
Abstract
The relationship between fish and amphibians is thought to be antagonistic because of the frequent amphibian extirpations after fish introductions, although some field observations show that amphibians and fish often co-occur in rivers. Here, we used surveys in north-eastern Spain (99,700 km2, 15 river catchments, N = 535 sites) to identify the most frequent fish–amphibian associations and the riverine conditions that might concurrently promote the diversity of native fish and of five widely distributed amphibian species. Overall, there was little congruence between native fish and amphibian-diversity measures (species richness and the Shannon, Simpson and Pielou diversity indices). Different riverine conditions appeared to be important for the two vertebrate groups. Alien fish richness, which was highly correlated with alien fish abundance, was negatively associated with amphibian richness and Shannon diversity but was positively associated with native fish richness. River water depth was negatively associated with amphibian occurrence. While our snap-shot surveys may be a transitional stage in the fish–amphibian relationships, we found that some widely distributed amphibian species co-occur with fish in rivers in north-eastern Spain. Small rivers, such as tributaries often have the most intact fish assemblages, and probably are the best locations to explore fish–amphibian associations in greater depth.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Human activities are rapidly changing biodiversity patterns around the world (Gaston, 2000; Dirzo et al., 2014; Valiente-Banuet et al., 2015). Many species are declining due to the effects of climate change, land-use transformation, overharvesting, chemical pollution, and the introduction of alien species, all of which are continuing pressures (Dirzo et al., 2014; Ricciardi et al., 2017). Although this biodiversity crisis is occurring in all ecosystems (Butchart et al., 2010; Waldron et al., 2017), riverine ecosystems are among the most severely affected (Vörösmarty et al., 2010). Given the likely on-going intensification of human impacts in rivers (Vörösmarty et al., 2010), identifying the main hazards to multiple taxa may help to improve the efficiency and cost-effectiveness of management strategies.
Fish are often considered a major threat to amphibians given reports of many amphibian extirpations after fish introductions (e.g. Kats & Ferrer, 2003; Denoël et al., 2005; Porej & Hetherington, 2005). However, native fish and amphibians may co-exist if they have co-evolved (Hecnar & M‘Closkey, 1997). Moreover, agonistic interactions (predation, interspecific competition) are likely to be ameliorated in more complex physical habitats because prey are more difficult to locate and catch (Diehl, 1992; Hartel et al., 2007). Nevertheless, poor water quality due to agricultural run-off and sewage effluents threatens riverine fauna (e.g. Kefford et al., 2012; Colin et al., 2016) and may diminish benefits arising from physical habitat quality on fish and amphibians. Common pollutants in poorly treated waters, such as ammonia and nitrite, are highly toxic to all aquatic taxa (Camargo & Alonso, 2006), and so, their effects may be similar across taxa. Relatively well-preserved rivers generally have high spatial heterogeneity, which often is the case for smaller tributaries (Pracheil et al., 2013; Maceda-Veiga et al., 2017). Even though riverine ecosystems are unlikely to provide suitable habitats for all amphibian species in a region (e.g. temporary pool-breeders), rivers may help to conserve those species that have at least some riverine populations (see de Vries & Marco, 2017).
Studies that establish the congruence of diversity measures between fish and amphibians in rivers are a first step to understanding the associations between the two faunal groups and the environmental conditions that might promote their co-occurrence. Cross-taxonomic congruence in diversity measures in rivers has been examined mostly for taxa typically used in monitoring, such as diatoms, plants, invertebrates, and fish (e.g. Heino et al., 2005; Tisseuil et al., 2013; Colin et al., 2016). However, many other taxa occur in rivers, including amphibians, which are among the most at-risk groups on Earth (Alroy, 2015). Moreover, studies on cross-taxonomic congruence in diversity patterns in rivers usually have focused on species richness (Spellerberg, 1991; Heino, 2010). Nevertheless, species richness per se does not account for the relative abundances of species, which underpin diversity indices such as the Shannon–Wiener index (Shannon, 1948).
We used extensive surveys in north-eastern Spain (99,700 km2, 15 catchments) to identify the fish species that co-occur with amphibians and the riverine conditions that might concurrently promote native fish and amphibian diversity. Rivers in this region are subject to the usual range of human impacts (e.g. nutrient pollution, sluggish waters due to water extractions, high conductivity) seen in many rivers around the world (e.g. Tockner et al., 2009). This region has a diverse fish fauna, including widespread native (e.g. Salmo trutta Linnaeus, 1758) and alien species (e.g. Cyprinus carpio Linnaeus, 1758, Doadrio, 2011), and has amphibian species that are widespread in western Europe (e.g. Alytes obstetricans (Laurenti, 1768), Sillero et al., 2014). Previous reports using these data identified different environmental variables to explain variation in native and alien fish richness (Maceda-Veiga et al., 2017). However, the combined effects of alien fish diversity and riverine environmental conditions on native fish and amphibian diversity have not yet been investigated. Moreover, the degree to which native fish and amphibians might co-occur in these riverine ecosystems is largely unknown (de Vries & Marco, 2017).
From a management perspective, efficiencies might be gained if native fish and amphibians co-occurred and responded similarly to environmental factors and to alien fish species. Commonalities would mean that management actions would jointly improve the condition of native-fish and amphibian populations.
Methods
Study region
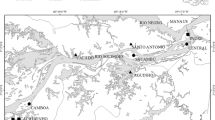
Data on river environmental conditions, fish and amphibians were collected in north-eastern Spain from 2002 to 2009 in several projects (e.g. Maceda-Veiga et al., 2017, 2018) to develop indices of biotic integrity in compliance with the EU Water Framework Directive (EU Directive 2000/60/EC). The study region included all rivers draining to the Mediterranean Sea from the Muga to Riudecanyes basins, the entire Ebro River and part of the Garonne basin (Fig. 1). Sampled sites (N = 535) were selected for accessibility and for representativeness of the full range of environmental conditions in the region (see Table 1). Most of the studied rivers are small or medium-size streams typical of Mediterranean-climate regions, often with low flows in summer and autumnal high flows and even floods. We surveyed in low-flow conditions, mostly from May to September, because this is when aquatic vertebrates can be sampled most effectively using electrofishing (see below). All rivers had continuous flow at the time of sampling. Seasonal droughts appear to induce relatively small, transient changes to Mediterranean stream fish assemblages (Magalhães et al., 2007). Most native fish in north-eastern Spain do not migrate long-distances apart from Anguilla anguilla Linnaeus, 1758 (Doadrio, 2011). Individuals of Barbus haasi Mertens, 1925, one of the most frequently encountered fish in these rivers (Appendix S1), rarely moved > 100 m in a year (Aparicio & de Sostoa, 1999).

Distribution of the 535 sampling sites surveyed in rivers of north-eastern Spain with protected areas in green
Fish and amphibian surveys
We used a standard fish-sampling method (CEN standards EN 14962 and EN 14011). We surveyed the whole wetted width of ≥ 100–m long reaches at each survey location in an upstream direction using single–pass electrofishing with a portable unit generating c. 200 V and 3 A pulsed DC (e.g. Maceda-Veiga et al., 2017). This sampling procedure detected 80–100% of the fish species and captured 60–90% of the individuals compared to estimates from four-pass electrofishing (A.S., unpublished data), so that our data are likely to be representative of the local fish assemblages. We used the same equipment at all sites to avoid potential bias in captures, and we expressed results as captures per unit of effort (CPUE, individuals caught divided by fishing time in minutes and the area surveyed in m2).
For amphibians, the four members of the fish-sampling crew visually inspected river water during electrofishing and all amphibians seen or stunned by electrofishing were recorded. These surveys were complemented by a second visual inspection of the study area and the wetted riverbank during the assessment of habitat quality and the measurement of in-stream hydraulic variables (see below). However, more species were not detected, and we only used the records of electrofishing surveys to be able to standardize the sampling effort. As for fish, amphibian captures were expressed as CPUEs to account for differences in the sampling area. While we may have missed species in a single survey due to the different species’ phenologies (e.g. Llorente et al., 1995; Richter-Boix et al., 2006), multiple surveys were not feasible in this extensive study area. However, the detectability of three of the five species examined in the present study [A. obstetricans; Salamandra salamandra (Linnaeus, 1758); Pelophylax perezi (López Seoane, 1885)] was reasonably high in our method (78–100% of the species detected) compared to multiple dip-net surveys in ten river reaches in four consecutive months.
Surveys were authorized by the Spanish Government and the Autonomous Government of Catalonia (AP/003). All animals were released at the site of capture. The status of fish species was defined as: (a) native, if the species historically occurred in the basin where captured according to regional faunal lists, and (b) alien, if the fish species did not historically occur in the basin of capture or elsewhere in the Iberian Peninsula (Appendix S1). Native species that historically occur only in the Iberian Peninsula are referred to as endemic species.
Geographical location and river environmental conditions
We used 16 geographical and environmental predictors to characterize the riverine conditions. Some environmental predictors (e.g. elevation, nutrient concentrations, pH, conductivity) have been used previously to explore associations between fish and riverine conditions (Maceda-Veiga et al., 2017, 2018). Other descriptors of physical habitat structure (e.g. algae cover, the presence of caves) were measured; these were selected because they may modulate interactions between fish and amphibians by changing habitat complexity.
We recorded the basin name and elevation (m.a.s.l.) using Google Earth®. Elevation was used as a surrogate for the position of the sampling site in the river, which is important because the composition of faunas changes naturally from upstream to downstream (Doadrio, 2011). We calculated the Strahler stream order number as a measure of river size (Strahler, 1964) using a 1:50,000 map. Rivers were ranked from small, first-order tributaries to the largest main river based on a hierarchy of tributaries.
Physical habitat complexity was characterized using six complementary measurements. We used the QBR index (Munné et al., 2003) as an integrated measure of river hydro-morphology and the conservation status of riparian vegetation. QBR consists of landscape and in-site variables, which are combined as described in Munné et al. (2003): presence and extent of riparian areas and their connections to adjacent woodlands; percentages of riparian trees, shrubs and emerged aquatic plants; presence of alien riparian vegetation species; and presence of weirs, channels, and walls. Other measures of habitat complexity were: the percentage of dead wood, macroalgae (e.g. Cladophora sp.) and caves and the percentage of river margins with ferns and herbaceous plants in each sampling site, all of which are not included in the QBR index. Last, we used a modified version of the U.S.A. Rapid Bioassessment protocol (Barbour et al., 1999) to assess microhabitat diversity, which ranks the presence and diversity of structural refuges in the riverbed (e.g. roots, submerged aquatic plants) on a scale from 0 to 20.
At each sampling site, we measured seven water-quality variables before each fish or amphibian survey. We used a digital multiparametric YSI® probe to quantify temperature (°C), conductivity (µS/cm) and pH, and the colorimetric test kit VISOCOLOR® to determine the concentrations of nutrients (ammonium, nitrite, nitrate and phosphate–P, all in mg/l). Agricultural runoff and sewage effluents cause silting that was estimated as the % of the streambed covered by sediment < 1 mm (Townsend et al., 2008). We calculated the mean current speed (m/s) and river water depth (cm) along three transects set perpendicular to the water flow at 20-m intervals over the 100-m long river reach surveyed in each sampling site.
Data analyses
Analyses were done in R (R Core Team, 2016) using the packages and functions outlined below. Four diversity measures, species richness and the indices of Shannon–Wiener (Shannon, 1948), Simpson’s diversity (Simpson, 1949) and Pielou’s evenness (Pielou, 1966), were calculated for native fish and amphibians separately. For amphibians, we also calculated the Chao richness estimates to account for the possibility that more species than those captured may be present because of the logistic limitations of our sampling design for amphibians. Nonparametric species estimators, such as Chao, estimate the probable number of species from species-abundance curves by using the number of species recorded just one time (singletons) or twice (doubletons) in a data set (Colwell & Coddington, 1994).
The three diversity indices (Shannon–Wiener, Simpson and Pielou) accounted for the number of species but treat relative abundances differently. The Shannon–Wiener index weights least abundant species most heavily, while Simpson’s diversity index emphasizes the most abundant species (Pielou, 1975; Moreno, 2001). Pielou’s index was a ratio between the Shannon–Wiener diversity and the natural logarithm of the maximum number of species and is a measure of the evenness of abundances among species (Moreno, 2001).
Description of the associations between fish and amphibian species
We recorded the number of sampling sites fishless and within the distributional range of each fish species at which amphibians were present (P, 1) or absent (A, 0). An analysis of similarities (ANOSIM; Clarke, 1993) tested statistically whether fish species composition using P/A data differed between sites with and without amphibians; we used 999 permutations. Spearman’s ρ correlation coefficients also were used to determine the associations among the abundances of the different species of amphibians and fish.
Data transformation and collinearity among explanatory variables
Visual inspection of the distribution of all predictor variables showed that continuous variables were right-skewed, and so, log-transformations were undertaken. Spearman’s ρ correlation coefficients were used to identify potential collinearity among predictors. Temperature was excluded because of its high correlation with elevation (Spearman’s ρ = − 0.85). The retained 15 environmental variables (Appendix S2) also had variation inflation factors (VIF) < 2, so that collinearity should be of little statistical concern (Zuur et al., 2010). All predictors (Table 1) were standardized (0 mean, unit variance), so that all predictors were on a comparable scale (i.e. values of each predictor are expressed in units of the standard deviation of the predictor).
The associations of alien fishes with native fish and with the five species of amphibians were summarized in the variables alien fish richness and abundance, both of which were highly correlated (Spearman’s ρ > 0.90; Maceda-Veiga et al., 2017). We retained alien fish richness rather than alien fish abundance because the latter had a highly skewed distribution (i.e. alien fish were absent in many sites but very abundant in others). Moreover, the use of alien fish richness rather than transformed abundance data (e.g. log, presence/absence) led to better model fits. The same rationale applies to the use of native fish richness as a potential predictor to explore the effects of native fish in amphibian models.
Overview of generalized linear models
We constructed generalized linear mixed models (GLMMs) to identify the riverine conditions that are likely to be important for predicting the diversity of native fish and that of five species of amphibians. Models had two steps for amphibians because of the highly skewed distribution (Appendix S3). We modelled the: (1) occurrence of amphibians; and (2) variation in the five diversity-measures only for those sites where amphibians were present. The Poisson error distribution was used for the observed richness and Chao estimates, the Gaussian for Shannon diversity, the binomial distribution for proportional data and Bernoulli distribution for occurrence. Predictors were collectively referred to as sets (i.e. geography, physical habitat, in-stream hydraulics, water quality and fish, Tables 2 and 3) to identify factors upon which environmental management for biodiversity can be implemented jointly. However, all predictors within each set of predictors were included individually in the models (Table 2).
Nutrient concentrations were aggregated based on their likely toxic severity (Noga, 2011), such as ammonium with nitrite (TN) and nitrate with phosphate–P (NP). An offset term was used in all GLMMs to account for the differences in the number of individuals captured in each sampling site because this can adversely affect diversity estimates (Moreno, 2001). All models included random factors (collectively referred to as ρ) for year and river basin, which accounted for within-year and within-basin variation (e.g. multiple sites surveyed in a basin).
Which river conditions best predict native diversity measures of fish and amphibians?
We used the function glmer in the lme4 package (Bates et al., 2015) for multimodel inference using the Akaike’s Information Criterion (AIC) to compare models. We ranked models with the random factors for year and basin and different combinations of the five sets of predictors (Table 2). The most probable model was one with the lowest AIC value but models differing by ≤ 2 AIC units were considered equally plausible. Overall, model fit of the best model(s) was assessed using the function r.squaredGLMM (pseudo-R2) in the MuMIn package (Barton, 2013). The final best model had the lowest AIC and the highest pseudo-R2. Significance of the predictors in these final models was assessed using the function Anova in the car package (Fox et al., 2012) with P ≤ 0.05 to obtain the simplest and most informative model. Moran’s index (the package spdep in R, Bivand et al., 2011) indicated that spatial autocorrelation of model residuals was not likely to affect inferences (Moran values ranging from − 0.05 to 0.09; P > 0.2). Diagnostic plots of model residuals were used to check for statistical assumptions (e.g. normality, homogeneity of variances) following Zuur et al. (2010).
Results
Native fishes and amphibians were found in a wide range of environmental conditions and diversities of alien fishes (Table 1). Amphibians were mostly captured in streams with Strahler orders ≤ 4 (Fig. 2). We caught 34 fish species, of which 14 native and 14 alien species each occurred in > 10 of the 535 sites surveyed (see also Appendix S4). The 14 alien fishes included omnivores (55% of the alien species, e.g. Alburnus alburnus Linnaeus 1758, C. carpio) and invertivores that as adults feed upon small vertebrates (45% of the alien species, e.g. Lepomis gibbosus (Linnaeus, 1758), Silurus glanis Linnaeus, 1758). We caught larvae of five species of amphibians (Alytes obstetricans; Salamandra salamandra; Bufo spinosus Daudin, 1803, Rana temporaria Linnaeus, 1758 and Pelophylax perezi, with P. perezi, A. obstetricans and S. salamandra accounting for most records (70%).

Percentages of sampling sites, classified by Strahler stream order (1, N = 149; 2, N = 133; 3, N = 145; 4, N = 57; 5, N = 36; 6, N = 5), at which amphibians were detected. Amphibians were not found at Strahler stream order numbers > 6 (7, N = 5; and 8, N = 4)
Co-occurrence patterns of fish and amphibians in reaches
Fish and amphibians were observed to co-occur in 30% of the 535 sampling sites, 17% of sites had neither fish nor amphibians recorded, 3% were fishless but amphibians were present, and 63% did not have amphibians but had fish. Amphibians were not reported in ≥ 50% of the distributional ranges of each of the 34 fish species (Fig. 3). There was a low congruence among the diversities of native fish and amphibians (Richness, Spearman’s ρ = 0.14; Shannon Diversity, Spearman’s ρ = 0.08; Simpson Diversity, Spearman’s ρ = − 0.10; Pielou’s evenness, Spearman’s ρ = − 0.03). Fish-species composition differed slightly (albeit statistically significantly) between sites with and without amphibians (ANOSIM R = 0.03, P = 0.04; Fig. 3). The highest percentages of co-occurrence between fish and amphibians (≥ 36%) were for the native fish Squalius laietanus Doadrio, Kottelat & de Sostoa, 2007, Achondrostoma arcasii Steindachner, 1966, Gasterosteus aculeatus Linnaeus, 1758 and Barbus meridionalis Risso, 1826, and for the alien fish Rutilus rutilus Linnaeus, 1758. All correlation coefficients between the abundances of fish and the five species of amphibians were low, with seven fish species having the highest Spearman’s ρ (i.e. ≤ 0.10 Appendix S5). The fish species with positive associations with any of the five species of amphibians were A. arcasii, S. laietanus, G. aculeatus, R. rutilus and Salaria fluviatilis Asso, 1801 (Appendix S5).

Percentages of sampling sites at which amphibians were present and absent for each native (top panel) and alien (bottom panel) fish species in north-eastern Spain. The total number of sampling sites at which each fish species was captured is shown in brackets (N)
Commonalities of responses of native fish and amphibians to riverine conditions
The explanatory power of the models that included any riverine condition(s) as important predictor(s) ranged from 0.10 to 0.35 (Table 3). The null models (i.e. just intercept and random factors) were the best models for the Pielou and Simpson indices for native fish and amphibians (Table 3), so that neither index was sensitive to riverine predictors. The null model was the best model for the estimated amphibian richness (Table 3). We only studied main effects of environmental and fish predictors on amphibians because we did not find ‘true’ replicates for testing interactive effects.
For native fish, species richness and Shannon diversity had significant associations with five and four predictors, respectively, but the models shared only Strahler stream order number (Ord) and water velocity (Vel) (Table 3; Fig. 4). Richness of native fish was associated positively with alien fish richness (Alienr; Table 3, Fig. 4).

Standardized regression-coefficient estimates (i.e. effect size) and 95% confidence limits for the generalized linear mixed models in native fish and amphibians. Only those riverine conditions significant at P < 0.05 are shown; all models included random effects. Note that abscissa is different for native fish and amphibians to facilitate visualization of the associations with predictors. Alien richness refers to alien fish richness (Alienr), which was highly correlated with alien fish abundance (Aliena). Strahler order is Strahler stream order number, which is used as proxy for river size (see Table 1 for further information on predictors)
The best model for amphibian occurrence (pseudo-R2 = 0.26) only had one significant association, which was a negative relationship with river-water depth (Wad; Table 3, Fig. 4). The model for variation in amphibian richness (pseudo-R2 = 0.10, Table 3, Fig. 4) only had a significant, although weak, negative association with Alienr. This was also the case of amphibian Shannon diversity (pseudo-R2 = 0.20, Table 3, Fig. 4). No model identified native fish richness as important factor to explain variation in amphibian diversity (Table 3, Fig. 4).
Discussion
Fish were observed to co-occur with ≥ 1 amphibian species in 160 of the 535 sampling sites (30%), which was 25% of the sampling sites, on average, within the distributional ranges of individual native fish species. Alien fish richness, which was correlated strongly with alien fish abundance, may influence the diversity of native fish and amphibians. However, the association between total native fish and amphibian diversity was not significant, notwithstanding the prevailing idea that fish have an antagonistic relationship with amphibians (Kats & Ferrer, 2003; Denoël et al., 2005; Porej & Hetherington, 2005). Results should be interpreted with caution owing to relatively low model fits (pseudo-R2s < 0.36), which possibly are due to rivers being highly dynamic, fish being highly mobile and patchily distributed, and that just part of amphibians’ life-cycles is in water. Such levels of explained variance are common in complex ecological models at broad scales but still provide important insights (e.g. Low-Décarie et al., 2014; Russildi et al., 2016). The best model for fish and amphibians differed by just 0.08 in explained variance, so that there was similar confidence for both taxa in their associations with riverine conditions.
Associations between fish and amphibian species in rivers
Most native and alien fish species co-occurred with ≥ 1 of the five species of amphibians (A. obstetricans, S. salamandra, B. spinosus, P. perezi and R. temporaria), but only the abundances of the first four amphibians were positively associated (albeit weakly) with fish (i.e. A. arcasii, S. laietanus, G. aculeatus, R. rutilus and S. fluviatilis). All fish species are potential predators with amphibians because fish often prey on their eggs, larvae or cause injuries in amphibian larvae (e.g. Kats & Ferrer, 2003; Manenti & Pennanti, 2016). However, local adaptation of amphibians to fish competition or predation has been reported (Hecnar & M‘Closkey, 1997), and the stated five fish species are native in the study area (Aparicio et al., 2000; Maceda-Veiga et al., 2017). While we cannot make strong inferences about the mechanisms underlying the positive association between fish and amphibians, it might be the result of indirect effects, such as fish predation on other amphibian competitors (e.g. dragonfly larvae) or fish being an alternative prey for predators of amphibians (e.g. Werner & McPeek, 1994). Tadpoles of common toad (Bufo spinosus) are toxic to fish (Manteifel & Reshetnikov, 2002), so that native fish might ignore them as prey. Significant associations between fish and amphibians mostly occurred between species of sites at low to medium elevations, either for fish (Doadrio, 2011) or amphibians (Llorente et al., 1995), which may reflect that the median elevation of the study area was 405 m.a.s.l.
The associations found between fish and amphibians are consistent with the observation that most frequently captured fish were omnivorous-invertivorous species, apart from S. trutta, which preys upon small vertebrates including even adult amphibians (Gillespie, 2001). Alien fish piscivores have been introduced into north-eastern Spain (e.g. Sander lucioperca Linnaeus, 1758, Esox lucius Linnaeus, 1758, Silurus glanis) but were rare in our samples. These predators mostly occur in reservoirs and in mainstem large rivers (Doadrio, 2011), neither of which were surveyed because of the different sampling equipment and methods needed (e.g. fishing boats). This is not to say that alien omnivorous fish do not affect amphibians because the alien carp Cyprinus carpio profoundly alters habitats, including the destruction of submerged vegetation (Bajer et al., 2009) often used by amphibians for shelter and for breeding (Llorente et al., 1995). Frequently translocated native fish species (e.g. Luciobarbus graellsii (Steindachner, 1866), Parachondrostoma miegii (Steindachner, 1866)) may compete with amphibians for food, but this has not yet been investigated. This is an important topic for future research because translocated native species often are the only non-local species and often dominate the ‘native’ fish assemblages when present in the study area (Maceda-Veiga, 2013).
Can native fish and amphibian diversities be jointly managed?
Results for models on fish and amphibian diversity were probably driven by the most frequently captured fish (e.g. S. trutta, Barbus haasi, B. meridionalis, Gobio lozanoi Doadrio & Madeira, 2004; Appendix S1) and amphibian species (P. perezi, A. obstetricans and S. salamandra) and the unavoidable limitations of snap-shot surveys. Most of the studied fish species are sedentary (e.g. Aparicio & de Sostoa, 1999; Doadrio, 2011), and so, results are unlikely to be much affected by fish phenology. However, we captured five amphibian species (i.e. those with long aquatic stages) from a pool of 16 species known to occur in temporary or permanent waters in north-eastern Spain (Llorente et al., 1995). We used Chao richness estimates to overcome the sampling limitation and the range values of estimated and observed amphibian richness were not inconsistent. Therefore, our study is likely to be a reasonable first step to understanding the potential role of rivers as habitat for amphibians in north-eastern Spain, even though we were unable to conduct multiple visits to each sampling location due to costs and logistics.
Overall, there was a poor congruence among native fish and amphibian diversity, supporting previous conclusions that no single taxonomic group can be used as a surrogate for all others in conservation planning (Mac Nally et al., 2002; Heino, 2010; Guareschi et al., 2015; Maceda-Veiga et al., 2017). Amphibians may prefer to breed in pond-like habitats, such as those on the riverbank, where their larvae might experience reduced competition from, and predation by, fish and where the larvae are less likely to be swept away by river current (AMV pers. observ.). Fish often occupy deeper waters in Mediterranean rivers, including species of Cyprinidae (Aparicio & de Sostoa, 1999; Doadrio, 2011), the dominant fish family in Spain and elsewhere in Europe (Kottelat & Freyhof, 2007).
Native-fish and amphibian models for diversity measures (e.g. richness, Shannon) had different predictors while diversity components often did not correlate even within the same taxon (reviewed by Heino, 2010). Diversity models did not much improve in fit compared to richness-based models (Table 3), which may explain the main use of richness as the predominant diversity measure (Gaston, 1996; Moreno, 2001; Heino, 2010). The Shannon index, which gives more weight to the rare species compared to those of Simpson and Pielov (Moreno, 2001), was the only index that highlighted riverine conditions as significant predictors. This finding suggests that the least frequent species in the data-set might have more limited environmental requirements than the more widely distributed species. Nevertheless, our inferences need further testing by comparing the riverine requirements among amphibian species using more comprehensive amphibian surveys in rivers to reduce the number of probable false absences, which prevent accurate species-specific modelling.
How organisms respond to environmental conditions is related to body size and life history (Allen et al., 1999; Heino, 2010; Velghe & Gregory-Eaves, 2013). Fish are necessarily more likely to be affected by riverine conditions, but patterns for larval amphibians depend on responses of both aquatic larvae and terrestrial adults, which may differ. Our findings suggest that native fish would benefit from restoring water and physical habitat quality (see also Maceda-Veiga et al., 2017). Although similar benefits might be expected for amphibians (e.g. Hartel et al., 2007; Egea-Serrano et al., 2012; Polo-Cavia et al., 2016), we found no evidence of associations. Moreover, little is known about the relative importance of rivers for the conservation of amphibian populations compared to other habitats (e.g. temporary pools, ponds, ephemeral streams) in the Mediterranean-climate region (see de Vries & Marco, 2017).
Our deductions may be at least partially occluded by general patterns in which alien fish tend to occupy larger streams and rivers (high Strahler stream order numbers) while amphibians mostly occur in shallow waters. Large rivers are the most altered by human impacts (Aparicio et al., 2000; Tockner et al., 2009; Pracheil et al., 2013), which make it difficult for correlative studies, such as ours, to distinguish between the effects of environmental degradation and those of alien fish species. Moreover, our findings may be for a transitional stage in the fish–amphibian association due to natural fluctuations in environmental conditions and biological communities. This includes ongoing changes in the diversity and abundances of alien species and their effects (Parker et al., 1999), which might alter the positive association we found between alien and native fish diversity. Smaller rivers, such as tributaries, have the lowest diversities and abundances of alien fish species in the study region (Maceda-Veiga et al., 2017), so that small streams might be treated as priority areas in the context of whole-catchment conservation management (see also Clarke et al., 2008; Pracheil et al., 2013).
References
Allen, A. P., T. R. Whittier, P. R. Kaufmann, D. P. Larsen, R. J. O’Connor, R. M. Hughes, R. S. Stemberger, S. S. Dixit, R. O. Brinkhurt, A. T. Herlihy & S. G. Paulsen, 1999. Concordance of taxonomic richness patterns across multiple assemblages in lakes of the northeastern United States. Canadian Journal of Fisheries and Aquatic Sciences 56: 739–754.
Alroy, J., 2015. Current extinction rates of reptiles and amphibians. Proceedings of the National Academy of Sciences USA 112: 13003–13008.
Aparicio, E. & A. de Sostoa, 1999. Pattern of movements of adult Barbus haasi in a small Mediterranean stream. Journal of Fish Biology 55: 1086–1095.
Aparicio, E., M. J. Vargas, J. M. Olmo & A. de Sostoa, 2000. Decline of native freshwater fishes in a Mediterranean watershed on the Iberian Peninsula: a quantitative assessment. Environmental Biology of Fishes 59: 11–19.
Bajer, P. G., G. Sullivan & P. W. Sorensen, 2009. Effects of a rapidly increasing population of common carp on vegetative cover and waterfowl in a recently restored Midwestern shallow lake. Hydrobiologia 632: 235–245.
Barbour, M. T., J. Gerritsen, B. D. Snyder & J. B. Stribling, 1999. Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish. US Environmental Protection Agency, Office of Water, Washington, DC.
Barton, K., 2013. MuMIn: multi-model inference. R package ver. 1(13): 6.
Bates, D., M. Maechler, B. Bolker & S. Walker, 2015. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software 67: 1–48.
Bivand, R., M. Anselin, L. Anselin, R. Assuncao, O. Berke & A. Bernat, 2011. spdep: Spatial dependence: weighting schemes, statistics and models.
Butchart, S. H., M. Walpole, B. Collen, A. Van Strien, J. P. Scharlemann, R. E. Almond, et al., 2010. Global biodiversity: indicators of recent declines. Science 328: 1164.
Camargo, J. A. & Á. Alonso, 2006. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: a global assessment. Environment International 32: 831–849.
Clarke, K. R., 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18: 117–143.
Clarke, A., R. Mac Nally, N. Bond & P. S. Lake, 2008. Macroinvertebrate diversity in headwater streams: a review. Freshwater Biology 53: 1707–1721.
Colin, N., A. Maceda-Veiga, N. Flor-Arnau, J. Mora, P. Fortuño, C. Vieira, N. Prat, J. Cambra & A. de Sostoa, 2016. Ecological impact and recovery of a Mediterranean river after receiving the effluent from a textile dyeing industry. Ecotoxicology and Environmental Safety 132: 295–303.
Colwell, R. K. & J. A. Coddington, 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London Series B 345: 101–118.
de Vries, W. & A. Marco, 2017. The importance of fluvial habitats for amphibian conservation in the Mediterranean climate. Basic and Applied Herpetology 31: 5–16.
Denoël, M., G. Dzukic & M. L. Kalezic, 2005. Effects of widespread fish introductions on paedomorphic newts in Europe. Conservation Biology 19: 162–170.
Diehl, S., 1992. Fish predation and benthic community structure: the role of omnivory and habitat complexity. Ecology 73: 1646–1661.
Dirzo, R., H. S. Young, M. Galetti, G. Ceballos, N. J. Isaac & B. Collen, 2014. Defaunation in the Anthropocene. Science 345: 401–406.
Doadrio, I. (ed.), 2011. Ictiofauna española. Bases para su seguimiento. Ministerio de Agricultura y Pesca, Madrid. (In Spanish).
Egea-Serrano, A., R. A. Relyea, M. Tejedo & M. Torralva, 2012. Understanding of the impact of chemicals on amphibians: a meta-analytic review. Ecology and Evolution 2: 1382–1397.
Fox, J., S. Weisberg, D. Adler, D. Bates, G. Baud-Bovy, S. Ellison, et al., 2012. Package ‘car’. R Foundation for Statistical Computing, Vienna.
Gaston, K. J., 1996. Species richness: measure and measurement. In Gaston, K. J. (ed.), Biodiversity, a biology of numbers and difference. Blackwell, Cambridge: 77–113.
Gaston, K. J., 2000. Global patterns in biodiversity. Nature 405(6783): 220.
Gillespie, G. R., 2001. The role of introduced trout in the decline of the spotted tree frog (Litoria spenceri) in south-eastern Australia. Biological Conservation 100: 187–198.
Guareschi, S., P. Abellán, A. Laini, A. J. Green, J. A. Sánchez-Zapata, J. Velasco & A. Millán, 2015. Cross-taxon congruence in wetlands: assessing the value of waterbirds as surrogates of macroinvertebrate biodiversity in Mediterranean Ramsar sites. Ecological Indicators 49: 204–215.
Hartel, T., S. Nemes, D. Cogălniceanu, K. Öllerer, O. Schweiger, C. I. Moga & L. Demeter, 2007. The effect of fish and aquatic habitat complexity on amphibians. Hydrobiologia 583: 173.
Hecnar, S. J. & R. T. M‘Closkey, 1997. The effects of predatory fish on amphibian species richness and distribution. Biological Conservation 79: 123–131.
Heino, J., 2010. Are indicator groups and cross-taxon congruence useful for predicting biodiversity in aquatic ecosystems? Ecological Indicators 10: 112–117.
Heino, J., R. Paavola, R. Virtanen & T. Muotka, 2005. Searching for biodiversity indicators in running waters: do bryophytes, macroinvertebrates, and fish show congruent diversity patterns? Biodiversity & Conservation 14: 415–428.
Kats, L. B. & R. P. Ferrer, 2003. Alien predators and amphibian declines: review of two decades of science and the transition to conservation. Diversity and Distributions 9: 99–110.
Kefford, B. J., G. L. Hickey, A. Gasith, E. Ben-David, J. E. Dunlop, C. G. Palmer, K. Allan, S. C. Choy & C. Piscart, 2012. Global scale variation in the salinity sensitivity of riverine macroinvertebrates: eastern Australia, France,Israel and South Africa. PLoS ONE 7: e35224.
Llorente, G. A., A. Montori, X. Santos & M. A. Carretero, 1995. Atlas dels amfibis i rèptils de Catalunya i Andorra. El Brau, Figueres.
Mac Nally, R., A. F. Bennett, G. W. Brown, L. F. Lumsden, A. Yen, S. Hinkley, P. Lillywhite & D. Ward, 2002. How well do ecosystem-based planning units represent different components of biodiversity? Ecological Applications 12: 900–912.
Maceda-Veiga, A., 2013. Towards the conservation of freshwater fish: Iberian Rivers as an example of threats and management practices. Reviews in Fish Biology and Fisheries 23: 1–22.
Maceda-Veiga, A., A. Baselga, R. Sousa, M. Vilà, I. Doadrio & A. de Sostoa, 2017. Fine-scale determinants of conservation value of river reaches in a hotspot of native and non-native species diversity. Science of the Total Environment 574: 455–466.
Maceda-Veiga, A., R. Mac Nally & A. de Sostoa, 2018. Water-quality impacts in semi-arid regions: can natural ‘green filters’ mitigate adverse effects on fish assemblages? Water Research 144: 628–641.
Magalhães, M. F., P. Beja, I. J. Schlosser & M. J. Collares-Pereira, 2007. Effects of multi-year droughts on fish assemblages of seasonally drying Mediterranean streams. Freshwater Biology 52: 1494–1510.
Manenti, R. & R. Pennati, 2016. Environmental factors associated with amphibian breeding in streams and springs: effects of habitat and fish occurrence. Amphibia-Reptilia 37: 237–242.
Manteifel, Y. B. & A. N. Reshetnikov, 2002. Avoidance of noxious tadpole prey by fish and invertebrate predators: adaptivity of a chemical defence may depend on predator feeding habits. Archiv fur Hydrobiologie 153: 657–668.
Moreno, C. E., 2001. Métodos para medir la biodiversidad. M&T–Manuales y Tesis SEA, vol. 1. Zaragoza, 84 pp (In Spanish).
Munné, A., N. Prat, C. Sola, N. Bonada & M. Rieradevall, 2003. A simple field method for assessing the ecological quality of riparian habitat in rivers and streams: QBR index. Aquatic Conservation: Marine and Freshwater Ecosystems 13: 147–163.
Noga, E. J., 2011. Fish disease: diagnosis and treatment. Wiley, Hoboken.
Parker, I. M., D. Simberloff, W. M. Lonsdale, K. Goodell, M. Wonham, P. M. Kareiva, M. H. Williamson, B. Von Holle, P. B. Moyle, J. E. Byers & L. Goldwasser, 1999. Impact: toward a framework for understanding the ecological effects of invaders. Biological Invasions 1: 3–19.
Pielou, E. C., 1966. The measurement of diversity in different types of biological collections. Journal of Theoretical Biology 13: 131–144.
Pielou, E. C., 1975. Ecological diversity. Wiley, New York.
Polo-Cavia, N., P. Burraco & I. Gomez-Mestre, 2016. Low levels of chemical anthropogenic pollution may threaten amphibians by impairing predator recognition. Aquatic Toxicology 172: 30–35.
Porej, D. & T. E. Hetherington, 2005. Designing wetlands for amphibians: the importance of predatory fish and shallow littoral zones in structuring of amphibian communities. Wetlands Ecology and Management 13: 445–455.
Pracheil, B. M., P. B. McIntyre & J. D. Lyons, 2013. Enhancing conservation of large-river biodiversity by accounting for tributaries. Frontiers in Ecology and the Environment 11: 124–128.
R Core Team, 2016. R: a language and environment for statistical computing (https://www.R-project.org/). R Foundation for Statistical Computing, Vienna, Austria.
Ricciardi, A., T. M. Blackburn, J. T. Carlton, J. T. Dick, P. E. Hulme, J. C. Iacarella, J. M. Jeschke, A. M. Leibhold, J. L. Lockwood, H. J. MacIsaac, P. Pysek, D. M. Richardson, G. M. Ruiz, D. Simberloff, W. J. Sutherland, D. A. Wardle & D. C. Aldrige, 2017. Invasion science: a horizon scan of emerging challenges and opportunities. Trends in Ecology & Evolution 32: 464–474.
Richter-Boix, A., G. A. Llorente & A. Montori, 2006. Breeding phenology of an amphibian community in a Mediterranean area. Amphibia-Reptilia 27: 549–559.
Russildi, G., V. Arroyo-Rodríguez, O. Hernández-Ordóñez, E. Pineda & V. H. Reynoso, 2016. Species-and community-level responses to habitat spatial changes in fragmented rainforests: assessing compensatory dynamics in amphibians and reptiles. Biodiversity and Conservation 25: 375–392.
Shannon, C. E., 1948. A mathematical theory of communication. Bell System Technical Journal 27: 379–423.
Sillero, N., J. A. Campos, C. Bonardi, R. Corti, P. Creemers, J. Crochet, M. Crnobrnja Isailovic, G. F. Denoël, J. Ficetola, S. Gonçalves, P. Kuzmin, P. Lymberakis, A. de Pous, R. Rodríguez, J. Sindaco, B. Speybroeck, B. Toxopeus, D. R. Vieites & M. Vences, 2014. Updated distribution and biogeography of amphibians and reptiles of Europe. Amphibia-Reptilia 35: 1–31.
Simpson, E. H., 1949. Measurement of diversity. Nature 163: 688.
Spellerberg, I. F., 1991. Monitoring ecological change. Cambridge University Press, Cambridge: 334.
Strahler, A. N., 1964. Quantitative geomorphology of drainage basin and channel networks. Handbook of Applied Hydrology. McGraw-Hill, New York.
Tisseuil, C., J. F. Cornu, O. Beauchard, S. Brosse, W. Darwall, R. Holland, B. Hugueny, P. A. Tedesco & T. Oberdorff, 2013. Global diversity patterns and cross-taxa convergence in freshwater systems. Journal of Animal Ecology 82: 365–376.
Tockner, K., U. Uehlinger & C. T. Robinson, 2009. Rivers of Europe. Academic Press, Cambridge.
Townsend, C. R., S. S. Uhlmann & C. D. Matthaei, 2008. Individual and combined responses of stream ecosystems to multiple stressors. Journal of Applied Ecology 45: 1810–1819.
Valiente-Banuet, A., M. A. Aizen, J. M. Alcántara, J. Arroyo, A. Cocucci, M. Galetti, M. B. García, D. García, J. M. Gómez, P. Jordano, R. Mendel, L. Navarro, J. R. Obeso, R. Oviedo, N. Ramírez, P. J. Rey, A. Traveset, M. Verdú & R. Zamora, 2015. Beyond species loss: the extinction of ecological interactions in a changing world. Functional Ecology 29: 299–307.
Velghe, K. & I. Gregory-Eaves, 2013. Body size is a significant predictor of congruency in species richness patterns: a meta-analysis of aquatic studies. PLoS ONE 8: e57019.
Vörösmarty, C. J., P. B. McIntyre, M. O. Gessner, D. Dudgeon, A. Prusevich, P. Green, et al., 2010. Global threats to human water security and river biodiversity. Nature 467: 555.
Waldron, A., D. C. Miller, D. Redding, A. Mooers, T. S. Kuhn, N. Nibbelink & J. L. Gittleman, 2017. Reductions in global biodiversity loss predicted from conservation spending. Nature 551: 364.
Zuur, A. F., E. N. Ieno & C. S. Elphick, 2010. A protocol for data exploration to avoid common statistical problems. Methods in Ecology and Evolution 1: 3–14.
Acknowledgements
We thank three anonymous reviewers and the associated editor Katya Kovalenko for their helpful suggestions. We are grateful to all people who assisted in the field and Mari Piñero-Rodríguez for acting as catalyst for this manuscript. River surveys were funded by Natural Parks (Collserola and Sant Llorenç del Munt i Serra de l’Obac), the town of Sabadell and the Spanish water authorities ‘Agència Catalana de l’Aigua’ (A.C.A.) and ‘Confederación Hidrográfica del Ebro’ (C.H.E.).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Katya E. Kovalenko
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Maceda-Veiga, A., Mac Nally, R. & de Sostoa, A. Congruence in riverine conditions and associations between native fish and several species of amphibians in a region prone to fish invasions. Hydrobiologia 836, 109–122 (2019). https://doi.org/10.1007/s10750-019-3945-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-019-3945-4