
Abstract
Trophic dynamics and the relative importance of allochthonous versus autochthonous production in supporting invertebrate consumers have been studied from headwaters to large rivers but rarely in springs. Our goals were to examine spatial and temporal invertebrate feeding pathways in three U.S. karst springs differing in canopy coverage and type (coniferous, open, or deciduous). Based on current food web models for headwaters, we initially hypothesized that: (a) overall biomass production would primarily depend on allochthonous carbon; (b) terrestrial carbon would be most important following leaf litter input; and (c) autochthonous carbon would be responsible for most of the biomass production in open canopy areas. To test these predictions, we analyzed trophic pathways using carbon and nitrogen stable isotopes, biomass measurements, and stoichiometric analyses. Contrary to our initial hypotheses, the proportional contribution of autochthonous carbon to consumer biomass exceeded that of allochthonous in each spring along temporal and spatial scales. Autochthony increased spatially along the gradient between spring sources and lower springbrook reaches and was highest in the open canopy spring. Allochthony increased slightly during the winter following litter fall. Despite the reported importance of periphyton to zoobenthic consumers, macroinvertebrate biomass was primarily supported by moss and water cress in these springs.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Deciphering the relative importance of diverse carbon sources supporting food webs is both a historical (e.g., Vannote et al., 1980; Pimm, 2002) and modern focus in aquatic systems (e.g., Pingram et al., 2012; Cross et al., 2013). In particular, the relative importance of terrestrial detritus versus instream primary production in fueling consumer biomass has often been questioned (Finlay, 2001; Bunn et al., 2003; Yam & Dudgeon, 2005; McNeely et al., 2007; Lau et al., 2009). The predominance of evidence has identified small forested headwater streams as principally heterotrophic and dependent on allochthonous leaf litter (Fisher & Likens, 1973; Vannote et al., 1980; Wallace et al., 1997). However, a significant degree of autochthony has been reported in some small forested streams (Bunn et al., 1999; Finlay et al., 2002; March & Pringle, 2003; Mantel et al., 2004; Lau et al., 2009) regardless of the relative standing stock of allochthonous and autochthonous matter available (Mayer & Likens, 1987; Salas & Dudgeon, 2001; Yam & Dudgeon, 2005). Little information exists, however, regarding the relative importance of autochthonous and allochthonous carbon to consumers in spring ecosystems.
Springs represent headwater systems that are considered “hot spots” for aquatic diversity (e.g., Barquín & Death, 2006; Cantonati et al., 2006). Despite their generally small size, springs can contribute as much as one third to the regional biodiversity of benthic species (Danks & Williams, 1991). Their permanency of flow and relative constancy of other physical and chemical conditions contribute to the success of species that reproduce asynchronously throughout the year (e.g., many amphipods). These conditions also account for the disproportionately large number of rare, relict, and endemic species in springs, and they enable springs to function as refugia for downstream epigean communities during floods and droughts (Teal, 1957; Hynes, 1983; Williams & Hogg, 1988; Erman & Erman, 1995).
We examined spatial (longitudinal) and temporal (seasonal) trophic pathways of benthic invertebrates in four reaches of three similar, neighboring karst springs in the Ozarks region of Missouri, USA. The goal of the work reported here was to determine the relative contribution of different food sources in supporting consumer biomass by employing carbon and nitrogen stable isotope ratios, macroinvertebrate biomass estimates, and stoichiometric (C:N) analyses.
Our three springs have similar geographic locations, subsurface and surface geologies, points of origin, flow magnitudes and permanencies, inorganic substrates, and physical and chemical stability. Our previous work on seasonal and longitudinal patterns of biotic community structure (richness, evenness, biomass, and density) and function (functional feeding groups and habit traits) in these springs showed negligible physical and chemical variability along each spring source to springbrook gradient (Carroll & Thorp, 2014). We concluded that physical and chemical parameters had little influence on the significant pattern of increasing biotic and functional diversity along these lengths (cf. Smith et al., 2003; Von Fumetti et al., 2006; Staudacher & Füreder, 2007; Kubíkoá et al., 2012) and focused this paper on other factors potentially affecting dominant invertebrate feeding pathways.
While very similar in stream habitat characteristics, our three spring systems have distinctly different riparia (deciduous forest, coniferous forest, and a combination of deciduous forest and open, manicured grassland). Based on these observations, prior epigean food web studies on headwater systems, and our previous study of diversity patterns in these springs, we hypothesized that: (a) the dominant organic source supporting macroinvertebrate biomass would be allochthonous, but the dependence on autochthonous sources (primarily algae) would increase in lower springbrook reaches; (b) feeding pathways would shift seasonally to a heavier reliance on allochthonous carbon in response to increased terrestrial-litter inputs; and (c) the relative importance of food sources would vary seasonally among the three springs due to relative differences in their riparia.
Methods
Study area and site descriptions
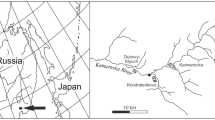
The study was conducted in three karst springs, Haseltine, Danforth, and Steury, each situated in the Ozark uplift region of Greene County, Missouri, USA (Fig. 1). All three springs lie in the Springfield Plateau physiographic region, a subdivision of the deeper Ozarks Plateau, and are all underlain by the Burlington-Keokuk Limestone of Mississippian age and older, slightly deeper Ordovician dolomites (Thomson, 1986). The springs are located in adjoining watersheds: Haseltine in the Sac River Basin (altitude 350 m a.s.l.), and Danforth and Steury in the James River Basin (altitude 410 and 396 m a.s.l., respectively). Each spring is defined as a third magnitude (mean discharge 0.04–0.17 m3/s−1), cool, freshwater, permanent rheocrene (Danks & Williams, 1991). The spring source (eucrenon) is clearly defined, as water issues from the subterranean habitat in each spring from a cave situated within a limestone bluff. The springbrook (hypocrenon) in each spring represented the point below the issuance at the cave, to the point just before it was influenced by input from any tributaries. Haseltine (37°15′N, 93°26′W) lies just northwest of Springfield, Missouri, and issues from a small cave (area = ~39 m2) into an open, mowed grassland with only one side of the springbrook lined with deciduous trees for a short distance adjacent to the cave (~30 m). The springbrook flows beyond the cave mouth to a point ~ 103 m downstream, just before it empties into a terminal lake. Danforth (37°23′N, 93°15′W) issues from a small cave (area = ~12 m2) into a planted coniferous forest that supports 17 different conifer genera, and Steury (37°20′N, 93°21′W) issues from a larger cave (area = ~64 m2) into a deciduous forest. Below the cave mouth, each is eventually influenced by input from other tributaries (145 & 101 m for Steury and Danforth, respectively). Nearly 80% of the channel in both Danforth and Steury is shaded (see Carroll & Thorp, 2014 for further details).

Map of location of three Ozarks springs in Springfield Plateau physiographic region, Greene County, Missouri, USA
Invertebrate sampling and identification
Three replicate zoobenthic samples were collected from the cave and three reaches downstream in each springbrook (reach 1-4; R1-R4) during two seasons (summer and winter), resulting in 12 invertebrate samples/spring/season. Because these springs are small, benthic sampling represented a potentially substantial disturbance to these fragile systems and was, therefore, limited to these two sampling periods to avoid unnecessary destruction of biota in the eucrenal and hypocrenal zones (cf. Erman, 2002). Macroinvertebrates were collected using a Surber sampler (0.093 m2; 500 μm mesh) while the substrate was being disturbed to a depth of ~15 cm for ~2 min. Samples were placed on ice for transport to the laboratory where they were sorted into major taxonomic groups and kept at 4.0°C for 24 h to clear their digestive tracts. Samples were then frozen for later identification, determinations of abundance and biomass, and stable isotope analysis. Invertebrates were identified and classified into functional feeding groups (FFGs: shredder, collector, scraper, filterer) using Merritt et al. (2008) and Thorp & Covich (2010). Most invertebrates were identified to species but chironomid midges were identified to subfamily, and oligochaete worms were identified to family. Secondary invertebrate consumers and vertebrate predators are rare in these systems and thus were not included in our analyses.
Invertebrate biomass determinations
Invertebrate biomass was estimated in two ways. For taxa having densities >100 specimens per Surber sample, we established taxon-specific length dry-mass equations (cf. Benke et al., 1999) by measuring the lengths and mass of 100 individuals. Before drying, body length (exclusive of appendages) was measured to the nearest 0.05 mm using a dissecting microscope and an ocular micrometer. Specimens were dried at 60°C for 48 h, and then individuals were weighed using a microbalance (sensitivity 0.002 mg) to determine dry mass. Subsequent biomass estimates for remaining specimens (>100) within a given taxa were then determined by measuring the length of each organism and determining its mass from the equation. Dry-mass measurements for taxonomic groups with <100 specimens per sample were determined directly by weighing each individual (2,600 total organisms measured directly). Crayfish were excluded from biomass comparisons because of both their disproportionate weight compared to other zoobenthic organisms and their rarity in the springs.
Collection of potential food sources
Replicates (n = 3) of potential organic food sources were collected from random locations in each reach within the riparian zone (5–6 C3 plants as well as C4 grasses) and stream proper during both summer and winter sampling periods. Terrestrial samples included leaves from dominant tree species (deciduous in two springs, coniferous in Danforth), grasses, and dominant herbaceous vegetation. Aquatic sources included: (a) suspended-fine-transported organic matter (FTOM; 0.1-1 mm); (b) dissolved organic matter (DOM; <0.1 mm); (c) coarse benthic organic matter from within the spring channel (BOM; consisting primarily of terrestrial leaves, pine needles, and grass); (d) epiphytic and epilithic biofilms; and (e) macrophytes (mosses: Fissidens debilis Schwäegrichen, Drepanocladus aduncus (Hedwig) Warnstorf, Fontinalis antipyretica Hedwig, and water cress: Nasturtium officinale W.T. Aiton). Ambient concentrations of coarse-transported organic matter (>1 mm) were minimal (too low to gather sufficient sample for isotopic measurements) and were not considered further.
Aquatic macrophytes were clipped above the roots, and algae were shaken free from plant hosts (Lowe & LaLiberte, 2006) and brushed from rock substrates. BOM was collected with a Surber sampler in each reach and wet-sieved to obtain a coarse (>1 mm) particulate organic matter fraction. Water for analysis of particulate and dissolved carbon was collected during both summer and winter sampling periods from three random transects in each sampling reach. Replicate water samples within each reach were pooled into a single, 20 l composite sample, thereby providing a cross-channel representation of organic matter in the water column (cf. Delong & Thorp, 2006). Terrestrial samples were placed in sterilized plastic bags and aquatic samples in acid-washed plastic vials and stored on ice for transport to the laboratory.
In the laboratory, water samples were passed through a fine (0.1 mm) sieve to collect FTOM for stable isotope analysis. Dissolved organic matter was obtained by filtering 3 l of this pre-filtered water through an ultra-fine (1-µm pore) glass-fiber filter, adjusting the pH to 4.3, and bubbling the sample with oxygen to remove inorganic forms of carbon (cf. Thorp et al., 1998). Primary food sources were cleaned and rinsed in distilled water. Invertebrate consumers were prepared whole except for crayfish where pure muscle tissue was extracted for stable isotope analysis. Samples were dried at 60°C for 48 h, ground to a fine powder with a Wig-L-Bug®, weighed to approximately 4 mg for food sources and 0.8 mg for animals, and placed into Ultra-Pure tin capsules (Costech Analytical, Valencia, California). Because carbonate concentrations are high in karst systems, FTOM and DOM samples were acidified with 1 N HCL after drying and grinding to remove this source of isotopic carbon.
Stable isotope and SIAR analyses
Prepared stable isotope samples were sent to the Colorado Plateau Stable Isotope Laboratory at Northern Arizona University, Flagstaff, Arizona. Ground samples for δ 13C (13C/12C) and δ 15N (15N/14N) analysis were processed with a DELTA V Advantage continuous flow mass spectrometer.
Isotope values are expressed as δ 13C or δ 15N according to the following equation:
where R = 13C/12C or 15N/14N. International standards used were Pee Dee belemnite for carbon and atmospheric nitrogen. Measurement error for the DELTA V was ± 0.05 ‰ for δ 13C and ± 0.13‰ for δ 15N. As part of the stable isotope analytical procedures, stoichiometric data for C/N mass percentages of samples were obtained. A total of 907 stable isotope samples from the three spring sites were analyzed: 462 samples were basal food sources, and 445 were consumer samples.
We used the Stable Isotope Analysis in R (SIAR) mixing model (Parnell et al., 2010; R Core Team, 2016) to calculate contributions of each source to consumer biomass. Prior to running SIAR models, we examined variation in C:N ratios and δ 13C and δ 15N signatures among food sources to evaluate their inclusion in the models.
The C:N values of dead organic food sources (FTOM, DOM, and BOM) were used to determine whether the sources were primarily derived from allochthonous or autochthonous materials. The C:N ratios of autochthonous food sources are lower than those of allochthonous sources, and C:N ratios >15 of organic matter are an indication of significant allochthonous contributions (Kendall et al., 2001). The C:N ratios of autochthonous carbon sources (water cress and moss) were consistently lower than those of allochthonous sources (Table 1), with water cress values ranging from 3.9 to 9.3 and moss values ranging from 6.6 to 12.1. The C:N ratios of allochthonous food sources (C3 plants and C4 grasses) ranged from 14.4 to 99.8, while ratios for DOM were consistently low among springs and seasons (range 1.7–7.7). Substantial seasonal variability in FTOM and BOM C:N ratios generally grouped them with autochthonous sources during the summer and allochthonous sources during the winter (Table 1).
Likewise, δ 13C and δ 15N were effective in discriminating among food sources. In all springs, moss was lower in δ 13C than all other food sources (range −45.6 to −35.6, Table 2). Water cress also tended to have a low δ 13C signature in all the springs (range −33.8 to −28.0). DOM had a low δ 13C signature (range −31.7 to −25.0) that was similar to the δ 13C signature of water cress and biofilm in many of the springs. DOM had a high δ 15N signature that identified it as a unique food source (Table 2). BOM and FTOM, however, had variable δ 13C signatures and could have originated from any of the food sources but moss. BOM ranged from −29.0 (Steury winter) to −10.0 (Haseltine summer) and FTOM ranged from −33.3 (Danforth summer) to −7.9 (Haseltine winter).
Based on these results, we included a total of seven food sources in SIAR models: moss, water cress, C3 plants, DOM, epilithic biofilm, epiphytic biofilm, and C4 grasses. Because C:N ratios and δ 13C indicated that BOM and FTOM were composed of a variety of sources, we did not include them as potential food sources in SIAR models. We used the summer and winter average and standard deviation of δ 13C and δ 15N of each of these seven food sources collected from each of the study reaches as an input value for the SIAR model. As an input for trophic fractionation of δ 13C, we used the average and standard deviation found for invertebrates (0.2‰ ± 1.4) in an analysis of fractionation in aquatic systems (Vander Zanden & Rasmussen, 2001). As an input for trophic fractionation of δ 15N, we used the average and standard deviation calculated for herbivorous invertebrates (0.6‰ ± 1.7) in an analysis of fractionation in rivers across Australia and New Guinea (Bunn et al., 2013). For each model, we ran 500,000 iterations, discarded 50,000 of the original iterations, and retained one of every 15 remaining iterations (Parnell et al., 2010). We used the lower and upper 95% credibility intervals of the seven food sources to calculate mean 95% credibility intervals for autochthonous (moss, water cress, DOM, epilithic biofilm, and epiphytic biofilm) and allochthonous (C3 plants, C4 grasses) sources.
Data analysis
We used the mean lower and upper 95% credibility intervals of food sources calculated using the SIAR program and invertebrate biomass measurements to estimate the contribution of autochthonous and allochthonous food sources to consumer biomass (Biomassautochthonous) for each FFG. We used the following biomass-weighted equation (cf. Jardine et al., 2013): Biomassautochthonous = (ΣmeanAUTOconsumer * Biomassconsumer), where ΣmeanAUTOconsumer was the sum of the mean 95% credibility intervals of autochthonous food sources for each invertebrate taxa, and BIOMASSconsumer was the biomass of that taxa. Comparative summations were performed for allochthonous food sources using the following equation: Biomassallochthonous = (ΣmeanALLOCconsumer * Biomassconsumer). The biomass-weighted calculations were then log (x + 1)-transformed and used as dependent variables in a three-way analysis of variance (ANOVA; NCSS, Kaysville, Utah, 2004) to test for effects of reach (distance from the spring source), spring, and season (fixed factors) on the relative importance of autochthonous and allochthonous food sources in supporting consumer biomass. Because of opposite downstream trends in biomass and density for crustaceans (decrease) versus insect/non-insect invertebrates (increase), ANOVA analyses related to spatial and temporal biomass-weighted contributions were analyzed separately for the two groups per individual taxa. Post hoc Fisher’s LSD Multiple-Comparison tests were performed to explore significant differences. Statistical significance was determined at α = 0.05.
Results
Summary of longitudinal changes in community composition
The spring source area in each of the three springs supported a depauperate faunal diversity with typically two species of Amphipoda (G. minus Say and C. forbesi Hubricht and Mackin) and one species of Isopoda (Lirceus hoppinae Faxon); these areas were generally devoid of aquatic insects. Diversity rose significantly with increasing distance from the spring source during both seasons in all three springs as peracaridans were replaced in springbrooks by a moderate diversity of aquatic insect and non-insect macroinvertebrates. The community composition of crustaceans and insects differed substantially from reaches 1–4, with peracaridans representing as high as 99% of the invertebrates in the spring source but as low as 0% in reach 1. In contrast, insects were as low as 0% in reach 1 and as high as 88% in reach 4. As a result, density (#/m2) and biomass (mg/m2) of peracaridans decreased, and insect and non-insect macroinvertebrates increased significantly along this gradient. Consequently, shredders were the primary or only FFG represented in each spring source (with the exception of a few aquatic worms—collectors). The relative abundance of representatives from all other FFGs, however, increased significantly with distance from the spring source (see Carroll & Thorp, 2014 for full zoobenthic community analysis).
Relationships between consumers and primary food sources
Examination of standard comparative bi-plots indicated that there was considerable variability in food source δ 13C and δ 15N values among springs and seasons (Fig. 2). Epilithic and epiphytic biofilm δ 13C signatures ranged from −38.5 to −10.7‰ and −32.8 to −19.0‰, respectively. C4 grasses tended to be higher in δ 13C than most other food source (average = −15.0‰). Among all springs, DOM had a high δ 15N signature (average = 9.0‰) that was distinct in δ 15N from many of the other food sources. Water cress also had a high δ 15N signature (average = 8.0‰) that distinguished it from other potential food sources. Among the springs, mean δ 15N values of several food sources, including C3 plants, DOM, and water cress, were lower in Danforth Spring than in Haseltine and Steury. In Haseltine, δ 15Nvalues of food sources tended to be higher in winter than in summer; however in Steury, δ 15N values of several food sources were higher in the summer. Some food sources also showed seasonal shifts in δ 13C values. For example, in all three springs, water cress and DOM had lower mean δ 13C values in winter than in summer. In all the springs, C4 grass was more enriched in 13C than all invertebrates. Epilithic algae were more enriched in 13C than invertebrates in Haseltine and Steury Springs.

Stable isotope ratios (δ 13C and δ 15N) of organic matter sources (squares, mean ± 1 SE) and invertebrate primary consumers (circles, mean) from three Ozarks Springs during summer and winter sampling periods. Circles: Black open Reach 1, gray solid Reach 2, gray open Reach 3, black solid Reach 4. Triangles represent crayfish (omnivorous consumer) with same color scheme per reach. Food sources: DOM dissolved organic matter, FPOM fine particulate organic matter, BOM benthic organic matter, WC water cress, C 3 terrestrial organic matter, C 4 C4 grasses
During the summer, δ 13C signatures for most invertebrates fell between −42 and −37‰ (Fig. 2). Most consumers had δ 13C signatures that were similar to moss (−45 to −39‰). However, in all springs, worms and crayfish had δ 13C values that were similar to water cress (−33 to −29‰). In Haseltine and Steury, δ 13C signatures of isopods resembled terrestrial plant material (−31 to −27‰). In Danforth, the δ 13C signatures of amphipods and snails resembled δ 13C of epilithic biofilm (−37 to −36‰). During the winter, δ 13C signatures for most consumers were substantially enriched compared to summer and fell between −35 and −29‰ (Fig. 2). For example, the δ 13C signatures of most consumers in Steury Spring were enriched on average by ~6‰ over summer. This trend, however, was not the case in Danforth Spring where most consumer δ 13C signatures remained in the range between −42 and −37‰. Crayfish were distinctly 15N enriched relative to primary producers and other primary consumers—indicative of their omnivorous feeding preferences.
SIAR models
The SIAR model estimates indicated that all invertebrate species assimilated a substantially greater fraction of autochthonous than allochthonous material in each of the three springs. In Haseltine Spring in both seasons, allochthonous contributions were low for invertebrate consumers. In the summer, “lower 95% credibility intervals” for allochthonous material were <0.05 for all invertebrate taxa in all four reaches. The maximum upper 95% credibility interval was 0.71 for isopods collected from the cave. For all other consumers, however, the average maximum upper 95% credibility was 0.35. In the winter, the lower 95% credibility intervals for all invertebrates were consistently <0.12. The maximum upper 95% credibility interval was 0.88 for isopods in the cave reach. For all other consumers, the average maximum upper 95% credibility was 0.39. Analysis of functional feeding groups showed that in Haseltine, all feeding groups predominantly assimilated carbon originating from moss in summer and winter (Tables 3, 4).
Contributions of allochthonous material to invertebrate biomass were also relatively low in Danforth Spring, although contributions increased slightly in the winter. In the summer, all consumers had lower 95% credibility intervals = 0 for allochthonous material except for isopods in the cave reach which had lower 95% credibility intervals = 0.15. Isopods in this reach also had the greatest upper 95% credibility intervals (0.35). For all other invertebrate taxa, upper 95% credibility intervals were <0.22. In the winter, contributions of allochthonous material increased slightly for some consumers. Several consumer taxa had lower 95% credibility intervals >0, including craneflies, isopods, snails, and riffle beetles. Isopods in the cave reach assimilated some allochthonous material, with lower 95% credibility intervals = 0.06 and upper 95% credibility intervals = 0.31. Craneflies in the most downstream reach assimilated some material from terrestrial plants, with lower 95% credibility intervals = 0.49 and upper 95% credibility intervals = 0.72. In Danforth, similar to Haseltine Spring, all functional feeding groups derived the majority of their carbon from moss in both seasons. However, in the cave reach in the summer, water cress was an important source of organic matter for collectors (Table 3). In the winter, DOM was a dominant source of organic matter for shredders and epilithic algae was an important source for collectors (Table 4).
In Steury Spring, material from terrestrial plants contributed to the biomass of several consumer taxa. In the summer, isopods and amphipods collected from the two reaches nearest the cave had relatively high upper 95% credibility intervals. Furthermore, snails collected from the most downstream reach had a wide range of 95% credibility intervals (lower 95% credibility interval = 0.09, upper 95% credibility interval = 0.73). Contributions of allochthonous material to consumer biomass likely increased in the winter. In the two reaches nearest the cave, isopods, chironomids, and oligochaetes had wide ranges of 95% credibility intervals and likely assimilated fractions of material from terrestrial plants. In the two downstream reaches, upper 95% credibility intervals for allochthonous material increased by more than two-fold for oligochaetes and net caddisflies in the winter. Analysis of functional feeding groups showed that in the summer, shredders, filterers, and scrapers assimilated organic carbon from moss and collectors assimilated much of their carbon from DOM (Table 3). In the winter, shredders derived material from moss and collectors, filterers, and scrapers derived material from moss and terrestrial C3 plants (Table 4).
Biomass-weighted contributions of autochthonous versus allochthonous carbon sources
Because large variability in the number of individuals/m2 collected among springs (i.e., Haseltine individuals/m2 twice that captured in Steury) and seasons (5,600 more individuals/m2 captured in summer vs. winter) equates to the amount of consumer biomass attributed to any given food source, ANOVA comparisons were performed on the relative proportion of autochthonous versus allochthonous biomass-weighted contributions to the total, per individual taxa.
Biomass-weighted source proportions indicated a clear reliance on autochthonous food sources by invertebrate consumers in all springs during both seasons (Appendix 1 in Supplementary material). Insect and non-insect invertebrate production supported by autochthonous food sources increased significantly with distance from the spring source (reach 1–4; Fig. 3) in all springs (ANOVA, reach factor: F (3,115) = 17.64, P < 0.001). Contributions also varied significantly among springs (ANOVA, spring factor: F (2,115) = 27.35, P < 0.001), with a greater amount of biomass supported by autochthonous food sources in Haseltine (primarily open canopy) followed by Danforth (coniferous) and Steury (deciduous).

Biomass-weighted contributions per taxa for a summer insect/non-insect invertebrate taxa and crustaceans b winter insect/non-insect invertebrate taxa and crustaceans. Numbers 1–4 represent reaches 1–4 along the spring source-springbrook continuum
Patterns for allochthonous support of insect consumer biomass were spatially similar to those found for autochthonous in each spring (increase R1-R4; ANOVA, reach factor: F (3,115) = 7.32, P < 0.001). Moreover, the amount of biomass supported by allochthonous sources was significantly greater during the winter than it had been in the summer (ANOVA, season factor: F (1,115) = 6.05, P = 0.02) and significantly greater in Steury followed by Haseltine and Danforth (ANOVA, spring factor: F (2,115) = 8.02, P = 0.001).
Biomass-weighted spatial trends for crustaceans were reversed, as these organisms virtually disappeared downstream with distance from the spring source (Fig. 3). Thus, biomass supported by autochthonous sources decreased significantly along the spring source-springbrook gradient (ANOVA, reach factor: F (3,47) = 3.90, P = 0.02). Spring effects were also found for variation in crustacean biomass supported by autochthonous contributions (ANOVA, spring factor: F (2,47) = 4.86, P = 0.01). Autochthonous contributions were significantly highest in Haseltine (open canopy) followed by Danforth (coniferous) and Steury (deciduous).
Similar to biomass-weighted autochthonous patterns, crustacean biomass supported by allochthonous sources also decreased significantly along the spring source-springbrook gradient (ANOVA, reach factor: F (3, 47) = 4.52, P = 0.01). Allochthonous spring-effect patterns were similar for autochthonous support of crustacean biomass (ANOVA, spring factor: F (2,47) = 9.98, P = 0.001) with dependence greatest in Steury followed by Haseltine and Danforth.
Discussion
The relative importance of autochthonous and allochthonous food sources
There is increasing evidence that autochthony is the dominant trophic pathway in lotic systems regardless of the amount of canopy cover (McCutchan & Lewis, 2002) or relative standing stock of allochthonous or autochthonous matter available (Yam & Dudgeon, 2005; Marcarelli et al., 2011). Our analyses showed that consumers in all three headwater springs depended for the majority of their biomass production on autochthonous food sources despite differences in their riparian cover. This conclusion was unexpected for the two heavily canopied springs for three reasons. First, headwater systems are typically classified in the scientific literature as being heterotrophic and dependent on input of allochthonous leaf litter. Second, the largest biomass of potentially available organic matter in most forested headwater streams comes from allochthonous leaf litter (Vannote et al., 1980). Third, the dominant functional feeding group, in terms of both abundance and biomass in each spring, was shredders (amphipods and isopods), which are known to rely primarily on allochthonous detritus for energy (Merritt et al., 2008).
Also contrary to initial hypotheses, our data showed that autochthonous support of consumer biomass in these three springs came from autochthonous sources other than periphyton, specifically moss and water cress. The heavy reliance on these macrophytes by amphipods and isopods was suggested by results from our previous zoobenthic community and functional feeding group analysis of these springs (Carroll & Thorp, 2014) and the work of others who have found moss and water cress to be important sources of carbon for these aquatic consumers (Winterbourn et al., 1986; Rosenfeld & Roff, 1992; Robinson et al, 2008). This could also be explained by the fact that peracaridans generally exhibit higher cellulase activities and, therefore, greater efficiencies in assimilating these plants (Kesler, 1983; Chamier, 1991; Glazier, 1991).
Despite the use of these resources by peracaridan shredders, the heavy reliance on submerged macrophytes by collectors, filterers, and scrapers was unexpected. Recent studies have confirmed that stream animals do not necessarily “prefer” leaf litter, but purposefully make opportunistic diet shifts to higher-quality algae or macrophytes even when leaf litter is abundant (Friberg & Jacobsen, 1994; Leberfinger et al., 2011; Marcarelli et al., 2011). Hence, several factors in our study suggest that regardless of the relative accessible quantities of autochthonous or allochthonous foods available to these consumers, their dependence on primarily moss and water cress was based on relative differences in food quality.
First, periphyton is thought to be the most important autochthonous carbon source in small streams due to its more nutritious nature as indicated by its relatively lower C:N ratios (Finlay, 2001; Lau et al., 2008). However, when epilithic periphyton C:N ratios are as high as those in Haseltine and Steury Springs, it may not be the more nutritious food choice. In both springs C4 grasses were prevalent, and dense mats of their clippings lined the springbrooks. Our data showed that C4 grasses had the most enriched δ 13C values and high C:N ratios, and are indications of their chemically low nutritional quality (Lau et al., 2008) and physically constraining anatomy. Because the algal biofilm samples processed here, as in many studies, likely contain a matrix of algae, bacteria, fungi, and accumulated terrestrial organic matter (Eminson & Moss, 1980; Allan, 1999; Bunn et al., 2013), instream benthic storage of the C4 detritus, along with riparian inputs during winter storm events could explain the C-enriched nature of biofilms in these two springs during both seasons, their lack of contribution to consumer diets, and suggests consumers selected foods of higher quality (cf. Clapcott & Bunn, 2003). The substantially lower C:N ratio observed for epilithic periphyton in Danforth may suggest it was of higher quality and explain why consumer biomass was more dependent on this source of organic matter in this spring during both seasons. Similar to our study, McWilliam-Hughes et al. (2009) predicted that epilithic periphyton would be an important autochthonous food fueling their food webs but concluded, based on strong correlations found between bryophytes and collector-gathers and scrapers, that bryophytes were the most important food source.
Second, bryophytes form a complex substrate which, along with its large surface area, provides protection from shear stress while also trapping detritus and providing a substrate for epiphytic algal growth. Microbes can progressively reduce the cellulose content and increase nitrogen content of these macrophytes, making them more nutritious in detrital form (Fenchel & Jorgensen, 1977; Mann, 1988). Consumption of microbially processed detritus from these plants has been reported elsewhere (Bayley, 1989; Hamilton et al., 1992). Hence, the diversity of potential food sources provided by bryophytes (living and detritus) and their attached periphyton may explain why some authors have reported invertebrate consumer preference for moss and other macrophytes (Minckley, 1963; Stern & Stern, 1969; Glazier, 1991; Suren & Winterbourn, 1991; Alvarez & Peckarsky, 2013). As the isotopic composition of organic matter changes little upon decomposition (Fry & Sherr, 1984), even with microbial colonization, consumer δ 13C values would still track the detrital forms of these macrophytes. This would also explain why crayfish were found to be dependent on moss, as they often feed on living or detrital forms of vascular plant material (Cumberlidge et al., 2014).
Finally, a number of recent studies have found that aquatic consumers actually feed directly on aquatic bryophytes and were important sources of carbon, even for scraper/grazers (Winterbourn et al., 1986; Lodge, 1991; Newman, 1991; Glime, 2006; McWilliam-Hughes et al., 2009). Additionally, snails and caddisflies are known to consume moss from the same genera (Fontinalis) found in our springs (Lohammer, 1954; Winterbourn & Davis, 1976). Fontinalis is not considered nutritionally inferior to other plants (Glime, 2006) even though aquatic mosses are often regarded as having low nutritional value. Regardless of whether consumers ate moss and water cress directly, fed off detrital forms of these plants, or relied on associated periphyton, autochthonous food sources represented the dominant trophic pathway in our springs.
Longitudinal and seasonal trends in the food base
Our results also demonstrated that the relative importance of autochthonous feeding pathways increased significantly along the spring source-springbrook gradient. We had expected a shift from dependence on allochthonous material closer to the spring source to a greater dependence on autochthonous resources (primarily algae) with distance from the spring source. Our data, instead, showed that autochthony was dominant, supporting the contention of Minshall (1978) and others (Bunn et al., 1999; Zah et al., 2001; March & Pringle, 2003; Douglas et al., 2005; McNeely et al., 2007; Lau et al., 2008, 2009; Dudgeon et al., 2010) that the importance of this trophic pathway is often underestimated. Consistent with our predictions, however, autochthony increased from upstream to downstream and was most intense in the spring with the most open canopy.
Finally, our results demonstrate that autochthonous organic matter serves as the main energy base throughout the entire year. This result supports the premise that even during times when leaf litter input makes it the most accessible food source for invertebrate consumers, it is of such poor quality that most select strongly against it in favor of higher-quality autochthonous materials (Marcarelli et al., 2011). Nonetheless, and consistent with our expectations, allochthony did increase slightly during winter, especially in the heavy deciduous-canopied spring. Furthermore, autochthony was highest in Haseltine, the stream with the open canopy, probably because algal primary production was highest in this system. This result illustrates that these systems rely, to some extent, on temporal subsidies of nutrients and organic matter from the terrestrial environment. The disproportional assimilation of autochthonous material reported in this study, however, suggests, as others have found, that resource quality and animal preference are likely mechanisms regulating the effect a subsidy has on a recipient ecosystem, rather than the magnitude of the flux (Baxter et al., 2005; Marcarelli et al., 2011).
Like other headwater ecosystems, springs are highly vulnerable to environmental change (Cantonati et al., 2006). As there are virtually no ecosystems free from human alteration (Vörösmarty et al., 2010), knowledge of ecosystem-level processes such as the fate of terrestrial and instream sources of organic matter in spring food webs should be beneficial in protecting these fragile systems from changes in quantity and composition of organic carbon inputs due to human alteration (e.g., riparian shading, sedimentation; Bunn et al., 1999).
References
Allan, J. D., 1999. Stream Ecology. Structure and Function of Running Waters. Kluwer Academic Publishers, Dordrecht.
Alvarez, M. & B. L. Peckarsky, 2013. The influence of moss on grazers in high-altitude streams: food, refuge or both? Freshwater Biology 58: 1982–1994.
Barquín, J. & R. G. Death, 2006. Spatial patterns of macroinvertebrate diversity in New Zealand springbrooks and rhithral streams. Journal of the North American Benthological Society 25: 768–786.
Baxter, C. V., K. D. Fausch & W. C. Saunders, 2005. Tangled webs: reciprocal flows of invertebrate prey link streams and riparian zones. Freshwater Biology 50: 201–220.
Bayley, P. B., 1989. Aquatic environments in the Amazon Basin, with an analysis of carbon sources, fish production, and yield. In: Dodge, D. P. (ed.), Proceedings of the International Large Rivers Symposium. Canadian Special Publication of Fisheries and Aquatic Science 106: 399-408.
Benke, A. C., A. D. Huryn, L. A. Smock & J. B. Wallace, 1999. Length-mass relationships for freshwater macroinvertebrates in North America with particular reference to the southeastern United States. Journal of the North American Benthological Society 18: 308–343.
Bunn, S. E., P. M. Davies & T. D. Mosisch, 1999. Ecosystem measures of river health and their response to riparian and catchment degradation. Freshwater Biology 41: 333–345.
Bunn, S. E., P. M. Davies & M. Winning, 2003. Sources of organic carbon support the food web of an arid zone floodplain river. Freshwater Biology 48: 619–635.
Bunn, S. E., C. Leigh & T. D. Jardine, 2013. Diet-tissue fractionation of δ15N by consumers from streams and rivers. Limnology and Oceanography 58: 765–773.
Cantonati, M., R. Gerecke & E. Bertuzzi, 2006. Springs of the Alps – sensitive ecosystems to environmental change: from biodiversity assessments to long-term studies. Hydrobiologia 562: 59–96.
Carroll, T. M. & J. H. Thorp, 2014. Ecotonal shifts in diversity and functional traits in zoobenthic communities in karst springs. Hydrobiologia 738: 1–20.
Chamier, A. C., 1991. Cellulose digestion and metabolism in the freshwater amphipod Gammarus pseudolimnaeus Bousfield. Freshwater Biology 25: 33–40.
Clapcott, J. E. & S. E. Bunn, 2003. Can C4 plants contribute to aquatic food webs of subtropical streams? Freshwater Biology 48: 1105–1116.
Cross, W. F., C. V. Baxter, E. J. Rosi-Marshall, R. O. Hall Jr, T. A. Kennedy, K. C. Donner, H. A. Kelly, S. E. Z. Seegert, K. W. Behn & M. D. Yard, 2013. Food-web dynamics in a large river discontinuum. Ecological Monographs 83: 311–337.
Cumberlidge, N., H. H. Hobbs & D. M. Lodge, 2014. Class Malacostraca, order Decapoda. In Thorp, J. H. & D. C. Rogers (eds), Thorp and Covich’s Freshwater Invertebrates, Vol. I, 4th ed., Ecology and General Biology Academic Press, Boston, MA: 797–847.
Danks, H. V. & D. D. Williams, 1991. Arthropods of springs, with particular reference to Canada. Memoirs of the Entomological Society of Canada 155: 203–217.
Delong, M. D. & J. H. Thorp, 2006. Significance of instream autotrophs in trophic dynamics of the Upper Mississippi River. Oecologia 147: 76–85.
Douglas, M. M., S. E. Bunn & P. M. Davies, 2005. River and wetland food webs in Australia’s wet-dry tropics: general principles and implications for management. Marine and Freshwater Research 56: 329–342.
Dudgeon, D., F. K. W. Cheung & S. K. Mantel, 2010. Foodweb structure in small streams: do we need different models for the tropics? Journal of the North American Benthological Society 29: 395–412.
Eminson, D. & B. Moss, 1980. The composition and ecology of periphyton communities in freshwaters. British Phycology Journal 15: 429–446.
Erman, N. A., 2002. Lessons from a long-term study of springs and spring invertebrates. (Sierra Nevada, California, U.S.A.) and implications for conservation and management. Conference Proceedings. Spring-fed Wetlands: Important Scientific and Cultural Resources of the Intermountain Region.
Erman, N. A. & D. C. Erman, 1995. Spring permanence, species richness and the role of drought. In: Ferrington, L.C. Jr., (ed), Biodiversity of Aquatic Insects and Other Invertebrates in Springs. Journal of the Kansas Entomological Society 68: 50–64.
Fenchel, T. M. & B. B. Jorgensen, 1977. Detritus food chains of aquatic ecosystems: the role of bacteria. Advances in Microbiological Ecology 1: 1–58.
Finlay, J. C., 2001. Stable-carbon-isotope ratios of river biota: implications for energy flow in lotic food webs. Ecology 82: 1052–1064.
Finlay, J. C., S. Khandwala & M. E. Power, 2002. Spatial scales of carbon flow in a river food web. Ecology 83: 1845–1859.
Fisher, S. G. & G. E. Likens, 1973. Energy flow in Bear Brook, New Hampshire: an integrative approach to stream ecosystem metabolism. Ecological Monographs 43: 421–439.
Fry, B. & E. Sherr, 1984. 13Cmeasurements as indicators of carbon flow in marine and freshwater ecosystems. Contributions in Marine Science 27: 13–47.
Friberg, N. & D. Jacobsen, 1994. Feeding plasticity of two detritivore-shredders. Freshwater Biology 32: 133–142.
Glazier, D. S., 1991. The fauna of North American temperate cold springs: patterns and hypotheses. Freshwater Biology 26: 527–542.
Glime, J. M., 2006. Bryophytes and herbivory. Cryptogamie Bryologie 27: 191–203.
Hamilton, S. K., W. M. Lewis Jr & S. J. Sippel, 1992. Energy sources for aquatic animals in the Orinoco River floodplain: evidence from stable isotopes. Oecologia 89: 324–330.
Hynes, H. B. N., 1983. Groundwater and stream ecology. Hydrobiologia 100: 93–99.
Jardine, T. D., R. J. Hunt, S. J. Faggotter, D. Valdez, M. A. Burford & S. E. Bunn, 2013. Carbon from periphyton supports fish biomass in waterholes of a wet-dry tropical river. River Research and Applications 29: 560–573.
Kendall, C., S. R. Silva & V. J. Kelly, 2001. Carbon and nitrogen isotopic compositions of particulate organic matter in four large river systems across the United States. Hydrological Processes 15: 1301–1346.
Kesler, D. H., 1983. Variation in cellulase activity in Physa heterostropha (Gastropoda) and other species of gastropods in a New England pond. American Midland Naturalist 109: 280–287.
Kubíkoá, L., O. Simon, K. Tichá, K. Douda, M. Maciak & M. Bíly, 2012. The influence of mesoscale habitat conditions on the macroinvertebrate composition of springs in a geologically homogeneous area. Freshwater Science 31: 668–679.
Lau, D. C. P., K. M. Y. Leung & D. Dudgeon, 2008. Experimental dietary manipulations for determining the relative importance of allochthonous and autochthonous food resources in tropical streams. Freshwater Biology 53: 139–147.
Lau, D. C. P., K. M. Y. Leung & D. Dudgeon, 2009. What does stable isotope analysis reveal about trophic relationships and the relative importance of allochthonous and autochthonous resources in tropical streams? A synthetic study from Hong Kong. Freshwater Biology 54: 127–141.
Leberfinger, K., I. Bohman & J. Herrmann, 2011. The importance of terrestrial resource subsidies for shredders in open-canopy streams revealed by stable isotope analysis. Freshwater Biology 56: 470–480.
Lodge, D. M., 1991. Herbivory on freshwater macrophytes. Aquatic Botany 41: 195–224.
Lohammer, G., 1954. The distribution and ecology of Fissidens julianus in northern Europe. Svensk botanisk tidskrift 58: 162–173.
Lowe, R. L. & G. D. LaLiberte, 2006. Benthic stream algae: distribution and structure. In Hauer, F. R. & G. A. Lamberti (eds), Methods in Stream Ecology. Academic Press, San Diego: 327–339.
Mann, K. H., 1988. Production and use of detritus in various freshwater, estuarine, and coastal marine ecosystems. Limnology and Oceanography 33: 910–930.
Mantel, S. K., M. Salas & D. Dudgeon, 2004. Foodweb structure in a tropical Asian forest stream. Journal of the North American Benthological Society 23: 728–755.
Marcarelli, A. M., C. V. Baxter, M. M. Mineau & R. O. Hall Jr, 2011. Quantity and quality: unifying food web and ecosystem perspectives on the role of resource subsidies in freshwaters. Ecology 92: 1215–1225.
March, J. G. & C. M. Pringle, 2003. Food web structure and basal resource utilization along a tropical island stream continuum, Puerto Rico. Biotropica 35: 82–93.
Mayer, M. S. & G. E. Likens, 1987. The importance of algae in a shaded headwater stream as food for an abundant caddisfly (Trichoptera). Journal of the North American Benthological Society 6: 262–269.
McCutchan, J. H. & W. M. Lewis Jr, 2002. Relative importance of carbon sources for macroinvertebrates in a Rocky Mountain stream. Limnology and Oceanography 47: 742–752.
McNeely, C., J. C. Finlay & M. E. Power, 2007. Grazer traits, competition, and carbon sources to a headwater-stream food web. Ecology 88: 391–401.
McWilliam-Huges, S. M., T. D. Jardine & R. A. Cunjak, 2009. Instream C sources for primary consumers in two temperate, oligotrophic rivers: possible evidence of bryophytes as a food source. Journal of the North American Benthological Society 28: 733–743.
Merritt, R. W., J. B. Cummins & M. B. Berg (eds), 2008. An Introduction to the Aquatic Insects of North America, 4th ed. Kendall/Hunt Publishing Company, Dubuque, IA.
Minckley, W. L., 1963. The ecology of a spring stream Doe Run, Meade County, Kentucky. Wildlife Monographs 11: 1–124.
Minshall, G. W., 1978. Autotrophy in stream ecosystems. Bioscience 28: 767–771.
Newman, R. M., 1991. Herbivory and detritivory on freshwater macrophytes by invertebrates: a review. Journal of the North American Benthological Society 10: 89–114.
Parnell, A. C., R. Inger, S. Bearhop & A. L. Jackson, 2010. Source partitioning using stable isotopes; coping with too much variation. PLoS One 5: e9672. doi:10.1371/journal.pone.0009672.PMID:20300637.
Pimm, S. L., 2002. Food Webs. University of Chicago Press, Chicago, IL.
Pingram, M. A., K. J. Collier, D. P. Hamilton, B. O. David & B. J. Hicks, 2012. Carbon sources supporting large river food webs: a review of ecological theories and evidence from stable isotopes. Freshwater Reviews 5: 85–103.
R Core Team, 2016. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna.
Robinson, C. T., D. Schmid, S. Matthias & S. M. Bernasconi, 2008. Functional measures and food webs of high elevation springs in the Swiss Alps. Aquatic Sciences 70: 432–445.
Rosenfeld, J. S. & J. C. Roff, 1992. Examination of the carbon base in southern Ontario streams using stable isotopes. Journal of the North American Benthological Society 11: 1–10.
Salas, M. & D. Dudgeon, 2001. Stable-isotope determination of mayfly (Insecta: Ephemeroptera) food sources in three tropical Asian streams. Archiv für Hydrobiologie 151: 17–32.
Smith, J., P. J. Wood & J. Gunn, 2003. The influence of habitat structure and flow permanence on invertebrate communities in karst springs systems. Hydrobiologia 510: 53–66.
Stern, M. S. & D. H. Stern, 1969. A limnological study of a Tennessee cold springbrook. American Midland Naturalist 82: 62–82.
Staudacher, K. & L. Füreder, 2007. Habitat complexity and invertebrates in selected Alpine springs (Schütt, Carinthia, Austria). International Review of Hydrobiologia 92: 465–479.
Suren, A. M. & M. J. Winterbourn, 1991. Consumption of aquatic bryophytes by alpine stream invertebrates in New Zealand. New Zealand Journal of Marine and Freshwater Research 25: 331–343.
Teal, J. M., 1957. Community metabolism in a temperate cold spring. Ecological Monographs 27: 283–302.
Thomson, K. C., 1986. Geology of Greene County. Missouri, Watershed Management Coordinating Committee, Springfield, Missouri.
Thorp, J. H. & A. P. Covich (eds), 2010. Ecology and Classification of North American Freshwater Invertebrates. Academic Press, San Diego, CA.
Thorp, J. H., M. D. Delong, K. S. Greenwood & A. F. Casper, 1998. Isotopic analysis of three food web theories in constricted and floodplain regions of a large river. Oecologia 117: 551–563.
Vander Zanden, M. J. & J. B. Rasmussen, 2001. Variation in δ15N & δ13C trophic fractionation: implications for aquatic food web studies. Limnology and Ocenography 46: 2061–2066.
Vannote, R. L., G. W. Minshall, K. W. Cummins, J. R. Sedell & C. E. Cushing, 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37: 130–137.
Von Fumetti, S., P. Nagel, N. Scheifhacken & B. Baltes, 2006. Factors governing macrozoobenthic assemblages in perennial springs in north-western Switzerland. Hydrobiologia 568: 467–475.
Vörösmarty, C. J., P. B. McIntyre, M. O. Gessner, D. Dudgeon, A. Prusevich, P. Green, S. Glidden, S. E. Bunn, C. A. Sullivan, C. R. Liermann & P. M. Davies, 2010. Global threats to human water security and river biodiversity. Nature 467: 555–561.
Wallace, J. B., S. L. Eggert, J. L. Meyer & J. R. Webster, 1997. Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 277: 102–104.
Williams, D. D. & I. D. Hogg, 1988. Ecology and production of invertebrates in a Canadian coldwater spring-springbrook system. Holarctic Ecology 11: 41–54.
Winterbourn, M. J. & S. F. Davis, 1976. Ecology role of Zelandopsyche ingens (Trichoptera: Oeconesidae) in a beech forest stream ecosystem. Australian Journal of Marine and Freshwater Research 27: 197–215.
Winterbourn, M. J., J. S. Rounick & A. G. Hildrew, 1986. Patterns of carbon resource utilization by benthic invertebrates in two British river systems: a stable carbon isotope study. Hydrobiologia 107: 349–361.
Yam, R. S. W. & D. Dudgeon, 2005. Stable isotopes investigation of food use by Caridina spp. (Decapoda: Atyidae) in Hong Kong streams. Journal of the North American Benthological Society 24: 68–81.
Zah, R., P. Burgher, S. M. Bernasconi & U. Uehlinger, 2001. Stable isotope analysis of macroinvertebrates and their food sources in a glacier stream. Freshwater Biology 46: 871–882.
Acknowledgments
We thank R.T. Pavlowsky, Marc Owen, and staff at the Ozarks Environmental and Water Resources Institute (OEWRI) at Missouri State University for their support. Special thanks are owed to spring owners T. Lynch, R. Campbell, and R. Lovett. We appreciate the help of L. Bullard from the Watershed Committee of the Ozarks for his assistance with locating the spring sites and sharing his expertise on spring ecosystems. We thank L.A. Bennett, M.A. Blackwood, and C. Freeman from the University of Kansas and J. Shepard and C. Copper for lab and field assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: David J. Hoeinghaus
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Carroll, T.M., Thorp, J.H. & Roach, K.A. Autochthony in Karst Spring Food Webs. Hydrobiologia 776, 173–191 (2016). https://doi.org/10.1007/s10750-016-2750-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-016-2750-6