
Abstract
In nonhuman primates, infanticide by adult males can occur when the leader male is ousted from a one-male, multifemale group, or when male dominance rank changes within a multimale, multifemale group. According to the sexual selection hypothesis, this behavior may be adaptive if perpetrators increase their reproductive success by killing unrelated, unweaned infants, thus shortening the interbirth interval of the mother, and then siring her next infant. Under an alternative hypothesis, infanticide is a byproduct of aggressive male–male competition and these predictions do not hold. Direct observations of the context surrounding infanticide in free-ranging primate populations that allow a test of these predictions are rare. Here, we document four cases of male infanticide and report paternity data for a group of golden snub-nosed monkeys (Rhinopithecus roxellana) at Shennongjia, China. Three cases of infanticide by new leader males supported the predictions of the sexual selection hypothesis, while another provides partial support for the sexual selection hypothesis, but can also be explained via a nonadaptive hypothesis. In this latter case, a male from an all-male group killed an infant during an aggressive episode that appeared to be accidental, as it took place 7 mo before a male takeover happened, and the perpetrator did not obtain any reproductive advantage. We conclude that most male infanticide events in golden snub-nosed monkeys are consistent with the adaptive selection sexual hypothesis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Infanticide has been observed in a variety of mammals, such as primates, carnivores, and rodents (Lukas and Huchard 2014) and is especially prevalent in primates (van Schaik 2000). The sexual selection hypothesis holds that male infanticide is adaptive (Hrdy 1974, 1979; Palombit 2012; van Schaik 2000). Under this model, infanticidal males improve their reproductive success by killing an unrelated and unweaned infant, thereby shortening the mother’s interbirth interval. These males then sire the next offspring of the victim’s mother (Hrdy 1979). As lactation suppresses sexual receptivity in many mammals (Bronson 1989), the death of an unweaned infant causes the mother to resume cycling, at least in nonseasonally breeding species. An alternative, the nonadaptive hypothesis holds that male infanticide is a byproduct of generalized male–male aggression (Bartlett et al. 1993) and occurs accidentally during aggressive episodes (Sussman et al. 1995). Under this hypothesis, the killer does not gain any reproductive advantage.
It has been assumed that the sexual selection hypothesis does not apply to seasonal breeders, because females will not resume cycling earlier as a result of the loss of an infant, so there is no reproductive advantage for the perpetrator (Wright 1995). However, two studies have shown that this is not necessarily the case (Borries et al. 1999; Soltis et al. 2000), and that if mean interbirth intervals are >1 yr, then killing a dependent infant may still reduce the interbirth interval, even in seasonal species, and therefore be advantageous for the male.
Although infanticide has been directly observed or inferred in a variety of primate taxa (Bellemain et al. 2006; Cords and Fuller 2010; Lukas and Huchard 2014; Palombit 2012; van Schaik 2000), few studies have presented the behavioral and molecular evidence needed to test the sexual selection hypothesis. In a study of Hanuman langurs (Semnopithecus entellus), the male was excluded as the father of the infant he attacked or killed in all 16 cases of attempted or successful infanticide where DNA samples were available for both adult males and infants; in the four cases of births following infanticides, the presumed killers were the likely fathers of these offspring (Borries et al. 1999). Male infanticide in Japanese macaques (Macaca fuscata) also met the conditions specified by the sexual selection hypothesis, because the study authors were able to exclude the attacker as the father of the victim in 22 of the 23 cases examined (Soltis et al. 2000).
The golden snub-nosed monkey (Rhinopithecus roxellana) is an Asian colobine that features a multilevel social organization in which a number of one-male, multifemale units (OMUs) form a stable and cohesive band numbering up to several hundred individuals (Grueter 2013). In addition to the constituent OMUs, one or several all-male units of up to 25 adult and subadult males associate closely with the band (Grueter 2013) but are usually prevented from mating (Xiang et al. 2014). Aggression between males is intense, especially during the mating season (Xiang et al. 2014; Zhao et al. 2005). Solitary males and males in all-male units periodically invade OMUs, replacing the former leader or mating with resident females (Qi et al. 2009; Yao et al. 2011; Zhao et al. 2005). Male OMU leaders cannot effectively monopolize access to resident females, and extra-unit males sire more than half of the offspring in the Qingling population (Guo et al. 2010). Copulations occur throughout the year, but particularly between May and November, and conception appears to be limited to the period from September to November with a corresponding birth from March to May (Xiang et al. 2014). Although infanticide has been reported in snub-nosed monkeys (R. roxellana: Zhang et al. 1999; R. bieti: Ren et al. 2011; Xiang and Grueter 2007), no study has yet tested evidence for whether this infanticide might be adaptive.
We here describe four instances of male infanticide that occurred during >8 yr of observation of a group of golden snub-nosed monkeys, with genetic analysis of paternity. We test the predictions of the sexual selection hypothesis: 1) killers are unrelated to the victims; 2) the victims’ mothers conceived shortly after infanticide, and infant death shortens the length of the mother’s interbirth interval; and 3) the killer sires the subsequent offspring of the victim’s mother. We compare this adaptive hypothesis against the null mode, whereby infanticide occurs as a byproduct of aggression between a leader male and individuals from an all-male unit, and the infant is killed by a male that gains no reproductive benefit.
Methods
Study Site
We conducted the study on a free-ranging, provisioned group of golden snub-nosed monkeys at Dalongtan in Shennongjia Nature Reserve, Hubei province, China (coordinates: 31°29′N, 110°18′E). Reserve staff fed the monkeys two or three times a day at different sites with lichen, pine seeds, apples, carrots, oranges, and peaches; they did not herd the monkeys to the provisioning sites.
From January 2006 to March 2014, we conducted daily observations at distances between 3 and 50 m from 0800 to 1600 h in winter (December–February) and spring (March–May) and 07:00 to 19:00 h in summer (June–August) and autumn (September–November) so long as weather permitted (Xiang et al. 2014; Yu et al. 2013). We observed and recorded male infanticides ad libitum (Altmann 1974).
We collected the dead bodies of four infants attacked by males. We preserved tissue samples at −20°C in a freezer before use in paternity exclusion analysis. We collected hairs with attached roots from all reproductively mature adult males in the focal group and newborn infants of an infanticide victim’s mother for paternity analysis (for details, see Xiang et al. 2014).
Data Analysis
We used 16 microsatellite loci to determine the paternity of the dead infants and the subsequent infant of the victim's mother, using all reproductively mature adult males (N = 12) in the study group as candidate sires (Xiang et al. 2014; Yang et al. 2014). We used CERVUS3.0 to determine paternity (Kalinowski et al. 2007) with confidence levels of >80% (relaxed confidence) in one case and >95% (strict confidence) in four cases.
To determine whether the victims’ mothers became sexually receptive shortly after infanticide, we estimated conception dates by subtracting gestation length from birth dates. As there are no accurate records of gestation length for wild individuals, we used the mean gestation of 202 days (SD = 1.1 days, N = 5) for captive individuals (Qi et al. 1995; Yan et al. 2003).
To test whether infanticide shortens the length of the interbirth interval, we used two-sample t-tests to compare the interbirth interval of females that lost infants to infanticide with those that did not suffer infanticide.
Ethical Note
All research methods adhered to Chinese legal requirements and complied with protocols approved by the State Forestry Administration of China. Because the field studies involved endangered and protected species, we obtained approval from the state and local governments. We collected data in a minimally invasive manner and recorded naturally occurring behavior.
Results
We observed three infants being killed by adult males, and inferred one further case of infanticide based on strong evidence (Table I). Autopsy confirmed that attacks by males contributed to the infant’s death in all cases.
Infanticide Cases 1 and 2
Infanticide cases 1 and 2 occurred in the same OMU and the killer was the same adult male (male DD). Case 1 occurred 7 mo before the killer ousted the old leader male; case 2 took place 4 d after the killer became the new leader male.
In December 2006, the study group consisted of three OMUs with a total of 37 individuals and one all-male unit (five adults and three juveniles). Male CM was the leader male of an OMU that consisted of three adult females (JJ, HZ, and JZ), three juveniles, and one unweaned infant in December 2006. Adult males XY and DD from the all-male unit chased male CM together at 13:20 h on December 23, 2006. CM mounted a successful defense against XY and DD with the help of other leader males (HT and BT). However, during the aggressive incident, male DD was observed biting female HZ’s unweaned infant (HZI06), who was aged 269 days. HZ106 bled from bites to the abdomen and the left thigh and died four days later (infanticide case 1).
On 26 June 26 2007, male XY evicted leader male CM from his OMU and XY became the leader of the OMU, which consisted of three adult females (JJ, HZ, and JZ); three juveniles; and one unweaned infant, born on March 26, 2007. On August 5, male DD was observed chasing and stalking leader male XY. On August 6, a serious fight between males DD and XY resulted in XY leaving the unit and joining the all-male unit, and DD became the new leader of the OMU. At 13:10 h on August 10, DD attacked female JZ’s unweaned infant (JZI07), aged 142 days, although females JJ and JZ attempted to protect it. The infant suffered serious injuries to the abdomen and neck and died several minutes later (infanticide case 2).
Infanticide Cases 3 and 4
Infanticide cases 3 and 4 occurred in the same OMU in 2012 after it was taken over by a male from the all-male unit (male NN).
In May 2012, the study group consisted of four OMUs with a total of 43 individuals and one all-male unit (four adults and seven subadults and juveniles). On May 15, 2012, male NN from the all-male unit evicted leader male DY and became the new leader of an OMU composed of three adult females (XH, XB, and HH3); two juveniles; and two unweaned infants. Female XB had given birth to XBI12 on April 19. Female HH3 gave birth to HH3I12 on April 2, but abandoned the infant at the age of 2 days due to a serious eye illness (to female HH3). Infant HH3I12 was adopted by its older sister female XH, who was lactating, but whose own infant had just died.
Infant HH3I12’s biological mother, female HH3, could not keep up with her social unit and was separated from other members and usually associated with an adult male (CM) from the all-male unit from May 25, 2012. Leader male NN often tried to herd her to his social unit, but was unsuccessful. When her eyesight partly recovered female HH3 returned to nurse her baby from time to time after April 14, 2012 (16% of nursing time; Z. F. Xiang, in preparation). Female HH3 was observed to mate with leader male NN on July 3, 2012 and returned to her original OMU on September 22, 2012, when she again mated with leader male NN.
At 14:53 h on August 20, 2012, female XH was grooming female XB. Female XH’s adopted infant (HH3I12) and XB’s infant (XBI12), both unweaned, were playing with a juvenile in close proximity to their mothers; male NN, the new leader male, was also near them. Suddenly, male NN rushed toward and bit HH3I12. Females HH3, XH, and XB rushed toward male NN and retrieved their offspring. Male NN attacked infant HH3I12 again at 15:52 h on August 29 and at 15:22 h on September 3, but females XH and HH3 saved the infant. On the morning of September 15, infant HH3I12 was found dead at the sleeping site, with 13 punctures and slash wounds on the body, at least eight of them from teeth (infanticide case 3).
At 10:08 h on September 20, 2012, new leader male NN attacked and bit infant XBI12, although females XB and XH fought male NN. Female XB held infant XBI12 tightly against her chest, but the infant died within a few minutes (infanticide case 4). Female XB carried the corpse for 10 min, then abandoned it. The injuries were two obvious holes in the right abdomen (Fig. 1).

An infant golden snub-nosed monkey with a fatal injury caused by a new leader male.
Predictions of the Sexual Selection Hypothesis
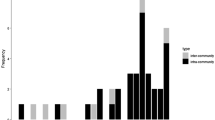
We could exclude the perpetrator as the father of the victims based on genotype in all four cases of infanticide (Table II). To assess potential effects of infanticide on interbirth intervals, we assessed all five mothers, including both the biological and foster mothers from case 3, and compared their interbirth intervals to those of other mothers. The mean interbirth interval of mothers whose infant were killed by a male (N = 5) was significantly shorter than that of mothers with surviving infants (N = 37) (Fig. 2, t 4,36 = 4.06, P < 0.001). However, in case 1 the interbirth interval (749 days) was similar to that of a mother with a surviving infant, while in cases 2, 3, and 4, the interval was much shorter (Fig. 2). The victims’ mothers conceived 53.2 ± 11.8 days (mean ± SD) after the infanticide and all gave birth in the following birth season (Table I). In cases 1, 2, and 4, the infanticidal male sired the next infant born to the victim’s mother (Table II). In case 3, the adopted infant, the infanticidal male did not sire the next infant of the victim’s biological mother. However, the infanticidal male did sire the stepmother’s next offspring.

Median interbirth interval in females with and without infanticide.
Support for the Nonadaptive Hypothesis
In case 1, the infant was killed by an adult male from the all-male unit 7 mo before the killer took over the mother’s OMU, while in cases 2, 3, and 4, infanticides occurred after the new male leader evicted the older one.
Discussion
In this study, we documented four instances of male infanticide in a group of golden snub-nosed monkeys. Our results support the sexual selection hypothesis in three cases, but although the fourth provides partial support for the sexual selection hypothesis, it can also be explained via a nonadaptive hypothesis.
In two cases (2 and 4), the context in which the behaviors were observed and the genetic data support the sexual selection hypothesis. The infanticidal male killed an unrelated, unweaned infant; the infant death resulted in a shorter interbirth interval than in females with surviving infants; and the killer sired the next infant of the victim’s mother’s (Table II). These findings are consistent with both observed and inferred incidents of male infanticide in other primates, such as Hanuman langurs (Borries 1997; Borries et al. 1999).
In case 3 the killer did not sire the subsequent offspring of the victim’s biological mother although she resumed cycling. At first glance, this result does not support the sexual selection hypothesis. However, the victim was adopted and nursed primarily by its older sister, the death of the infant led to that female resuming cycling, and the killer did sire the adoptive mother’s next infant, supporting the sexual selection hypothesis.
In case 1, the killer killed an unrelated infant and sired the subsequent infant of the victim’s mother, but infanticide did not shorten the interval to that infant’s birth (Table I, Fig. 2), so the killer did not increase his reproductive success beyond that accrued as new leader male. This case offers only partial support for the sexual selection hypothesis. It is also consistent with the byproduct of aggression hypothesis, as the infant’s death resulted from male–male aggression well before the infanticidal male’s eventual takeover of the unit 7 mo later. In this case, killing a female infant may also have harmed the killer’s future reproductive success, as he was the leader of the victim’s OMU from August 2007 to October 2013 (Z. Xiang, unpubl. data). If the female infant had survived and reached maturity within the unit, she could have mated with him and given birth between 2010 and 2013.
In sum, three of four male infanticide events in golden snub-nosed monkeys are consistent with the adaptive selection sexual hypothesis, while another provides partial support for the sexual selection hypothesis, but can also be explained via a nonadaptive hypothesis. These results provide further evidence that sexually selected infanticide can occur in seasonally breeding primates and provide the first evidence for sexually selected infanticide in a species with a multilevel social organization.
References
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour, 49, 227–267.
Bartlett, T. Q., Sussman, R. W., & Cheverud, J. M. (1993). Infant killing in primates: a review of observed cases with specific reference to the sexual selection hypothesis. American Anthropologist, 95, 958–990.
Bellemain, E., Swenson, J. E., & Taberlet, P. (2006). Mating strategies in relation to sexually selected infanticide in a non-social carnivore: the Brown Bear. Ethology, 112, 238–246.
Borries, C. (1997). Infanticide in seasonally breeding multimale groups of Hanuman langurs (Presbytis entellus) in Ramnagar (South Nepal). Behavioral Ecology and Sociobiology, 41, 139–150.
Borries, C., Launhardt, K., Epplen, C., Epplen, J. T., & Winkler, P. (1999). DNA analyses support the hypothesis that infanticide is adaptive in langur monkeys. Proceedings of the Royal Society of London B: Biological Sciences, 266, 901–904.
Bronson, F. H. (1989). Mammalian reproductive biology. Chicago: University of Chicago Press.
Cords, M., & Fuller, J. L. (2010). Infanticide in Cercopithecus mitis stuhlmanni in the Kakamega Forest, Kenya: variation in the occurrence of an adaptive behavior. International Journal of Primatology, 31, 409–431.
Grueter, C. C. (2013). The biology of snub-nosed monkeys, douc langurs, proboscis monkeys and simakobu. Hauppauge: Nova.
Guo, S., Ji, W., Li, M., Chang, H., & Li, B. (2010). The mating system of the Sichuan snub-nosed monkey (Rhinopithecus roxellana). American Journal of Primatology, 72, 25–32.
Hrdy, S. B. (1974). Male-male competition and infanticide among the langurs (Prebytis entellus) of Abu, Rajasthan. Folia Primatologica, 22, 19–58.
Hrdy, S. B. (1979). Infanticide among animals: a review, classification, and examination of the implications for the reproductive strategies of females. Ethology and Sociobiology, 1, 13–40.
Kalinowski, S. T., Taper, M. L., & Marshall, T. C. (2007). Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Molecular Ecology, 16, 1099–1106.
Lukas, D., & Huchard, E. (2014). The evolution of infanticide by males in mammalian societies. Science, 346(6211), 841–844.
Palombit, R. A. (2012). Infanticide: Male strategies and female counterstrategies. In J. C. Mitani, J. P. Call, P. M. Kappeler, R. A. Palombit, & J. B. Silk (Eds.), Evolution of primate societies (pp. 432–468). Chicago: University of Chicago Press.
Qi, H. J., Liang, B., Bao, W. Y., Jia, Y. C., Hama, N., Czekala, N. M., & Harvey, N. C. (1995).The hormone change in the urine of female snub-nosed monkeys. ActaTheriologica Sinica, 15(2), 106–112.
Qi, X. G., Li, B. G., Garber, P. A., Ji, W., & Watanabe, K. (2009). Social dynamics of the golden snub-nosed monkey (Rhinopithecusroxellana): female transfer and one-male unit succession. American Journal of Primatology, 71, 670–679.
Ren, B., Li, D., He, X., Qiu, J., & Li, M. (2011). Female resistance to invading males increases infanticide in Langurs. PLoS ONE, 6(4), e18971.
Soltis, J., Thomsen, R., Matsubayashi, K., & Takenaka, O. (2000). Infanticide by resident males and female counter-strategies in wild Japanese macaques (Macacafuscata). Behavioral Ecology and Sociobiology, 48, 195–202.
Sussman, R. W., Cheverud, J. M., & Bartlett, T. Q. (1995). Infant killing as an evolutionary strategy: reality or myth? Evolutionary Anthropology, 3, 149–151.
van Schaik, C. P. (2000). Vulnerability to infanticide by males: Patterns among mammals. In C. P. van Schaik & C. H. Janson (Eds.), Infanticide by males and its implications (pp. 61–71). Cambridge: Cambridge University Press.
Wright, P. C. (1995). Demography and life history of free-ranging Propithecus diadema edwardsii in Ranomafana National Park, Madagascar. International Journal of Primatology, 16, 835–854.
Xiang, Z. F., & Grueter, C. C. (2007). First direct evidence of infanticide and cannibalism in wild snub-nosed monkeys (Rhinopithecus bieti). American Journal of Primatology, 69, 249–254.
Xiang, Z. F., Yang, B. H., Yu, Y., Yao, H., Grueter, C. C., Garber, P. A., & Li, M. (2014). Males collectively defend their one-male units against bachelor males in a multi-level primate society. American Journal of Primatology, 76, 609–617.
Yan, C. E., Jiang, Z. G., Li, C. W., Zen, Y., Tan, N. N., & Xia, S. Z. (2003). Monitoring the menstrual cycle and pregnancy in the Sichuan golden monkey (Rhinopithecus roxellana) by measuring urinary estradiol and progesterone. Acta Zoologica Sinica, 49, 693–697.
Yang, B., Ren, B., Xiang, Z., Yang, J., Yao, H., Garber, P. A., & Li, M. (2014). Major histocompatibility complex and mate choice in the polygynous primate: the Sichuan snub-nosed monkey (Rhinopithecus roxellana). Integrative Zoology. doi:10.1111/1749-4877.12084.
Yao, H., Liu, X., Standford, C., & Li, Y. (2011). Male dispersal in a provisioned multilevel group of Rhinopithecus roxellana in Shennongjia Nature Reserve, China. American Journal of Primatology, 73, 1280–1288.
Yu, Y., Xiang, Z. F., Yao, H., Grueter, C. C., & Li, M. (2013). Female snub-nosed monkeys exchange grooming for sex and infant handling. PLoS ONE, 8(9), e74822.
Zhang, S. Y., Liang, B., & Wang, L. X. (1999). Infanticide within captive groups of Sichuan golden snub-nosed monkeys (Rhinopithecus roxellana). Folia Primatologica, 70, 274–276.
Zhao, D., Li, B., Li, Y., & Wada, K. (2005). Extra-unit sexual behaviour among wild Sichuan snub-nosed monkeys (Rhinopithecus roxellana) in the Qinling Mountains of China. Folia Primatologica, 76, 172–176.
Acknowledgments
This study was supported by the National Key Technology R & D Program of China (2013BAD03B03), the Opening Project of Hubei Key Laboratory of Conservation of the Golden Snub-nosed Monkey (2014snjac003), and the State Forestry Administration of China. We would like give special thanks to Dr. Joanna Setchell for her constructive suggestions and extensive editing of the manuscript. We also thank Dominic Cram, Dieter Lukas, and Alicia Krzton for useful suggestions and editing the English in the manuscript. We thank James Higham, Elise Huchard, and one anonymous reviewer for their valued suggestions for revisions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Hui Yao and Huiliang Yu contributed equally to this work.
Rights and permissions
About this article

Cite this article
Yao, H., Yu, H., Yang, B. et al. Male Infanticide in the Golden Snub-Nosed Monkey (Rhinopithecus roxellana), a Seasonally Breeding Primate. Int J Primatol 37, 175–184 (2016). https://doi.org/10.1007/s10764-016-9892-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-016-9892-2