pulp (Simmen & Sabatier, 1996). The Pitheciinae subfamily of the Pitheciidae, which comprises of the uacaris (Cacajao), cuxiús (Chiropotes), and sakis (Pithecia), is an example of the former, being specialist seed predators with diets composed of 60-80% unripe seeds (Norconk, 2007). This type of resource is low in protein, because storage protein is deposited in the seed only as it nears maturity, thus leaving only harder-to-digest structural proteins available in younger seeds (Hill & Breidenbach, 1974). The solution adopted by pitheciins is to select those seeds infested with insect larvae (Cacajao: Ballantyne, 2018; Barnett et al., 2017; Chiropotes: Barnett et al., n.d.). Such larvae can contribute up to 86% of a seed’s weight (Barnett et al., 2017). Given the high protein and lipid levels of larval insects (Bukkens, 1997; Table II), they are likely to provide a substantial nutrient supplement (Barnett et al., 2017). For Cacajao ouakary (Spix 1823), Barnett et al. (2017) reported that 26% of species eaten for their seeds included infested seeds; together, these comprised 40.8% of seed-feeding records. Of the 19 statistically evaluated species in that study, uacaris showed active selection for 47% of infested fruit species (Barnett et al., 2017).
Members of the genus Ateles are an example of the second group, with fruit pulp comprising up to 70% of the diet (Di Fiore et al., 2008; Stevenson & Link, 2010), where protein supplements are considered to come from shoots and buds (Cant, 1990).
Insectivory is generally considered a minor component of Ateles diets, not in the least because Ateles have specializations for arboreal locomotion, such as long, curved fingers and a vestigial thumb (Nelson & Boeving, 2015). (In fact, the name "Ateles" derives from the Greek ατελής for "imperfect": Rosenberger et al., 2008). While the hand’s structure is sufficient to manipulate fruit (Dew, 2005), it may compromise the fine levels of dexterity necessary to capture free-living animals, because a full precision grip is an anatomically demanding task (Lindshield & Rodrigues, 2009). Therefore, when insectivory is reported in Ateles species, it is limited to a few free-living (i.e., nonembedded) species, such as of leaf-rolling caterpillars, meliponine (stingless) bees, and termites (Di Fiore et al., 2008). It also is only ever considered a minor, or supplementary, diet component (González-Zamora et al., 2009; van Roosmalen & Klein, 1988), although individual bouts of insectivory may be protracted (Link, 2003).
Given the diet specialization on ripe fruits with succulent pulp by the Ateles clade, we hypothesize that Ateles, like the seed specialists pitheciins, actively select infested fruits to offset protein shortfalls in their diets. It would appear that such requirements may well be met by the protein content of fig fruits, at least at some sites (Felton et al., 2009a, b in Bolivia). However, there are parts of Amazonia where figs are rare (Berg et al., 1984; Pitman et al., 2001; Wittmann et al., 2006). At such locations, therefore, given the differences between the of ateline and pitheciin diets, the selection by Ateles will involve insect larvae in the pulp, rather than within seeds. Consequently, we tested the following predictions: (i) a considerable proportion of fruit species eaten by Ateles will be infested with insect larvae at the maturational stage at which they are consumed; and (ii) such fruit will be actively selected by Ateles so that they appear at a higher percentage in the diet than they do on the tree.
Methods
Study Site and Study Species
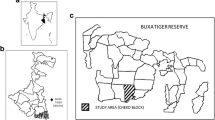
We conducted the study in two sites along the mid-Tapajós River, including the mouth of Jamanxim River, Pará State, Brazil (Fig. 1), an area where fig trees are uncommon (Pinto, 2008). We collected data along the margins of igapó seasonally flooded forest (sensu, Prance, 1979), and in the never-flooded terra firme forest bordering the igapó, using a system of predefined trails.

Location of study site within South America (inset), positions of sampling locations at the survey site, and the presence of the two Ateles study species, one on either side of the Tapajós River, Brazil (main panel). Ateles images by Stephan Nash, and used with permission.
We studied two species of Ateles. Following the taxonomy of Morales-Jimenez et al. (2015), these were black-faced black spider monkey (Ateles chamek, found on the western bank of the Tapajós River) and white-cheeked spider monkey (A. marginatus, found on the eastern bank).
Data Collection
Monkeys are messy eaters (Howe, 1980), and large volumes of partially-eaten fruits generally accumulate under feeding trees. These fruits are commonly used in ecological studies (Russo, 2003; Wehncke et al., 2003). To distinguish such fruits from whole ones that have been knocked down by the movements of feeding animals, or by other elements, we continue the use of the term “ort,” defined as “a fragment of food, fallen from a table. A meal remnant” (see https://www.merriam-webster.com/dictionary/ort), first proposed by Barnett et al. (2017).
We performed spider monkey surveys between October 2013 and December 2014. In a survey team of two to three people, we searched for these primates between 05.30 and 18.30 hr from boats. From 06.00 to 10.00 hr and 14.00 to 18.00 hr, we searched for the monkeys on a system of seven terrestrial trails (Fig. 1), which totaled 36 km in length. Individual trail lengths were between 4 and 9 km, and were a mixture of specially cut trails and extensions to existing ones. We covered each transect three times a month. Groups were not habituated to humans, but hunting pressure is low in the region (de Oliveira et al., 2016), and Ateles showed little fear. Nevertheless, we never approached a feeding group closer than 10 m to avoid causing the group to flee. Whenever we noted animals feeding, we recorded the feeding tree species (whenever field conditions permitted), and its GPS location. To allow the animals time to feed and for the accumulation of maximum number of ort-fruits for analysis, we did not access such feeding trees until 1-2 hours after the initial observation.
For trees that the monkeys visited, we collected samples (mean: 1.9 samples per feeding tree; SD ± 0.86, range: 1-4) of fallen, noneaten fruits (including material brought down by wind or rainstorms). We also obtained fruit samples directly from trees (mean: 1.9 samples per tree; SD ± 0.78, range: 1-3) by branch trimming. These data were obtained from a mean of 2.81 individuals per species (SD ± 1.38, range 1-7). For this, we sampled trees with fruits beneath them that had distinctive signs of very recent feeding (i.e., orts still oozing sap and not-discolored), or at which feeding had very recently been observed. We did not sample fruits that we knew from discoloration had been on the ground for more than 1-2 hours, as such fruits’ insect content was likely to have been substantially reduced by foraging ants, potentially leading to erroneous conclusions.
When orts were collected from trees without direct feeding observations for Ateles, the potential exists for such material to be confused with the feeding remnants of other large primates in the region, such as the two Alouatta species present in the region (A. nigerrima, western bank; A. discolor, eastern bank: Jucá et al., 2020). However, the tooth marks made by Alouatta and Ateles are distinct, and consistently so, allowing differentiation in the field.
We stored sampled material in plastic bags, labeled as either ort, fallen-uneaten, or on-tree. We added a small volume of 70% alcohol to each fruit sample bag to ensure any infestation remained contained until analyzed. This procedure ensured that seed-inhabiting larvae did not invade the pulp and bias the results.
While monkeys can be messy eaters, components found on the ground may have been discarded for a reason. It is, therefore, possible that the food remains (orts) on the ground may not truly reflect what the animals were eating. However, given that the methodological alternatives would involve marking every fruit and monitoring both their presence and any change in the variable being analyzed, several previous studies have used ort-based approaches to explore diet and selectivity patterns. These include Regan et al. (2001) who sampled the discarded items by Alouatta seniculus, Ateles paniscus, and Sapajus apella, as well as from the trees in which feeding occurred, Bowler and Bodmer (2011) for Cacajao calvus ucayalii, Gutiérrez-Granados and Dirzo (2010) who used orts to study Ateles geoffroyi selectivity when feeding on Manilkara zapota (Sapotaceae), and Julliot (1996) who used fallen, eaten, fruits when constructing a diet list for Alouatta seniculus and Boubli (1999) who did the same for Cacajao melanocephalus.
Testing Prediction I
When the collected fruit were infested, we identified all larvae present to Order. We identified trees as close to species level as possible with Gentry (1993), Ribeiro et al. (1999), van Roosmalen (1985), relevant volumes of Flora Neotropica (Pennington (1990) for Sapotaceae), and specialist treatments (Ramos (2020) for the genus Heisteria, Coulaceae). We used Harris and Harris (2001) and Jackson (2004) as guides to botanical terminology. We then checked our list of plant species against that of Pinto (2008) for terra firme, and Ferreira and Prance (1998) for igapó. To help in identification, we also compared common names given by local guides to those listed in Freitas da Silva et al. (1977, 2004). Where needed, we updated the nomenclature based on The Plant List (www.theplantlist.org). Additionally, we photographed leaves, fruits, flowers (when available), bark, and (when possible) entire trees, and later compared the images with those in virtual herbaria (Neotropical Herbarium Specimens https://www.fieldmuseum.org/node/4781; New York Botanic Garden http://sweetgum.nybg.org/science/vh/; Tropicos https://www.tropicos.org/home), as well as the Flore de Guyane site (https://floredeguyane.piwigo.com).
Testing Prediction II
To test for selectivity, we followed Felton et al. (2008) and Barnett et al. (2017) and calculated Electivity Indices (Ivlev, 1961) for each fruit species, such that:
Where Oi = percent of ort-fruit insect-infestation, and Ti = percent of on-tree fruit insect-infestation (in fruits at the same maturation level as those eaten by Ateles spp.). Electivity values may range from −1 to +1, with −1 indicating complete avoidance, 0 indicating no preference, and +1 indicating complete selection.
For comparative purposes, and to provide a conservative selection estimate, we used the same categories as Barnett et al. (2017): values falling between −0.3 and +0.3 were taken as showing no selection was occurring; the selection was considered active when values were larger than 0.33. Finally, values smaller than −0.33 indicate negative selection (i.e., active avoidance).
Ethical Note
All research complied with Brazilian law. Institutional Animal Care and Use Committee consent was not sought as data collection was ad hoc, non-invasive, and purely observational. We adhered to Association for the Study of Animal Behaviour guidelines for research animal treatment (ASAB, 2012), to the Code of Best Practices for Field Primatology of the American Society of Primatologists and International Primatological Society (www.asp.org/resources/docs/ Code%20of_Best_Practices%20Oct%202014.pdf). We did not trap or handle study animals, and maintained a minimum 2-m distance from individuals to minimize stress. None of the authors have a conflict of interest to declare.
Data Availability
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Results
We analyzed 2,836 fruits from 74 trees, representing 27 species in 21 genera and 14 families. The samples included 1,525 (53.8%) fruits from trees and 1,311 (46.2%) orts. We sampled 34 trees by direct observation, and sampled 40 more trees where feeding had occurred very recently (i.e., fruit not discolored, and/or covered in ants). In addition, we recorded four species (Cordia sp., Boraginaceae; Protium sp. Burseraceae; Dimorphandra sp. and Swarzia sp., both Fabaceae-Caesalpinioideae) consumed by Ateles in igapó forest, but which we could not sample for logistical reasons. All records represent data from the two spider monkey species combined (Table II).
We observed Ateles chamek and A. marginatus feeding on a combined total of 31 species. Of these, we collected samples of fruit from the trees of 27 of these species. We also collected orts for 20 of the 27 species. Four of the 27 species were eaten whole and so orts could not be collected, while three had no recorded infestation.
We found that 23 species of fruits (85%) consumed by Ateles were infested with insect larvae at the maturational stage at which Ateles ate them, supporting prediction I. In 11 species (40.7%), high levels of individual fruits (35-78%) were infested (Table III). Partially supporting prediction II, we found that for the 20 species for which we had collected orts of fruits, 12 showed positive selection of infested fruits (60%), 4 showed aversions to infested fruits (20%), while no form of selection (e.g., infested and uninfested fruits were eaten at levels similar to their availability) appeared to be occurring for the four remaining species (20%; Table III). All infestation records involved larvae of Coleoptera, Diptera, and Lepidoptera. We found no adult insects within the fruit pulp tissues.
Discussion
We found that two species of spider monkey actively selected infested fruits across a variety of species in their diets, supporting suggestions that covert ingestion of animal protein may be a widespread feature of the Ateles diet (Felton et al., 2008; Felton, Felton, Raubenheimer, et al., 2009b). In many spider monkey populations, it appears that ingestion of animal protein would be met by eating figs, whose associations with fig wasps guarantee that a ripe fruit is an infested fruit. However, at locations (such as along the Tapajós River), where figs are rare both in numbers of species and individuals, spider monkeys instead appear to get their protein supplements from other fruits, where infestation is not a given (as it is with figs). This results in the selectivity seen in the current study.
The plant genera we recorded as diet items in this study are well-known components of the Ateles diet (Russo et al., 2005), and the extent of insect infestation lay well within ranges reported for Amazonian fruits (Jesus-Barros et al., 2012). However, the response of the study species to fruit infestation was non-uniform. Although spider monkeys selected infested fruits of some species (e.g., species within the genera Licania, Spondias, Tetragastis), they appeared to strongly avoid infested fruits within the genera Inga (Fabaceae-Mimosoideae) and Xylopia (Annonaceae)—both of which have multiseeded pod-like fruit. An individual selection of uninfested seeds occurred in each case, with infested seeds (and often those adjacent to them) avoided. We do not know whether such avoidance results from the infesting insects producing inimical or unpleasant-tasting chemicals or whether the specific area was made unpalatable by chemicals deposited by the host plant (Kaplan et al., 2008). Although we only analyzed infestation of the sarcotesta in this study, infested seeds of both Inga and Parkia (members of the same legume subfamily) were avoided by Cacajao (Barnett et al., 2017).
Seeds of the plant family Annonaceae are very toxic (a fact well-known to the rural Amazonians who grind them up for use as an insecticide: de Cássia Seffrin et al., 2010). Xylopia fruits are small, single-seeded, and with a thin covering of pulp. Thus, their infestation by small dipteran larvae may have activated some defensive response in the seed, making the infested seed/pulp assemblage unpalatable to primates. This, indeed, reflects a broader methodological problem because (except here for Xylopia and for Inga, which also were eaten by Chiropotes), we did not check whether seeds of eaten fruits also were infested. Infested seeds may have influenced our findings and may explain some of the results where negative selection (avoidance) occurred, as any larvae in the seeds might have changed fruit pulp chemistry (or resulted in the plant changing such chemistry). Such an effect may also explain observations that Ateles geoffroyi avoided seeds of Dialium (Fabaceae-Caesalpinioideae) when infested with beetle larvae (Benítez-Malvido et al., 2016).
The three analysed members of the Myristacaceae (Iryanthera, Osteophloeum, and Virola) were the only species to have no infestation of the analysed part (the aril), which may be linked to the high levels of insecticidal compounds present in these tissues (Nakamura et al., 1988; Su, 1989). For four of the analysed species, the spider monkeys showed neither preference nor avoidance. These four species contained the highest (Passiflora cf. costata, 78.6%) and lowest (Pouteria macrophylla, 8%) infestation values. Thus, it is possible that, in scenarios of both very high and very low infestation, the gain in protein intake derived from selecting infested fruit is not offset by the extra energy and time required to achieve such gains (Krebs et al., 1977); because (i) noninfested fruits are rarely encountered when fruits have high levels of infestation, so the time spent rejecting them may not be recuperated by the opportunity cost that the next fruit will be infected, while (ii) in situations where infested fruits are rare, searching for them may involve more expenditure of more energy than accrues from the ingestion of insect larvae in occasionally encountered infested fruit. This makes this an example of “hunting by expectation” (Hodges, 1981), a situation also known for nectar feeding with tamarins (Saguinus mystax and Saguinus fuscicollis: Garber, 1988). Thus, while it is important to consider the nutrient-balancing framework of primate foraging decision-making (Raubenheimer et al., 2009, 2015), this and optimality are complimentary, because spatially aggregated foods must be found and then, once located, distinguished between for the highest nutrient reward with the lowest time/energy expenditure.
One of the potential limitations of the ort fruit collection method is that primates can be highly selective in their consumption of plant foods/plant parts. Thus, while large amounts of potential food may fall to the ground, at least some of this may have been discarded for a reason, which may result in an over-representation within available orts of foodstuffs in the rejected category. Given that unambiguous quantification under such circumstances would require numbering every fruit or leaf, followed by regular monitoring of their presence and the status of the variable of study interest (phytochemical composition, insect infestation, etc.), it is common to use ort-based methods to explore diet and selectivity patterns (Barnett et al., 2017: Cacajao ouakary; Bowler & Bodmer, 2011; Cacajao calvus ucayalii; Boubli, 1999: Cacajao melanocephalus; Gutiérrez-Granados & Dirzo, 2010: Ateles geoffroyi; Julliot, 1996: Alouatta seniculus; Regan et al., 2001: Alouatta seniculus, Ateles paniscus and Sapajus apella; Take, 2017: Pithecia chrysocephala, Saimiri sciureus and Saguinus bicolor). Until a nontime- and nonlabor-intensive method for monitoring whole canopies and their fruits or leaves becomes available, studies are likely to continue with this potential, and currently unquantifiable, bias as an inherent constituent.
Consumption of larval insects is an especially efficient means of accessing protein, which can be 60-80% by wet weight (Bukkens, 1997), as well as a variety of free amino acids (Drew, 1988), lipids, and minerals (Finke, 2013). In addition, accessing such insects is energetically efficient since larvae, generally being more lightly sclerotized, often are easier to digest than fully developed individuals (imagoes) (Raubenheimer & Rothman, 2013). Furthermore, the use of such larvae can be energetically effective as they can form a spatially aggregated resource, unlike free-living imagoes that often are dispersed or may have defensive behaviors.
There are, however, potential downsides to ingesting infested fruits. For example, if fruit infestation is extensive, or large volumes of the fruit have already been ingested by larvae, the energetic and nutritive values of infested fruits may be lower than comparably sized noninfested ones. This point has been used to explain why infested fruits are sometimes avoided by rodents (Muñoz & Bonal, 2008; Steele et al., 1996), and bats (Utzurrum & Heideman, 1991). In addition, damaged plant tissues may become toxic due to fungal infection. Such toxicity is hazardous, as some spoilage fungi synthesize toxic molecules, such as aflatoxins and patulin, which are some of the most potent carcinogens known (Janzen, 1977). Thus, spoilt fruits (discolored or blotched) are widely avoided by frugivores (Borowicz, 1988; Buchholz & Levey, 1990). This, and changes in palatability due to accumulation of excretory products and alteration of chemosynthetic pathways (Tollrian & Harvell, 1999), may explain other examples of frugivores avoiding infested fruits (primates: Bravo, 2012; Julliot, 1996; bats: Engriser, 1995; birds: Palacio et al., 2020; Traveset et al., 1995). However, some cases of avoidance appear to be due to the direct influence of the infesting insect. For example, larvae of the holly berry midge (Asphondylia ilicicola, Cecidomyiidae) chemically prevent the berry of the American holly (Ilex opaca, Aquioliaceae) from turning red, which results in avoidance of infested fruits by avian frugivores that use red as a cue for edibility (Krischik et al., 1989). All of these potential downsides and complex mutualisms indirectly affect plant-consumer interactions (Krischik et al., 1989), which may be responsible for the non-uniform responses of ripe-fruit specialists to fruit infestation.
There also is the peculiar case of figs (genus Ficus); their pollination ecology involves fig-wasps (Agaonidae), with larvae of both sexes developing in the maturing synconium. Having fertilized females, the wingless males die without leaving the fruit (Janzen, 1979; Weiblen, 2002). Thus, consumption of wild fig fruits inevitably involves covert carnivory (sensu, Barnett et al., 2017) of, at least, dead male and developing larval wasps. Even though the fig-agaonid wasp relation is biologically well known, the nutritional significance of the concealed larval component of the fruit is rarely (Link, 2003) commented on or acknowledged, even when figs make up truly significant parts of the diet (e.g., hornbills, primates: Choudhury, 1989; Kinnaird & O’Brien, 2005; Kinnaird et al., 1996; Wrangham et al., 1993). It is therefore pertinent to note that figs are a major component of Ateles diets in certain parts of their geographic range (e.g., 50% for A. chamek in Bolivian semihumid forests: Felton et al., 2008).
With the exception of fig wasps, their braconid parasites, and some gall-forming wasp species (Dsouza & Ravishankar, 2014; Weiblen, 2002), it seems unlikely that covert carnivory is a contributory component in figs, because very few species of insect appear to infest figs on the tree. Fallen figs are rapidly colonized by yeasts, beetles, and flies (Palmieri & Pereira, 2018), but Ateles spp. rarely forage on the ground (Campbell et al., 2005), so that this infestation is unlikely to be relevant to spider monkeys. Additionally, although they represent one of the major groups of fruit-infesting insects in the tropics (Ajuja & Liedo, 1993), Anastrepha fruit flies (Tephritidae) are not known to infest wild figs (Hernández-Ortiz & Pérez-Alonso, 1993; Jesus-Barros et al., 2012). This is likely due to the presence of digestive enzymes (ficins) with strong larvicidal properties (Kitajima et al., 2018) in the latex of many Moraceae (Zare et al., 2013).
Nutritional modelling of protein requirements of a Bolivian population of A. chamek (Felton, Felton, Wood, et al., 2009c) matched observed protein intake and nutritional profiles closely, so that Felton and coworkers concluded that Ficus was a nutritionally balanced food for Ateles. However, while eating figs is common in Ateles species, in no other population studied do ingestion levels reach those reported from Bolivia (Felton et al., 2008, 2009a,b,c), where almost 50% of total time spent feeding involved ingesting figs. This may reflect underlying differences in study site floral composition, where four Ficus species occurred, two of which (Ficus boliviana, Ficus trigona) were common and produced large fruit crops for 8–9 months/year, so providing fruits when other trees did not (Felton et al., 2008; Felton, Felton, Wood, et al., 2009c). In contrast, in the Tapajós National Forest (a protected area close to the current study sites), two independent studies (de Lima Francisco & Cordeira, 2004; Pinto, 2008) reported only one (uncommon) Ficus species. The high level of Ficus ingestion reported by Felton et al. (2008) appears unusual, as a comparison of sites in Surinam, Ecuador, Colombia, and Panama found Ficus comprised no more than 15.4% of the diet at the individual sites and, at two sites, less than 3% of the diet (Russo et al., 2005).
That very few figs were consumed at our study sites may simply be because figs are not common enough in Tapajós basin forests for their fruits to dominate the diet of large-bodied primates travelling in large groups (30-50 individuals: Symington, 1990). Nevertheless, given the importance of phylogeny in determining physiology (Garland Jr. et al., 2005; Herrera, 1992a), and the metabolic underpinning of overall ecology in a genus (Herrera, 1992b; McNab, 2002), it is very likely that the protein-balancing metabolic model advanced by Felton et al. (2008) is practiced by A. chamek and A. marginatus on the Tapajós, but that, in the absence of large volumes of nutritionally-balanced figs, these primates adopt a strategy of selecting infested fruits as an energetically efficient alternative. This hypothesis could be tested on other spider monkey populations in forests of varying Ficus abundance.
Thus, while we have shown that two members of the genus Ateles practice covert carnivory, it is possible that other species, generally considered highly frugivorous, also may adopt this strategy. Accordingly, we suggest that researchers check the pulp (or seeds) being eaten for such evidence. Other sources of insects that merit investigation, include the leaf bases of epiphytic bromeliads, buds of leaves and flowers (see Barnett et al., 2020 for infestation levels of Eschweilera flower buds), and young leaves. Spider monkeys use bromeliads as a water source (Di Fiore et al., 2008), for their succulent leaf bases (Campbell, 2008; Dew, 2005), or for the small vertebrate prey they conceal (Valero, 2004). However, they also contain larvae of a specialist genus of bromeliad-eating weevils (Metamasius, Dryophthoridae: Cave et al., 2006), which could provide an additional reason for their exploitation. Buds also should be considered due to the ubiquity of bud-boring insect larvae (Sugiura & Yamazaki, 2009) and, while young leaves are widely considered a good source of protein (Felton et al., 2008; Felton, Felton, Wood, et al., 2009c), they are commonly attacked by leaf-mining insects (Sinclair & Hughes, 2010), which could contribute supplemental levels of protein, as could the insects within phytophilous galls (Askew, 1980; Raman, 2012).
Further complexities arise because a species may select infested fruit from one plant species but not another (e.g., common bush tanagers, Chlorospingus ophthalmicus: Valburg, 1992; black-and-gold howler monkeys, Alouatta caraya: Bravo, 2012; Bravo & Zunino, 1998; golden-backed uacaris, Cacajao ouakary: Barnett et al., 2017). This likely results from the heterogeneity of plant responses, including those where plants selectively deposit toxic compounds into infested fruits (Ibanez et al., 2009), as well as insect countermeasures where larvae sequester such compounds for their own defense (Ferro et al., 2006). Just as such higher-order interactions (sensu, Mayfield & Stouffer, 2017) finesse our view of both potential seed predators (Barnett et al., 2012, 2017; Herrera, 1989), it also may do so with seed dispersers, because ingestion of seed- or pulp-eating insects generally destroys them (Bravo, 2008; Herrera, 1989; Jordano, 1987), although there are some exceptions (Di Iorio, 2015). In situations where such seed- or pulp-eating insects are destroyed during ingestion, there is reduction in the overall population of organisms that can depress plant reproductive success (da Silva & Pinheiro, 2009; Wilson, 2008); however, in some cases selective predation on insect-infested fruits leads to the destruction of the fruits (Scott & Black, 1981).
As a caveat, because spider monkeys often ingest fruit whole (Russo et al., 2005), discarded parts of a fruit might be those that were overripe or immature, potentially leading to erroneous estimates of selectivity. This possibility should be tested, either in the wild (where it would be observationally challenging—although perhaps resolved by camera traps in favored fruiting trees, with premarked fruit) or in captivity with choice tests of fruits of known infestation levels.
Plant species generally have multiple seed dispersers (Willson & Traveset, 2000; Zamora, 2000), and these may not react in the same way to infesting insects: some may prefer infested fruits or seeds, while others avoid them (Barnett et al., 2017; Valburg, 1992). Passage through Ateles guts rarely kills seeds and, indeed, often enhances germination rates (see Fuzessy et al., 2016 for topic review). Thus, by eating insect-infested fruit that other species might refuse, spider monkeys ensure that individual fruits that might otherwise not be dispersed achieve a zoochoric interaction, enhancing the plants reproductive potential. In addition, our study shows that, for some populations at least, Ateles engage in active insectivory, via the selection of infested fruits. Given the elegant interplay Felton et al. (2009a, b, c) demonstrated between energy intake and diet item composition, and the central role of protein as a key element in macronutrient balancing, it is possible that distinct frugivore populations do things differently, with the abundance of protein-rich resources in the region inhabited by different study populations, as the main variable. This may parallel the differences observed in time spent foraging for nonembedded arthropod prey displayed by different species and populations of Lagothrix (Gonzalez et al., 2016), the ateline genus closely related to Ateles (Ange-van Heugten, 2008), because our findings reinforce the observation of Felton et al. (2008, p. 393) that “both intra- and interspecific distinctions in ateline feeding ecology are best explained by differences in the local food resources” (see also Russo et al., 2005). Thus, studies comparing the incidence of covert carnivory in areas where figs either do or do not constitute significant parts of the diet of resident Ateles would both clarify the current situation and help open future avenues of research, as well as providing larger samples for genera such as Xylopia, where numbers of sampled individuals were low in the current study. Because a particular stage of fruit ripeness generally corresponds to a particular insect maturation stage, future studies could investigate whether infesting insect species, and if their different life cycles influence protein/lipid/micronutrient availability for individual fruit species. Additional studies could investigate how variation in fruit morphology and size affects insect infestation.
References
Ajuja, M., & Liedo, P. (1993). Fruit flies, biology and management. Springer.
Ange-van Heugten, K. D. (2008). Nutritional analyses and intervention in the captive woolly monkey (Lagothrix lagotricha) Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands.
Arroyo-Rodríguez, V., Aguilar-Barajas, E., González-Zamora, A., Rocha-Ramirez, V., González-Rodríguez, A., et al (2017). Parent-parent and parent-offspring distances in Spondias radlkoferi seeds suggest long-distance pollen and seed dispersal: evidence from latrines of the spider monkey. Journal of Tropical Ecology, 33, 95–106. https://doi.org/10.1017/S0266467417000050.
ASAB. (2012). Guidelines for the treatment of animals in behavioural research and teaching. Animal Behaviour, 83, 301–309.
Askew, R. R. (1980). The diversity of insect communities in leafmines and plant galls. Journal of Animal Ecology, 49, 817–829. https://doi.org/10.2307/4229.
Ballantyne, J. (2018). Insect-infested fruit preference and detection by the golden-backed uacari monkey (Cacajao ouakary). Masters Dissertation, Roehampton University, London, UK.
Barnett, A. A., Boyle, S. A., Pinto, L. P., Lourenço, W. C., Almeida, T., et al (2012). Primary seed dispersal by three Neotropical seed-predators (Cacajao melanocephalus, Chiropotes and Chiropotes albinasus). Journal of Tropical Ecology, 28, 543–555. https://doi.org/10.1017/S0266467412000600.
Barnett, A. A., Ronchi-Teles, B., Silva, W. S., Andrade, R., Almeida, T., et al (2017). Covert carnivory: a seed-predating primate, the golden-backed uacari, selects insect-infested fruits. Journal of Zoological Research, 1, 16–31 https://www.sryahwapublications.com/journal-of-zoological-research/volume-1-issue-1/3.php.
Barnett, A. A., Boyle, S. A., dos Santos-Barnett, T. C., Tuma, T., Piedade, M. T. F., et al (2020). Buds, bugs and bienniality: the floral biology of Eschweilera tenuifolia (O. Berg) Miers (Lecythidaceae) in blackwater flooded forest (igapó), central Amazonian Brazil. Forests, 11, 1251. https://doi.org/10.3390/f11121251.
Barnett, A. A., dos Santos-Barnett, T. C., Muir, J., Tománek, P., Gregory, T., Matte, A. L. L., Bezerra, B. M., de Oliveira, T. G., Norconk, M., & Boyle, S. A. (n.d.). Beans with bugs: covert insectivory and infested seed selection by the red-nosed cuxiu monkey, Chiropotes albinasus. At Biotropica (BITR-22-018-R1).
Bello, C., Galetti, M. M., Pizo, A. L. F., Magnago, L. F., Rocha, M. F., et al (2015). Defaunation affects carbon storage in tropical forests. Science Advances, 1, e1501105. https://doi.org/10.1126/sciadv.1501105.
Benítez-Malvido, J., Zermeño-Hernández, I. A., González-DiPierro, A. M., Lombera, R., & Estrada, E. (2016). Frugivore choice and escape from pre-dispersal seed predators: the case of Dialium guianense and two sympatric primate species in southern Mexico. Plant Ecology, 217, 923–933. https://doi.org/10.1007/s11258-016-0617-6.
Berg, C. C., Avila, M. V., & Kooy, F. (1984). Ficus species of Brazilian Amazonia and the Guianas. Acta Amazonica, 14, 159–194. https://doi.org/10.1590/1809-43921984145194.
Borowicz, V. A. (1988). Do vertebrates reject decaying fruit? An experimental test with Cornus amomum fruits. Oikos, 53, 74–78. https://doi.org/10.2307/3565665.
Boubli, J. P. (1999). Feeding ecology of black-headed uacaris (Cacajao melanocephalus melanocephalus) in Pico da Neblina National Park, Brazil. International Journal of Primatology, 20, 719–749. https://doi.org/10.1023/A:1020704819367.
Bowler, M., & Bodmer, R. E. (2011). Diet and food choice in Peruvian red uakaris (Cacajao calvus ucayalii): selective or opportunistic seed predation? International Journal of Primatology, 32, 1109–1122. https://doi.org/10.1007/s10764-011-9527-6.
Bravo, S. P. (2008). Seed dispersal and ingestion of insect-infested seeds by black howler monkeys in flooded forests of the Parana River, Argentina. Biotropica, 40, 471–476. https://doi.org/10.1111/j.1744-7429.2008.00400.x.
Bravo, S. P. (2012). From which Ocotea diospyrifolia trees does Alouatta caraya (Primates, Atelidae) eat fruits? Journal of Tropical Ecology, 28, 417–420. https://doi.org/10.1017/S0266467412000272.
Bravo, S. P., & Zunino, G. E. (1998). Effects of black howler monkey (Alouatta caraya) seed ingestion on insect larvae. American Journal of Primatology, 45, 411–415. https://doi.org/10.1002/(SICI)1098-2345(1998)45:4%3C411::AID-AJP7%3E3.0.CO;2-Z.
Buchholz, R., & Levey, D. J. (1990). The evolutionary triad of microbes, fruits, and seed dispersers: an experiment in fruit choice by cedar waxwings, Bombycilla cedrorum. Oikos, 59, 200–204. https://doi.org/10.2307/3545535.
Bukkens, S. G. F. (1997). The nutritional value of edible insects. Ecology of Food and Nutrition, 36, 287–319. https://doi.org/10.1080/03670244.1997.9991521.
Campbell, C. J. (2008). Spider monkeys: behavior, ecology and evolution of the genus Ateles. Cambridge University Press.
Campbell, C. J., Aureli, F., Chapman, C. A., Ramos-Fernández, G., Matthews, K., et al (2005). Terrestrial behavior of Ateles spp. International Journal of Primatology, 26, 1039–1051. https://doi.org/10.1007/s10764-005-6457-1.
Cant, J. G. H. (1990). Feeding ecology of spider monkeys (Ateles geoffroyi) at Tikal, Guatemala. Human Evolution, 5, 269–281. https://doi.org/10.1007/BF02437243.
Carlson, B. A., Rothman, J. M., & Mitani, J. C. (2013). Diurnal variation in nutrients and chimpanzee foraging behavior. American Journal of Primatology, 75, 342–349. https://doi.org/10.1002/ajp.22112.
Cave, R. D., Duetting, P. S., Creel, O. R., & Branch, C. L. (2006). Biology of Metamasius mosieri (Coleoptera: Dryophthoridae), with a description of the larval and pupal stages. Annals of the Entomological Society of America, 99, 1146–1153. https://doi.org/10.1603/0013-8746(2006)99[1146:BOMMCD]2.0.CO;2.
Choudhury, A. (1989). Ecology of the capped langur (Presbytis pileatus) in Assam, India. Folia Primatologica, 52, 88–92. https://doi.org/10.1159/000156385.
Cifuentes, E. F., Ramírez, M. A., León, J., Galvis, N., Vargas, S., & Stevenson, P. R. (2013). Dieta de los churucos colombianos (Lagothrix lagothricha lugens) en relación con la productividad de frutos en el Parque Nacional Natural Cueva de Los Guácharos. In T. R. Defler, P. R. Stevenson, M. L. Bueno, & D. C. Guzmán-Caro (Eds.), Primates Colombianos en Peligro de Extinción (pp. 294–312).
da Silva, A. L. G., & Pinheiro, M. C. B. (2009). Reproductive success of four species of Eugenia L. (Myrtaceae). Acta Botanica Brasilica, 23, 526–534. https://doi.org/10.1590/S0102-33062009000200024.
Dasilva, G. L. (1994). Diet of Colobus polykomos on Tiwai Island: selection of food in relation to its seasonal abundance and nutritional quality. International Journal of Primatology, 15, 655–680. https://doi.org/10.1007/BF02737426.
de Cássia Seffrin, R., Shikano, I., Akhtar, Y., & Isman, M. B. (2010). Effects of crude seed extracts of Annona atemoya and Annona squamosa L. against the cabbage looper, Trichoplusia ni in the laboratory and greenhouse. Crop Protection, 29, 20–24. https://doi.org/10.1016/j.cropro.2009.09.003.
de Lima Francisco, A., & Cordeira, A. (2004). Floresta Nacional do Tapajós: plano de manejo. Volume I – Informações gerais. IBAMA-MMA.
de Oliveira, T. G., Mazim, D., Quixaba Vieira, O., Barnett, A. A., do Nascimento Silva, G., et al (2016). Nonvolant mammal megadiversity and conservation issues in a threatened Central Amazonian hotspot in Brazil. Tropical Conservation Science, 9, 1–16. https://doi.org/10.1177/1940082916672340.
Devescovi, F., Liendo, M. C., Bachmann, G. E., et al (2015). Fruit infestation patterns by Anastrepha fraterculus and Ceratitis capitata reveal that cross-recognition does not lead to complete avoidance of interspecific competition in nature. Agricultural and Forest Entomology, 17, 325–335. https://doi.org/10.1111/afe.12111.
Dew, J. L. (2005). Foraging, food choice, and food processing by sympatric ripe-fruit specialists: Lagothrix lagotricha poeppigii and Ateles belzebuth. International Journal of Primatology, 26, 1107–1135. https://doi.org/10.1007/s10764-005-6461-5.
Di Fiore, A., & Rodman, P. S. (2001). Time allocation patterns of lowland woolly monkeys (Lagothrix lagotricha poeppigii) in a neotropical terra firma forest. International Journal of Primatology, 22, 449–480. https://doi.org/10.1023/A:1010759729567.
Di Fiore, A., Link, A., & Dew, J. L. (2008). Diets of wild spider monkeys. In C. J. Campbell (Ed.), Spider monkeys: behavior, ecology and evolution of the genus Ateles (pp. 81–137). Cambridge University Press.
Di Iorio, O. R. (2015). Survival and emergence of adult seed beetles (Coleoptera: Chrysomelidae: Bruchinae) from legume seeds egested by vertebrates. Boletín de la Sociedad Entomológica Aragonesa, 57, 286–292.
Dias da Silva, R. H. P., Castro Sá, M. J., Baccaro, F. B., Tománek, P., & Barnett, A. A. (2020). Juggling options: Manipulation ease determines primate optimal fruit-size choice. Biotropica, 52, 1275–1285. https://doi.org/10.1111/btp.12835.
Drew, R. A. I. (1988). Amino acid increases in fruits infested by fruit flies of the family Tephritidae. Zoological Journal of the Linnean Society, 93, 107–112. https://doi.org/10.1111/j.1096-3642.1988.tb01529.x.
Dsouza, M. R., & Ravishankar, B. E. (2014). Nutritional sink formation in galls of Ficus glomerata Roxb. (Moraceae) by the insect Pauropsylla depressa (Psyllidae, Hemiptera). Tropical Ecology, 55, 129–136.
Dunn, J. C., Cristóbal-Azkarate, J., & Veà, J. J. (2009). Differences in diet and activity pattern between two groups of Alouatta palliata associated with the availability of big trees and fruit of top food taxa. American Journal of Primatology, 71, 654–662. https://doi.org/10.1002/ajp.20700.
Engriser, E. M. (1995). The effect of insect larvae infestation on fruit choice in phyllostomid fruit bats: an experimental study. Biotropica, 27, 523–525. https://doi.org/10.2307/2388967.
Felton, A. M., Felton, A., Wood, J. T., & Lindenmayer, D. B. (2008). Diet and feeding ecology of Ateles chamek in a Bolivian semihumid forest: the importance of Ficus as a staple food resource. International Journal of Primatology, 29, 379–403. https://doi.org/10.1007/s10764-008-9241-1.
Felton, A. M., Felton, A., Lindenmayer, D. B., & Foley, W. J. (2009a). Nutritional goals of wild primates. Functional Ecology, 23, 70–78. https://doi.org/10.1111/j.1365-2435.2008.01526.x.
Felton, A. M., Felton, A., Raubenheimer, D., Simpson, S. J., Foley, W. J., et al (2009b). Protein content of diets dictates the daily energy intake of a free-ranging primate. Behavioral Ecology, 20, 685–690. https://doi.org/10.1093/beheco/arp021.
Felton, A. M., Felton, A., Wood, J. T., Foley, W. J., Raubenheimer, D., et al (2009c). Nutritional ecology of Ateles chamek in lowland Bolivia: how macronutrient balancing influences food choices. International Journal of Primatology, 30, 675–696. https://doi.org/10.1007/s10764-009-9367-9.
Ferreira, L. V., & Prance, G. T. (1998). Structure and species richness of low-diversity floodplain forest on the Rio Tapajós, eastern Amazonia, Brazil. Biodiversity & Conservation, 7, 585–596. https://doi.org/10.1023/A:1008848200441.
Ferro, V. G., Guimarães Jr., P. R., & Trigo, J. R. (2006). Why do larvae of Utetheisa ornatrix penetrate and feed in pods of Crotalaria species? Larval performance vs. chemical and physical constraints. Entomologia Experimentalis et Applicata, 121, 23–29. https://doi.org/10.1111/j.1570-8703.2006.00450.x.
Finke, M. D. (2013). Complete nutrient content of four species of feeder insects. Zoo Biology, 32, 27–36. https://doi.org/10.1002/zoo.21012.
Fleming, T. H., & Kress, W. J. (2011). A brief history of fruits and frugivores. Acta Oecologica, 37, 521–530. https://doi.org/10.1016/j.actao.2011.01.016.
Freitas da Silva, M., Lisbôa, P. L. B., & Lisbôa, R. C. L. (1977). Nomes vulgares de plantas amazônicas. Conselho Nacional de Desenvolvimiento Científico and Instituto Nacional de Pesquisas da Amazônia, Manaus, AM.
Freitas da Silva, M., Gomes de Souza, L. A., & de Madeiros Carreira, L. M. (2004). Nomes populares das Leguminosas do Brasil. Instituto Nacional de Pesquisas da Amazônia, Manaus, AM.
Fricke, E. C., Simon, M. J., Reagan, K. M., Levey, D. J., Riffell, J. A., et al (2013). When condition trumps location: seed consumption by fruit-eating birds removes pathogens and predator attractants. Ecology Letters, 16, 1031–1036. https://doi.org/10.1111/ele.12134.
Fuzessy, L. F., Cornelissen, T. G., Janson, C., & Silveira, F. A. (2016). How do primates affect seed germination? A meta-analysis of gut passage effects on neotropical plants. Oikos, 125, 1069–1080.
Garber, P. A. (1988). Foraging decisions during nectar feeding by tamarin monkeys (Saguinus mystax and Saguinus fuscicollis, Callitrichidae, Primates) in Amazonian Peru. Biotropica, 20, 100–106. https://doi.org/10.2307/2388181.
Garber, P. A., Righini, N., & Kowalewski, M. M. (2015). Evidence of alternative dietary syndromes and nutritional goals in the genus Alouatta. In M. M. Kowalewski, P. A. Garber, L. Cortés-Ortiz, B. Urbani, & D. Youlatos (Eds.), Howler monkeys (pp. 85–109). Springer.
Garland Jr., T., Bennett, A. F., & Rezende, E. L. (2005). Phylogenetic approaches in comparative physiology. Journal of Experimental Biology, 208, 3015–3035. https://doi.org/10.1242/jeb.01745.
Gautier-Hion, A., Sourd, C., & Quris, R. (1984). Fruit choice by frugivorous forest monkeys—Relations with fruit morphology and nutrient content. International Journal of Primatology, 5, 340. https://doi.org/10.1007/BF02738971.
Gentry, A. H. (1993). A field guide to the families and genera of woody plants of Northwest South America (Colombia, Ecuador, Peru) with supplementary notes. University of Chicago Press.
Gonzalez, M., Clavijo, L., Betancur, J., & Stevenson, P. R. (2016). Fruits eaten by woolly monkeys (Lagothrix lagothricha) at local and regional scales. Primates, 57, 241–251. https://doi.org/10.1007/s10329-016-0524-4.
González-Zamora, A., Arroyo-Rodríguez, V., Chaves, Ó. M., Sánchez-López, S., Stoner, K. E., et al (2009). Diet of spider monkeys (Ateles geoffroyi) in Mesoamerica: current knowledge and future directions. American Journal of Primatology, 71, 8–20. https://doi.org/10.1002/ajp.20625.
Gutiérrez-Granados, G., & Dirzo, R. (2010). Indirect effects of timber extraction on plant recruitment and diversity via reductions in abundance of frugivorous spider monkeys. Journal of Tropical Ecology, 26, 45–52.
Harris, J. G., & Harris, M. W. (2001). Plant identification terminology: an illustrated glossary (2nd ed.). Spring Lake Publishing.
Hemingway, C. A. (1998). Selectivity and variability in the diet of Milne-Edwards’ sifakas (Propithecus diadema edwardsi): Implications for folivory and seed-eating. International Journal of Primatology, 19, 355–737. https://doi.org/10.1023/A:1020344018670.
Hernández-Ortiz, V., & Pérez-Alonso, R. (1993). The natural host plants of Anastrepha (Diptera: Tephritidae) in a tropical rain forest of Mexico. Florida Entomologist, 76, 447–460. https://doi.org/10.2307/3495645.
Herrera, C. M. (1989). Vertebrate frugivores and their interaction with invertebrate fruit predators: supporting evidence-from a Costa Rican dry forest. Oikos, 54, 185–188. https://doi.org/10.2307/3565264.
Herrera, C. M. (1992a). Historical effects and sorting processes as explanations for contemporary ecological patterns: character syndromes in Mediterranean woody plants. American Naturalist, 140, 421–446. https://doi.org/10.1086/285420.
Herrera, C. M. (1992b). Interspecific variation in fruit shape: allometry, phylogeny, and adaptation to dispersal agents. Ecology, 73, 1832–1841. https://doi.org/10.2307/1940034.
Hill, J. E., & Breidenbach, R. W. (1974). Proteins of soybean seeds: II. Accumulation of the major protein components during seed development and maturation. Plant Physiology, 53, 747–751. https://doi.org/10.1104/pp.53.5.747.
Hladik, C. M., Hladik, A., Bousset, J., Valdebouze, P., Viroben, G., et al (1971). Le régime alimentaire des primates de l’Île de Barro Colorado (Panama). Folia Primatologia, 16, 85–122. https://doi.org/10.1159/000155393.
Hodges, C. M. (1981). Optimal foraging in bumblebees: hunting by expectation. Animal Behaviour, 29, 1166–1171. https://doi.org/10.1016/S0003-3472(81)80068-1.
Houle, A., Chapman, C. A., & Vickery, W. L. (2007). Intra-tree variation in fruit production and implications for primate foraging. International Journal of Primatology, 28, 1197–1217. https://doi.org/10.1007/s10764-007-9214-9.
Howe, H. F. (1980). Monkey dispersal and waste of a neotropical fruit. Ecology, 61, 944–959. https://doi.org/10.2307/1936763.
Howe, H. F. (1986). Seed dispersal by fruit-eating birds and mammals. In D. R. Murray (Ed.), Seed dispersal (pp. 123–189). Academic Press.
Ibanez, S., Gallet, C., Dommanget, F., & Després, L. (2009). Plant chemical defence: a partner control mechanism stabilising plant-seed-eating pollinator mutualisms. BMC Evolutionary Biology, 9, 261. https://doi.org/10.1186/1471-2148-9-261.
Instituto Brasileiro de Geografia e Estatística (1979). Estudo Nacional de Despesa Familiar: Tabela de composição dos alimentos. Instituto Brasileiro de Geografia e Estatística.
Ivlev, V. S. (1961). Experimental ecology of the feeding of fishes. Yale University Press.
Jackson, B. D. (2004). Dictionary of botanical names and terms. Laurier Books New York.
Janzen, D. H. (1977). Why fruits rot, seeds mold and meat spoils. American Naturalist, 111, 691–711. https://doi.org/10.1086/283200.
Janzen, D. H. (1979). How to be a fig. Annual Review Ecology and Systematics, 10, 13–51. https://doi.org/10.1146/annurev.es.10.110179.000305.
Jesus-Barros, C. R., Adaime, R., Oliveira, M. N., Silva, W. R., Cost-Neto, S. V., et al. (2012). Anastrepha (Diptera: Tephritidae) species, their hosts and parasitoids (Hymenoptera: Braconidae) in five municipalities of the state of Amapá, Brazil. Florida Entomologist, 95, 694–705. https://doi.org/10.1653/024.095.0320.
Jordano, P. (1987). Avian fruit removal: effects of fruit variation, crop size, and insect damage. Ecology, 68, 1711–1723. https://doi.org/10.2307/1939863.
Jordano, P. (1995). Angiosperm fleshy fruits and seed dispersers: a comparative analysis of adaptation and constraints in plant-animal interactions. American Naturalist, 145, 163–191. https://doi.org/10.1086/285735.
Jucá, T., Boyle, S., Cavalcanti, G., Cavalcante, T., Tomanek, P., Clemente, S., de Oliveira, T., & Barnett, A. A. (2020). Being hunted high and low: do differences in nocturnal sleeping and diurnal resting sites of howler monkeys (Alouatta nigerrima and Alouatta discolor) reflect safety from attack by different types of predator? Biological Journal of the Linnean Society, 131, 203–219. https://doi.org/10.1093/biolinnean/blaa102.
Julliot, C. (1996). Fruit choice by red howler monkeys (Alouatta seniculus) in a tropical rain forest. American Journal of Primatology, 40, 261–282. https://doi.org/10.1002/(SICI)1098-2345(1996)40:3%3C261::AID-AJP4%3E3.0.CO;2-W.
Julliot, C., & Sabatier, D. (1993). Diet of the red howler monkey (Alouatta seniculus) in French Guiana. International Journal of Primatology, 14, 527–550. https://doi.org/10.1007/BF02215446.
Kaplan, I., Halitschke, R., Kessler, A., Sardanelli, S., & Denno, R. F. (2008). Constitutive and induced defenses to herbivory in above-and belowground plant tissues. Ecology, 89, 392–406. https://doi.org/10.1890/07-0471.1.
Kato, M. J. (1995). The chemistry of Amazonian Myristicaceae: developmental, ecological, and pharmacological aspects. In P. R. Seidl, O. R. Gottlieb, & M. A. C. Kaplan MAC (Eds.), Chemistry of the Amazon: biodiversity, natural products, and environmental issues (pp.168–179). American Chemical Society. https://doi.org/10.1021/bk-1995-0588.ch014
Kinnaird, M. F., & O’Brien, T. G. (2005). Fast foods of the forest: the influence of figs on primates and hornbills across Wallace's line. In J. L. Drew & J. P. Boubli (Eds.), Tropical fruits and frugivores (pp. 155–184). Springer. https://doi.org/10.1007/1-4020-3833-X_9.
Kinnaird, M. F., O'Brien, T. G., & Suryadi, S. (1996). Population fluctuation in Sulawesi red-knobbed hornbills: tracking figs in space and time. The Auk, 113, 431–440. https://doi.org/10.2307/4088909.
Kitajima, S., Aoki, W., Shibata, D., Nakajima, D., Sakurai, N., et al (2018). Comparative multi-omics analysis reveals diverse latex-based defense strategies against pests among latex-producing organs of the fig tree (Ficus carica). Planta, 247, 1423–1438. https://doi.org/10.1007/s00425-018-2880-3.
Krebs, J. R., Erichsen, J. T., Webber, M. I., & Charnov, E. L. (1977). Optimal prey selection in the great tit (Parus major). Animal Behaviour, 25, 30–38. https://doi.org/10.1016/0003-3472(77)90064-1.
Krischik, V., McCloud, E. S., & Davidson, J. A. (1989). Selective avoidance by vertebrate frugivores of green holly berries infested with acecidomyiid fly (Diptera: Cecidomyiidae). American Midland Naturalist, 121, 350–354. https://doi.org/10.2307/2426039.
Laurent, P., Braekman, J. C., & Daloze, D. (2005). Insect chemical defense. In S. Schultz (Ed.), The chemistry of pheromones and other semiochemicals (Vol. II, pp. 167–229). Springer.
Lindshield, S. M., & Rodrigues, M. A. (2009). Tool use in wild spider monkeys (Ateles geoffroyi). Primates, 50, 269–272. https://doi.org/10.1007/s10329-009-0144-3.
Link, A. (2003). Insect-eating by spider monkeys. Neotropical. Primates, 11, 104–107.
Link, A., & Di Fiore, A. (2006). Seed dispersal by spider monkeys and its importance in the maintenance of Neotropical rain-forest diversity. Journal of Tropical Ecology, 22, 235–246. https://doi.org/10.1017/S0266467405003081.
Martins, M. M. (2008). Fruit diet of Alouatta guariba and Brachyteles arachnoides in Southeastern Brazil: comparison of fruit type, color, and seed size. Primates, 49, 1–8. https://doi.org/10.1007/s10329-007-0050-5.
Maruyama, P. K., Melo, C., Pascoal, C., Vicente, E., Cardoso, J. C. F., et al (2019). What is on the menu for frugivorous birds in the Cerrado? Fruiting phenology and nutritional traits highlight the importance of habitat complementarity. Acta Botanica Brasilica, 33, 572–583. https://doi.org/10.1590/0102-33062019abb0221.
Masette, M., Isabirye-Basuta, G., Baranga, D., Chapman, C. A., & Rothman, J. M. (2015). The challenge of interpreting primate diets: mangabey foraging on Blighia unijugata fruit in relation to changing nutrient content. African Journal of Ecology, 53, 259–267. https://doi.org/10.1111/aje.12174.
Mayfield, M. M., & Stouffer, D. B. (2017). Higher-order interactions capture unexplained complexity in diverse communities. Nature Ecology & Evolution, 1, 0062. https://doi.org/10.1038/s41559-016-0062.
McConkey, K. R., Aldy, A., Ario, A., & Chivers, D. J. (2002). Selection of fruit by gibbons (Hylobates muelleri x agilis) in the rain forests of central Borneo. International Journal of Primatology, 23, 123–145. https://doi.org/10.1023/A:1013253909046.
McGrew, W. C. (2014). The ‘other faunivory’ revisited: insectivory in human and non-human primates and the evolution of human diet. Journal of Human Evolution, 71, 4–11.
McNab, B. K. (2002). The physiological ecology of vertebrates: a view from energetics. Comstock Publishing Associates.
Ministério da Saúde, Brasil (2015). Alimentos regionais brasileiros (2nd ed.). Ministério da Saúde.
Moermond, T. C., & Denslow, J. S. (1983). Fruit choice in neotropical birds: effects of fruit type and accessibility on selectivity. Journal of Animal Ecology, 52, 407–420. https://doi.org/10.2307/4562.
Morales-Jimenez, A. L., Disotell, T., & Di Fiore, A. (2015). Revisiting the phylogenetic relationships, biogeography, and taxonomy of spider monkeys (genus Ateles) in light of new molecular data. Molecular Phylogenetics & Evolution, 82, 467–483. https://doi.org/10.1016/j.ympev.2014.09.019.
Muñoz, A., & Bonal, R. (2008). Seed choice by rodents: learning or inheritance? Behavioral Ecology and Sociobiology, 62, 913–922. https://doi.org/10.1007/s00265-007-0515-y.
Nakamura, N., Kiuchi, F., Tsuda, Y., & Kondo, K. (1988). Studies on crude drugs effective on visceral larva migrans. V.: the larvicidal principle in mace (aril of Myristica fragrans). Chemical and Pharmaceutical Bulletin, 36, 2685–2688. https://doi.org/10.1248/cpb.36.2685.
Nelson, E. L., & Boeving, E. R. (2015). Precise digit use increases the expression of handedness in Colombian spider monkeys (Ateles fusciceps rufiventris). American Journal of Primatology, 77, 1253–1262. https://doi.org/10.1002/ajp.22478.
NEPA-UNICAMP (2011). Tabela brasileira de composição de alimentos (4th ed.). NEPA–UNICAMP.
Neri-Numa, I. A., Soriano Sancho, R. A., Aparecida Pereira, A. P., & Pastore, G. M. (2018). Small Brazilian wild fruits: nutrients, bioactive compounds, health-promotion properties and commercial interest. Food Research International, 103, 345–360. https://doi.org/10.1016/j.foodres.2017.10.053.
Norconk, M. A. (2007). Sakis, uacaris and titi monkeys: behavioral diversity in a radiation of primate seed predators. In C. J. Campbell, A. Fuentes, K. C. MacKinnon, M. Panger, & S. K. Bearder (Eds.), Primates in perspective (pp. 123–138). Oxford University Press.
Norconk, M. A., & Kinzey, W. G. (1994). Challenge of neotropical frugivory: travel patterns of spider monkeys and bearded sakis. American Journal of Primatology, 34, 171–183. https://doi.org/10.1002/ajp.1350340208.
Oftedal, O. T. (1991). The nutritional consequences of foraging in primates—The relationship of nutrient intakes to nutrient-requirements. Philosophical Transactions of the Royal Society of London, Series B-Biological Sciences, 334, 161–170. https://doi.org/10.1098/rstb.1991.0105.
Palacio, F. X., Siepielski, A. M., Lacoretz, M. V., & Ordano, M. (2020). Selection on fruit traits is mediated by the interplay between frugivorous birds, fruit flies, parasitoid wasps, and seed-dispersing ants. Journal of Evolutionary Biology, 33, 874–886. https://doi.org/10.1111/jeb.13656.
Palmieri, L., & Pereira, R. A. S. (2018). The role of non-fig-wasp insects on fig tree biology, with a proposal of the F phase (Fallen figs). Acta Oecologica, 90, 132–139. https://doi.org/10.1016/j.actao.2017.10.006.
Pennington, T. D. (1990). Sapotaceae. Flora Neotropica Monograph (Vol. 52). New York Botanic Garden.
Pinto, L. B. (2008). Ecologia alimentar do cuxiú-de-variz-vermelho Chiropotes albinasus (Primates: Pitheciidae) na Floresta Nacional do Tapajós, Pará. PhD thesis, Univesidade de Campinas, São Paulo, Brazil.
Pitman, N. C., Terborgh, J. W., Silman, M. R., Núñez, V. P., Neill, D. A., et al (2001). Dominance and distribution of tree species in upper Amazonian terra firme forests. Ecology, 82, 2101–2117. https://doi.org/10.1890/0012-9658(2001)082[2101:DADOTS]2.0.CO;2.
Ponce-Santizo, G., Andresen, E., Cano, E., & Cuarón, A. D. (2006). Primary seed dispersal by primates and secondary dispersal by dung beetles in Tikal, Guatemala. Biotropica, 38, 390–397. https://doi.org/10.1111/j.1744-7429.2006.00144.x.
Prance, G. T. (1979). Notes on the vegetation of Amazonia III. The terminology of Amazonian forest types subject to inundation. Brittonia, 31, 26–38. https://doi.org/10.2307/2806669.
Raman, A. (2012). Gall induction by hemipteroid insects. Journal of Plant Interactions, 7, 29–44. https://doi.org/10.1080/17429145.2011.630847.
Ramos, R. S. (2020). Heisteria Jacq. (Olacaceae) na Amazônia Brasileira. Masters dissertation, Universidade Federal Rural da Amazônia-Museu Paraense Emilio Goeldi, Belém, PA.
Raubenheimer, D., & Rothman, J. M. (2013). Nutritional ecology of entomophagy in humans and other primates. Annual Review of Entomology, 58, 141–160. https://doi.org/10.1146/annurev-ento-120710-100713.
Raubenheimer, D., & Simpson, S. J. (1997). Integrative models of nutrient balancing: application to insects and vertebrates. Nutrition Research Reviews, 10, 151–179. https://doi.org/10.1079/NRR19970009.
Raubenheimer, D., Simpson, S. J., & Mayntz, D. (2009). Nutrition, ecology and nutritional ecology: toward an integrated framework. Functional Ecology, 23, 4–16. https://doi.org/10.1111/j.1365-2435.2009.01522.x.
Raubenheimer, D., Machovsky-Capuska, G. E., Chapman, C. A., & Rothman, J. M. (2015). Geometry of nutrition in field studies: an illustration using wild primates. Oecologia, 177, 223–234. https://doi.org/10.1007/s00442-014-3142-0.
Redford, K. H., da Fonseca, G. A. B., & Lacher Jr., T. E. (1984). The relationship between frugivory and insectivory in primates. Primates, 25, 433–440. https://doi.org/10.1007/BF02381666.
Regan, B. C., Julliot, C., Simmen, B., Viénot, F., Charles–Dominique, P., et al. (2001). Fruits, foliage and the evolution of primate colour vision. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 356, 229-283. https://doi.org/10.1098/rstb.2000.0773
Ribeiro, S. J. E. L., Hopkins, M. J. G., Vicentini, A., Sothers, C., Costa, M. A. S., et al (1999). Flora da Reserva Ducke: guia de identificação das plantas vasculares de uma floresta de terra firme na Amazonia Central. DFID-Instituto Nacional de Pesquisas da Amazônia.
Rosenberger, A. L., Halenar, L., Cooke, S. B., & Hartwig, W. C. (2008). Morphology and evolution of the spider monkey, genus Ateles. In C. Campbell (Ed.), Spider monkeys: behaviour, ecology and evolution of the genus Ateles (pp. 19–49). Cambridge University Press.
Russo, S. E. (2003). Responses of dispersal agents to tree and fruit traits in Virola calophylla (Myristicaceae): implications for selection. Oecologia, 136, 80–87. https://doi.org/10.1007/s00442-003-1239-y.
Russo, S. E., Campbell, C. J., Dew, J. L., Stevenson, P. R., & Suarez, S. A. (2005). A multi-forest comparison of dietary preferences and seed dispersal by Ateles spp. International Journal of Primatology, 26, 1017–1037. https://doi.org/10.1007/s10764-005-6456-2.
Ryan, A. M., Chapman, C. A., & Rothman, J. M. (2013). How do differences in species and part consumption affect diet nutrient concentrations? A test with red colobus monkeys in Kibale National Park, Uganda. African Journal of Ecology, 51, 1–10. https://doi.org/10.1111/j.1365-2028.2012.01346.x.
Scott, J. K., & Black, R. (1981). Selective predation by white-tailed black cockatoos on fruit of Banksia attenuata containing the seed-eating weevil Alphitopis nivea. Australian Wildlife Research, 8, 421–430. https://doi.org/10.1071/WR9810421.
Simmen, B., & Sabatier, D. (1996). Diets of some French Guianan primates: composition and food choices. International Journal of Primatology, 17, 661–693. https://doi.org/10.1007/BF02735260.
Sinclair, R. J., & Hughes, L. (2010). Leaf miners: the hidden herbivores. Austral Ecology, 35, 300–313. https://doi.org/10.1111/j.1442-9993.2009.02039.x.
Steele, M. A., Hadj-Chikh, L. Z., & Hazeltine, J. (1996). Caching and feeding decisions by Sciurus carolinensis: responses to weevil-infested acorns. Journal of Mammalogy, 77, 305–314. https://doi.org/10.2307/1382802.
Stevenson, P. (2004). Fruit choice by woolly monkeys in Tinigua National Park, Colombia. International Journal of Primatology, 25, 367–381. https://doi.org/10.1023/B:IJOP.0000019157.35464.a0.
Stevenson, P. R., & Link, A. (2010). Fruit preferences of Ateles belzebuth in Tinigua Park, northwestern Amazonia. International Journal of Primatology, 31, 393–407. https://doi.org/10.1007/s10764-010-9392-8.
Stevenson, P. R., Quinones, M. J., & Ahumada, J. Á. (1994). Ecological strategies of woolly monkeys (Lagothrix lagotricha) at Tinigua National Park, Colombia. American Journal of Primatology, 32, 123–140. https://doi.org/10.1002/ajp.1350320205.
Su, H. C. F. (1989). Effects of Myristica fragrans fruit (Family: Myristicaceae) to four species of stored-product insects. Journal of Entomological. Science, 24, 168–173. https://doi.org/10.18474/0749-8004-24.2.168.
Sugiura, S., & Yamazaki, K. (2009). Gall-attacking behavior in phytophagous insects, with emphasis on Coleoptera and Lepidoptera. Terrestrial Arthropod Reviews, 2, 41–61. https://doi.org/10.1163/187498309X435658.
Symington, M. M. (1990). Fission-fusion social organization in Ateles and Pan. International Journal of Primatology, 11, 47–61. https://doi.org/10.1007/BF02193695.
Take, M. (2017). Comparative ecology of three primate species in urban forest fragments in Brazilian Amazonia. MSc thesis, Kyoto University, Japan. 87 pages.
Tollrian, R., & Harvell, C. D. (1999). The ecology and evolution of inducible defenses. Princeton University Press.
Traveset, A., Willson, M. F., & Gaither Jr., J. C. (1995). Avoidance by birds of insect-infested fruits of Vaccinium ovalifolium. Oikos, 73, 381–386. https://doi.org/10.2307/3545962.
U.S. Department of Agriculture. (2019). Agricultural Research Service. FoodData Central. http://fdc.nal.usda.gov/
Uriarte, M., Anciães, M., Silva, M. T. B., Rubim, P., Johnson, E., et al (2011). Disentangling the drivers of reduced long-distance seed dispersal by birds in an experimentally fragmented landscape. Ecology, 92, 924–937. https://doi.org/10.1890/10-0709.1.
Utzurrum, R. C. B., & Heideman, P. D. (1991). Differential ingestion of viable vs. nonviable Ficus seeds by fruit bats. Biotropica, 23, 311–312.
Valburg, L. K. (1992). Feeding preferences of common bush-tanagers for insect-infested fruits: avoidance or attraction? Oikos, 65, 29–33. https://doi.org/10.2307/3544884.
Valero, A. (2004). Spider monkey (Ateles geoffroyi yucatanensis) travel patterns in a subtropical forest of Yucatan, Mexico. PhD Thesis, St. Andrews University.
van Roosmalen, M. G. M. (1985). Fruits of the Guianan flora. University of Utrecht Press.
van Roosmalen, M. G., & Klein, L. L. (1988). The spider monkeys, genus Ateles. In R. A. Mittermeier, A. B. Rylands, A. Coimbra-Filho, & G. A. B. Fonseca (Eds.), Ecology and Behavior of Neotropical Primates (Vol. 2, pp. 455–539). World Wildlife Fund.
Virgolin, L. B., Seixas, F. R. F., & Janzantti, N. S. (2017). Composition, content of bioactive compounds, and antioxidant activity of fruit pulps from the Brazilian Amazon biome. Pesquisa Agropecuária Brasileira, 52, 933–941. https://doi.org/10.1590/S0100-204X2017001000013.
Wehncke, E. V., Hubbell, S. P., Foster, R. B., & Dalling, J. W. (2003). Seed dispersal patterns produced by white-faced monkeys: implications for the dispersal limitation of neotropical tree species. Journal of Ecology, 91, 677–685. https://doi.org/10.1046/j.1365-2745.2003.00798.x.
Weiblen, G. D. (2002). How to be a fig wasp. Annual Review of Entomology, 47, 299–330. https://doi.org/10.1146/annurev.ento.47.091201.145213.
Willson, M. F., & Traveset, A. (2000). The ecology of seed dispersal. In M. Fenner (Ed.), Seeds: the ecology of regeneration in plant communities (pp. 85–110). CAB International.
Wilson, A. (2008). Insect frugivore interactions: the potential for beneficial and neutral effects on host plants. Queensland University of Technology.
Wittmann, F., Schöngart, J., Montero, J. C., Motzer, T., Junk, W. J., et al (2006). Tree species composition and diversity gradients in white-water forests across the Amazon Basin. Journal of Biogeography, 33, 1334–1347. https://doi.org/10.1111/j.1365-2699.2006.01495.x.
Worman, C. O. D., & Chapman, C. A. (2005). Seasonal variation in the quality of a tropical ripe fruit and the response of three frugivores. Journal of Tropical Ecology, 21, 689–697. https://doi.org/10.1017/S0266467405002725.
Wrangham, R. W., Conklin, N. L., Etot, G., Obua, J., Hunt, K. D., et al (1993). The value of figs to chimpanzees. International Journal of Primatology, 14, 243–256. https://doi.org/10.1007/BF02192634.
Xiaoming, C., Ying, F., Hong, Z., & Zhiyong, C. (2010). Review of the nutritive value of edible insects. In P. B. Durst, D. V. Johnson, R. N. Leslie, & K. K. Shono (Eds.), Edible forest insects: humans bite back (pp. 85–92). FAO.
Yuyama, L. K., Aguiar, J. P., Yuyama, K., Clement, C. R., Macedo, S. H. M., et al (2003). Chemical composition of the fruit mesocarp of three peach palm (Bactris gasipaes) populations grown in Central Amazonia, Brazil. International Journal of Food Sciences and Nutrition, 54, 49–56. https://doi.org/10.1080/0963748031000061994.
Zamora, R. (2000). Functional equivalence in plant-animal interactions: ecological and evolutionary consequences. Oikos, 88, 442–447 https://www.jstor.org/stable/3547039.
Zare, H., Moosavi-Movahedi, A. A., Salami, M., Mirzaei, M., Saboury, A. A., et al (2013). Purification and autolysis of the ficin isoforms from fig (Ficus carica cv. Sabz) latex. Phytochemistry, 87, 16–22. https://doi.org/10.1016/j.phytochem.2012.12.006.
Acknowledgments
TCS-B thanks the staff and members of the Department of Nutrition, University Centre Manaus-FAMETRO. TdeO thanks CNEC/WorleyParsons and ELETROBRÁS for financial support of the mammal survey in the Tapajós river basin. AAB thanks Maracajá Ecological Consulting for the invitation to conduct the primate survey work. BB is partially supported by a CNPq-Productivity grant (309256/2019-4), ALM is supported by Capes (Finance code 0001), AAB is supported by PNPD/CAPES (Process no. 8887.470331/2019-00). This paper is contribution XX from the Amazonian Mammals Research Group, and contribution number XX from the Igapó Study Project. We thank the anonymous reviewers for their helpful comments, and Stephan Nash for permission to use his Ateles images in Fig. 1.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Joanna Setchell
Rights and permissions
About this article

Cite this article
dos Santos-Barnett, T.C., Cavalcante, T., Boyle, S.A. et al. Pulp Fiction: Why Some Populations of Ripe-Fruit Specialists Ateles chamek and A. marginatus Prefer Insect-Infested Foods. Int J Primatol 43, 384–408 (2022). https://doi.org/10.1007/s10764-022-00284-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-022-00284-0