
Abstract
Perch Perca fluviatilis is a widespread predator in European reservoirs, frequent in open waters but also known to spend a lot of time in the littoral zones. To get insight into how adult perch used and selected their habitat in an environment subject to water-level (WL) fluctuations, 21 perch were continuously tracked using acoustic telemetry over 2 years in the Bariousses Reservoir (France). The different available habitats were characterized by depth classes and substrate types, presence of emerging trees and presence of tree stumps in the littoral zone. We showed that perch habitat preferences were strongly dependent on the season, except for substrate type, and in line with their habitat use. Surprisingly we did not find any influence of the WL which, however, reduced the structural complexity of the littoral zone when lowering. In spring and summer, whatever the WL, we observed a strong preference for the littoral zone and complex habitats. In autumn and winter, perch migrated into deeper waters. However, the individual variability of the habitat preferences was quite high. This type of research helps to understand the spatial ecology of fish and provides useful guidance to hydromorphological restoration for fish populations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Whatever their origin, lake littoral zones are transitional areas between terrestrial and aquatic habitats that generally host a high biodiversity (Schmieder, 2004). They provide resources available nowhere else in the lacustrine ecosystem (Zohary & Gasith, 2014) and most lake fishes use them during their life cycle (Winfield, 2004). Littoral habitats can also be composed of complex structures (e.g. rocks, woody debris, vegetation) that provide both spawning grounds and refuges to a diverse assemblage of microorganisms, algae, invertebrates (Gasith & Gafny, 1998) as well as fish larvae and juveniles (Diehl, 1993; Stoll et al., 2008).
Littoral habitats are highly influenced by water-level fluctuations (WLFs) which can be particularly pervasive in lentic systems exploited by humans for water use and hydropower. Habitat complexity changes with water level (WL), particularly in the littoral zone (Zohary & Ostrovsky, 2011). For instance, high WLF (frequent and wide) may engender coarser littoral substrate with less macrophyte coverage in the shallow zones of lakes (Evtimova & Donohue, 2016), which in turn reduce the productivity and biodiversity of these areas (Wetzel, 1990; Evtimova & Donohue, 2014). More specifically, studies have shown that WLF can affect various ecological aspects of fish species such as their growth, distribution and behaviour (e.g. Gaboury & Patalas, 1984; Rogers & Bergersen, 1995; Fischer & Ohl, 2005; Sutela & Vehanen, 2008; Logez et al., 2016). Indeed, WLF can lead to loss of potential prey (Winfield, 2004), loss of refuge areas (Kaczka & Miranda, 2014) and loss of spawning habitats (Hudon et al., 2005) for fish fauna. Finally, WLF may also decrease fish recruitment if WL drops after spawning (Kahl et al., 2008) and fish eggs desiccate (Michaletz, 1997; Winfield, 2004). Therefore, Coops et al. (2003) and more recently Tao et al. (2016) highlighted the urgency of collecting ecological data on fish–habitat relationship. Fish habitat use is a key driver of population dynamics (Hayes et al., 2009) and an essential knowledge for predicting how populations will respond to management interventions (Koster et al., 2015; Cooke et al., 2016). In particular, the identification of crucial habitats is important for efficient aquatic conservation in areas with strong human influence (Halpern et al., 2005; Sale et al., 2005).
A thorough in situ study of WLF effects on the behaviour of a species has rarely been conducted and can contribute to valuable knowledge on management decisions, as Gardner et al. (2015) recently showed on the common bream (Abramis brama L.) which altered their home range size according to the WL while keeping the same activity level. In the present work, we propose to study habitat-use patterns of the Eurasian perch (Perca fluviatilis L.), a species present in most lowland reservoirs (Irz et al., 2006) and widespread in Europe (Kottelat & Freyhof, 2007) but on which the effects of WLF have never been investigated. Although frequent in open waters, their dependence on the littoral zone is high (Zamora & Moreno-Amich, 2002; Jacobsen et al., 2015). At night, they rest in the littoral zone on the bottom (Imbrock et al., 1996). They are highly selective in their choice of spawning habitat, generally preferring shallow and sheltered areas with rigid and structurally complex substrates (Probst et al., 2009; Snickars et al., 2010; Čech et al., 2012a) but can spawn in deeper waters depending on the environmental conditions (Čech et al., 2012a, b). Perch spawning usually takes place when water temperatures reach about 5°C (Souchon & Tissot, 2012).
Given that the littoral zone is a critical habitat for the perch, we designed the present study to evaluate whether and how WLF affects its habitat preferences, here defined as the higher likelihood that an individual chooses a habitat type if offered on an equal basis to others (Johnson, 1980). As WLF can modify the availability of the different littoral habitat types (Zohary & Ostrovsky, 2011; Evtimova & Donohue, 2016), we expected a shift of perch habitat preferences with WL. For example, when an habitat becomes very scarce, if perch keeps on frequently using it, then its preference will automatically raise; on the contrary, if perch switches to another habitat, its preference will decline for this scarce habitat and raise for the other. In particular, we could expect this to happen during the spawning season, in spring in our case, when the habitat choice appears crucial. To answer this question, we set up a high spatial and temporal resolution study over a French reservoir, subject to human-induced WLF ranging from days to seasons. More specifically, we tracked 21 large perch by acoustic telemetry over a 2-year period. We first evaluated how WLF affected the relative availability of the littoral and pelagic areas in the reservoir. Then, we assessed the effects of WLF and seasons on individual habitat preference. Lastly, perch habitat use and preferences were analyzed in detail.
Materials and methods
Study site
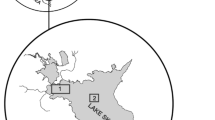
The study was conducted in the Bariousses Hydropower Reservoir (45.33°N, 1.49°E) in the west central part of France (Fig. 1). The reservoir is located in a rural and natural environment, in a forest land cover-dominated catchment with low anthropogenic activities (Logez et al., 2016). This reservoir, with a 229 km2 watershed, is an impoundment of the Vezere River. It is the second dam on this river and it is located 42 km downstream the source; the average flow is 4.37 m3/s at Bugeat, 20 km upstream from the dam. The main inflow to Bariousses Lake comes from deep waters of the first dam located 12 km upstream. As these waters are colder than those of the lake, especially in spring and summer, they probably flow at the bottom of the lake along the original river track. At the mean WL, altitude of 511.5 m, its area covers 86.6 ha, mean depth is 7.1 m and maximum depth reaches 19.4 m. The bathymetry of the reservoir was measured with a multibeam sounder [Electricité de France (EDF), personal source], giving a 2 × 2-m resolution GIS map. The reservoir mean renewal time is 12 days.

Location of the study site on the inset map of France and bathymetric map of the reservoir with the location of the receivers and synchronizing tags
The thermal regime of the reservoir is monomictic with four distinct temperature regimes. In spring (April–June), the water temperatures raise rapidly and the stratification is taking place; in summer (July–September), waters are warmer and stratified and the thermocline about 4.5 m deep; the autumn (October–December) corresponds to a rapid decrease of water temperatures when the mixing is taking place and destratification is in progress and, in winter (January–March), the waters are homogeneously cold over the whole water column (Table 1). The summer thermocline is associated with an oxycline that separates saturated surface waters from deep waters; the deep layer has an oxygen saturation rate of 40%. These regimes can be linked with perch ecology. The broad range of optimum temperatures for perch spawning lies between 5 and 19°C, depending on the region, and the limited one between 8 and 15°C (Souchon & Tissot, 2012). Rising temperatures appeared to be the major factor inducing spawning (Hokanson, 1977; Thorpe, 1977; Craig, 2000) which suites well with spring. In addition, in the Bariousses Reservoir, we observed some perch eggs laying on the shore in April and perch egg ribbons were also usually seen by regular anglers in early May in very shallow zones dewatered by the WLF. Perch activity was shown to raise with temperature (Craig, 1977) and to peak concomitantly with high summer temperatures (Jacobsen et al., 2002).
Depending both on human energy needs and hydrology, WLFs are very variable in the Bariousses Reservoir (Fig. 2). Over the study period (June 2012–March 2014), the hourly WL, measured by EDF, ranged from 507.1 to 513.5 m. The tertiles of the hourly WL distribution over the study period were used to split WL into low, mean and high WL. The mean lake area is 78.1, 86.6 and 90.5 ha, respectively, at low, mean and high WL. Typically, WL can shift from one class to a neighbouring class in a few days, though main annual features emerge. In spring, the high WL is by far the most frequent, whereas the low one is in autumn because, at the beginning of this season, the WL is usually lowered in anticipation of rains (Logez et al., 2016). In winter, the occurrence of the three WL classes is more evenly distributed. In summer, the WL is kept stable around its mean value 95% of the time to sustain recreational activities which are concentrated close to a sandy beach located on the west shore in front of the island (Fig. 1), motorboats being forbidden.

Hourly water level in the Bariousses Reservoir from 29 June 2012 to 10 March 2014. The solid (respectively dotted) black line corresponds to periods with (respectively without) fish positions. The dashed horizontal lines represent the first and second tertiles of this water-level distribution which were used to split water levels into low, mean and high
The water conductivity is low over the whole water column all over the year. The Secchi transparency depth lies between 1.3 and 2.5 m. The characterization of the phytoplankton community qualifies the reservoir as oligotrophic. Based on diversity, abundance and sensitivity to pollutants of invertebrates sampled in shallow waters, the reservoir appears in good condition (unpublished data).The fish fauna of the reservoir was determined with a standardized procedure (CEN, 2005) in 2010 and comprises 15 species. It is dominated by Cyprinids and Percids, characteristic of a lowland reservoir (Irz et al., 2002). The most dominant species, in terms of catch per unit effort, are roach (Rutilus rutilus), ruffe (Gymnocephalus cernua), Eurasian perch (P. fluviatilis), pikeperch (Sander lucioperca) and common bream (A. brama). Besides Eurasian perch and pikeperch, another dominant predator, pike (Esox lucius), is present. This community has been very little manipulated since a lake drainage in 1997. Fishing is allowed all along the year and only outside of spawning periods for pike and pikeperch.
Fish tagging
Following the suggestions of Thiem et al. (2011), below is detailed the followed surgical procedure. A total of 29 adult perch were caught with gillnets set at dawn, day and dusk for maximally 2 h over four sampling campaigns: 16 in spring 2012, 1 in summer 2012, 7 in autumn 2012 and 5 in spring 2013. Once captured, to check their condition, they spent 3–6 h in an aerated tank of lake water prior tagging, after a half an hour trip on the boat in another aerated tank to join the tagging site located in a building on the lake shore. Fishes were individually anaesthetized, which took 8–10 min, by immersion in a 20-l tank filled with an aerated solution composed of 90% diluted clove oil (0.03–0.05% in lake water) and 10% ethanol. Once the fish had lost its balance (ventral side up), did not respond to stimuli anymore and had a very slow and steady operculum rate with large amplitude, it was weighted, measured and placed ventral side up on a V-shaped surgical table. The same anaesthetic solution but less concentrated in clove oil (0.003%) was used to irrigate the gills during surgery. A 10–15-mm-long incision was made posterior to the pelvic girdle to insert an acoustic transmitter, previously sterilized in surgical spirit, in the peritoneal cavity. We used Vemco V9P-2L (47 mm long, 6.3 g in the air, 90-s mean burst interval, mean battery life 385 days) or V8-4L (20.5 mm long, 2 g in the air, 90-s mean burst interval, mean battery life 163 days) acoustic transmitters. The transmitter weight in the air did not exceed 2% of the fish body weight (Winter, 1983). The incision was closed using two to three simple surgical sutures (3-0 Polydioxanone Ethicon Monofilament Ltd.) placed 5 mm apart. An antiseptic and antibiotic cream (Fucidine 2%) was applied on the incision wound to help healing and limit the risk of infection. The surgical procedure took 5–6 min. The same person always carried out the surgery. Fishes were then put in an aerated recovery tank, where they were continually observed until the opercular activity, swimming ability, balance and behavioural response to stimuli became normal again, usually after 10 min. Then, in a couple of minutes, they were transferred to a net set in the lake where they spent 6–12 h. Lastly, they were transported by boat within half an hour in an aerated tank back to their capture site to be released.
Fish tracking
An array of 40 underwater VR2W 69-kHz omnidirectional acoustic receivers (Vemco) with their associated synchronization tag (V13-1L) was anchored at the bottom throughout the reservoir from January 2012 until March 2014 (Fig. 1). Eight additional synchronization tags were settled in the reservoir to detect anomalies in the tracking system. On average, neighbouring receivers were positioned 150 m from each other (range 72–223 m), 6 m deep (range 2–15 m) (Roy et al., 2014). Roughly every 6 months, receivers were removed from the lake to download fish detections. Fish positions were calculated by Vemco with their Vemco positioning system (VPS) algorithm (Smith, 2013). The horizontal position error, a dimensionless parameter calculated by the VPS for each position that gives information on the quality of the position estimate, was used to filter the dataset (Espinoza et al., 2011). In this study, we followed the recommendations of Roy et al. (2014) for the same system and only positions with horizontal position error not exceeding 15 were retained; this limit represented a good compromise between the mean position error (3.3 m throughout the reservoir) and the percentage of positions kept (79%) (see Roy et al., 2014 for detailed calculations of these error and percentage). Moreover, the probability of location map, estimated by Roy et al. (2014), showed that some parts of the lake were not well sampled, all located at the ends of the lake or on the shore. In parallel, in some of these areas very few locations were recorded. Not to introduce biases, we then removed from our study areas where the probability of location was below 2.5%; this threshold appeared as a good compromise between the representativeness of the sampling and the number of removed positions. So as not to include positions affected by behavioural modification following the surgery, only the positions recorded at least 2 days after release were included in the analyses (Bridger & Booth, 2003; Vehanen & Lahti, 2003). Some fish were already tagged in March 2012 but a problem of receiver memory saturation only fixed on late June 2012 caused the loss of data; the same problem caused the loss of data from August to October 2012. The downloading of receivers led to an interruption of the tracking for a few days in May 2013. In early October 2013, due to a sharp lowering of the WL for dam inspection, receivers were again downloaded and the experiment interrupted as the shallowest receivers could be dewatered; the interruption lasted till late November as the receivers could not have been put back into water sooner because of extended unavailability of divers and staff.
In the end, among the 29 perch initially tagged, 2 stationary fish were supposed to be dead rapidly after release, and 6 others, sparsely located from a few days to a few weeks, were then never located. These eight individuals were omitted from the analyses. Hence, 21 adult perch, 320–486 mm long (Table 1), corresponding to 12–18 individuals depending on the season, were followed. The time series of their positions used in this study are represented on Online Resource 1.
Data analysis
We first define several terms used in the following. The use of an habitat is the quantity that is utilized by perch; the availability of an habitat is the quantity accessible to the perch; the selection of an habitat is the process in which perch choose an habitat and preference is a reflection of the likelihood that perch choose an habitat if offered on an equal basis to others (Johnson, 1980). Habitat availability, use, selection and preference were evaluated at the reservoir scale as most of the individuals used the whole reservoir (Online Resource 2).
Habitat description and availability
Based on the Secchi transparency depth, ranging from 1.3 to 2.5 m, we defined the littoral area of the reservoir as the lake area connected to the bank with a depth lower than 2.5 m. The substrate types observed in the Bariousses were silt, sand, gravel, pebble, stone, boulder and rock; at high WL, lawn was also present thanks to flooded grasslands. Other available habitats were tree stumps, coming from tree felling at the time of the impounding, emerging trees, i.e. living shrubs or trees with roots and trunk in the water at least in some periods, helophytes and undercut banks. The last two categories were very scarcely present and thus not used further in the study. Thus, the different available habitats were characterized by four variables: depth (7 classes: [0, 2.5[, [2.5, 5[, [5, 7.5[, [7.5, 10[, [10, 12.5[, [12.5, 15[ and [15, 22[ m) and, in the littoral zone, main substrate, emerging trees and tree stumps.
All the habitats listed above were mapped on October 2013 by visual observation all around the lake when the WL was 507.5 m. Dewatered habitats between 513.5 and 507.5 m were described as well as those down to 506.5 m thanks to the water transparency. For each habitat variable, homogeneous polygonal areas were delimited with a differential GPS (Leica 1200®) and the habitat map was discretized in 10 × 10 m2 squares. In each grid square, for each habitat variable the habitat type which was selected was the one that covered more than 50% of the square. The mean square depth was used for depth. All habitats were mapped with ArcGis 10.0.
The relative availability of habitat type i, π i , was defined as the ratio of the number of grid squares with habitat type i on the total number C of 10 × 10 m2 grid squares in the study area (the whole lake if the considered habitat is depth, the littoral area for other habitat variables) at the considered WL. The different available habitats of the lake were quantified at low, mean and high WL. For each hourly WL of the study period and each habitat variable, the relative availability of each habitat type was estimated. Mean and standard deviation of these relative availabilities were calculated in grouping the data by the three WL classes.
Environmental variables impacting habitat preference
For each fish position, the used habitat corresponded to the habitat of the grid square in which the fish was. The number of positions in a grid square was corrected with the probability of location in this grid square, estimated by Roy et al. (2014), to minimize the spatial variability of the sampling inherent to a telemetry system. The proportion of each type was calculated for each habitat variable.
The corrected number of positions of fish j in habitat type i is then given by
where \(n_{ij}^{c}\) is the number of positions of fish j in the grid square c containing the habitat type i and p c is the probability for a tagged fish present in the grid square c to be located (see Roy et al., 2014 for more details).
The corrected number of positions of fish j in all classes of habitat type i (1,…,I) is
where I is the number of types in the considered habitat variable (for example, the “main substrate” habitat variable contains eight types).
The relative use of habitat type i by fish j is the ratio of u ij on u +j .
The selection ratios were then used to quantify the habitat preference (Manly et al., 2002). For individual j and habitat type i, the selection ratio is as follows:
For each habitat variable, the effects of habitat type, season, WL and two-way interactions on perch individual selection ratios were explored using generalized additive mixed-effects models (GAMMs; Zuur et al., 2009). The fish identity was considered as a random effect to explicitly account for individual heterogeneity. To take into account the skewed distribution of individual selection ratios towards zero, a Tweedie family function with a log-link was used (Gilman et al., 2012). As water temperature regimes were very similar over the 2-year study period (Table 1), data from the same seasons of different years were merged.
For one habitat variable, the full model could be written as follows:
where \(\overline{{{\text{SR}}_{\text{ind}} }}\) is the expected mean individual selection ratio, strictly positive, α is the overall intercept, HAB is the habitat variable (depth, main substrate, emerging trees, tree stumps) split into different types corresponding to the variable classes, SEASON is the season (four classes, Table 1), WL is the water level (three classes, Fig. 2), s(ind) is a smoothing function modelling the individual effects (Wood, 2008) having the advantage to get a significance test of these effects and an evaluation of the explained deviance of the model, and ε is the error term following a normal distribution with zero mean.
For each habitat variable, the most parsimonious simple model was selected by running a forward stepwise-based procedure (Venables & Ripley, 2002) and applying the recommendations of Richards (2008): all models having an AIC value within a range of 6 from the lowest AIC value were initially selected and, among them, the more complex models that did not have an AIC value lower than all the simpler models within which they were nested were removed.
The model fitting was assessed with regard to the homogeneity and normality of the residuals (Zuur et al., 2009) and to the percentage of explained deviance (Hastie & Tibshirani, 1990).
Habitat use and preference
For each of the habitat variables, habitat use and preference were explored according to those of the environmental variables that had significant effects in the GAMM.
The compositional analysis as proposed by Aebischer et al. (1993) was applied on each habitat variable to test for habitat selection and to investigate habitat use. For each habitat variable, this analysis tests with a Wilk’s lambda if the different habitat types are used more or less than expected from their availability and ranks the habitat types in order of use by comparing them two by two. For each habitat variable, the compositional analysis was also used to test for the significance of the use of each habitat type compared to all other combined.
Although the aforementioned analysis (Aebischer et al., 1993) allows testing the significance of the relative habitat use and the occurrence of habitat selection, no absolute preference per habitat type is calculated (Pauwels et al., 2016). Therefore, we calculated selection ratios for the pool of individuals, hereafter called mean selection ratios, and their associated Bonferroni-adjusted 95% confidence intervals as proposed by Manly et al. (2002).
The mean selection ratio is given by
where u i+ is the corrected number of positions of all fish in habitat type i,
with n the total number of fish, and u ++ is the corrected total number of positions
The variance of \(\hat{w}_{i}\) is estimated as
Simultaneous Bonferroni confidence intervals for mean selection ratios can then be constructed with an overall confidence level of 100(1 − α)%, so that the probability of all the intervals containing the true value is 1 − α. These intervals are of the form
where z ∝/2I is the value exceeded with probability ∝/2I by a standard normal random variable.
The mean selection ratio pools observations from all fish in the sample, but the confidence interval takes the variation in resource selection from individual to individual into account (Manly et al., 2002). When a selection ratio and accompanying confidence interval is higher than 1.0, habitat preference is considered significant (Rogers & White, 1990; Manly et al., 2002).
Compositional analysis and selection ratios were generated in R 3.0.1 (R Core Team, 2013) using adehabitatHS package (Calenge, 2006). GAMM were implemented using the mgcv package (Wood, 2006).
Methodological considerations
We can mention that the pool of individuals was not strictly the same from one season to another but we took this into account both in the GAMM, by modelling individual effects, and in the confidence interval of mean selection ratios. By construction, the habitat use analysis also considers this.
We also paid great attention to the crucial step of telemetry experiment design (Kessel et al., 2014; Steel et al., 2014) and evaluated its performance (Roy et al., 2014) as recommended by Biesinger et al. (2013). The spatial variability of the performance was assessed in a prior study (Roy et al., 2014): the performance (positioning error and probability to get a position) was lower in the littoral zone than in the pelagic one. At least two reasons can explain this: firstly, the littoral zone is a structurally complex zone what has been shown to affect the system performance in terms of detection and error (Baktoft et al., 2015); secondly, by construction of the receiver network, the littoral zone is mainly outside this network, whereas the pelagic one is inside and the VPS theory tells that the performance of the system is lower outside the network (Smith, 2013). Hence, we brought a correction to smooth this artificial spatial variability using the probabilities of positioning calculated by Roy et al. (2014) in winter when the mixing of water was complete. In addition, the performance of a system can vary in time due to the modifications of the thermocline gradient and thermocline depth (Huveneers et al., 2016). We checked this over the 40 synchronizing tags and eight reference tags and no strong seasonal pattern of detection was found in this reservoir making our positioning adequate to study seasonal variability of habitat preferences.
Results
WL influence on habitat availability
When the WL rose, the relative availability of the littoral area ([0, 2.5[ m) dropped from 19.5% at low level to 13.3% at high level (Fig. 3a). These proportions, applied to the mean lake area at the different WL, also showed that the surface of the littoral area diminished when the WL rose. In the littoral zone, the relative availabilities of stone and boulder/rock were quite low (less than 10%) and little influenced by WL (Fig. 3b). Similarly, the relative availability of tree stumps was not strongly impacted by WL and remained stable around 20% (Fig. 3d). On the contrary, the WL influenced the relative availability of finer substrates and lawn (Fig. 3b). The relative availability of silt prevailed at the lowest WL and dropped rapidly when the WL rose; conversely, the relative availability of gravel/pebble and lawn (class almost not present at low WL) increased with increasing WL to reach 19.5 and 27.6%, respectively, whereas the relative availability of sand peaked at the mean WL (Fig. 3b). The relative availability of emerging trees increased with increasing WL, roughly from 18.9% at low WL to 59.6% at high WL (Fig. 3c).

Relative availability (%) of the different habitat types at low, mean and high water levels: a the depth classes, b the main substrate types, c the presence of emerging trees and d the presence of tree stumps. The error bar represents two standard deviations
Which variables influence perch habitat preference?
Depth selection ratios were the best modelled with 44.4% of the deviance explained (Table 2). This model showed that perch preferences were different between depth classes and influenced by the season with individual variability. The WL did not impact significantly the depth preferences. The models selected for tree stumps and emerging trees selection ratios explained, respectively, 33.4 and 24.3% of the deviance (Table 2) without significant individual differences. Perch preference for tree stumps and emerging trees had a significant seasonal component but was not influenced by WL. Main substrate selection ratios were the most poorly modelled with only 11.2% of deviance explained by the selected model in which neither the season nor the WL had a significant effect. The individual variability was not significant.
Depth use and preference
The selection of depth occurred in the four seasons. Whatever the season, perch used the [2.5, 5[ m zone the most, significantly in spring, summer and autumn (Table 3). In spring and summer, the littoral zone use ranked second and was significantly more used than other depths only in summer, whereas in autumn and winter the [5, 7.5[ m depth ranked second. In autumn and winter, the littoral zone use only ranked four.
In spring and summer, perch preferred littoral and [2.5, 5[ m zones (Fig. 4a), in good agreement with the high use of these depths (Table 3). In autumn, perch preferred the [2.5, 5[ m zone and tended to prefer the [5, 7.5[ m depth, again in line with the high use of these depths, whereas they tended to avoid the littoral zone and avoided all other depths. In winter, the littoral zone was avoided and areas between 2.5 and 10 m deep tended to be preferred, and also more used. Over the four seasons, the areas deeper than 10 m were avoided or rarely explored by perch, except by some individuals in the winter.

Perch mean selection ratios of a depth and, in the littoral zone, of b the main substrate, c emerging trees and d tree stumps. The sample of perch used is given in the upper left corner. The 95% Bonferroni confidence intervals (vertical dashed bars) of the selection ratios are represented. The 1 threshold value, corresponding to “no preference”, is represented by a horizontal dashed line
Use and preference of littoral habitats
Perch selected the littoral substrate (Table 3). In the littoral zone, boulder/rock was significantly the most used substrate type, followed by stone and sand which were not significantly more used than other substrates (Table 3). In line with their high use, boulder/rock and stone tended to be preferred (Fig. 4b), as lawn although ranked second last regarding its relative use (Table 3). Perch tended to avoid finer substrates (silt, sand and gravel/pebble) (Fig. 4b). However, no preference/avoidance was significant due to a high individual variability, highlighted by large confidence intervals, especially for stone and boulder/rock.
Perch selected littoral zones with emerging trees in spring and summer; they were significantly more used than those without in these both seasons and also in winter (Table 3) but preferred only in spring and summer (Fig. 4c). In autumn and winter, perch also tended to prefer them but the individual variability was high (Fig. 4c).
The tree stumps were also selected in spring and summer and significantly more used (Table 3). Not completely in line with the habitat use analysis, they were preferred in spring and autumn and only tended to be in summer and winter, with a very high individual variability in spring and winter (Fig. 4d).
Discussion
Variability of habitat availability with WL
Using a precise qualitative and quantitative description of the different habitats, we highlighted an influence of WL on the structure of the available littoral habitats of the Bariousses Reservoir: they tended to become more homogeneous with a lowering structural complexity when the WL dropped. These results, based on the entire lake, are in agreement with other studies (Gasith & Gafny, 1990, 1998), and mean that lowering WL corresponds to fewer refuge areas and probably fewer food resources for perch (Zohary & Ostrovsky, 2011; Zohary & Gasith, 2014). They also confirmed the trends observed by Logez et al. (2016) with a point sampling on the same lake, but brought additional information as the stable availability of littoral tree stumps with WL and the rise of the relative availability and surface of the littoral area when the WL dropped. The highest diversity of habitats was observed in spring, which corresponds to the perch spawning period, while the lowest diversity was noted in autumn. It was intermediate almost throughout the summer when WLF was very limited around the mean WL, and very contrasted in winter when WL was quite evenly distributed between low, mean and high classes.
Influence of WL on habitat preference
WLF directly impacted the relative availability of the different littoral habitats; however, we did not observe any significant effect of WL on the habitat preference of adult perch. This suggests that the preference of littoral habitats remained the same whatever their availability, in the ranges and timing encountered in this reservoir. The scarcity of some preferred resources probably did not reach any critical threshold. Low WL was the most frequent in autumn and winter and corresponded to the highest relative availability of the littoral zone which was, however, much less used in these two seasons than in spring and summer.
In the same reservoir in summer and autumn, Logez et al. (2016) observed that the littoral fish assemblages, composed of juveniles and adults, were dependent on the WL and tended to homogenize when the habitat complexity lowered, suggesting variations in habitat use when its availability changed, even if adults seemed to be the less affected. Considering the advantages of the littoral zone for fish fauna (Schiemer et al., 1995; Schmieder, 2004; Lewin et al., 2014) and the importance of the habitats impacted by WLF for perch (Imbrock et al., 1996; Zamora & Moreno-Amich, 2002; Pekcan-Hekim et al., 2005; Cech et al., 2009; Muska et al., 2013), we expected some changes in the habitat preferences at different WL. As shown on some terrestrial species, variations in habitat availability can lead to changes in habitat preferences (Godvik et al., 2009; Hansen et al., 2009; Pellerin et al., 2010). With the loss of some emerging trees and boulder/rock habitat types when the WL dropped, we could have expected a higher attractiveness of tree stumps, whose availability remained stable, for feeding and spawning. In the Bariousses Reservoir, the temporal scales of WLF (days) mainly changed the habitat availability without imposing great physical stress on organisms living in the littoral zone as short-term WLF would (Hofmann et al., 2008). Moreover, the relative availability and surface of the littoral area rose when the WL dropped, which could mitigate the effects of the WLF. Above all, the relative availability of the complex preferred habitats by adult perch was the highest in spring and summer, respectively, the period of spawning (Craig, 2000) and of highest activity (Jacobsen et al., 2002), when this type of habitat is the most required regarding perch ecology. In autumn and winter, when food requirements lessen because perch activity is reduced (Jacobsen et al., 2002), perch could cope with a limited availability of complex habitats. Aside this uneven distribution of WL between seasons, the lack of a net effect of WL on adult perch habitat preferences can also be related to their plastic nature regarding the environment (Craig, 2000). Besides, Čech et al. (2012b) have shown that, being able to spawn at various depths depending on the period and temperature, perch have evolved a mechanism to cope with a large spectrum of conditions.
Individual variability
Finally, even if the individual effect was globally significant only for depth selection, high individual variability, quantified by the confidence interval length of selection ratios, was observed on numerous habitat types. It is, however, important to emphasize that no link appeared between the individual variability and the number of individuals used in the sample. Regarding the substrates, the individual variability was very high especially for the most complex ones what could be linked to their relatively reduced availability in this reservoir.
Such variability is frequent among fish (Magurran, 1993) and contributes to the population adaptation to rapid changes in the environment. This variability could reflect different strategies adopted by perch, possibly related to sex-specific responses to environmental changes (Estlander et al., 2015) or to predation risk (Estlander & Nurminen, 2014) that also exists even if limited on these large individuals, as well as to the existence of different fish personalities in the population, for example bold or shy individuals, which have been shown to forage or manage the predation risk differently (Kekalainen et al., 2014; Harkonen et al., 2016). Unfortunately, despite the surgical operation, the gender of perch was seldom determined because the smallest possible incision was favoured to provide the greatest chance of full healing.
Seasonal pattern of habitat use and preference
The high use of and preference for the shallow zones of the reservoir ([0, 5[ m in depth) in spring and summer shown with high-resolution individual tracking are in agreement with previous conclusions obtained with fishing data (Craig, 1977; Muska et al., 2013). They also confirm results obtained with a comparable approach implemented in another temperate lake (Zamora & Moreno-Amich, 2002). However, even if these shallow zones were highly used and preferred, the individual variability was also significant. In stratified reservoirs, fish are usually distributed in the surface waters due to the attraction of warmer temperatures in spring and to avoid deoxygenated hypolimnion in summer (Kubecka & Wittingerova, 1998). What is more, the preference for shallow depths by adult perch could be related to the numerous juveniles of different fish species that live in this sheltered area and that provide food (Degiorgi & Grandmottet, 1993; Stoll et al., 2008) and, in spring, also to the search for spawning support (Cech et al., 2009). Emerging trees and tree stumps, which are rigid, structurally complex and strongly used and preferred in spring, correlate to the type of spawning substrate sought by perch (Gillet & Dubois, 1995; Cech et al., 2009; Snickars et al., 2010). This preference for complex habitats observed throughout the year for the substrate type and to a lesser extent in summer for emerging trees and tree stumps can probably be related to feeding given that they can shelter macro-invertebrates and fish (Czarnecka et al., 2014). With the beginning of autumnal mixing, the perch migrated into slightly deeper waters as already described (Craig, 1977; Imbrock et al., 1996). We showed that, in the Bariousses Reservoir, the most used and preferred depths shifted to [2.5, 7.5[ m in autumn and winter. The low use of the littoral zone in these seasons was accompanied by a decrease of the attractiveness of emerging trees and tree stumps. Regarding the preferences, some exceptions could appear in winter, when littoral zones with emerging trees were still significantly more used than those without (Table 3) or in autumn, when the compositional analysis did not see any significant use or selection of tree stumps (Table 3), whereas the selection ratios showed a significant preference (Fig. 4d). In this last case, the significance was, however, not very strong and a small variation in the sample could probably have led to a non-significant result, especially as the availability of emerging trees was the lowest in autumn.
Such studies would be worth conducting on different species to improve knowledge of the impacts of human-induced WLF in reservoirs. Even if we did not show any effect of WLF on adult perch habitat preference, they could act differently on other species such as pike, a more demanding species in terms of spawning substrate (Craig, 2008). Very recently, Cooke et al. (2016) submitted that: “quantifying and describing the spatial ecology of fish and their habitat is an important component of freshwater fishery assessment and management”. Fish habitat requirement is the basis for rehabilitation (Müller & Stadelmann, 2004; Cooke et al., 2016) and could help to adapt hydraulic management to efficiently preserve fish populations. Operational tools to quantify human influence (through hydraulic management, stocking, recreational use) and environmental drivers (temperature, water quality) on fish populations in reservoirs still remain scarce. In environments where the littoral zone is altered, hydromorphological rehabilitation programmes are often implemented (Gonzalez et al., 2015). Any study that could provide insight into the impact of a hydromorphological restoration on the fish population is valuable (e.g. Boromisza et al., 2014; Rose et al., 2015), including species-habitat preference studies.
In conclusion, our study indicated that large perch selected zones at different depths in the reservoir mainly according to seasons without any effect of WL. In spring and summer, they highly used and preferred the littoral zone and the complex habitats it hosts, independently of the WL, even if the structural complexity of the littoral zone was reduced when the WL lowered. In autumn and winter, they migrated to deeper zones. Our results also highlighted quite a high individual variability in the habitat preferences. This study contributes to the understanding of the spatial ecology of fish, which is essential to leading efficient management actions (Cooke et al., 2016). Based on these findings, even if WLFs were shown not to impact perch habitat preferences, we can propose the following management actions to prevent eggs from being dewatered: in spring, to keep the WL stable at mean or high level or at least to avoid sharp lowering of WL from mid-April, the likely start of the spawning season, to mid-June when the majority of eggs have hatched; to add spawning substrates, woody structures for example, at mean and low WL to encourage perch to spawn deeper.
References
Aebischer, N. J., P. A. Robertson & R. E. Kenward, 1993. Compositional analysis of habitat use from animal radio-tracking data. Ecology 74: 1313–1325.
Baktoft, H., P. Zajicek, T. Klefoth, J. C. Svendsen, L. Jacobsen, M. W. Pedersen, D. M. Morla, C. Skov, S. Nakayama & R. Arlinghaus, 2015. Performance assessment of two whole-lake acoustic positional telemetry systems – is reality mining of free-ranging aquatic animals technologically possible? PLoS ONE. https://doi.org/10.1371/journal.pone.0126534.
Biesinger, Z., B. M. Bolker, D. Marcinek, T. M. Grothues, J. A. Dobarro & W. J. Lindberg, 2013. Testing an autonomous acoustic telemetry positioning system for fine-scale space use in marine animals. Journal of Experimental Marine Biology and Ecology 448: 46–56.
Boromisza, Z., E. P. Torok & T. Acs, 2014. Lakeshore-restoration-landscape ecology-land use: assessment of shore-sections, being suitable for restoration, by the example of Lake Velence (Hungary). Carpathian Journal of Earth and Environmental Sciences 9: 179–188.
Bridger, C. J. & R. K. Booth, 2003. The effects of biotelemetry transmitter presence and attachment procedures on fish physiology and behavior. Reviews in Fisheries Science 11: 13–34.
Calenge, C., 2006. The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling 197: 516–519.
Cech, M., J. Peterka, M. Riha, T. Juza & J. Kubecka, 2009. Distribution of egg strands of perch (Perca fluviatilis L.) with respect to depth and spawning substrate. Hydrobiologia 630: 105–114.
Čech, M., J. Peterka, M. Říha, L. Vejřík, T. Jůza, M. Kratochvíl, V. Draštík, M. Muška, P. Znachor & J. Kubečka, 2012a. Extremely shallow spawning of perch (Perca fluviatilis L.): the roles of sheltered bays, dense semi-terrestrial vegetation and low visibility in deeper water. Knowledge and Management of Aquatic Ecosystems. https://doi.org/10.1051/kmae/2012026.
Čech, M., L. Vejřík, J. Peterka, M. Říha, M. Muška, T. Jůza, V. Draštík, M. Kratochvíl & J. Kubečka, 2012b. The use of artificial spawning substrates in order to understand the factors influencing the spawning site selection, depth of egg strands deposition and hatching time of perch (Perca fluviatilis L.). Journal of Limnology 71: 18.
CEN, 2005. Water Quality – Sampling of Fish with Multi-mesh Gillnets EN 14757.27.
Cooke, S. J., E. G. Martins, D. P. Struthers, L. F. Gutowsky, M. Power, S. E. Doka, J. M. Dettmers, D. A. Crook, M. C. Lucas & C. M. Holbrook, 2016. A moving target – incorporating knowledge of the spatial ecology of fish into the assessment and management of freshwater fish populations. Environmental Monitoring and Assessment 188: 1–18.
Coops, H., M. Beklioglu & T. L. Crisman, 2003. The role of water-level fluctuations in shallow lake ecosystems – workshop conclusions. Hydrobiologia 506: 23–27.
Craig, J. F., 1977. Seasonal changes in day and night activity of adult perch, Perca fluviatilis L. Journal of Fish Biology 11: 161–166.
Craig, J. F., 2000. Percid Fishes: Systematics, Ecology and Exploitation. Blackwell Science, Oxford.
Craig, J. F., 2008. A short review of pike ecology. Hydrobiologia 601: 5–16.
Czarnecka, M., F. Pilotto & M. T. Pusch, 2014. Is coarse woody debris in lakes a refuge or a trap for benthic invertebrates exposed to fish predation? Freshwater Biology 59: 2400–2412.
Degiorgi, F. & J. P. Grandmottet, 1993. Relations entre la topographie aquatique et l’organisation spatiale de l’ichtyofaune lacustre : définition des modalités spatiales d’une stratégie de prélèvement reproductible. Bulletin Français de Pêche et de Pisciculture 329: 199–220.
Diehl, S., 1993. Effects of habitat structure on resource availability, diet and growth of benthivorous perch, Perca fluviatilis. Oikos. https://doi.org/10.2307/3545353.
Espinoza, M., T. J. Farrugia & C. G. Lowe, 2011. Habitat use, movements and site fidelity of the gray smooth-hound shark (Mustelus californicus Gill 1863) in a newly restored southern California estuary. Journal of Experimental Marine Biology and Ecology 401: 63–74.
Estlander, S. & L. Nurminen, 2014. Feeding under predation risk: potential sex-specific response of perch (Perca fluviatilis). Ecology of Freshwater Fish 23: 478–480.
Estlander, S., L. Nurminen, T. Mrkvicka, M. Olin, M. Rask & H. Lehtonen, 2015. Sex-dependent responses of perch to changes in water clarity and temperature. Ecology of Freshwater Fish 24: 544–552.
Evtimova, V. V. & I. Donohue, 2014. Quantifying ecological responses to amplified water level fluctuations in standing waters: an experimental approach. Journal of Applied Ecology 51: 1282–1291.
Evtimova, V. V. & I. Donohue, 2016. Water-level fluctuations regulate the structure and functioning of natural lakes. Freshwater Biology 61: 251–264.
Fischer, P. & U. Ohl, 2005. Effects of water-level fluctuations on the littoral benthic fish community in lakes: a mesocosm experiment. Behavioral Ecology 16: 741–746.
Gaboury, M. N. & J. W. Patalas, 1984. Influence of water level drawdown on the fish populations of Cross Lake, Manitoba. Canadian Journal of Fisheries and Aquatic Sciences 41: 118–125.
Gardner, C. J., D. C. Deeming & P. E. Eady, 2015. Seasonal water level manipulation for flood risk management influences home-range size of common bream Abramis brama L. in a lowland river. River Research and Applications 31: 165–172.
Gasith, A. & S. Gafny, 1990. Effects of water level fluctuation on the structure and function of the littoral zone. In Large Lakes. Springer, New York: 156–171.
Gasith, A. & S. Gafny, 1998. The importance of physical structure in lakes: the case study of Lake Kinneret and general implications. In Jeppesen, E., M. Sondergaard, M. Sondergaard & K. Christoffersen (eds), The Structuring Role of Submerged Macrophytes in Lakes. Springer, Berlin: 331–338.
Gillet, C. & J. P. Dubois, 1995. A survey of the spawning of perch (Perca fluviatilis), pike (Esox lucius), and roach (Rutilus rutilus), using artificial spawning substrates in lakes. Hydrobiologia 300–301: 409–415.
Gilman, E., M. Chaloupka, A. Read, P. Dalzell, J. Holetschek & C. Curtice, 2012. Hawaii longline tuna fishery temporal trends in standardized catch rates and length distributions and effects on pelagic and seamount ecosystems. Aquatic Conservation: Marine and Freshwater Ecosystems 22: 446–488.
Godvik, I. M. R., L. E. Loe, J. O. Vik, V. Veiberg, R. Langvatn & A. Mysterud, 2009. Temporal scales, trade-offs, and functional responses in red deer habitat selection. Ecology 90: 699–710.
Gonzalez, E., A. A. Sher, E. Tabacchi, A. Masip & M. Poulin, 2015. Restoration of riparian vegetation: a global review of implementation and evaluation approaches in the international, peer-reviewed literature. Journal of Environmental Management 158: 85–94.
Halpern, B. S., S. D. Gaines & R. R. Warner, 2005. Habitat size, recruitment, and longevity as factors limiting population size in stage-structured species. The American Naturalist 165: 82–94.
Hansen, B. B., I. Herfindal, R. Aanes, B.-E. Saether & S. Henriksen, 2009. Functional response in habitat selection and the tradeoffs between foraging niche components in a large herbivore. Oikos 118: 859–872.
Harkonen, L., P. Hyvarinen, P. T. Niemela & A. Vainikka, 2016. Behavioural variation in Eurasian perch populations with respect to relative catchability. Acta Ethologica 19: 21–31.
Hastie, T. J. & R. J. Tibshirani, 1990. Generalized additive models. In Monographs on Statistics and Applied Probability, Vol. 43. Chapman and Hall, London.
Hayes, D., M. Jones, N. Lester, C. Chu, S. Doka, J. Netto, J. Stockwell, B. Thompson, C. K. Minns, B. Shuter & N. Collins, 2009. Linking fish population dynamics to habitat conditions: insights from the application of a process-oriented approach to several Great Lakes species. Reviews in Fish Biology and Fisheries 19: 295–312.
Hofmann, H., A. Lorke & F. Peeters, 2008. Temporal scales of water-level fluctuations in lakes and their ecological implications. Hydrobiologia 613: 85–96.
Hokanson, K. E. F., 1977. Temperature requirements of some Percids and adaptations to the seasonal temperature cycle. Journal of the Fisheries Research Board of Canada 34: 1524–1550.
Hudon, C., P. Gagnon, J. P. Amyot, G. Letourneau, M. Jean, U. Plante, D. Rioux & M. Deschenes, 2005. Historical changes in herbaceous wetland distribution induced by hydrological conditions in Lake Saint-Pierre (St. Lawrence River, Quebec, Canada). Hydrobiologia 539: 205–224.
Huveneers, C., C. A. Simpfendorfer, S. Kim, J. M. Semmens, A. J. Hobday, H. Pederson, T. Stieglitz, R. Vallee, D. Webber, M. R. Heupel, V. Peddemors & R. G. Harcourt, 2016. The influence of environmental parameters on the performance and detection range of acoustic receivers. Methods in Ecology and Evolution 7: 825–835.
Imbrock, F., A. Appenzeller & R. Eckmann, 1996. Diel and seasonal distribution of perch in Lake Constance: a hydroacoustic study and in situ observations. Journal of Fish Biology 49: 1–13.
Irz, P., A. Laurent, S. Messad, O. Pronier & C. Argillier, 2002. Influence of site characteristics on fish community patterns in French reservoirs. Ecology of Freshwater Fish 11: 123–136.
Irz, P., M. Odion, C. Argillier & D. Pont, 2006. Comparison between the fish communities of lakes, reservoirs and rivers: can natural systems help define the ecological potential of reservoirs? Aquatic Sciences 68: 109–116.
Jacobsen, L., S. Berg, M. Broberg, N. Jepsen & C. Skov, 2002. Activity and food choice of piscivorous perch (Perca fluviatilis) in a eutrophic shallow lake: a radio-telemetry study. Freshwater Biology 47: 2370–2379.
Jacobsen, L., S. Berg, H. Baktoft & C. Skov, 2015. Behavioural strategy of large perch Perca fluviatilis varies between a mesotrophic and a hypereutrophic lake. Journal of Fish Biology 86: 1016–1029.
Johnson, D. H., 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology 61: 65–71.
Kaczka, L. J. & L. E. Miranda, 2014. Size of age-0 crappies (Pomoxis spp.) relative to reservoir habitats and water levels. Journal of Freshwater Ecology 29: 525–534.
Kahl, U., S. Hulsmann, R. J. Radke & J. Benndorf, 2008. The impact of water level fluctuations on the year class strength of roach: implications for fish stock management. Limnologica 38: 258–268.
Kekalainen, J., T. Podgorniak, T. Puolakka, P. Hyvarinen & A. Vainikka, 2014. Individually assessed boldness predicts Perca fluviatilis behaviour in shoals, but is not associated with the capture order or angling method. Journal of Fish Biology 85: 1603–1616.
Kessel, S. T., S. J. Cooke, M. R. Heupel, N. E. Hussey, C. A. Simpfendorfer, S. Vagle & A. T. Fisk, 2014. A review of detection range testing in aquatic passive acoustic telemetry studies. Reviews in Fish Biology and Fisheries 24: 199–218.
Koster, W. M., D. R. Dawson, P. Clunie, F. Hames, J. McKenzie, P. D. Moloney & D. A. Crook, 2015. Movement and habitat use of the freshwater catfish (Tandanus tandanus) in a remnant floodplain wetland. Ecology of Freshwater Fish 24: 443–455.
Kottelat, M. & J. Freyhof, 2007. Handbook of European Freshwater Fishes. Publications Kottelat, Cornol.
Kubecka, J. & M. Wittingerova, 1998. Horizontal beaming as a crucial component of acoustic fish stock assessment in freshwater reservoirs. Fisheries Research 35: 99–106.
Lewin, W. C., T. Mehner, D. Ritterbusch & U. Bramick, 2014. The influence of anthropogenic shoreline changes on the littoral abundance of fish species in German lowland lakes varying in depth as determined by boosted regression trees. Hydrobiologia 724: 293–306.
Logez, M., R. Roy, L. Tissot & C. Argillier, 2016. Effects of water-level fluctuations on the environmental characteristics and fish–environment relationships in the littoral zone of a reservoir. Fundamental and Applied Limnology 189: 37–49.
Magurran, A. E., 1993. Individual differences and alternative behaviours. In Pitcher, T. J. (ed.), Behaviour of Teleost Fishes, 2nd edn. Chapman and Hall, London: 441–477.
Manly, B. F. J., L. L. McDonald, D. L. Thomas, T. L. MacDonald & W. P. Erikson, 2002. Resource Selection by Animals: Statistical Design and Analysis for Field Studies, 2nd ed. Kluwer Academic Publishers, Boston.
Michaletz, P. H., 1997. Factors affecting abundance, growth, and survival of age-0 gizzard shad. Transactions of the American Fisheries Society 126: 84–100.
Müller, R. & P. Stadelmann, 2004. Fish habitat requirements as the basis for rehabilitation of eutrophic lakes by oxygenation. Fisheries Management and Ecology 11: 251–260.
Muska, M., M. Tuser, J. Frouzova, V. Drastik, M. Cech, T. Juza, M. Kratochvil, T. Mrkvicka, J. Peterka, M. Prchalova, M. Riha, M. Vasek & J. Kubecka, 2013. To migrate, or not to migrate: partial diel horizontal migration of fish in a temperate freshwater reservoir. Hydrobiologia 707: 17–28.
Pauwels, I. S., P. L. M. Goethals, J. Coeck & A. M. Mouton, 2016. Habitat use and preference of adult pike (Esox lucius L.) in an anthropogenically impacted lowland river. Limnologica: Ecology and Management of Inland Waters. https://doi.org/10.1016/j.limno.2016.10.001.
Pekcan-Hekim, Z., J. Horppila, L. Nurminen & J. Niemistö, 2005. Diel changes in habitat preference and diet of perch (Perca fluviatilis), roach (Rutilus rutilus) and white bream (Abramis björkna). Archiv für Hydrobiologie Special Issues in Advances in Limnology 59: 173–187.
Pellerin, M., C. Calenge, S. Said, J. M. Gaillard, H. Fritz, P. Duncan & G. Van Laere, 2010. Habitat use by female western roe deer (Capreolus capreolus): influence of resource availability on habitat selection in two contrasting years. Canadian Journal of Zoology: Revue Canadienne De Zoologie 88: 1052–1062.
Probst, W. N., S. Stoll, H. Hofmann, P. Fischer & R. Eckmann, 2009. Spawning site selection by Eurasian perch (Perca fluviatilis L.) in relation to temperature and wave exposure. Ecology of Freshwater Fish 18: 1–7.
R Core Team, 2013. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0.
Richards, S. A., 2008. Dealing with overdispersed count data in applied ecology. Journal of Applied Ecology 45: 218–227.
Rogers, K. B. & E. P. Bergersen, 1995. Effects of a fall drawdown on movement of adult northern pike and largemouth bass. North American Journal of Fisheries Management 15: 596–600.
Rogers, K. B. & G. C White, 2007. Analysis of movement and habitat use from telemetry data. In Guy, C. S., M. L. Brown (eds), Analysis and interpretation of freshwater fisheries data. American Fisheries Society, Bethesda: 625–676.
Rose, K. A., S. Sable, D. L. DeAngelis, S. Yurek, J. C. Trexler, W. Graf & D. J. Reed, 2015. Proposed best modeling practices for assessing the effects of ecosystem restoration on fish. Ecological Modelling 300: 12–29.
Roy, R., J. Beguin, C. Argillier, L. Tissot, F. Smith, S. Smedbol & E. De-Oliveira, 2014. Testing the VEMCO Positioning System: spatial distribution of the probability of location and the positioning error in a reservoir. Animal Biotelemetry. https://doi.org/10.1186/2050-3385-2-1.
Sale, P. F., R. K. Cowen, B. S. Danilowicz, G. P. Jones, J. P. Kritzer, K. C. Lindeman, S. Planes, N. V. Polunin, G. R. Russ & Y. J. Sadovy, 2005. Critical science gaps impede use of no-take fishery reserves. Trends in Ecology and Evolution 20: 74–80.
Schiemer, F., M. Zalewski & J. E. Thorpe, 1995. Land and inland water ecotones – intermediate habitats critical for conservation and management. Hydrobiologia 303: 259–264.
Schmieder, K., 2004. European lake shores in danger – concepts for a sustainable development. Limnologica 34: 3–14.
Smith, F., 2013. Understanding HPE in the VPS Telemetry System. VEMCO Tutorials, Halifax.
Snickars, M., G. Sundblad, A. Sandström, L. Ljunggren, U. Bergström, G. Johansson & J. Mattila, 2010. Habitat selectivity of substrate-spawning fish: modelling requirements for the Eurasian perch Perca fluviatilis. Marine Ecology Progress Series 398: 235–243.
Souchon, Y. & L. Tissot, 2012. Synthesis of thermal tolerances of the common freshwater fish species in large Western Europe rivers. Knowledge and Management of Aquatic Ecosystems. https://doi.org/10.1051/kmae/2012008.
Steel, A., J. Coates, A. Hearn & A. P. Klimley, 2014. Performance of an ultrasonic telemetry positioning system under varied environmental conditions. Animal Biotelemetry 2: 15.
Stoll, S., P. Fischer, P. Klahold, N. Scheifhacken, H. Hofmann & K. O. Rothhaupt, 2008. Effects of water depth and hydrodynamics on the growth and distribution of juvenile cyprinids in the littoral zone of a large pre-alpine lake. Journal of Fish Biology 72: 1001–1022.
Sutela, T. & T. Vehanen, 2008. Effects of water-level regulation on the nearshore fish community in boreal lakes. Hydrobiologia 613: 13–20.
Tao, J., D. S. Wang, K. Q. Chen & X. Sui, 2016. Productive capacity of fish habitats: a review of research development and future directions. Environmental Earth Sciences. https://doi.org/10.1007/s12665-015-5056-5.
Thiem, J. D., M. K. Taylor, S. H. McConnachie, T. R. Binder & S. J. Cooke, 2011. Trends in the reporting of tagging procedures for fish telemetry studies that have used surgical implantation of transmitters: a call for more complete reporting. Reviews in Fish Biology and Fisheries 21: 117–126.
Thorpe, J. E., 1977. Morphology, physiology, behavior, and ecology of Perca fluviatilis L. and P. flavescens Mitchill. Journal of the Fisheries Board of Canada 34: 1504–1514.
Vehanen, T. & M. Lahti, 2003. Movements and habitat use by pikeperch (Stizostedion lucioperca (L.)) in a hydropeaking reservoir. Ecology of Freshwater Fish 12: 203–215.
Venables, W. N. & B. D. Ripley, 2002. Random and mixed effects. In Modern Applied Statistics with S. Springer, New York: 271–300.
Wetzel, R. G., 1990. Reservoir ecosystems: conclusions and speculations. In Thornton, K. W., B. L. Kimmel & F. E. Payne (eds), Reservoir Limnology: Ecological Perspective. Wiley, New York: 227–238.
Winfield, I. J., 2004. Fish in the littoral zone: ecology, threats and management. Limnologica 34: 124–131.
Winter, J.-D., 1983. Underwater biotelemetry. In Nielsen, L.-A. & D.-L. Johnson (eds), Fisheries Techniques. American Fisheries Society, Bethesda: 371–395.
Wood, S., 2006. Generalized Additive Models: An Introduction with R. CRC Press, Boca Raton.
Wood, S. N., 2008. Fast stable direct fitting and smoothness selection for generalized additive models. Journal of the Royal Statistical Society: Series B (Statistical Methodology) 70: 495–518.
Zamora, L. & R. Moreno-Amich, 2002. Quantifying the activity and movement of perch in a temperate lake by integrating acoustic telemetry and a geographic information system. Hydrobiologia 483: 209–218.
Zohary, T. & A. Gasith, 2014. The Littoral Zone. In Lake Kinneret. Springer: 517–532. https://doi.org/10.1007/978-94-017-8944-8_29.
Zohary, T. & I. Ostrovsky, 2011. Ecological impacts of excessive water level fluctuations in stratified freshwater lakes. Inland Waters 1: 47–59.
Zuur, A. F., E. N. Ieno, N. J. Walker, A. A. Saveliev & G. M. Smith, 2009. In Gail, M., K. Krickeberg, J. M. Samet, A. Tsiatis & W. Wong (eds), Mixed Effects Models and Extensions in Ecology with R. Springer, New York.
Acknowledgements
We thank Tiphaine Peroux, Julien Dublon, Marie-Laure Acolas, Mario Lepage, Virginie Raymond and Charlie Roqueplo for their assistance in the field, Yann Le Coarer for dGPS treatments, Nathalie Reynaud for GIS treatments and Hervé Capra, Nicolas Lamouroux and Hervé Pella for useful discussions. We also thank numerous other people who occasionally helped in the field. Furthermore, we thank Electricité de France, which manages the Bariousses Reservoir, for permission to use the lake and water-level data. The authors would like to thank two anonymous reviewers, whose advice substantially improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Fernando M. Pelicice
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Westrelin, S., Roy, R., Tissot-Rey, L. et al. Habitat use and preference of adult perch (Perca fluviatilis L.) in a deep reservoir: variations with seasons, water levels and individuals. Hydrobiologia 809, 121–139 (2018). https://doi.org/10.1007/s10750-017-3454-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-017-3454-2