
Abstract
Invasive alien aquatic plant species (IAAPs) cause serious ecological and economic impact and are a major driver of changes in aquatic plant communities. Their invasive success is influenced by both abiotic and biotic factors. Here, we summarize the existing knowledge on the biology of 21 IAAPs (four free-floating species, eight sediment-rooted, emerged or floating-leaved species, and nine sediment-rooted, submerged species) to highlight traits that are linked to their invasive success. We focus on those traits which were documented as closely linked to plant invasions, including dispersal and growth patterns, allelopathy and herbivore defence. The traits are generally specific to the different growth forms of IAAPs. In general, the species show effective dispersal and spread mechanisms, even though sexual and vegetative spread differs strongly between species. Moreover, IAAPs show varying strategies to cope with the environment. The presented overview of traits of IAAPs will help to identify potential invasive alien aquatic plants. Further, the information provided is of interest for developing species-specific management strategies and effective prevention measures.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Invasive alien aquatic plants (IAAPs) are considered a serious threat to aquatic ecosystems throughout the world. Due to their nuisance growth, IAAPs have both significant economic and ecological impact (Holm et al., 1969, Halstead et al., 2003, Stiers et al., 2011a; Santos et al., 2011). Consequently, management options are ongoing to reduce the impact of IAAPs, which are highly cost-intensive and resource demanding (Hussner et al., 2017; Simberloff, in press).
Interestingly, the proportion of introduced plant species, which become invasive is much higher in aquatic than in terrestrial habitats (Hussner, 2012). The reasons for this strong imbalance are still not clear, even though the uniformity of freshwater habitats is commonly considered as important for the broad distribution ranges of aquatic plants (Santamaria, 2002; but see Murphy et al., 2019). Moreover, it must be borne in mind that most alien aquatic plant introductions, similar to terrestrial plants (Reichard & White, 2001), occur via the horticultural trade (Brunel, 2009; Hussner et al., 2014a). In contrast to terrestrial plants, which are commonly cultivated in gardens and often escape from cultivation into natural habitats via seeds (e.g. Heracleum mantegazzianum; Thiele & Otte, 2008), particularly aquatic plants cultivated in aquaria require a human-mediated, intended or unintended, transport into water bodies (Maki & Galatowitsch, 2004; Champion et al., 2010; Hussner et al., 2010; Hill et al., 2020). In fact, most traded aquatic plant species are particularly robust and able to grow under varying conditions in aquaria and under different field conditions, suggesting that a wide ecological amplitude is a relevant trait-promoting invasiveness of IAAPs (Azan et al., 2015).
Nonetheless, numerous studies deal with the question, why do some introduced aquatic plants become invasive, but not others? These studies consider varying aspects of alien aquatic plant invasions, particularly focussing on
-
(i)
pathways of introduction (Maki & Galatowitsch, 2004; Brunel, 2009; Martin & Coetzee, 2011; Hussner et al., 2014a);
-
(ii)
the empty niche theory (Fleming & Dibble, 2015);
-
(iii)
the role of environmental (particularly nutrients) and climatic conditions (James et al., 2006; Gillard et al., 2017a);
-
(iv)
changes in the invasive success of IAAPs due to global change (Wu & Ding, 2019); including climate change (Netten et al., 2010, 2011; Calvo et al., 2019);
-
(v)
the presence/absence of herbivores in its introduced range and the enemy release hypothesis (Fleming & Dibble, 2015; Grutters et al., 2017a; Petruzzella et al., 2017; Redekop et al., 2018; Pulzatto et al., 2018); and
-
(vi)
the role of aquatic plants and plant communities for the establishment of introduced alien aquatic plants (Thiebaut & Martinez, 2015; Chadwell & Engelhardt, 2008; Thouvenot et al., 2019).
There is general concern about questioning the role of species biology on their invasive success (Goodwin et al., 1999; Kolar & Lodge, 2001). The species’ biology and the capacity to acclimate to new/changing conditions determine the growth and competitive strength and thus contribute to the invasiveness of a species. In fact, many studies use growth and competition experiments between native and invasive alien aquatic plant species to identify plant traits explaining the success or failure of the alien species. The outcomes of such studies are limited due to species selection and experimental growth conditions, but do give an idea of relative competitive ability. Furthermore, the role of competitive strength for the success of invasive species differs between the different stages of the invasion process (Vilà & Weiner, 2004).
The performance of a given plant in an environment depends on its capacity to respond to abiotic environmental factors or to avoid detrimental effects originating from biotic interactions such as competition, herbivory or pathogen attack. Specific plant characteristics or attributes linked to morphological, anatomical, physiological, biochemical or phenological “traits” will determine the overall performance of a species (Kattge et al., 2011).
Identifying and understanding plant traits that contribute to the invasiveness of alien species is a key component for the identification of further potential invasive species (Kolar & Lodge, 2001; Jacobs & MacIsaac, 2009). The higher the tolerance of a species to varying environmental conditions, the higher the likelihood to become a successful invader (Higgins & Richardson, 2014). Even though this has been studied intensely for terrestrial plants (e.g. van Kleunen et al. 2011), only a few studies focussed on plant attributes, characteristics or traits (terms often used synonymously) that might be linked to successful aquatic plant invasions (Fleming & Dibble, 2015; Brundu, 2015), and a comprehensive overview for aquatic invasive plants is still lacking.
Nevertheless, it seems reasonable that a large variety of plant traits contribute to the invasiveness of an alien aquatic plant, even though the relevance of a specific trait might differ between habitats, environmental conditions and growth form of the species. A recent paper by Azan et al. (2015), focussing on aquatic invasive plants in Canada, identified some traits associated with the most nuisance species, causing significant negative effects to aquatic ecosystems. These include growth form, growth rate, seed (or spore) and asexual propagule production and dispersal mechanisms, tolerance to environmental conditions (i.e light, temperature, water availability, salinity, pH, nutrients) and phenotypic plasticity, including growth (leaf length, stem length) and fruit parameters (e.g. fruit length, fruit type, time of flowering).
Here, we summarize the existing knowledge on aquatic plant traits, which are reported to contribute to the invasiveness of IAAPs, and discuss their relevance under different environmental and growth conditions. We focus on the IAAPs which were selected as the most invasive IAAPs worldwide in a recent review on management aspects by Hussner et al. (2017), and focus on the traits identified by Azan et al. (2015) (Table 1). Our species list included four free-floating species (Azolla filiculoides Lam., Eichhornia crassipes (Mart.) Solms, Pistia stratiotes L. and Salvinia molesta D. Mitch), eight sediment-rooted species with emerged and/or floating leaves (Alternanthera philoxeroides (Mart.) Griseb., Crassula helmsii (Kirk) Cockayne, Hydrocotyle ranunculoides L. fil., Hygrophila polysperma T. Anderson, Ludwigia grandiflora (Michx.) Greuter & Burdet, Ludwigia peploides (Kunth) P.H. Raven, Myriophyllum aquaticum (Vell.) Verdc. and Trapa natans L.) as well as nine sediment-rooted, submerged species (Cabomba caroliniana A. Gray, Ceratophyllum demersum L., Egeria densa Planch., Elodea canadensis Michx., Elodea nuttallii H. St. John, Hydrilla verticillata (L.f.) Royle, Lagarosiphon major (Ridl.) Moss, Myriophyllum heterophyllum Michx. and Myriophyllum spicatum L.).
Methods
We reviewed the scientific and grey literature, unpublished papers and reports. A comprehensive web search was carried out using google scholar from January to May 2020, and from August to September 2020, as this database includes both published scientific and grey literature. We searched for “trait*” or “attribute*” and “invasive aquatic plant*” or “alien aquatic plant*” or “aquatic weed*” or “invasive macrophyte*”. In addition, we searched for references to each specific trait or attribute considered here by using “specific trait*” or “specific attribute*” and “invasive aquatic plant*” or “alien aquatic plant*” or “aquatic weed*”.
The search for allelochemical traits was based on a search for (“macrophyte*” or “aquatic plant*”) and “allelo*” or (“herbivor*” and (“deterrent*” or “induc*” or “inhib*”).
The list of traits, including dispersal or spread patterns and growth characteristics, was compiled following the suggestions made by Azan et al. (2015) or Fleming & Dibble (2015). We added “clonal integration” as a new trait, which has received increasing attention during the last years in IAAPs research.
Biological traits which contribute to the invasiveness of IAAPs
Growth form
IAAPs occur in different growth forms, which can be simply differentiated into (i) free-floating, (ii) sediment rooted with emerged and/or floating leaves and (iii) submerged (see Hussner et al., 2017). The growth form determines the habitat types which can be invaded by an IAAP (Table 2). Free-floating species can generally grow on any type of water body irrespective of water depth (or even under drained conditions, see section on drought resistance) (EPPO, 2017a, c). The invasion by submerged IAAPs is limited to a certain depth of the water column (due to the species-specific light requirements; Sand-Jensen, 1989); and sediment-rooted emerged IAAPs invasions are usually limited to shallow water bodies and habitats with waterlogged sediments or moist soils (Thouvenot et al., 2013b).
However, despite having a predominant growth form, most IAAPs are able to grow or at least survive in another growth form and withstand unfavourable habitat conditions for a certain period of time (Table 1). Even submerged or free-floating IAAPs may have terrestrial forms, for example M. heterophyllum (Gross et al., 2020) or E. crassipes (Wright & Purcell, 1995; Venter et al., 2017) (Table 1). These IAAPs can invade habitats with rapid changes in water availability and, moreover, can regrow after disposal on the shore of water courses during mechanical control measures. Nevertheless, the ability to occur as more than one growth form extends the list of habitats which are prone to invasion by a given IAAP, as has been documented for, e.g. M. aquaticum (from drained habitats to fast flowing river sections; Hussner & Lösch, 2005; Wersal & Madsen, 2011; Table 2).
The traits conferring invasiveness might differ between species due to the different growth form. For example, submerged IAAPs might have to cope with reduced light availability (e.g. due to enhanced phytoplankton growth) or insufficient access to suitable forms of inorganic carbon for photosynthesis (Sand-Jensen, 1989), while emerged and free-floating IAAPs use air CO2 and thus usually do not face photosynthetic carbon limitation. In contrast, for sediment-rooted plants with floating or emerged leaves, the capacity to respond to reduced water availability is particularly important (Geng et al., 2006; Hussner, 2010; Wersal et al., 2013; Thouvenot et al., 2013a).
When separating the species according to their predominant growth form(s) (Table 1), we can see that certain traits are relevant for a specific growth form and will help explaining the invasiveness of an IAAP in different habitat types (see details below). In addition, the presented knowledge of species biology is crucial for pest risk assessments (Gordon et al., 2012) and the selection of the most appropriate management measure to achieve the highest management success (Hussner et al., 2016a). The question posed in Hussner et al. (2017) when an introduced aquatic plant species becomes invasive is answered here in this review in much more detail by the proposed information network on different plant traits.
Dispersal and spread
The propagule pressure is considered as one of the most important factors determining the invasive success of IAAPs (Thomaz et al., 2015; Fig. 1). Aquatic plants spread sexually (via seeds or spores) and asexually (via whole plants, plant fragments or duration organs, i.e. turions). There are strong differences in the relevance of sexual and asexual dispersal for the spread success, as the presence and number of produced seeds, spores or vegetative dispersal organs largely differ between the growth form and species (Table 3). For example, while seed production is widely reported for most free-floating and sediment-rooted, emerged or floating-leaved species, the majority of submerged IAAPs do not produce viable seeds within their introduced ranges (Table 3). In contrast, the number of vegetative propagules has been found to be much higher in submerged IAAPs than in sediment-rooted, emerged or floating-leaved IAAPs (Heidbüchel et al., 2016).

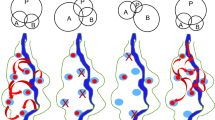
Schematic illustrations of the traits and the role in the invasion process
Vegetative plant parts, particularly stem fragments, are considered as the major pathway of spread of most IAAPs (Boedeltje et al., 2003). These fragments are produced either by autofragmentation or allofragmentation (Riis et al., 2009). Any kind of disturbance, for example control measure such as mechanical control, herbicide treatment or biological control, hydrology, wind and wave action, can strongly increase their number (Anderson, 1998; Dugdale et al., 2010).
The dispersal distance of plant fragments depends on both habitat and fragment characteristics (Heidbüchel et al., 2020), but fragment buoyancy in particular has been documented as a key parameter (Sarneel, 2013; Cornacchia et al., 2019). The retention of drifting fragments mostly occurred at deadwood and existing riparian and aquatic vegetation, but colonization (except for free-floating species) is limited to those sites where fragments are able to sink down to the sediment to anchor through root production (Bickel, 2017; Heidbüchel & Hussner, 2019; Heidbüchel et al., 2020). Moreover, the likelihood for regeneration and colonization of dispersed vegetative propagules when stranded on the shore differs between growth forms, where conditions are suitable for plant regrowth and regeneration. Most of the floating plant parts commonly strand at the shore and banks of water bodies, where emergent species in particular have a high likelihood for regeneration and colonization (Hussner, 2009; Thouvenot et al., 2013b).
The likelihood for colonization by vegetative propagules of submerged species seems less likely and depends on the buoyancy of the propagules. They need to sink down to the sediment at those sites, where light conditions are suitable for growth. However, surprisingly little is known about seasonal or diurnal changes or other influencing parameters for fragment buoyancy (but see e.g. Cattaneo & Kalff, 1980; Creed et al., 1992; 1995). The propagules of free-floating plant species are rarely limited by sites for their regrowth due to their growth form.
While no additional vectors are needed for dispersal within a water body, the spread into new unconnected water bodies relies on vectors for the spread, where water sport equipment (i.e. boats and trailers) is the most common (Bruckerhoff et al., 2015). Recreational boaters and fishermen may inadvertently transport the plant in trailers or boat hulls between water bodies, while subsistence fishermen and recreational anglers are known to move plants around to increase habitat for fish fry (Hill, 2003). The endozoochorous (documented only for very small floating Wolffia spp. so far; Silva et al., 2018) and ectozoochorous overland transport of vegetative plant parts (Agami & Weisel, 1986; Coughlan et al., 2015) via waterfowl seems possible, but rather unlikely, for example for the submerged species listed in Table 1, considering their rapid desiccation in air (Barnes et al., 2013; Heidbüchel et al., 2019b). Moreover, it will be usually limited to small pleustophytes, as the waterfowl-mediated dispersal of larger vegetative propagules is considered as rare. In contrast, viable seeds could be easily transported via waterfowl (see following sections on free-floating and on sediment-rooted plant species with emerged or floating leaves) (Reynolds et al., 2015). Seed production is rather uncommon in submerged IAAPs and has been found only for three out of the nine submerged species considered here (Table 3).
In addition to the general factors determining the capacity for propagule regeneration and establishment at new sites, the resistance and tolerance of the propagules to desiccation during overland transport are crucial, as these affect the fitness of the propagules at the time entering a new water body. Desiccation resistance strongly differs between growth forms, as in contrast to free-floating and emerged plants, submerged plants lack a cuticle, which reduces water loss (Sculthorpe, 1967; Heidbüchel et al., 2019b). Consequently, submerged plants have in general a lower desiccation resistance. However, the fitness of the propagules at the time of entrance determines the likelihood for establishment and regrowth, but has been studied in detail only for submerged IAAPs (Barnes et al., 2013; Bickel, 2015; Coughlan et al., 2018; Heidbüchel et al., 2019b).
Free-floating species
The production of viable seeds or spores of the free-floating E. crassipes, P. stratiotes and A. filiculoides is reported within their introduced ranges (Table 3; Janes, 1998a; Albano Pérez et al., 2011; Hussner et al., 2014b). Both seeds and spores are dispersal organs, but both will sink down to the sediment and thus their spread seems limited, although seeds of E. crassipes are considered hydrochorous, dispersed by rain wash, downstream flow and floods (Albano Pérez et al., 2011). Furthermore, seeds and spores can be transported into new water bodies by waterfowl and mammals when caught up in mud, illustrating the high potential for dispersal over large distances (Brochet et al., 2012; Garcia-Alvarez et al., 2015). In contrast to E. crassipes, A. filiculoides and P. stratiotes, spore production is lacking in S. molesta, a sterile pentaploid for which dispersal is thus limited to viable fragments (Julien et al., 2009) (Table 3).
Water flow is the main mode of natural dispersal of vegetative organs of free-floating IAAPs within a waterbody. Daughter plants break off from the parent plant and are moved downstream (Table 3). While wind moves the plants around systems too, creating large mats in the direction of the prevailing winds, birds and mammals may disperse daughter plants within and between water bodies even over larger distances (Coetzee et al., 2017).
Sediment-rooted, emerged or floating-leaved species
Most sediment-rooted, emerged or floating-leaved species from our list of IAAPs are known to disperse sexually and asexually (Table 3), but the asexual spread via plant fragments is predominant (Hussner, 2009; Schooler et al., 2012; Thouvenot et al., 2013b). Surprisingly, the documented fragmentation rates (the number of produced fragments per unit plant mass) were lower for H. ranunculoides, H. polysperma and M. aquaticum than for submerged growing species like M. spicatum or C. demersum (Heidbüchel et al., 2016), indicating that the fragmentation rate is much lower in sediment-rooted, emerged or floating-leaved than in submerged IAAPs. Nevertheless, the likelihood for the establishment of new infestations is higher than for submerged plant fragments, as the floating fragments often strand at the shore of water bodies, where new root formation might easily result in the anchorage of the fragments and the establishment of a new population. In conclusion, stem fragments of the rooted, emerged and floating-leaved IAAPs studied here have a high regeneration capacity (Table 3) and thus a high likelihood for establishment when stranded at suitable habitats, which will supersede the effect of relatively low fragmentation rates.
Submerged species
For submerged IAAPs, vegetative dispersal organs, i.e. stem fragments, play the major role for the spread within and into new water bodies (Boedeltje et al., 2003; Heidbüchel et al., 2016), even though seed production is documented for M. heterophyllum (reviewed in Gross et al., 2020), M. spicatum (Hartleb et al., 1993) and H. verticillata (Langeland & Smith, 1984; but see Sousa, 2011) within, in at least parts of, their introduced ranges (Table 3). Despite strong differences in the number of stem fragments produced by a species (Anderson, 1998; Redekop et al., 2016; Heidbüchel et al., 2016, 2019a; Heidbüchel & Hussner, 2020), the number of fragments produced strongly increases if management, especially mechanical management, of IAAPs is applied (Anderson et al., 1998).
In general, the likelihood for regeneration of stem fragments increases with fragment size (Redekop et al., 2016; Bickel, 2017; Heidbüchel et al., 2019a) and the presence of an apical bud (Riis et al., 2009; Umetsu et al., 2012; Heidbüchel & Hussner, 2019). Surprisingly, high desiccation tolerance was found for almost all of the studied submerged IAAPs, even though there were differences between the fragment type (e.g. with or without apical tips) (Barnes et al., 2013; Bickel, 2015; Coughlan et al., 2018; Xie et al., 2018; Heidbüchel & Hussner, 2019). This explains the high likelihood for overland transport of submerged plants with plant fragments. In addition to the most common stem fragments, some submerged IAAPs produce storage organs such as tubers or turions (Madsen & Smith, 1999, Hofstra et al., 1999), which act as additional dispersal organs.
Growth
In general, the invasiveness of alien plants has been linked to their maximum growth rate (Dawson et al., 2010), which allows IAAPs to outcompete slower growing species and to form large stands within a short period of time. Nevertheless, the growth rates of IAAPs differ strongly between habitats, depending on abiotic factors, such as nutrients, light, temperature and carbon availability for photosynthetic uptake (Reddy et al., 1990; Thouvenot et al., 2013b) and biotic factors (i.e. competition and herbivory). Surprisingly, for some IAAPs, comparatively low growth rates are reported (e.g. for C. helmsii and M. heterophyllum; Hussner, 2009; Hussner & Jahns, 2015), indicating that other traits can compensate for the slow growth and contribute to the invasive success. Reasonably, the seasonality of species determines the importance of fast growth rates, as strong seasonal species must form their invasive stands within a limited period of time, while evergreen species accumulate their biomass in the long term (Greulich & Bornette, 2003). In addition, the different growth forms of IAAPs colonize different zones within the aquatic habitat. While free-floating species are not strongly bound to a specific zone of the aquatic ecosystem, rooted, emerged and floating-leaved species usually grow in the transition zone between land and water and are thus often exposed to rapid and strong changes in environmental conditions (i.e. water-level fluctuations). Thus, the acclimation capacity or the phenotypic plasticity seems to be more relevant than for free-floating species (see section on phenotypic plasticity below).
However, the growth of IAAPs is determined by a number of growth traits, including the growth response capacity to varying environmental conditions (phenotypic plasticity) and the rapid vegetative and sexual reproduction (see separate sections). A few emergent IAAPs are known to grow much taller in the invaded habitat than in their native habitat (e.g. Lythrum salicaria (Chun et al., 2007), but little is known for the species investigated in our review, warranting more studies on the phenotypic plasticity of IAAPs between native and invaded habitats. Moreover, intraspecific and interspecific competition can strongly modify growth rates (Agami & Reddy, 1990). In the following, we provide a general overview about growth rates and preferred growth conditions.
Free-floating species
The availability of nitrogen and phosphorus in the water column is the primary factor determining the growth rate of free-floating IAAPs (Agami & Reddy, 1990; Henry-Silva et al., 2008), with eutrophic waters in particular offering favourable conditions for the invasion by free-floating IAAPs. When readily available, growth of floating macrophytes can increase unchecked because they absorb available nutrients through their root systems directly from the water column. In addition, E. crassipes, P. stratiotes and S. molesta are native to subtropical and tropical regions and thus profit from high air temperatures of 25 to 30°C (Owens & Madsen, 1995; McFarland et al., 2004; Henry-Silva et al., 2008), while A. filiculoides, native to warm temperate and subtropical regions, shows fastest growth at about 20°C; Janes, 1998a). Biomass doubling times of less than 1 week are reported for, e.g. E. crassipes and A. filiculoides (Edwards & Musil, 1975; Lumpkin & Plucknett, 1980; Gopal, 1987; Brouwer et al., 2018). However, doubling times or growth rates vary strongly, for example doubling times for biomass and plant number of E. crassipes are in the range of 5.9–28.1 and 3.7– 57.8 days, respectively (reviewed in Gopal, 1987). At very high densities, self-thinning (density declines and biomass increases) regulates E. crassipes density (Center & Spencer, 1981; Madsen, 1993). However, higher growth rates were found for P. stratiotes than for E. crassipes in monoculture experiments under central Florida climate (Reddy & DeBusk, 1983). In contrast, greenhouse experiments revealed similar (but much lower than in Reddy & DeBusk, 1983) growth rates for P. stratiotes and S. molesta than for E. crassipes, all being stimulated in growth by high nutrients (Henry-Silva et al., 2008), indicating that growth rates differ strongly due to differences in the growing conditions. Salvinia molesta is able to double the infestation size in about three days at optimal growth conditions (about 30°C and high nutrients; Room, 1992; EPPO, 2017a) and shows a high tolerance to environmental stress (Thomas & Room, 1986; Tipping, 2004). Its high growth rate is characterized by three distinct growth stages, the primary, secondary and tertiary phases, which are differentiated largely by the size of the leaves (Mitchell & Tur, 1975). The tertiary growth stage is the mat-forming stage and the only stage to bear sterile sporocarps (Mitchell & Tur, 1975).
Sediment-rooted, emerged or floating-leaved species
For the eight rooted, emerged or floating-leaved species considered here, similar maximum relative growth rates (RGRs) are reported, ranging from 0.06 to 0.18 g g−1 day−1 and resulting in doubling times under optimal growth conditions of about 1–2 weeks and RGR of 0.05–0.11 g g−1 day−1. The highest RGR > 0.1 g g−1 day−1 is reported for H. ranunculoides, C. helmsii and L. peploides (Rejmànková, 1992; Hussner, 2009), but RGRs between 0.025 and 0.07 are common for rooted, emerged or floating-leaved IAAPs (Doyle et al., 2003; Hussner, 2009; Thouvenot et al.; 2013a, b; Ayi et al., 2019). In general, the highest RGRs are found when plants are growing in shallow water or under waterlogged conditions and high nutrients (Geng et al., 2006; Hussner et al., 2009; Hussner & Meyer, 2009; Hussner, 2010; Wersal & Madsen, 2011; Thouvenot et al., 2013a, b), illustrating that eutrophic, shallow waters and wetlands in particular are prone to invasion by these emerged species. However, in the transition zone between land and water, changes in environmental conditions (i.e. water-level fluctuations) are common, and thus, plant growth and invasive success are strongly determined by the acclimation capacity of the species (see section on phenotypic plasticity and drought tolerance). All rooted, emerged or floating-leaved IAAPs considered here showed relatively high light optima and high temperature maxima of about 25 to 30°C, with lowest temperature and light optima for C. helmsii and H. polysperma and highest for A. philoxeroides and L. peploides (Hussner, 2009; Nault & Mikulyuk, 2009a; Thouvenot et al., 2013b; EPPO, 2016a).
Submerged species
The growth of the majority of submerged IAAPs is largely determined by nutrients, temperature, light and carbon availability (Barko & Smart, 1981; Sand-Jensen, 1989; Riis et al. 2012; Hussner & Jahns, 2015). The light availability within the water column is highly variable due to water depth, mineral and biotic (algae) turbidity, shading by vegetation (including self-shading) or epiphyton, and wave focussing (Hofmann et al., 2008; Pedersen et al., 2013). For plant growth, both optimum and minimum light requirements (determining the maximum colonization depths; Middelboe & Markager, 1997) are highly relevant (Sand-Jensen, 1989). Several studies reported the invasion of submerged IAAPs in oligotrophic, clear water lakes (Howard-Williams & Davies, 1988; Kozhova & Izhboldina, 1993; Wells et al., 1997), illustrating that invasion of submerged IAAPs can occur even under low nutrient conditions and thus relatively high light availabilities. Recent studies documented decreasing nitrogen requirements under increasing DIC availabilities (Dülger et al., 2017), which might explain the invasion success of submerged IAAPs (which usually require eutrophic conditions) in high alkaline but nutrient-poor volcanic lakes (Howard-Williams & Davies, 1988; Wells et al., 1997). However, for submerged IAAPs, light optima and compensation points seem to be generally low, but highly variable due to fast acclimation to overall differences in the light availability, as documented for submerged aquatic plant species (Barko & Smart 1981; Madsen & Sand-Jensen, 1994; Riis et al., 2012; Hussner et al., 2011, 2015). Not surprisingly, the light compensation point of submerged plants is influenced by the carbon availability and will decrease with increasing carbon availability due to a higher light use efficiency (Madsen & Sand-Jensen, 1994). The temperature optima of almost all submerged IAAPs are within the range of 20–30°C (Barko & Smart, 1981; Riis et al., 2012; Hussner et al., 2015; Hussner & Jahns, 2015), with L. major at the lower and H. verticillata at the upper end. The relatively high temperature optimum in H. verticillata might be linked to its single-cell C4-mechanism (Bowes, 2011). Overall, under optimal conditions, biomass doubling times of about 1–2 weeks (RGR of about 0.05–0.11 g g−1 day−1) for submerged IAAPs are reported (Nielsen & Sand-Jensen, 1991; James et al., 2006; Hussner et al., 2015, 2016b; Riis et al., 2012), but strong variations occur, largely due to variations in carbon affinity and carbon availability (Nielsen & Sand-Jensen, 1991; see also section on carbon-concentrating mechanisms).
Clonal growth and integration
Clonal growth can improve plant growth and competitive strength in both homogenous and heterogeneous environments (You et al., 2016b, Wang et al., 2016a, b). Because ramets are connected, clonal plants are able to share nutrients, carbohydrates and water between different habitat types, from terrestrial to aquatic (You et al., 2016b). Clonal growth and clonal integration are important traits for IAAPs (You et al. 2016b). Clonal integration allows sediment-rooted plants in particular to spread into unfavourable habitat conditions with limitation in light, nutrients or water, or to grow even in an unfavourable growth form (e.g. totally submerged) and to withstand periodic submergence (Wei et al., 2018). The donor and recipient ramets from different habitats might differ in plant traits (e.g. mass ratios of leaves, stems and roots; Wang et al., 2017), which must be borne in mind when studying the plant traits of IAAPs.
Clonal growth has been widely studied for rooted, emerged or floating-leaved and free-floating species (Alpert et al., 1991). Surprisingly, there is a lack of information on clonal integration in obligate submerged IAAP species considered in this study (Table 4), even though clonal integration in submerged plant species occurs (Xiao et al., 2007; Wolfer & Straile, 2012).
Free-floating species
Clonal integration is well documented for the large free-floating species, E. crassipes and P. stratiotes (Wang et al. 2014, 2016a; Yu et al., 2019). In E. crassipes, carbon transport between clonal integrated plants is documented (Alpert et al., 1991), but no measurements of internal transport between connected ramets for P. stratiotes appear to have been published. Using fragmentation as a means to prevent clonal integration, Wang et al. (2016a) demonstrated that the relative interspecific competitive abilities of these two species is increased if the daughter plants remain attached to the mother plant. Further, the relative competitive ability of P. stratiotes was reduced through fragmentation, but integration enabled the smaller P. stratiotes daughter plants to compete more effectively against the larger E. crassipes daughter plants. Lower rates of fragmentation may therefore select for production of more numerous, smaller ramets in clonal species.
Clonal integration has not been documented for A. filiculoides or S. molesta to date.
Sediment-rooted, emerged or floating-leaved species
For IAAPs growing in the transition zone between land and water, clonal integration seems of high relevance, and connection between terrestrial and aquatic ramets supports the growth of the species even under unfavourable conditions (Glover et al., 2015). In addition, clonal integration might help to withstand periodic changes in habitat conditions, e.g. from flooding to dry conditions (You et al., 2013). Numerous studies on A. philoxeroides, document the major role of clonal integration within this species (e.g. Wang et al., 2008; Xu et al., 2010; You et al., 2016b). Clonal integration is also reported for L. grandiflora (Glover et al., 2015), M. aquaticum (You et al., 2013) and for the genus Hydrocotyle (Wang et al. 2016b, 2017), and is thus most likely present in H. ranunculoides as well. No studies were found on clonal integration in T. natans and H. polysperma.
Seasonality and evergreen life cycle
There is some evidence that evergreen species show generally lower growth rates than seasonal species, but being evergreen can compensate for the slow growth during the main growing season (Greulich & Bornette, 1999, 2003; Fig. 1). Being evergreen allows IAAPs to colonize sites formerly dominated by native seasonal species and to displace them (Hussner, 2014), and thus, there is no need for evergreen IAAPs to outcompete seasonal species within their limited growth period. Moreover, the long-term biomass accumulation might explain why evergreen IAAPs are able to invade even unfavourable habitats, such as C. helmsii in nutrient-poor moorland pools (Brouwer et al., 2017), and thus expand the list of potential habitats suitable for invasion by the species. Nevertheless, the aspect of being evergreen for IAAPs has hardly been studied to date (Table 4).
It seems reasonable that changes in climatic warming extend the growing season of seasonal species in temperature-limited ecosystems (Menzel & Fabian, 1999). This might increase the resistance of native plant communities to invasion by evergreen IAAPs, but will also increase the growth rates of evergreen IAAPs during the winter time. Surprisingly, fewer studies investigated the effects of climate warming on the competition between seasonal and evergreen species (but see e.g. Hussner et al., 2014c). Nevertheless, some of the species considered here are obligate evergreen even during frost events, and when water surfaces are covered by ice (Table 4), while others may stay green as long as the water surface is not covered by ice and are thus considered evergreen in parts of their introduced ranges (e.g. E. nuttalli; Kadono, 2004). Consequently, the seasonality of a given species may differ within is distribution range, making a clear distinction between evergreen and seasonal plants difficult (e.g. in E. densa, reviewed in Yarrow et al., 2009). Moreover, almost all seasonal species considered here can be found in overwintering stages (vital fragments or small plants) during the winter time, from which regrowth occurs in spring. Moreover, in some cases, a species might be seasonal in their predominant growth form but evergreen in the other, e.g. M. aquaticum with seasonal emerged, but evergreen submerged, plants. We thus consider only those IAAPs as evergreen, if they stay evergreen even in frost events and/or under ice in their predominant growth form (Table 4).
Free-floating species
Because the most invasive free-floating species are tropical in origin, their growth period is largely determined by frost and ice (see section on frost tolerance below). If no frost and ice events occur, the species are able to grow the whole year round (Hussner et al., 2014b), even though the growth rates are determined by changing temperature and light conditions. Large free-floating E. crassipes and P. stratiotes are more exposed to cold air temperatures than small and rather flat A. filiculoides due to their higher growth above the water surface. Interestingly, a small flat winter form of P. stratiotes with floating leaves was found, which allows the plant to survive freezing air temperatures as long as the water surface is not covered by ice (Hussner et al., 2014b).
Sediment-rooted, emerged or floating-leaved species
The sediment-rooted, emerged or floating-leaved species considered here are all seasonal (Hummel & Kiviat, 2004; Hussner, 2008; Thouvenot et al., 2013b; EPPO, 2016a, 2017b), except C. helmsii, which remains green and vital as long as the plants are not covered for extended periods by snow (EPPO, 2007; Hussner, 2008). Nevertheless, in M. aquaticum, H. ranunculoides and Ludwigia spp., seasonality largely depends on the winter temperatures. While under mild winter conditions, plants may overwinter in a small creeping form (Hussner, 2008), the plants usually overwinter during frost and ice with their rhizome and root system, from which plant regrowth occurs in spring (Hussner, 2008; Thouvenot et al., 2013b). Moreover, at least for M. aquaticum and H. ranunculoides, small overwintering submerged forms are reported even under winter air frost conditions (Hussner, 2008).
Submerged species
In contrast to free-floating and rooted, emerged and floating-leaved species, submerged growing IAAPs are not exposed to freezing temperatures within the water column. Some submerged IAAPs have been reported as at least partly evergreen in their introduced ranges even though plant parts enclosed in ice near the water surface are killed, including C. caroliniana, E. densa, L. major and M. heterophyllum (Kadono, 2004; Wilson et al., 2007; Caffrey et al., 2011; Bickel & Schooler, 2015; EPPO, 2016b). For other species, like E. nuttallii, being evergreen has been reported in at least parts of their introduced ranges (Kadono, 2004). However, some plants remain in a dormant life form during winter (e.g. E. densa, Matthews et al., 2014) or form dormant apices (e.g. E. canadensis; Janauer, 1981). Because seed production is rather uncommon in submerged IAAPs, plant regrowth of seasonal species must rely on overwintering organs, such as turions (e.g. in C. caroliana, Wilson et al., 2007; H. verticillata, reviewed in Sousa, 2011; and M. heterophyllum, even though this has not been reported for this species within its introduced range in Europe yet; Gross et al., 2020), or rhizomes and roots.
Phenotypic plasticity
Phenotypic plasticity is the variability of the phenotype expressed by a single genotype, which is a plant response to changing abiotic and biotic conditions (Gratani, 2014). In IAAPs, phenotypic plasticity has been reported particularly as a response to differing and changing habitat conditions (Riis et al., 2010). In general, plant populations with an optimal plastic response show higher growth rates than non-plastic species; still, growth rates are not higher than for plant species with different phenotypes, which are perfectly adjusted to the environment at all times (Xue & Leibler, 2018). IAAPs with a high phenotypic plasticity may provide a competitive advantage over (native) species with a lower plasticity (Geng et al., 2006). In addition, the ability to respond to varying habitat conditions allows species to grow and survive under unfavourable conditions and/or habitats (Fig. 1). Consequently, phenotypic plasticity seems to be an important trait of IAAPs (Table 4), increasing the number of potential habitats and enhancing the competitive strength of a species.
Free-floating species
The morphology of free-floating invasive macrophytes tends to be plastic particularly in response to crowding and/or nutrient conditions. For example, the basal rosette leaves of E. crassipes have elongated petioles (up to 1 m) and circular leaves in dense stands, while in sparse infestations the plants have short (< 30 cm) and bulbous petioles and kidney-shaped leaves (Center & Spencer, 1981), resulting in a higher weight per plant in high densities (Agami & Reddy, 1990). This more erect and taller growth in dense stands allows E. crassipes to overgrow other free-floating species, including P. stratiotes (Agami & Reddy, 1990).
In addition, growth plasticity occurs within the root system, for example, the rhizome and the feathery roots of E. crassipes are longer and denser at low nutrients (particularly phosphorus) than at high nutrients (Xie & Yu, 2003). Moreover, the root–shoot ratio varies inversely with nutrient, particularly nitrogen, availability.
Similarly, S. molesta demonstrates considerable phenotypic plasticity in response to crowding (Julien et al., 2009), with the primary form having small leaves, typical of plants invading open water, the secondary form having large, slightly folded leaves, and the tertiary form having large, deeply folded and densely packed leaves, typical of mature stands (Mitchell & Tur, 1975). These different morphological forms may all occur within a mat, as a response to local conditions in space and time, showing a high dynamic in phenotypic plasticity (Julien et al., 2009).
In addition to crowding and nutrients, temperature may cause changes in plant phenology. Freezing air temperatures cause reduced leaf size and changes in leaf position in P. stratiotes, when plants form a small floating-leaved winter form to protect the leaves from freezing air temperatures (Hussner et al., 2014b). In A. filiculoides, a very small and dark red form was found at temperatures around 0°C, and this red colour, caused by anthocyanin, is also known as a response to warm temperatures and high light conditions (Janes, 1998a). But in comparison to E. crassipes, P. stratiotes and S. molesta, there is only a low growth plasticity in A. filiculoides.
Sediment-rooted, emerged or floating-leaved species
Along the shoreline of lakes and rivers, rooted species are adapted to changing water levels and usually show phenotypical acclimations to both submergence and emergence. Decreasing water levels and water limitations lead to reduced erect growth, shorter internodes and petioles and smaller leaves and specific leaf area (SLA), but higher leaf dry matter contents (LDMC) and larger root systems (Geng et al., 2006; Fast et al., 2008; Hussner & Meyer, 2009; Hussner et al., 2009; Hussner, 2010; Thouvenot et al., 2013a). In contrast, plant submergence causes a reduced leaf thickness and the lack of a cuticle in M. aquaticum and A. philoxeroides (Salvucci & Bowes, 1982). Similarly, in H. ranunculoides, A. philoxeroides and L. grandiflora, the leaves get softer under submergence, indicated by a lower leaf dry matter content or increased specific leaf area (Botts et al., 1990; Hussner, 2008; Fan et al., 2015).
Similar growth responses are documented for Ludwigia spp. and M. aquaticum under varying nutrient availability, when plants showed reduced erect growth but larger root systems at low nutrient levels (Hussner et al., 2009; Hussner, 2010).
Submerged species
For submerged IAAPs such as E. canadensis, E. densa and L. major, strong phenotypic variations occur between plants from different water bodies, which largely disappear after growth under similar conditions (Riis et al., 2012). Such strong phenotypic plasticity has been reported as a response to varying nutrients, light, temperature and DIC and/or CO2 availability for all submerged species considered here (e.g. Barko & Smart, 1981; Sousa, 2011; Riis et al., 2012; Eusebio Malheiro et al., 2013). Light limitation causes more erect growth with longer internodes and less branching (Barko & Smart, 1981; Riis et al., 2012), but also high plant density stimulates shoot elongation. Increasing leaf length and shorter internodes were observed in submerged M. aquaticum under CO2 limitation, when plants increased the leaf surface to improve the diffusional carbon uptake, which resulted in decreasing leaf dry matter contents (LDMC, Eusebio Malheiro et al., 2013). The biomass allocation to roots is largely driven by the nutrient, light and DIC/CO2 availability than by temperature (Hussner et al., 2015; Hussner & Jahns, 2015). Nutrient limitation and high carbon availability, with roots acting as a sink for the produced starch (Dülger et al., 2017; Dülger & Hussner, 2017), will cause increased biomass allocation to roots and thus increasing root: shoot ratios (Hussner et al., 2015; Hussner & Jahns, 2015).
Allelopathy, allelochemical reactions and herbivore defences
Allelopathy is a means of interference competition between different plant species, linked to the release of active compounds by the producing organism and subsequent effects on target species. The inhibition of other competing primary producers, specifically phytoplankton but also epiphytic algae and microorganisms in general, allows a better exploitation of resources by the producing species (Gross, 2003). A wider definition talks about allelochemical interactions, including the production of feeding deterrents or inhibitors against herbivores (Gross, 2009).
Although quite a few studies investigate biological control by herbivores or pathogens of IAAPs (Newman, 1991; Gross & Bakker, 2012), not much is known about possible herbivore deterrents. Such allelochemicals might be active against generalist herbivores but fail to control specialists (see, e.g. Gross & Bakker, 2012).
A few more studies report new secondary metabolites, sometimes with antibacterial or antifungal compounds in the listed macrophytes (Della Greca et al., 1991, 1994; Smida et al., 2015). It is unclear if allelochemical interference against herbivores and pathogens accounts for the invasion success of IAAPs. Given that many of these invaders seem to suffer less from pathogens or herbivores (sensu the “Enemy Release Hypothesis”) (Keane & Crawley, 2002), it might be possible that these IAAPs are not only lacking their natural enemies but are also better defended than native plants. A similar direction takes the “Novel Weapons Hypothesis” (NWH), which suggests that alien invasive plants possess a higher allelopathic activity in the new habitat (Bais et al., 2003). While proof for such patterns is heavily debated for the few terrestrial case studies (Blair et al., 2006; Bais, 2010), equal evidence is, to our knowledge, still lacking for aquatic alien invasive plants and thus warrants further investigation. Some studies have compared native and invasive alien aquatic plants for their allelopathic or allelochemical potential. No difference in epiphyte density was observed between native and non-native plants (Grutters et al., 2017b). Growth form, but not origin (native or non-native) explained the allelopathic potential of aquatic plants, which was strongly linked to the content of phenolic compounds and a high C:P ratio (Grutters et al., 2017c). Non-native plants exhibited no higher defence against larvae of an herbivorous aquatic moth (Grutters et al., 2016). Other studies using crayfish or snails found diverging results on the preference of non-native macrophytes: crayfish preferred exotic plants (Parker & Hay, 2005) while an aquatic snail preferred native plants (Xiong et al., 2008) when each was offered a pair of similar native and non-native macrophytes.
Free-floating species
Two of the four species show allelopathic activity against algae or cyanobacteria. Eichhornia crassipes and P. stratiotes contain antialgal and/or anticyanobacterial secondary metabolites (Aliotta et al., 1991; Della Greca et al., 1991, 1992, 1998; Kato-Noguchi et al., 2014; Wu et al., 2013, 2015, 2019). No reports on allelopathic activity are available for S. molesta and A. filiculoides (Table 4).
The same pattern holds for herbivore defences. They have been observed in E. crassipes and P. stratiotes, but not in A. filiculoides and S. molesta. Different species of the genus Neochetina induce resistance against herbivory in E. crassipes (Buchanan, 2013). Low-nutrient plants of P. stratiotes have a higher tissue toughness, negatively affecting potential biocontrol agents (Wheeler et al., 1998).
Sediment-rooted, emerged or floating-leaved species
Myriophyllum aquaticum produces allelopathically active compounds against cyanobacteria and/or eukaryotic algae (Saito et al., 1989). A few reports on interference competition or allelopathy exist for H. polysperma, L. grandiflora (hexapetala) and L. peploides (Dandelot et al., 2008; Doyle et al., 2003). Many different studies have been performed with A. philoxeroides, outlining strong allelopathic and antibacterial activity, the isolation of active compounds and the effect of environmental conditions on the production of active compounds (Zuo et al., 2012a; b; Kleinowski et al., 2016; Huang et al., 2017; Ge et al., 2018; Xiao et al., 2019).
Potential herbivore defence has been described for A. philoxeroides against crayfish, apple snail and native insect herbivores (Cronin et al., 2002; Wong et al., 2010; Dai et al., 2014). Phenolic compounds present in M. aquaticum are considered to reduce snail herbivory (Qiu & Kwong, 2009; Wong et al., 2010). No reports on allelopathic or allelochemical interactions are published to our knowledge for H. ranunculoides and T. natans.
Submerged species
Hydrocharitaceae and Haloragaceae contribute the most species to submerged IAAPs. Probably, the best investigated species among them is M. spicatum, known to produce allelochemically active polyphenols, inhibiting algae and cyanobacteria (Gross et al., 1996; Leu et al., 2002) and affecting feeding by larvae of the aquatic moth Acentria ephemerella (Choi et al., 2002; Walenciak et al., 2002; Fornoff & Gross, 2014). Although no studies to date have shown allelopathic effects or herbivore deterrents in M. heterophyllum (Table 4), such effects are very likely given similar contents in bioactive polyphenols (Choi et al., 2002).
Elodea species (E. canadensis and E. nuttallii) are also well known to contain allelopathically active compounds (Erhard & Gross, 2006). The same can possibly been deduced for the multiple members from the Hydrocharitaceae, such as E. densa, H. verticillata and L. major, as they most likely all contain similar phenolic compounds or other secondary metabolites than those found in E. nuttallii. Among them are flavonoids, which are presumably involved in allelopathic effects (Erhard & Gross, 2006) and act as herbivore deterrents (Erhard et al., 2007). A range of species, among them C. caroliniana, H. verticillata and M. aquaticum, inhibited the germination of lettuce or the growth of Lemna minor (Elakovich, 1989). C. caroliniana showed moderate to low inhibition of cyanobacterial growth compared with, e.g. M. spicatum in coexistence experiments (Nakai et al., 1999). Relatively high contents in alkaloids, potential herbivore deterrents, have been found in C. caroliniana, C. demersum, E. canadensis and M. spicatum (Ostrofsky & Zettler, 1986). No publications on allelopathy or herbivore deterrents have been found for C. helmsii.
Drought tolerance and avoidance strategies
IAAPs face significant challenges to survival during periods of water loss, such as seasonal drought or other periods of water drawdowns (Barrat-Segretain & Cellot, 2007; Barnes et al., 2013). They also experience desiccation during dispersal between water bodies as they travel as hitchhikers on terrestrial or semiaquatic organisms (Figuerola & Green, 2002) or recreational boats (Johnson et al. 2001) (for more information on desiccation tolerance and its role for dispersal see section above). However, several strategies allow aquatic plants to withstand temporary water drawdowns (Arthaud et al., 2012). Some species are able to tolerate the drawdown event, with or without modification of the plant growth form, e.g. leaf plasticity, a decrease in size, or tissue modifications to cuticles, stomata, aerenchyma and lignin (Wells & Pigliucci, 2000); or some persist as stunted forms without any specific developmental strategy, enabling them to survive but not to grow. For others, aboveground biomass dies, and the species remains dormant in the sediment as vegetative propagules (Barrat-Segretain, 2001), or through extended root growth and acclimation to reduce photosynthetic water loss (Hussner et al., 2009; Hussner & Meyer, 2009; Hussner, 2010). Some also flower during the drought event and die following seed production, producing a dormant seed bank that establishes following the return of inundation (Casanova & Brock, 2000; Brock et al., 2003).
Aquatic plant species rely on significant periods of inundation in order to survive and reproduce, so a reduction in water availability during periods of water loss is one of the most drastic changes that may affect invasive aquatic species because it represents a major habitat shift from aquatic to terrestrial environment (Madsen & Sand-Jensen, 1991). Increased evapotranspiration and rapid leaf desiccation, which prevents gas exchange, occur soon after water drawdown because most aquatic species have thin leaves with reduced cuticles and stomata. In addition, particularly submerged plants are unable to support themselves because their tissues do not have supportive structures comparable to terrestrial species (Wells & Pigliucci, 2000; Rascio, 2002). Apart from the obvious desiccation challenges, solar radiation and gravitational forces encountered by the plants increase substantially, and access to nutrients is modified (Rattray et al., 1991; Baldwin & Mitchell, 2000; Rascio, 2002). On the other hand, water drawdown increases light and CO2 availability in the transition zone between land and water, potentially enhancing photosynthesis of especially homophyllous riparian plant species (Sand-Jensen & Frost-Christensen 1999). In response to water drawdowns, species with adaptations that include the development of self-supporting aboveground organs with higher dry matter content enabling plants to withstand gravity, and smaller leaves with thicker cuticle to reduce evapotranspiration, leading to lower specific leaf area, higher leaf-construction costs and consequently higher leaf life span are more likely to survive (De Wilde et al., 2014).
Free-floating species
The world’s worst invasive free-floating macrophytes are native to the Amazon Basin, where desiccation and flooding are regular occurrences, as in most regions of the world. These IAAPs are tolerant to drought once water has receded, leaving the plants exposed on mud, and, because they are free-floating and mobile, are capable of surviving and flourishing on variable water levels. Germination occurs when substrates are exposed as water recedes, and also as dry substrates are moistened when water levels rise. Seeds and spores also survive in wet mud and are long-lived, and flowers of E. crassipes and P. stratiotes can be produced within 10–15 weeks after germination (Center & Spencer, 1981; Pieterse et al., 1981). Germination of E. crassipes seeds in sediments is prevented if the sediments are shaded or light levels and temperatures are low. In habitats where the water is shallow with a rooting medium suitable for initial seedling development, propagation by seeds may be crucial for the invasion of new areas (Edwards & Musil, 1975). Azolla filiculoides, on the other hand, reproduce sexually via spores in spring and summer, which overwinter and are resistant to extreme desiccation, thus enabling this fern to re-establish after any drought period (Hill & McConnachie, 2009).
While these floating species are capable of surviving desiccation during drought by sexual reproduction, they also have tolerance strategies enabling whole plants to survive water drawdowns. Venter et al. (2017) investigated responses of E. crassipes to periodic drought in controlled experiments, demonstrating that rooted plants (drought-stressed treatment) exhibited decreased leaf size (surface area) and increased root: shoot ratios, both interpreted as morphological adaptations to drought. By altering these morphological structures in response to being rooted, albeit in saturated-soil, E. crassipes probably reduced its overall plant water demands (Touchette et al., 2007). Furthermore, drought-stressed plants primarily exhibited a drought avoidance strategy whereby they continued photosynthesising during the drought by altering their physiology and morphology. By reducing stomatal conductance and subsequent transpiration, drought-stressed plants significantly increased their intrinsic water use efficiency (Venter et al., 2017).
Mats are produced during the dry season by S. molesta occuring in naturally systems with seasonal flooding. Those mats are flushed downstream, often until the sea, where salinity kills the plants, or they are deposited on floodplains, where they desiccate and die.
However, individual plants may survive through a dry season under a mulch of dead S. molesta plants, especially in lower, moist areas (Storrs & Julien, 1996). Neither P. stratiotes nor A. filiculoides have demonstrated mechanisms for whole plants withstanding desiccation during times of drought or water drawdown.
Sediment-rooted, emerged or floating-leaved species
These IAAPs are generally characterized by a high tolerance to water-level drawdowns and drought events (Table 4). Most species produce viable seeds in their introduced ranges, which withstand even prolonged drought periods in the sediment (Rector et al., 2015), and if the seeds are frost tolerant, even during the winter months. In addition, the species show strong growth responses to water-level fluctuations and drought (see section on phenotypic plasticity). The formation of large root systems and the reduction of biomass allocated to leaves are common growth responses to reduced water availability and allow the species, such as A. philoxeroides, Ludwigia spp., M. aquaticum and H. ranunculoides, to grow even under dry conditions (Geng et al., 2006; Hussner et al., 2009; Hussner & Meyer, 2009; Hussner, 2010; Thouvenot et al., 2013a). The responses of the root systems are lower in low-nutrient sediments (Hussner et al., 2009; Hussner, 2010), indicating that the drought tolerance might be lower in these sediments.
In contrast, C. helmsii does not enlarge the root system under dry conditions, but the plants have shorter internodes and more succulent leaves (Prinz et al., 2019). Crassula helmsii was found to grow even on dry and sandy sediments in a very small, creeping form (pers. obs. A.H. on the island of Norderney, Germany), documenting its high tolerance to drought.
Submerged species
In contrast to free-floating and sediment-rooted emerged or floating-leaved species, obligate submerged plants lack anatomical structures as adaptations to drought (e.g. cuticle). Submerged aquatic plants are strongly affected by water-level drawdowns and drought, and thus artificial water-level drawdowns have been used as a management tool for submerged IAAPs. But nevertheless, these management measured did not generally lead to the die-off of the target species (Hussner et al., 2017), as submerged IAAPs can withstand even a prolonged period of water-level drawdowns. Submerged biomass is clumped together and the amount of stranded vegetation mass influences the desiccation rate: if the top layer of plant material is thick enough to provide insulation to the plants underneath them (Clayton, 1996; Barrat-Segretain & Cellot, 2007), plants regrow through fragments that survive once the water level rises, as it has been documented for C. carolinaina, E. densa and Elodea spp. (Segretain & Cellot, 2007; Dugdale et al., 2012, 2013).
In addition, submerged IAAPs can survive drought events through different mechanisms. Viable seed formation can lead to seed banks, which are rarely killed by drawdown events, but seed formation is rare in most submerged IAAPs (but see Table 3). Moreover, terrestrial forms are reported for M. spicatum and M. heterophyllum (Hegi, 1975; Gross et al., 2020), which can consequently act as surviving strategy during drought events.
Perennation by rhizomes, tubers or other vegetative propagules submerged in the substrate is also a factor which can bring about rapid recovery of a population of submerged plants that may appear to have been totally destroyed after exposure to the air. Reproductive organs such as tubers and turions, characterized by high starch accumulation (Adamec et al., 2020) and high resistance to desiccation (Glisson et al., 2020), withstand drawdown events in the hydrosoil (Poovey & Kay, 1998). Turion formation is documented for, e.g. H. verticillata (Poovey & Kay, 1998) and controversial for M. heterophyllum (Gross et al., 2020), but also other forms such as turion-like stem apices in C. caroliniana (Wilson et al., 2007), or dormant apices in E. canadensis (Janauer, 1981). These act as duration organs and have been found in large numbers (several million subterranean turions of H. verticillata per hectare) within the sediment (Netherland, 1997). These allow reinfestation of a site after a drought and may remain quiescent in undisturbed sediment for up to 4 years (Netherland, 1997).
Frost tolerance
It has been widely assumed that climate change is a major driver of plant invasion, as it allows species to expand their range in formerly unsuitable regions (Hellmann et al., 2008). Recent studies document that the winter water temperatures in particular affect the establishment and invasive success of introduced alien aquatic plant species (Šajna et al., 2007; Hussner et al., 2014b, c). Consequently, IAAPs showing a tolerance to cooler temperatures or even frost and ice are more likely to expand their invasive range and establish. However, this tolerance will have a stronger effect on free-floating than on submerged IAAPs, as indicated by the different effects of increasing temperatures on alien aquatic plants with different growth forms (Netten et al., 2011). While single air frost events might have strong effects on free-floating and rooted, emerged and floating-leaved species, submerged plants will be reasonably less affected, as the water column provides a strong temperature buffer, but any ice enclosure of submerged plants leads to the die-off of the plant parts enclosed in ice. Consequently, we only consider the frost tolerance of free-floating and sediment-rooted, emerged and floating-leaved IAAPs.
Free-floating species
Free-floating IAAPs will generally die when enclosed in ice, but even air frost events affect plant growth. However, the effect of air frost will depend on the erect growth, as flat growth forms along the water surface provide protection, for example, the growth response of the usually more erect growing P. stratiotes to air frost (Hussner et al., 2014b). Nevertheless, of the four free-floating IAAPs considered here, E. crassipes showed the lowest tolerance to cold temperatures (Table 4), but withstood near-freezing temperature (< 5°C) for a limited period of time, and rooted plants are more resistant than free-floating plants (Owens & Madsen, 1995).
Salvinia molesta can withstand short periods (up to 48 h) of air frost of − 3°C, but plants are killed when cold temperatures persist (Owens et al., 2004; EPPO, 2017a). Pistia stratiotes shows a strong growth response to freezing air temperature. While its leaves above the water surface are killed by air frost, the plants survive in a small flat form with floating leaves even at air temperatures of − 5°C (Hussner et al., 2014b). Azolla filiculoides show the highest frost tolerance to air temperatures down to − 15°C, but plant parts above the water surface are killed as well as plants enclosed in ice (Janes, 1998a). Beside the frost effects on vegetative plant parts, seeds of E. crassipes and P. stratiotes and spores of A. filiculoides might allow plant regrowth in spring, as the seeds or spores have a higher resistance to freezing temperatures and are usually less exposed to cold temperatures when located at the sediment (Pieterse et al., 1981; Janes, 1998b).
Sediment-rooted, emerged or floating-leaved species
The eight sediment-rooted, emerged or floating-leaved IAAPs from our list do not exhibit frost tolerance (Table 4), and plant parts exposed to freezing air temperatures are usually killed, except for C. helmsii, which survives even coverage by snow (Hussner, 2008; Nault & Mikulyuk, 2009a; Thouvenot et al., 2013b; Smith & Buckley, 2020). But plants may overwinter with submerged plant forms or plant parts (e.g. M. aquaticum, Hussner, 2008; Ludwigia spp., Thouvenot et al., 2013b). In most cases, plant regrowth occurs due to persisting roots and rhizomes or produced seeds (Table 3).
Carbon-concentrating mechanism (CCM)
While free-floating and rooted, emerged or floating-leaved IAAPs are able to take up their carbon for primary production from the air, submerged plants must use the dissolved inorganic carbon within the water column or from the sediment (Winkel & Borum, 2009; Pedersen et al., 2013). Due to the lower diffusion rate of CO2 in water than in air, strong boundary effects and the fact that the portion of CO2 within the DIC pool in the water column is pH dependent (with HCO3− being the dominant DIC form in water with a pH of 7–10), DIC limitation for photosynthesis and growth might occur (Pedersen et al., 2013). To cope with limitations in CO2 and DIC, numerous submerged plant species develop mechanisms to use HCO3− as an additional carbon source (Maberly & Madsen, 2002; Yin et al., 2017). Some prior studies suggest that the HCO3− use capacity in submerged plants will increase the competitive strength under CO2 or DIC limitation and that this can be an important trait of submerged IAAPs (Spencer & Bowes, 1990; Bowes, 2011), allowing these species to outcompete (native) submerged species lacking this HCO3−- use capacity (Hussner et al., 2015). In fact, this capacity is documented for all submerged IAAPs species considered here (Table 4), even though it varies due to different underlying mechanisms (or differences in the efficiency within) between the species (see, e.g. Bowes, 2011; Hussner et al., 2016b; Table 4), even within a single genus. For Myriophyllum spp. it is reported that the HCO3− capacity differs strongly between species and clearly separates the two IAAPs, M. spicatum and, to a lesser extent, the evergreen M. heterophyllum, from non-invasive species within the genus, such as Myriophyllum verticillatum (Dülger & Hussner, 2017). In addition to obligate submerged IAAPs, CCMs are reported from submerged forms of some predominantly emerged growing IAAPs (i.e. C. helmsii: Klavsen & Maberly, 2009; M. aquaticum: Bowes, 2011; Eusebio Malheiro et al., 2013).
Nevertheless, it must be borne in mind that the relevance of the HCO3− use capacity for plant invasiveness is restricted to DIC or CO2-limited conditions. In CO2-rich waters, even obligate CO2 users, such as M. verticillatum (Caffrey & Monahan, 2006) can show nuisance growth.
Conclusions
IAAPs cause serious ecological and economic impact and are a major driver of changes in aquatic plant communities. The collected data show that differences in traits of IAAPs occur and that particularly the growth form determines the traits, which are of major relevance for a specific IAAP. The detailed knowledge on specific traits conferring a potential advantage to IAAPs is of major concern for several reasons: (i) Species traits could help identifying further potential IAAPs according to species traits considered here, which could then be used in Pest Risk Assessments (Gordon et al. 2012); (ii) Specific traits are linked to certain habitat preferences. A better knowledge on the link between such physiological, anatomical or morphological traits will help identifying habitats which are prone to the invasion of a given IAAP; and (iii) Knowing the specific traits of an IAAP will allow developing species-specific management plans and prevention measures (Hussner et al., 2017).
For the identification of potential further IAAPs, species biology should be assessed by studying the invasion history and potential changes in environmental tolerance traits (Gordon et al., 2012). The exhaustive compilation of different morphological, anatomical, biochemical and physiological traits described here provide a base to prepare and improve weed assessments following existing schemes (Champion & Clayton, 2001) as they would help assigning a score of invasiveness based on the combination of certain plant traits and potentially invaded habitats.
It is evident that certain traits are strongly linked to specific habitats that can be invaded, e.g. the growth form of IAAPs (Table 2) and physiological traits (e.g. CCM).
Finally, prevention and management of IAAPs require comprehensive knowledge on IAAPs biology (Hussner et al., 2017). For the development of prevention measures, the identification of pathways of spread is of major interest. The reviewed data on traits linked to the dispersal and spread of IAAPs illustrate that artificial traps for floating plant fragments would provide a relatively simple prevention measure to limit the flow-induced spread of IAAPs by fragments, particularly in river ecosystems. In contrast, measures to prevent the overland dispersal via boats and trailers will be largely based on the tolerance of plant fragments to desiccation (Barnes et al., 2013; Heidbüchel et al., 2019b) or any treatment methods (Hussner, 2019). The knowledge on IAAPs characteristics can be also used to develop site- and habitat-specific prevention and management measures. For example, native evergreen species can help to increase the biotic resistance of a habitat to the invasion of an evergreen IAAP (van der Loop et al., 2020). The review of IAAP traits presented in this study should therefore be a first comprehensive base highlighting and targeting the traits of those IAAPs that are likely to invade an aquatic habitat, based on both species-specific and site-specific characteristics, allowing policy-makers and managers to improve prevention and control measures.
References
Adamec, L., A. Kučerová & S. Janeček, 2020. Mineral nutrients, photosynthetic pigments and storage carbohydrates in turions of 21 aquatic plant species. Aquatic Botany 165: 103238.
Agami, M. & Y. Weisel, 1986. The role of mallard ducks (Anas platyrhynchos) in distribution and germination of seeds of the submerged hydrophyte Najas marina L. Oecologia 68: 473–475.
Agami, M. & K. R. Reddy, 1990. Competition for space between Eichhornia crassipes (Mart.) Solms and Pistia stratiotes L. cultured in nutrient-enriched water. Aquatic Botany 38: 195–208.
Aiken, S. G., P. R. Newroth & I. Wile, 1979. The biology of Canadian weeds. 34. Myriophyllum spicatum L. Canadian Journal of Plant Science 59: 201–215.
Albano Pérez, E., J. A. Coetzee, T. Ruiz-Téllez & M. P. Hill, 2011. A first report of water hyacinth (Eichhornia crassipes) soil seed banks in South Africa. South African Journal of Botany 77: 795–800.
Aliotta, G., P. Monaco, G. Pinto, A. Pollio & L. Previtera, 1991. Potential allelochemicals from Pistia stratiotes L. Journal of Chemical Ecology 17: 2223–2234.
Alpert, P., F. R. Warembourg & J. Roy, 1991. Transport of carbon among connected ramets of Eichhornia crassipes (Pontederiaceae) at normal and high levels of CO2. American Journal of Botany 78: 1459–1466.
Anderson, L.W.J., 1998. Dissipation and movement of Sonar and Komeen following typical applications for control of Egeria densa in the Sacramento–San Joaquin Delta and Production and viability of E. densa fragments following mechanical harvesting (1997/1998). Davis, CA U.S. Department of Agriculture–Agricultural Research Service, Environmental Impact Report for California Department of Boating and Waterways. 79.
Arthaud, F., D. Vallod, J. Robin & G. Bornette, 2012. Eutrophication and drought disturbance shape functional diversity and life-history traits of aquatic plants in shallow lakes. Aquatic Sciences 74: 471–481.
Ayi, Q., B. Zeng, K. Yang, F. Lin, X. Zhang, P. van Bogedom & J. H. C. Cornelissen, 2019. Similar growth performance but contrasting biomass allocation of root-flooded terrestrial plant Alternanthera philoxeroides (Mart.) Griseb. in response to nutrient versus dissolved oxygen stress. Frontiers in Plant Science 10: 111.
Azan, S., M. Bardecki & A. E. Laursen, 2015. Invasive aquatic plants in the aquarium and ornamental pond industries: a risk assessment for southern Ontario (Canada). Weed Research 55: 249–259.
Bais, H. P., 2010. Correction: allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 327: 781.
Bais, H. P., R. Vepachedu, S. Gilroy, R. M. Callaway & J. M. Vivanco, 2003. Allelopathy and exotic plant invasion: from molecules and genes to species interactions. Science 301: 1377–1380.
Baldwin, D. S. & A. M. Mitchell, 2000. The effects of drying and re-flooding on the sediment and soil nutrient dynamics of lowland river-floodplain systems. River Research and Applications 16: 457–467.
Baniszewski, J., J. P. Cuda, S. A. Gezan, S. Sharma & E. N. I. Weeks, 2016. Stem fragment regrowth of Hydrilla verticillata following desiccation. Journal of Aquatic Plant Management 54: 53–60.
Barko, J. W. & R. M. Smart, 1981. Comparative influences of light and temperature on the growth and metabolism of selected submersed freshwater macrophytes. Ecological Monographs 51: 219–235.
Barnes, M. A., C. L. Jerde, D. Keller, W. L. Chadderton, J. G. Howeth & D. M. Lodge, 2013. Viability of aquatic plant fragments following desiccation. Invasive Plant Science and Management 6: 320–325.
Barrat-Segretain, M. H., 2001. Biomass allocation in three macrophyte species in relation to the disturbance level of their habitat. Freshwater Biology 46: 935–945.
Barrat-Segretain, M. H. & B. Cellot, 2007. Response of invasive macrophyte species to drawdown: the case of Elodea sp. Aquatic Botany 87: 255–261.
Bickel, T. O., 2012. Ecology of the submersed aquatic weed Cabomba caroliniana in Australia. In: Eldershaw, V. (ed.), Eighteenth Australasian Weeds Conference, Melbourne, 8–11 October 2012. Weed Society of Victoria: 21–24.
Bickel, T. O., 2015. A boat hitchhiker’s guide to survival: Cabomba caroliniana desiccation resistance and survival ability. Hydrobiologia 746: 123–134.
Bickel, T. O., 2017. Processes and factors that affect regeneration and establishment of the invasive aquatic plant Cabomba caroliniana. Hydrobiologia 788: 157–168.
Bickel, T. O. & S. S. Schooler, 2015. Effect of water quality and season on the population dynamics of Cabomba caroliniana in subtropical Queensland, Australia. Aquatic Botany 123: 64–71.
Blair, A. C., S. J. Nissen, G. R. Brunk & R. A. Hufbauer, 2006. A lack of evidence for an ecological role of the putative allelochemical (±)-catechin in spotted knapweed invasion success. Journal of Chemical Ecology 32: 2327–2331.
Boedeltje, G., J. P. Bakker, R. M. Bekker, J. M. Van Groenendael & M. Soesbergen, 2003. Plant dispersal in a lowland stream in relation to occurrence and three specific life-history traits of the species in the species pool. Journal of Ecology 91: 855–866.
Botts, P. S., J. M. Lawrence, B. W. Witz & C. W. Kovach, 1990. Plasticity in Morphology, proximate composition, and energy content of Hygrophila polysperma (Roxb.) Anders. Aquatic Botany 36: 207–214.
Bowes, G., 2011. Single-cell C4 photosynthesis in aquatic plants. In: Photosynthesis and related CO2 concentrating mechanisms In: A.S. Rhagavendra & R.F. Sage (eds), pp. 63–80.
Bowmer, K. H., S. W. L. Jacobs & G. R. Sainty, 1995. Identification, biology and management of Elodea canadensis, hydrocharitaceae. Journal of Aquatic Plant Management 33: 13–19.
Brochet, A. L., J. B. Mouronval, P. Aubry, M. Gauthier-Clerc, A. J. Green, H. Fritz & M. Guillemain, 2012. Diet and feeding habitats of Camargue dabbling ducks: what has changed since the 1960s? Waterbirds 35: 555–576.
Brock, M.A., Nielsen, D.L., Shiel, R.J., Green, J.D. & J.D. Langley, 2003. Drought and aquatic community resilience: the role of eggs and seeds in sediments of temporary wetlands. Freshwater Biology 48: 1207–1218
Brouwer, E., L. Denys, E. C. H. E. T. Lucassen, M. Buiks & T. Onelinx, 2017. Competitive strength of Australian swamp stonecrop (Crassula helmsii) invading moorland pools. Aquatic Invasions. https://doi.org/10.3391/AI.2017.12.3.06.
Brouwer, P., H. Schluepmann, K. G. J. Nierop, J. Elderson, P. K. Kijl, I. van der Meer, W. de Visser, G. J. Reichart, S. Smeekens & A. van der Werf, 2018. Growing Azolla to produce sustainable protein feed: the effect of differing species and CO2 concentrations on biomass productivity and chemical composition. Journal of the Science of Food and Agriculture 98: 4759–4768.
Bruckerhoff, L., J. Havel & S. Knight, 2015. Survival of invasive aquatic plants after air exposure and implications for dispersal by recreational boats. Hydrobiologia 746: 113–121.
Brundu, G., 2015. Plant invaders in European and Mediterranean inland waters: profiles, distribution, and threats. Hydrobiologia 746: 61–79.
Brunel, S., 2009. Pathway analysis: aquatic plants imported in 10 EPPO countries. EPPO Bulletin 39: 201–213.
Buchanan, A. L., 2013. Damage by Neochetina weevils (Coleoptera: Curculionidae) induces resistance in Eichhornia crassipes (Commelinales: Pontederiacea). Florida Entomologist 96: 458–462.
Cabrera Walsh, G., M. Dalto, F. M. Mattioli, R. I. Carruthers & L. W. Anderson, 2013. Biology and ecology of Brazilian elodea (Egeria densa) and its specific herbivore, Hydrellia sp., in Argentina. BioControl 58: 133–147.
Caffrey, J. M. & C. Monahan, 2006. Control of Myriophyllum verticillatum L. in Irish canals by turion removal. Hydrobiologia 570: 211–215.
Caffrey, J. M., M. Millane, S. L. Evers & H. Moran, 2011. Management of Lagarosiphon major (Ridley) Moss in Lough Corrib: a review. Biology and Environment 111: 205–212.
Calvo, C., R. P. Mormul, B. R. S. Figueiredo, E. R. Cunha, S. M. Thomaz & M. Meerhoff, 2019. Herbivory can mitigate, but not counteract, the positive effects of warming on the establishment of the invasive macrophyte Hydrilla verticillata. Biological Invasions 21: 59–66.
Casanova, M. T. & M. A. Brock, 2000. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecology 147: 237–250.
Cattaneo, A. & J. Kalff, 1980. The relative contribution of aquatic macrophytes and their epiphytes to the production of macrophyte beds. Limnology and Oceanography 25: 280–289.
Center, T. D. & N. R. Spencer, 1981. The phenology and growth of water hyacinth (Eichhornia crassipes (Mart.) Solms) in a eutrophic north-central Florida lake. Aquatic Botany 10: 1–32.
Chadwell, T. B. & K. A. M. Engelhardt, 2008. Effects of pre-existing submersed vegetation and propagule pressure on the invasion success of Hydrilla verticillata. Journal of Applied Ecology 45: 515–523.
Champion, P. D. & J. S. Clayton, 2001. Border control for potential aquatic weeds: Stage 2: Weed risk assessment. Science for Conservation 185: 30.
Champion, P. D., J. S. Clayton & D. E. Hofstra, 2010. Nipping aquatic plant invasions in the bud: weed risk assessment and the trade. Hydrobiologia 656: 167–172.
Choi, C., C. Bareiss, O. Walenciak & E. M. Gross, 2002. Impact of polyphenols on growth of the aquatic herbivore Acentria ephemerella. Journal of Chemical Ecology 28: 2245–2256.
Chun, Y. J., M. L. Collyer, K. A. Moloney & J. D. Nason, 2007. Phenotypic plasticity of native vs. invasive purple loosestrife: a two-state multivariate approach. Ecology 88: 1499–1512.
Clayton, J. S., 1996. Aquatic weeds and their control in New Zealand lakes. Journal of Lake and Reservoir Management 12: 477–486.
Coetzee, J. A., M. P. Hill, T. Ruiz-Téllez, U. Starfinger & S. Brunel, 2017. Monographs on invasive plants in Europe N° 2: Eichhornia crassipes (Mart.) Solms. Botany Letters 164: 303–326.
Cook, C. D. K. & R. Lüönd, 1982. A revision of the genus Hydrilla (Hydrocharitaceae). Aquatic Botany 13: 485–504.
Cook, C. D. K. & K. Urmi-König, 1985. A revision of the genus Elodea (Hydrocharitaceae). Aquatic Botany 21: 111–156.
Cornacchia, L., D. van der Wal, J. van de Koppel, S. Puijalon, G. Wharton & T. J. Bouma, 2019. Flow-divergence feedbacks control propagule retention by in-stream vegetation: the importance of spatial patterns for facilitation. Aquatic Sciences 81: 17.
Coughlan, N. E., T. C. Kelly & M. A. K. Jansen, 2015. Mallard duck (Anas platyrhynchos)-mediated dispersal of Lemnaceae: a contributing factor in the spread of invasive Lemna minuta? Plant Biology 17: 108–114.
Coughlan, N. E., R. N. Cuthbert, T. C. Kelly & M. A. K. Jansen, 2018. Parched plants: survival and viability of invasive aquatic macrophytes following exposure to various desiccation regimes. Aquatic Botany 150: 9–15.
Creed, R. P. & S. P. Sheldon, 1995. Weevils and watermilfoil: did a North American herbivore cause the decline of an exotic plant? Ecological Applications 5: 1113–1121.
Creed, R. P., S. P. Sheldon & D. M. Cheer, 1992. The effect of herbivore feeding on the buoyancy of Eurasian watermilfoil. Journal of Aquatic Plant Management 30: 75–76.
Cronin, G., D. M. Lodge, M. E. Hay, M. Miller, A. M. Hill, T. Horvath, R. C. Bolser, N. Lindquist & M. Wahl, 2002. Crayfish feeding preferences for fresh water macrophytes: the influence of plant structure and chemistry. Journal of Crustacean Biology 22: 708–718.
Dai, H., X. Lu, J. Zhang & J. Ding, 2014. Responses of a native beetle to novel exotic plant species with varying invasion history. Ecological Entomology 39: 118–124.
Dandelot, S., R. Verlaque, A. Dutartre & A. Cazaubon, 2005. Ecological, dynamic and taxonomic problems due to Ludwigia (Onagraceae) in France. Hydrobiologia 551: 131–136.
Dandelot, S., C. Robles, N. Pech, A. Cazaubon & R. Verlaque, 2008. Allelopathic potential of two invasive alien Ludwigia spp. Aquatic Botany 88: 311–316.
Dawson, W., M. Fischer & M. van Kleunen, 2010. The maximum relative growth rate of common UK plant species is positively associated with their global invasiveness. Global Ecology and Biogeography 20: 299–306.
Della Greca, M., R. Lanzetta, L. Mangoni, P. Monaco & L. Previtera, 1991. A bioactive benzoindenone from Eichhornia crassipes Solms. Bioorganic & Medicinal Chemistry Letters 1: 599–600.
Della Greca, M., R. Lanzetta, A. Molinaro, P. Monaco & L. Previtera, 1992. Phenalene metabolites from Eichhornia crassipes. Bioorganic & Medicinal Chemistry Letters 2: 311–314.
Della Greca, M., A. Fiorentino, P. Monaco & L. Previtera, 1994. Polyoxygenated oleanane triterpenes from Hydrocotyle ranunculoides. Phytochemistry 35: 201–204.
Della Greca, M., A. Fiorentino, P. Monaco, G. Pinto, A. Pollio, L. Previtera & A. Zarrelli, 1998. Structural characterization and antialgal activity of compounds from Pistia stratiotes exudates. Allelopathy Journal 5: 53–66.
De Wilde, M., N. Sebei, S. Puijalon & G. Bornette, 2014. Responses of macrophytes to dewatering: effects of phylogeny and phenotypic plasticity on species performance. Evolutionary Ecology 28: 1155–1167.
Denys, L., J. Packet, W. Jambon & K. Scheers, 2014. Dispersal of the non-native invasive species Crassula helmsii (Crassulaceae) may involve seeds and endozoochorous transport by birds. New Journal of Botany 4: 104–106.
D’hondt, B., L. Denys, W. Jambon, R. de Wilde, T. Adriaens, J. Packet & J. L. H. C. van Valkenburg, 2016. Reproduction of Crassula helmsii by seed in western Europe. Aquatic Invasions 11: 125–130.
Dong, B. C., P. Alpert, W. Guo & F. H. Yu, 2012. Effects of fragmentation on the survival and growth of the invasive, clonal plant Alternanthera philoxeroides. Biological Invasions 14: 1101–1110.
Doyle, R. D., M. D. Francis & R. M. Smart, 2003. Interference competition between Ludwigia repens and Hygrophila polysperma: two morphologically similar aquatic plant species. Aquatic Botany 77: 223–234.
Dugdale, T. M., D. Clements, T. D. Hunt & K. L. Butler, 2010. Alligatorweed produces viable stem fragments in response to herbicide treatment. Journal of Aquatic Plant Management 48: 84–91.
Dugdale, T. M., D. Clements, T. D. Hunt & K. L. Butler, 2012. Survival of a submerged aquatic weed (Egeria densa) during lake drawdown within mounds of stranded vegetation. Lake and Reservoir Management 28: 153–157.
Dugdale, M., K. M. Butler, D. Clements & T. D. Hunt, 2013. Survival of cabomba (Cabomba caroliniana) during lake drawdown within mounds of stranded vegetation. Lake and Reservoir Management 29: 61–67.
Dülger, E. & A. Hussner, 2017. Differences in the growth and physiological response of eight Myriophyllum species to carbon dioxide depletion. Aquatic Botany 139: 25–31.
Dülger, E., P. Heidbüchel, T. Schumann, T. Mettler-Altmann & A. Hussner, 2017. Interactive effects of nitrate concentration and carbon dioxide on the stoichiometry, biomass allocation and growth rate of submerged aquatic plants. Freshwater Biology 62: 1094–1104.
Edwards, D. & C. J. Musil, 1975. Eichhornia crassipes in South Africa: a general review. Journal of the Limnological Society of Southern Africa 1: 23–27.
Elakovich, S. D., 1989. Allelopathic aquatic plants for aquatic weed management. Biologia Plantarum 31: 479–486.
EPPO, 2007. Crassula helmsii. EPPO Bulletin 37: 225–229.
EPPO, 2016a. Alternanthera philoxeroides (Mart.) Griseb. EPPO Bulletin 46: 8–13.
EPPO, 2016b. Myriophyllum heterophyllum Michaux. EPPO Bulletin 46: 20–24.
EPPO, 2017a. Salvinia molesta D.S. Mitch. EPPO Bulletin 47: 531–536.
EPPO, 2017b. Pest risk analysis for Hygrophila polysperma. EPPO, Paris: 51.
EPPO, 2017c. Pistia stratiotes L. EPPO Bulletin 47: 537–543.
Erhard, D. & E. M. Gross, 2006. Allelopathic activity of Elodea canadensis and Elodea nuttallii against epiphytes and phytoplankton. Aquatic Botany 85: 203–211.
Erhard, D., G. Pohnert & E. M. Gross, 2007. Chemical defense in Elodea nuttallii reduces feeding and growth of aquatic herbivorous Lepidoptera. Journal of Chemical Ecology 33: 1646–1661.
Espinosa-Rodriguez, C., L. Rivera-De la Parra, A. Martinez-Tellez, G. C. Gomez-Cabral, S. S. S. Sarma & S. Nandini, 2016. Allelopathic interactions between the macrophyte Egeria densa and plankton (alga, Scenedesmus acutus and cladocerans, Simocephalus spp.): a laboratory study. Journal of Limnology 75: 151–160.
Eusebio Malheiro, A. C., P. Jahns & A. Hussner, 2013. CO2 availability rather than light and temperature determines growth and phenotypical responses in submerged Myriophyllum aquaticum. Aquatic Botany 110: 31–37.
Evans, C. A., D. L. Kelting, K. M. Forrest & L. E. Steblen, 2011. Fragment viability and rootlet formation in Eurasian watermilfoil after desiccation. Journal of Aquatic Plant Management 49: 57–62.
Fan, S., H. Yu, D. Yu, Y. Han & L. Wang, 2015. The effects of complete submergence on the morphological and biomass allocation response of the invasive plant Alternanthera philoxeroides. Hydrobiologia 746: 159–169.
Fast, B. J., C. J. Gray, J. A. Ferrell, G. E. MacDonald & F. M. Fishel, 2008. Water regimen and depth affect Hygrophila growth and establishment. Journal of Aquatic Plant Management 46: 97–99.
Fornoff, F. & E. M. Gross, 2014. Induced defense mechanisms in an aquatic angiosperm to insect herbivory. Oecologia 175: 173–185.
Fernández-Zamudioa, R., P. García-Murilloa & S. Cirujano, 2010. Germination characteristics and sporeling success of Azolla filiculoides Lamarck, an aquatic invasive fern, in a Mediterranean temporary wetland. Aquatic Botany 93: 89–92.
Figuerola, J. & A. J. Green, 2002. Dispersal of aquatic organisms by waterbirds: a review of past research and priorities for future studies. Freshwater Biology 47: 483–494.
Fleming, J. P. & E. D. Dibble, 2015. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia 746: 23–37.
Franceschi, V. R. & P. A. Nakata, 2005. Calcium oxalate in plants: formation and function. Annual Review of Plant Biology 56: 41–71.
Garcia-Álvarez, A., C. H. A. Van Leeuwen, C. J. Luque, A. Hussner, A. Velez-Martin, A. Perez-Vasquez, A. J. Green & E. M. Castellanos, 2015. Internal transport of alien Ludwigia and Spartina seeds by geese and ducks. Freshwater Biology 60: 1316–1329.
Ge, Y., Q. Wang, L. Wang, W. Liu, X. Liu, Y. Huang & P. Christie, 2018. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Archives of Agronomy and Soil Science 64: 708–717.
Geng, Y. P., X. Y. Pan, C. Y. Xu, W. J. Zhang, B. Li & J. K. Chen, 2006. Phenotypic plasticity of invasive Alternanthera philoxeroides in relation to different water availability, compared to its native congener. Acta Oecologia 30: 380–385.
Gillard, M., G. Thiébaut & B. Leroy, 2017a. Present and future distribution of three aquatic plant taxa across the world: decreases in native and increases in invasive ranges. Biological Invasions 19: 2159–2170.
Gillard, M., B. J. Grewell, C. J. Futrell, C. Deleu & G. Thiebaut, 2017b. Germination and seedling growth of water primroses: a cross experiment between two invaded ranges with contrasting climates. Frontiers in Plant Science 8: 1677.
Glisson, W. J., C. K. Wagner, M. R. Verhoeven, R. Muthukrishnan, R. Contreras-Rangel & D. J. Larkin, 2020. Desiccation tolerance of the invasive alga starrystonewort (Nitellopsis obtusa) as an indicator of overland spread risk. Journal of Aquatic Plant Management 58: 7–18.
Global Invasive Species Database (GISD), 2020. Species profile: Lagarosiphon major. Downloaded from http://www.iucngisd.org/gisd/species.php?sc=403 on 25 Apr 2020.
Glover, R., R. E. Drenovsky, C. J. Futrell & B. J. Grewell, 2015. Clonal integration in Ludwigia hexapetala under different light regimes. Aquatic Botany 122: 40–46.
Goodwin, B. J., A. J. McAllister & L. Fahrig, 1999. Predicting invasiveness of plant species based on biological information. Conservation Biology 13: 422–426.
Gopal, B., 1987. Water Hyacinth. Elsevier Science Publishers, Amsterdam.
Gordon, D. R., C. A. Gantz, C. L. Jerde, W. L. Chadderton, R. P. Keller & P. D. Champion, 2012. Weed risk assessment for aquatic plants: modification of a New Zealand system for the United States. PLOS ONE. https://doi.org/10.1371/journal.pone.0040031.
Gratani, L., 2014. Plant phenotypic plasticity as a response to environmental factors. Advances in Botany 2014: 208747.
Greulich, S. & G. Bornette, 1999. Competitive abilities and related strategies in four aquatic plant species from an intermediately disturbed habitat. Freshwater Biology 41: 493–506.
Greulich, S. & G. Bornette, 2003. Being evergreen in an aquatic habitat with attenuated seasonal contrasts: a major competitive advantage? Plant Ecology 167: 9–18.
Gross, E. M., 2003. Allelopathy of aquatic autotrophs. Critical Reviews in Plant Science 22: 313–339.
Gross, E. M., 2009. Allelochemical Reactions. Encyclopedia of Inland Waters 20: 715–726.
Gross, E. M. & E. S. Bakker, 2012. The role of plant secondary metabolites in freshwater macrophyte-herbivore interactions: limited or unexplored chemical defences? In Iason, G. R., M. Dicke & S. E. Hartley (eds), The Integrative Role of Plant Secondary Metabolites in Ecological Systems. BES-British Ecological Society, Sussex.
Gross, E. M., H. Meyer & G. Schilling, 1996. Release and ecological impact of algicidal hydrolysable polyphenols in Myriophyllum spicatum. Phytochemistry 41: 133–138.
Gross, E.M., Erhard, D. & E. Iványi, 2003. Allelopathic activity of Ceratophyllum demersum L. and Najas marina ssp. intermedia (Wolfgang) Casper. Hydrobiologia 506: 583–589.
Gross, E. M., H. Groffier, C. Pestelard & A. Hussner, 2020. Ecology and environmental impact of Myriophyllum heterophyllum, an aggressive invasive in European waterways. Diversity 12: 127.
Groth, A. T., L. Lovett-Doust & J. Lovett-Doust, 1996. Population density and module demography in Trapa natans (Trapaceae), an annual, clonal aquatic macrophyte. American Journal of Botany 83: 1406–1415.
Grutters, B. M. C., E. M. Gross & E. S. Bakker, 2016. Insect herbivory on native and exotic aquatic plants: phosphorus and nitrogen drive insect growth and nutrient release. Hydrobiologia 778: 209–220.
Grutters, B. M. C., Y. O. A. Roijendijk, W. C. E. P. Verberk & E. S. Bakker, 2017a. Plant trait and plant biogeography control the biotic resistance provided by generalist herbivores. Functional Ecology 31: 1184–1192.
Grutters, B. M. C., E. M. Gross, E. van Donk & E. S. Bakker, 2017b. Periphyton density is similar on native and non-native plant species. Freshwater Biology 62: 906–915.
Grutters, B. M. C., B. Saccomanno, E. M. Gross, D. B. Van de Waal, E. van Donk & E. S. Bakker, 2017c. Growth strategy, phylogeny and stoichiometry determine the allelopathic potential of native and non-native plants. Oikos 126: 1770–1779.
Halstead, J. M., J. Michaud, S. Hallas-Burt & P. Gibbs, 2003. Hedonic analysis of effects of a nonnative invader (Myriophyllum heterophyllum) on New Hampshire (USA) lakefront properties. Environmental Management 32: 391–398.
Haramoto, T. & I. Ikusima, 1988. Life cycle of Egeria densa Planch., an aquatic plant naturalized in Japan. Aquatic Botany 30: 389–403.
Hartleb, F. C., J. D. Madsen & C. W. Boylen, 1993. Environmental factors affecting seed germination in Myriophyllum spicatum L. Aquatic Botany 45: 15–25.
Hegi, G., 1975. Illustrierte Flora von Mitteleuropa, V/2. Verlag Paul Parey, Berlin.
Heidbüchel, P. & A. Hussner, 2019. Fragment type and water depth determine the regeneration and colonization success of submerged aquatic plants. Aquatic Sciences 81: 6.
Heidbüchel, P. & A. Hussner, 2020. Falling into pieces: Influence of discharge on fragmentation rates of submerged aquatic plants in lowland streams. Aquatic Botany 160.
Heidbüchel, P., K. Kuntz & A. Hussner, 2016. Alien aquatic plants do not have higher fragmentation rates than native species: a field study from the River Erft. Aquatic Sciences 78: 767–777.
Heidbüchel, P., M. Sachs, N. Stanik & A. Hussner, 2019a. Species-specific fragmentation rates and colonization potential partly explain the successful spread of aquatic plants in lowland streams. Hydrobiologia 843: 107–123.
Heidbüchel, P., P. Jahns & A. Hussner, 2019b. Chlorophyll fluorometry sheds light on the role of desiccation resistance for vegetative overland dispersal of aquatic plants. Freshwater Biology 64: 1401–1415.
Heidbüchel, P., Sachs, M., Hamzehian, N. & A. Hussner, 2020. Go with the flow: Fragment retention patterns shape the vegetative dispersal of aquatic plants in lowland streams. Freshwater Biology 65: 1936–1949.
Hellmann, J. J., J. E. Byers, B. G. Bierwagen & J. S. Dukes, 2008. Five potential consequences of climate change for invasive species. Conservation Biology 22: 534–543.
Henry-Silva, G. G., A. F. M. Camargo & M. M. Pezzato, 2008. Growth of free-floating aquatic macrophytes in different concentrations of nutrients. Hydrobiologia 610: 153–160.
Higgins, S. I. & D. M. Richardson, 2014. Invasive plants have broader physiological niches. Proceeding of the National Academy of Sciences of the United States of America 29: 10610–10614.
Hill, M. P., 2003. The impact and control of alien aquatic vegetation in South African aquatic ecosystems. African Journal of Aquatic Sciences 28: 19–24.
Hill, M. P. & A. J. McConnachie, 2009. Azolla filiculoides Lamarck (Azollaceae). In Muniappan, R., G. V. P. Reddy & A. Raman (eds), Biological Control of Tropical Weeds using Arthropods. Cambridge University Press, Cambridge: 74–87.
Hill, M.P., Coetzee, J.A., Martin, G.D., Smith, R. & E.F. Strange, 2020. Invasive Alien Aquatic Plants in South African Freshwater Ecosystems. In: van Wilgen B., Measey J., Richardson D., Wilson J. & T. Zengeya (eds) Biological Invasions in South Africa. Invading Nature - Springer Series in Invasion Ecology, vol 14. Springer, Cham, pp. 97-114. https://doi.org/10.1007/978-3-030-32394-3_4
Hofmann, H., A. Lorke & F. Peeters, 2008. Wave-induced variability of the underwater light climate in the littoral zone. Verhandlungen des Internationalen Vereins der Limnolologie 30: 627–632.
Hofstra, D. E., J. S. Clayton & J. D. Green, 1999. Distribution and density of vegetative Hydrilla propagules in the sediments of two New Zealand lakes. Journal of Aquatic Plant Management 37: 41–44.
Holm, L. G., L. W. Weldon & R. D. Blackburn, 1969. Aquatic Weeds. Science 166: 699–709.
Howard-Williams, C. & J. Davies, 1988. The invasion of Lake Taupo by the submerged water weed Lagarosiphon major and its impact on the native flora. New Zealand Journal of Ecology 11: 13–19.
Huang, Y., Y. Ge, Q. Wang, H. Zhou, W. Liu & P. Christie, 2017. Allelopathic effects of aqueous extracts of Alternanthera philoxeroides on the growth of Zoysia matrella. Polish Journal of Environmental Studies 26: 97–105.
Hummel, M. & E. Kiviat, 2004. Review of world literature on Water Chestnut with implications for management in North America. Journal of Aquatic Plant Management 42: 17–28.
Hussner, A., 2008. Zur Ökologie und Ökophysiologie aquatischer Neophyten in Nordrhein-Westfalen. PhD thesis, Heinrich-Heine-University, Düsseldorf
Hussner, A., 2009. Growth and photosynthesis of four invasive aquatic plant species in Europe. Weed Research 49: 506–515.
Hussner, A., 2010. Growth response and root system development of the invasive Ludwigia grandiflora and Ludwigia peploides to nutrient availability and water level. Fundamental and Applied Limnology 177: 189–196.
Hussner, A., 2012. Alien aquatic plants in European countries. Weed Research 52: 397–406.
Hussner, A., 2014. Long-term macrophyte mapping documents a continuously shift from native to non-native aquatic plant dominance in the thermally abnormal River Erft (North Rhine-Westphalia, Germany). Limnologica 48: 39–45.
Hussner, A. 2019. Information on measures and related costs in relation to the species included on the Union list - Lagarosiphon major. Technical note prepared by IUCN for the European Commission.
Hussner, A. & R. Lösch, 2005. Alien aquatic plants in a thermally abnormal river and their assembly to neophyte-dominated macrophyte stands (River Erft, Northrhine-Westphalia). Limnologica 35: 18–30.
Hussner, A. & C. Meyer, 2009. The influence of water level on growth and photosynthesis of Hydrocotyle ranunculoides L.fil. Flora 204: 755–761.
Hussner, A. & P. Jahns, 2015. European native Myriophyllum spicatum showed a higher HCO -3 use capacity than alien invasive Myriophyllum heterophyllum. Hydrobiologia 746: 171–182.
Hussner, A., C. Meyer & J. Busch, 2009. Influence of water level on growth and root system development of Myriophyllum aquaticum (Vell.) Verdcourt. Weed Research 49: 73–80.
Hussner, A., K. van de Weyer, E. M. Gross & S. Hilt, 2010. Comments on increasing number and abundance of non-indigenous aquatic macrophyte species in Germany. Weed Research 50: 519–526.
Hussner, A., D. Hofstra & P. Jahns, 2011. Diurnal courses of net photosynthesis and photosystem II quantum efficiency of submerged Lagarosiphon major under natural light conditions. Flora 206: 904–909.
Hussner, A., S. Nehring & S. Hilt, 2014a. From first reports to successful control: a plea for improved management of alien aquatic plant species in Germany. Hydrobiologia 737: 321–331.
Hussner, A., P. Heidbüechel & S. Heiligtag, 2014b. Vegetative overwintering and reproduction by seeds explains the establishment of invasive Pistia stratiotes within the River Erft system (North Rhine-Westphalia, Germany). Aquatic Botany 119: 28–32.
Hussner, A., H. van Dam, J. E. Vermaat & S. Hilt, 2014c. Comparison of native and neophytic aquatic macrophyte developments in a geothermally warmed river and thermally normal channels. Fundamental and Applied Limnology 185: 155–166.
Hussner, A., Hofstra, D., Jahns, P. & J. Clayton, 2015. Response capacity to CO2 depletion rather than temperature and light effects explain the growth success of three alien Hydrocharitaceae compared with native Myriophyllum triphyllum in New Zealand. Aquatic Botany 120: 205–211
Hussner, A., M. Windhaus & U. Starfinger, 2016a. From weed biology to successful control: an example of successful management of Ludwigia grandiflora in Germany. Weed Research 56: 434–441.
Hussner, A., T. Mettler-Altmann, A. P. M. Weber & K. Sand-Jensen, 2016b. Acclimation of photosynthesis to supersaturated CO2 in aquatic plant bicarbonate users. Freshwater Biology 61: 1720–1732.
Hussner, A., I. Stiers, M. J. J. M. Verhofstad, E. S. Bakker, B. M. C. Grutters, J. Haury, J. L. C. H. van Valkenburg, G. Brundu, J. Newman, J. S. Clayton, L. W. J. Anderson & D. Hofstra, 2017. Management and control methods of invasive alien aquatic plants: a review. Aquatic Botany 136: 113–137.
Hussner, A., R. Smith, T. Mettler-Altmann, M. Hill & J. Coetzee, 2019. Simulated global increase in atmospheric CO2 alter the tissue composition, but not the growth of some submerged aquatic plant bicarbonate users growing in DIC rich waters. Aquatic Botany 153: 44–50.
Jacobs, M. J. & H. J. MacIsaac, 2009. Modelling spread of the invasive macrophyte Cabomba caroliniana. Freshwater Biology 54: 296–305.
James, C. S., J. W. Eaton & K. Hardwick, 2006. Responses of three invasive aquatic macrophytes to nutrient enrichment do not explain their observed field displacements. Aquatic Botany 84: 347–353.
Janauer, G. A., 1981. Elodea canadensis and its dormant apices: an investigation of organic and mineral constituents. Aquatic Botany 11: 231–243.
Janes, R., 1998a. Growth and survival of Azolla filiculoides in Britain. II. Sexual reproduction. New Phytologist 138: 377–384.
Janes, R., 1998b. Growth and survival of Azolla filiculoides in Britain. I. Vegetative reproduction. New Phytologist 138: 367–375.
Johnson, L.E., Ricciardi, A. & J.T. Carlton, 2001. Overland dispersal of aquatic invasive species: A risk assessment of transient recreational boating. Ecological Applications 11: 1789–1799.
Julien, M. H., M. P. Hill & P. W. Tipping, 2009. Salvinia molesta DS Mitchell (Salviniaceae). In Muniappan, R., G. V. Reddy & A. Raman (eds), Biological Control of Tropical Weeds using Arthropods. Cambridge University Press, Cambridge: 378–407.
Kadono, Y., 2004. Alien aquatic plants naturalized in Japan: history and present status. Global Environmental Research 8: 163–169.
Kato-Noguchi, H., M. Moriyasu, O. Ohno & K. Suenaga, 2014. Growth limiting effects on various terrestrial plant species by an allelopathic substance, loliolide, from water hyacinth. Aquatic Botany 117: 56–61.
Kattge, J., S. Diaz, S. Lavorel, I. C. Prentice, P. Leadley, G. Bönisch, E. Garnier, et al., 2011. TRY: a global database of plant traits. Global Change Biology 17: 2905–2935.
Keane, R. M. & M. J. Crawley, 2002. Exotic plant invasions and the enemy release hypothesis. Trends in Ecology and Evolution 17: 164–170.
Klavsen, S. K. & S. C. Maberly, 2009. Crassulacean acid metabolism contributes significantly to the in situ carbon budget in a population of the invasive aquatic macrophyte Crassula helmsii. Freshwater Biology 54: 105–118.
Kleinowski, A. M., G. A. Ribeiro, C. Milech & E. J. B. Braga, 2016. Potential allelopathic and antibacterial activity from Alternanthera philoxeroides. Hoehnea 43: 533–540.
Kolar, C. S. & D. M. Lodge, 2001. Progress in invasion biology: predicting invaders. Trends in Ecology & Evolution 16: 199–204.
Kozhova, O. M. & L. A. Izhboldina, 1993. Spread of Elodea canadensis in Lake Baikal. Hydrobiologia 259: 203–211.
Kulshretha, M. & B. Gopal, 1983. Allelopathic influence of Hydrilla verticillata (L.F.) Royle on the distribution of Ceratophyllum species. Aquatic Botany 16: 207–209.
Kuntz, K., P. Heidbüchel & A. Hussner, 2014. Effects of water nutrients on regeneration capacity of submerged aquatic plant fragments. Annales de Limnologie 50: 155–162.
Lal, C. & B. Gopal, 1993. Production and germination of seeds in Hydrilla verticillata. Aquatic Botany 45: 257–261.
Langeland, K. A. & D. L. Sutton, 1980. Regrowth of Hydrilla from axillary buds. Journal of Aquatic Plant Management 18: 27–29.
Langeland, K. A. & C. B. Smith, 1984. Hydrilla produces viable seed in North Carolina lakes. Aquatics 6: 20–22.
Leu, E., A. Krieger-Liszkay, C. Goussias & E. M. Gross, 2002. Polyphenolic allelochemicals from the aquatic angiosperm Myriophyllum spicatum inhibit photosystem II. Plant Physiology 130: 2011–2018.
Liu, M., F. Zhou, X. Pan, Z. Zhang, M. B. Traw & B. Li, 2018. Specificity of herbivore- induced responses in an invasive species, Alternanthera philoxeroides (alligator weed). Ecology and Evolution 8: 59–70.
Loop, J.M.M., Tjampens, J., Vogels, J.J., van Kleef, H.H., Lamers, L.P.M. & R.S.E.W. Leuven, 2020. Reducing nutrient availability and enhancing biotic resistance limits settlement and growth of the invasive Australian swamp stonecrop (Crassula helmsii). Biological Invasions 22: 3391–3402.
Lumpkin, T. A. & D. L. Plucknett, 1980. Azolla: botany, physiology, and use as a green manure. Economic Botany 34: 111–153.
Maberly, S. C. & T. V. Madsen, 2002. Freshwater angiosperm carbon concentrating mechanisms: processes and patterns. Functional Plant Biology 29: 393–405.
Madsen, J. D., 1993. Growth and biomass allocation patterns during waterhyacinth mat development. Journal of Aquatic Plant Management 31: 134–137.
Madsen, T. V. & K. Sand-Jensen, 1991. Photosynthetic carbon assimilation in aquatic macrophytes. Aquatic Botany 41: 5–40.
Madsen, T. V. & K. Sand-Jensen, 1994. The interactive effects of light and inorganic carbon on aquatic plant growth. Plant, Cell and Environment 17: 955–962.
Madsen, J. D. & D. H. Smith, 1999. Vegetative spread of dioecious Hydrilla colonies in experimental ponds. Journal of Aquatic Plant Management 37: 25–29.
Maki, K. & S. Galatowitsch, 2004. Movement of invasive aquatic plants into Minnesota (USA) through horticultural trade. Biological Conservation 118: 389–396.
Martin, G. & J. A. Coetzee, 2011. Pet stores, aquarists and the internet trade as modes of introduction and spread of invasive macrophytes in South Africa. Water SA 37: 371–380.
Matthews, J., Koopman, K.R., Beringen, R., Odé, B., Pot, R., van der Velde, G., van Valkenburg, J.L.H.C. & R.S.E.W. Leuven, 2014. Knowledge document for risk analysis of the non-native Brazilian waterweed (Egeria densa) in the Netherlands. Reports Environmental Science 468: 58.
McFarland, D.G., Nelson, S.L., Grodowitz, M.J., Smart, R.M. & C.S. Owens, 2004. Salvinia molesta D.S. Mitchell (Giant Salvinia) in the United States: A review of species ecology and approaches to management. Aquatic Plant Control Research Program. US Army Engineer Research and Development Center, Environmental Laboratory, Vicksbury.
Menzel, A. & P. Fabian, 1999. Growing season extend in Europe. Nature 397: 659.
Methe, B. A., R. J. Soracco, J. D. Madsen & C. W. Boylen, 1993. Seed production and growth of waterchestnut as influenced by cutting. Journal Aquatic Plant Management 31: 154–157.
Middelboe, A. L. & S. Markager, 1997. Depth limits and minimum light requirements of freshwater macrophytes. Freshwater Biology 37: 553–568.
Millane, M. & J. Caffrey, 2014. Risk Assessment of Ludwigia spp. (Water Primroses). Report prepared for Inland Fisheries Ireland and the National Biodiversity Data Centre.
Mitchell, D. S. & N. M. Tur, 1975. The rate of growth of Salvinia molesta (S. auriculata Auct.) in laboratory and natural conditions. Journal of Applied Ecology 12: 213–225.
Mikulyuk, A. & M.E. Nault, 2009. Water Chestnut (Trapa natans): A Technical Review of Distribution, Ecology, Impacts, and Management. Wisconsin Department of Natural Resources Bureau of Science Services, PUB-SS-10542009. Madison, Wisconsin.
Morrsion, W. E. & M. E. Hay, 2011. Induced chemical defenses in a freshwater macrophyte suppress herbivore fitness and the growth of associated microbes. Oecologia 165: 427–436.
Murphy, K., A. Efremov, T. A. Davidson, E. Molina-Navarro, K. Fidanza, E. M. C. Betiol, P. Chambers, et al., 2019. World distribution, diversity and endemism of aquatic macrophytes. Aquatic Botany 158: 103127.
Nakai, S., Y. Inoue, M. Hosomi & A. Murakami, 1999. Growth inhibition of blue-green algae by allelopathic effects of macrophyte. Water Science and Technology 39(8): 47–53.
Nault, M.E. & A. Mikulyuk, 2009a. East Indian Hygrophila (Hygrophila polysperma): A Technical Review of Distribution, Ecology, Impacts, and Management. Wisconsin Department of Natural Resources Bureau of Science Services, PUB-SS-10492009. Madison, Wisconsin.
Nault, M.E. & A. Mikulyuk, 2009b. African Elodea (Lagarosiphon major): A Technical Review of Distribution, Ecology, Impacts, and Management. Wisconsin Department of Natural Resources Bureau of Science Services, PUB-SS-10502009. Madison, Wisconsin.
Newman, R. M., 1991. Herbivory and detritivory on fresh-water macrophytes by invertebrates: a review. Journal of the North American Benthological Society 10: 89–114.
Netherland, M. D., 1997. Turion ecology of Hydrilla. Journal of Aquatic Plant Management 35: 1–10.
Netten, J. J. C., G. H. P. Arts, R. Gylstra, E. H. Van Nes, M. Scheffer & R. M. M. Roijackers, 2010. Effect of temperature and nutrients on the competition between free-floating Salvinia natans and submerged Elodea nuttallii in mesocosms. Fundamental and Applied Limnology 177: 125–132.
Netten, J. J. C., J. Van Zuidam, S. Kosten & E. T. H. M. Peeters, 2011. Differential response to climatic variation of free-floating and submerged macrophytes in ditches. Freshwater Biology 56: 1761–1768.
Nielsen, S. L. & K. Sand-Jensen, 1991. Variation in growth rates of submerged rooted macrophytes. Aquatic Botany 39: 109–120.
Neuenschwander, P., M. H. Julien, T. D. Center & M. P. Hill, 2009. Pistia stratiotes L. (Araceae). In Muniappan, R., G. V. P. Reddy & R. Raman (eds), Biological Control of Tropical Weeds Using Arthropods. Cambridge University Press, Cambridge: 332–352.
Ostrofsky, M. L. & E. R. Zettler, 1986. Chemical defences in aquatic plants. Journal of Ecology 74: 279–287.
Owens, C. S. & J. D. Madsen, 1995. Low temperature limits of Waterhyacinth. Journal of Aquatic Plant Management 33: 63–68.
Owens, C. S., J. D. Madsen, R. M. Smart & R. M. Stewart, 2001. Dispersal of native and nonnative aquatic plant species in the San Marcos River, Texas. Journal of Aquatic Plant Management 39: 75–79.
Owens, C. S., R. M. Smart & R. M. Stewart, 2004. Low temperature limits of giant Salvinia. Journal of Aquatic Plant Management 42: 91–94.
Parker, J. D. & M. E. Hay, 2005. Biotic resistance to plant invasions? Native herbivores prefer non-native plants. Ecology Letters 8: 959–967.
Pedersen, O., T. D. Colmer & K. Sand-Jensen, 2013. Underwater photosynthesis of submerged plants: recent advances and methods. Frontiers in Plant Science 4: 140.
Petruzzella, A., B. M. C. Grutters, S. M. Thomaz & E. S. Bakker, 2017. Potential for biotic resistance from herbivores to tropical and subtropical plant invasions in aquatic ecosystems. Aquatic Invasions 12: 343–353.
Phartyal, S. S., S. Rosbakh & P. Poschlod, 2018. Seed germination ecology in Trapa natans L., a widely distributed freshwater macrophyte. Aquatic Botany 147: 18–23.
Pieterse, A. H., L. Delange & L. Verhagan, 1981. A study of certain aspects of seed germination and growth of Pistia stratiotes L. Acta Botanica Neerlandica 30: 47–58.
Poovey, A. G. & S. H. Kay, 1998. The potential of a summer drawdown to manage monoecious Hydrilla. Journal of Aquatic Plant Management 36: 127–130.
Prinz, M., C. Peppler-Lisbach, A. Weidhüner & H. Freund, 2019. Crassula helmsii (T. Kirk) Cockayne–Habitat requirements, distribution and vegetation community composition of an alien invasive species on Norderney. Tuexenia 39: 267–286.
Pulzatto, M. M., L. A. Lolis, N. Louback-Franco & R. P. Mormul, 2018. Herbivory on freshwater macrophytes from the perspective of biological invasions: a systematic review. Aquatic Ecology 52: 297–309.
Qiu, J. W. & K. L. Kwong, 2009. Effects of macrophytes on feeding and life-history traits of the invasive apple snail Pomacea canaliculata. Freshwater Biology 54: 1720–1730.
Rao, P. N. & A. S. Reddy, 1984. Studies on the population biology of water lettuce: Pistia stratiotes L. Hydrobiologia 119: 15–19.
Rascio, N., 2002. The underwater life of secondarily aquatic plants: some problems and solutions. Critical Reviews in Plant Sciences 21(4): 401–427.
Rattray, M. R., C. Howard-Williams & J. M. A. Brown, 1991. Sediment and water as sources of nitrogen and phosphorus for submerged rooted aquatic macrophytes. Aquatic Botany 40: 225–237.
Rector, P. R., P. J. Nitzsche & S. S. Mangiafico, 2015. Temperature and herbicide impacts on germination of water chestnut seeds. Journal of Aquatic Plant Management 53: 105–112.
Reddy, K. R. & W. F. DeBusk, 1983. Growth characteristics of aquatic macrophytes cultured in nutrient-enriched water: I. Water Hyacinth, Water Lettuce, and Pennywort. Economic Botany 38: 229–239.
Reddy, K. R., M. Agami & J. C. Tucker, 1990. Influence of phosphorus on growth and nutrient storage by water hyacinth (Eichhornia crassipes (Mart.) Solms) plants. Aquatic Botany 37: 355–365.
Redekop, P., D. Hofstra & A. Hussner, 2016. Elodea canadensis shows a higher dispersal capacity via fragmentation than Egeria densa and Lagarosiphon major. Aquatic Botany 130: 45–49.
Redekop, P., E. M. Gross, A. Nuttens, D. Hofstra, J. S. Clayton & A. Hussner, 2018. Hygraula nitens, the only native aquatic caterpillar in New Zealand, prefers an alien submerged plant. Hydrobiologia 812: 13–25.
Reichard, S. H. & P. White, 2001. Horticulture as a pathway of invasive plant introductions in the United States. BioScience 51: 103–113.
Rejmànková, E., 1992. Ecology of creeping macrophytes with special reference to Ludwigia peploides (H. B. K.) Raven. Aquatic Botany 43: 283–299.
Reynolds, C., N. A. Miranda & G. S. Cumming, 2015. The role of waterbirds in the dispersal of aquatic alien and invasive species. Diversity and Distributions 21: 744–754.
Riede, W., 1920. Untersuchungen über Wasserpflanzen. Flora 114: 1–118.
Riis, T., T. V. Madsen & R. S. H. Sennels, 2009. Regeneration, colonisation and growth rates of allofragments in four common stream plants. Aquatic Botany 90: 209–212.
Riis, T., C. Lambertini, B. Olesen, J. S. Clayton, H. Brix & B. K. Sorrell, 2010. Invasion strategies in clonal aquatic plants: are phenotypic differences caused by phenotypic plasticity or local adaptation? Annals of Botany 106: 813–822.
Riis, T., B. Olesen, J. S. Clayton, C. Lambertini, H. Brix & B. K. Sorrell, 2012. Growth and morphology in relation to temperature and light availability during the establishment of three invasive aquatic plant species. Aquatic Botany 102: 56–64.
Room, P. M., 1992. Equations relying growth and uptake of nitrogen by Salvinia molesta to temperature and the availability of nitrogen. Aquatic Botany 24: 43–59.
Ruaux, B., S. Greulich, J. Haury & J. P. Berton, 2009. Sexual reproduction of two alien invasive Ludwigia (Onagraceae) on the middle Loire River, France. Aquatic Botany 90: 143–148.
Rybicki, N. B., D. G. McFarland, H. A. Ruhl, J. T. Reel & J. W. Barko, 2001. Investigations of the availability and survival of submersed aquatic vegetation propagules in the tidal Potomac River. Estuaries 24: 407–424.
Šajna, N., M. Haler, S. Škornik & M. Kaligarič, 2007. Survival and expansion of Pistia stratiotes L. in a thermal stream in Slovenia. Aquatic Botany 87: 75–79.
Saito, K., M. Matsumoto, T. Sekine, I. Murakoshi, N. Morisaki & S. Iwasaki, 1989. Inhibitory substances from Myriophyllum. Journal of Natural Products 52: 1221–1226.
Salvucci, M. E. & G. Bowes, 1982. Photosynthetic and photorespiratory responses of the aerial and submerged leaves of Myriophyllum brasiliense (Myriophyllum aquaticum). Aquatic Botany 13: 147–164.
Sand-Jensen, K., 1989. Environmental variables and their effects on photosynthesis of aquatic plant communities. Aquatic Botany 34: 5–25.
Sand-Jensen, K. & H. Frost-Christensen, 1999. Plant growth and photosynthesis in the transition zone between land and stream. Aquatic Botany 63: 23–35.
Santamaria, L., 2002. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment. Acta Oecologia 23: 137–154.
Santos, M. J., L. W. Anderson & S. L. Ustin, 2011. Effects of invasive species on plant communities: an example using submerged aquatic plants at the regional scale. Biological Invasions 13: 443–457.
Sarneel, J. M., 2013. The dispersal capacity of vegetative propagules of riparian fen species. Hydrobiologia 710: 219–225.
Schooler, S., 2012. Alternanthera philoxeroides (Martius) Grisebach (alligator weed). In Francis, R. A. (ed.), A Handbook of Global Freshwater Invasive Species. Earthscan Publisher, London: 25–35.
Sculthorpe, C. D., 1967. The Biology of Aquatic Vascular Plants. Arnold, London.
Silva, G. G., A. J. Green, V. Weber, P. Hoffmann, A. Lovas-Kiss, C. Stenert & L. Maltchik, 2018. Whole angiosperms Wolffia columbiana disperse by gut passage through wildfowl in South America. Biology Letters 14: 20180703.
Simberloff, D., in press. Maintenance management and eradication of established aquatic invaders. Hydrobiologia. https://doi.org/10.1007/s10750-020-04352-5
Smida, I., C. Charpy-Roubaud, S. Y. Cherif, F. Torre, G. Audran, S. Smiti & J. Le Petit, 2015. Antibacterial properties of extracts of Ludwigia peploides subsp montevidensis and Ludwigia grandiflora subsp hexapetala during their cycle of development. Aquatic Botany 121: 39–45.
Smith, T. & P. Buckley, 2020. Biological Flora of the British Isles: Crassula helmsii. Journal of Ecology 108: 797–813.
Sousa, W. T. Z., 2011. Hydrilla verticillata (Hydrocharitaceae), a recent invader threatening Brazil’s freshwater environments: a review of the extent of the problem. Hydrobiologia 669: 1–20.
Spencer, W. & G. Bowes, 1985. Limnophila and Hygrophila: a review and physiological assessment of their weed potential in Florida. Journal of Aquatic Plant Management 23: 7–16.
Spencer, W. & G. Bowes, 1990. Ecophysiology of the world’s most troublesome aquatic weeds. In Pieterse, A. H. & K. J. Murphy (eds), Aquatic Weeds – The Ecology and Management of Nuisance Aquatic Vegetation. Oxford University Press, Oxford: 39–73.
Stiers, I., N. Crohain, G. Josens & L. Triest, 2011a. Impact of three aquatic invasive species on native plants and macroinvertebrates in temperate ponds. Biological Invasions 13: 2715–2726.
Stiers, I., J. Njambuya & L. Triest, 2011b. Competitive abilities of invasive Lagarosiphon major and native Ceratophyllum demersum in monocultures and mixed cultures in relation to experimental sediment dredging. Aquatic Botany 95: 161–166.
Storrs, M. J. & M.H. Julien, 1996. Salvinia: A Handbook for the Integrated Control of Salvinia molesta in Kakadu National Park. Northern Landscapes Occasional Papers No. 1. Darwin, Australia: Australian Nature Conservation Agency, 58 pp.
Thiebaut, G. & L. Martinez, 2015. An exotic macrophyte bed may facilitate the anchorage of exotic propagules during the first stage of invasion. Hydrobiologia 746: 183–196.
Thiebaut, G., H. Rodriguez-Perez & O. Jambon, 2019. Reciprocal interactions between the native Mentha aquatica and the invasive Ludwigia hexapetala in an outdoor experiment. Aquatic Botany 157: 17–23.
Thiele, J. & A. Otte, 2008. Invasion patterns of Heracleum mantegazzianum in Germany on the regional and landscape scales. Journal for Nature Conservation 16: 61–71.
Thomas, P. A. & P. M. Room, 1986. Taxonomy and control of Salvinia molesta. Nature 320: 581–584.
Thomaz, S. M., R. P. Mormul & T. S. Michelan, 2015. Propagule pressure, invasibility of freshwater ecosystems by macrophytes and their ecological impacts: a review of tropical freshwater ecosystems. Hydrobiologia 746: 39–59.
Thouvenot, L., J. Haury & G. Thiebaut, 2013a. Seasonal plasticity of Ludwigia grandiflora under light and water depth gradients: an outdoor mesocosm experiment. Flora 208: 430–437.
Thouvenot, L., J. Haury & G. Thiebaut, 2013b. A success story: water primroses, aquatic plant pests. Aquatic Conservation: Marine and Freshwater Ecosystems 23: 790–803.
Thouvenot, L., C. Puech, L. Martinez, J. Haury & G. Thiébaut, 2013c. Strategies of the invasive macrophyte Ludwigia grandiflora in its introduced range: competition, facilitation or coexistence with native and exotic species? Aquatic Botany 107: 8–16.
Thouvenot, L., B. Gauzens, J. Haury & G. Thiebaut, 2019. Response of macrophyte traits to herbivory ad neighbouring species: integration of the functional trait framework in the context of ecological invasions. Frontiers in Plant Science 9: 1981.
Tipping, P.W., 2004. Giant Salvinia. In: Coombs, E.M., Clark, J.K., Piper, G.L. & A.F. Cofrancesco (eds): Biological control of invasive plants in the United States. Oregon State University Press, Corvallis, pp. 174–177.
Touchette, B. W., L. R. Iannacone, E. T. Gwendolyn & A. R. Frank, 2007. Drought tolerance versus drought avoidance: a comparison of plant-water relations in herbaceous wetland plants subjected to water withdrawal and repletion. Wetlands 27: 656–667.
Umetsu, C. A., H. B. A. Evangelista & S. M. Thomaz, 2012. The colonization, regeneration, and growth rates of macrophytes from fragments: a comparison between exotic and native submerged aquatic species. Aquatic Ecology 46: 443–449.
van Kleunen, M., W. Dawson & P. Dostal, 2011. Research on invasive-plant traits tells us a lot. Trends in Ecology & Evolution 26: 317.
Venter, N., B. W. Cowie, E. T. F. Witkowski, G. C. Snow & M. J. Byrne, 2017. The amphibious invader: Rooted water hyacinth’s morphological and physiological strategy to survive stranding and drought events. Aquatic Botany 143: 41–48.
Vilà, M. & J. Weiner, 2004. Are invasive plant species better competitors than native plant species? - evidence from pair-wise experiments. Oikos 105: 229–238.
Walenciak, O., W. Zwisler & E. M. Gross, 2002. Influence of Myriophyllum spicatum-derived tannins on gut microbiota of its herbivore Acentria ephemerella. Journal of Chemical Ecology 28: 2045–2056.
Wang, N., F. H. Yu, P. X. Li, W. M. He, F. H. Liu, J. M. Liu & M. Dong, 2008. Clonal integration affects growth, photosynthetic efficiency and biomass allocation, but not the competitive ability, of the alien invasive Alternanthera philoxeroides under severe stress. Annals of Botany 101: 671–678.
Wang, P., Y. S. Xu, B. C. Dong, W. Xue & F. H. Yu, 2014. Effects of clonal fragmentation on intraspecific competition of a stoloniferous floating plant. Plant Biology 16: 1121–1126.
Wang, P., P. Alpert & F. H. Yu, 2016a. Clonal integration increases relative competitive ability in an invasive aquatic plant. American Journal of Botany 103: 2079–2086.
Wang, Y. J., Y. F. Bai, S. Q. Zeng, B. Yao, W. Wang & F. L. Luo, 2016b. Heterogeneous water supply affects growth and benefits of clonal integration between co-existing invasive and native Hydrocotyle species. Scientific Reports 6: 29420.
Wang, Y. J., H. Müller-Schärer, M. van Kleunen, A. M. Cai, P. Zhang, R. Yan, B. C. Dong & F. H. Yu, 2017. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytologist 216: 1072–1078.
Wei, G. W., Q. Shu, F. L. Luo, Y. H. Chen, B. C. Dong, L. C. Mo, W. J. Huang & F. H. Yu, 2018. Separating effects of clonal integration on plant growth during submergence and de-submergence. Flora 246–247: 118–125.
Wells, C. L. & M. Pigliucci, 2000. Adaptive phenotypic plasticity: the case of heterophylly in aquatic plants. Perspectives in Plant Ecology, Evolution and Systematics 3: 1–18.
Wells, R., M. de Winton & J. S. Clayton, 1997. Successive macrophyte invasions within the submerged flora of Lake Tarawera, Central North Island, New Zealand. New Zealand Journal of Marine and Freshwater Research 31: 449–459.
Wersal, R. M. & J. D. Madsen, 2011. Comparative effects of water level variations on growth characteristics of Myriophyllum aquaticum. Weed Research 51: 386–393.
Wersal, R. M., J. D. Madsen & P. D. Gerard, 2013. Survival of parrotfeather following simulated drawdown events. Journal of Aquatic Plant Management 51: 22–26.
Wheeler, G. S., T. K. Van & T. D. Center, 1998. Herbivore adaptations to a low-nutrient food: weed biological control specialist Spodoptera pectinicornis (Lepidoptera: Noctuidae) fed the floating aquatic plant Pistia stratiotes. Environmental Entomology 27(4): 993–1000.
Wilson, C. E., S. J. Darbyshire & R. Jones, 2007. The biology of invasive alien plants in Canada. 7. Cabomba caroliniana A. Gray. Canadian Journal of Plant Science 87: 615–638.
Winkel, A. & J. Borum, 2009. Use of sediment CO2 by submersed rooted plants. Annals of Botany 103: 1015–1023.
Wolfer, S. & D. Straile, 2012. To share or not to share: clonal integration in a submerged macrophyte in response to light stress. Hydrobiologia 684: 261–269.
Wong, P. K., Y. Liang, N. Y. Liu & J.-W. Qiu, 2010. Palatability of macrophytes to the invasive freshwater snail Pomacea canaliculata: differential effects of multiple plant traits. Freshwater Biology 55: 2023–2031.
Wright, A.D. & M.F. Purcell, 1995. Eichhornia crassipes (Mart.) Solms-Laubach. In: Groves R.H., R.C.H. Shepherd & R.G. Richardson (eds) The Biology of Australian Weeds. R.G. & F.J. Richardson, Melbourne. pp. 111–121.
Wu, H. & J. Ding, 2019. Global change sharpens the double-edged sword effect of aquatic alien plants in China and beyond. Frontiers in Plant Science 10: 787.
Wu, X., H. Wu, J. R. Chen & J. Y. Ye, 2013. Effects of allelochemical extracted from water lettuce (Pistia stratiotes Linn.) on the growth, microcystin production and release of Microcystis aeruginosa. Environmental Science and Pollution Research 20: 8192–8201.
Wu, X., H. Wu, J. Y. Ye & B. Zhong, 2015. Study on the release routes of allelochemicals from Pistia stratiotes Linn., and its anti-cyanobacteria mechanisms on Microcystis aeruginosa. Environmental Science and Pollution Research 22: 18994–19001.
Wu, X. X., Z. Y. Zhang & Y. G. Jin, 2019. Physiological mechanism of Eichhornia crassipes in inhibiting the growth of Microcytis aeruginosa. Russian Journal of Plant Physiology 66: 433–439.
Xiao, K., D. Yu, X. Xu & W. Xiong, 2007. Benefits of clonal integration between interconnected ramets of Vallisneria spiralis in heterogeneous light environments. Aquatic Botany 86: 76–82.
Xiao, T., H. Yu, Y.-B. Song, Y.-P. Jiang, B. Zeng & M. Dong, 2019. Nutrient enhancement of allelopathic effects of exotic invasive on native plant species. Plos One 14: e0206165.
Xie, Y. & D. Yu, 2003. The significance of lateral roots in phosphorus (P) acquisition of water hyacinth (Eichhornia crassipes). Aquatic Botany 75: 311–321.
Xie, D., R. Hu, R. P. Mormul, H. Ruan, Y. Feng & M. Zhang, 2018. Fragment type and water nutrient interact and affect the survival and establishment of Myriophyllum aquaticum. Hydrobiologia 817: 205–213.
Xue, B. K. & S. Leibler, 2018. Benefits of phenotypic plasticity for population growth in varying environments. Proceedings of the National Academy of Sciences of the United States of America 115: 12745–12750.
Yarrow, M., V. H. Marin, M. Finlayson, A. Tironi, L. E. Delgado & F. Fischer, 2009. The ecology of Egeria densa Planchon (Liliopsida: Alismatales): A wetland ecosystem engineer? Revista Chilena de Historia Natural 82: 299–313.
Ye, X. Q., J. L. Meng, B. Zeng, M. Wu, Y. Y. Zhang & X. P. Zhang, 2016. Submergence causes similar carbohydrate starvation but faster post-stress recovery than darkness in Alternanthera philoxeroides plants. PLoS One 11: e0165193.
Yin, L., W. Li, T. V. Madsen, S. C. Maberly & G. Bowes, 2017. Photosynthetic inorganic carbon acquisition in 30 freshwater macrophytes. Aquatic Botany 140: 48–54.
You, W., D. Yu, C. Liu, D. Xie & W. Xiong, 2013. Clonal integration facilitates invasiveness of the alien aquatic plant Myriophyllum aquaticum L. under heterogeneous water availability. Hydrobiologia 718: 27–39.
You, W. H., C. M. Han, L. X. Fang & D. L. Du, 2016a. Propagule pressure, habitat conditions and clonal integration influence the establishment and growth of an invasive clonal plant, Alternanthera philoxeroides. Frontiers in Plant Science 7: 1–11.
You, W. H., C. M. Han, C. H. Liu & D. Yu, 2016b. Effects of clonal integration on the invasive clonal plant Alternanthera philoxeroides under heterogeneous and homogeneous water availability. Scientific Reports 6: 29767.
Yu, L. Q., Y. Fujii, Y. J. Zhou, J. P. Zhang, Y. L. Lu & S. N. Xuan, 2007. Response of exotic invasive weed Alternanthera philoxeroides to environmental factors and its competition with rice. Rice Science 14: 49–55.
Yu, H., N. Shen, D. Yu & C. Liu, 2019. Clonal integration increases growth performance and expansion of Eichhornia crassipes in littoral zones: a simulation study. Environmental and Experimental Botany 159: 13–22.
Xiong, W., D. Yu, Q. Wang, C. Liu & L. Wang, 2008. A snail prefers native over exotic freshwater plants: implications for the enemy release hypotheses. Freshwater Biology 53: 2256–2263.
Xu, C-Y., Schooler, S.S. & R.D. van Klinken, 2010. Effects of clonal integration and light availability in the gowth and physiology of two invasive herbs. Journal of Ecology 98: 833–844.
Zuo, S., Y. Ma & I. Shinobu, 2012a. Differences in ecological and allelopathic traits among Alternanthera philoxeroides populations. Weed Biology and Management 12: 123–130.
Zuo, S., H. Mei, L. Ye, J. Wang & S. Ma, 2012b. Effects of water quality characteristics on the algicidal property of Alternanthera philoxeroides (Mart.) Griseb. in an aquatic ecosystem. Biochemical Systematics and Ecology 43: 93–100.
Acknowledgement
We heartily acknowledge the helpful comments of the editor S. Thomaz and two anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Guest editors: Katya E. Kovalenko, Fernando M. Pelicice, Lee B. Kats, Jonne Kotta & Sidinei M. Thomaz / Aquatic Invasive Species III
Rights and permissions
About this article

Cite this article
Hussner, A., Heidbüchel, P., Coetzee, J. et al. From introduction to nuisance growth: a review of traits of alien aquatic plants which contribute to their invasiveness. Hydrobiologia 848, 2119–2151 (2021). https://doi.org/10.1007/s10750-020-04463-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-020-04463-z