
Abstract
Potassium (K+) plays a crucial role in plant homeostasis, and its deficiency significantly impacts photosynthesis, triggering a decrease in growth and crop production. K+ starvation induced a significant reduction in the net photosynthetic rate, and the drop is associated with resistance of CO2 diffusion through stomatal conductance, mesophyll conductance, and lowered carboxylase activity of Rubisco, electron transport rate of PSII, and with many biochemical limitations. The complex interaction of all the above factors contributed to limiting photosynthesis under K+ starved conditions. Low chlorophyll content and poor chloroplast structure may also define photosynthetic processes and causes a decrease in crop growth and productivity under K+ starvation. Under abiotic stresses such as drought, waterlogging, temperature, salinity, and heavy metal, the application of exogenous K+ promotes plant tolerance by activating the antioxidant system, which limits the overproduction of reactive oxygen species (ROS), avoiding the associated oxidative damages where other signaling molecules such as nitric oxide (NO) and hydrogen sulfide (H2S) may involve. The review highlights the decisive role of exogenous and endogenous K+ in modulating diverse physiological and biochemical processes in a healthy and stressful environment. Furthermore, this review appraises the involvement of K+ with another signaling molecule in enhancing abiotic stress tolerance in plants. Therefore, this review provides a comprehensive update on the relevance of K+ in higher plants. Its exogenous application should be a potential tool, especially in crops under adverse environmental conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Potassium is an essential macronutrient and the most abundant cation in plant cells (Pettigrew 2008; Singh and Reddy 2017). It is only found in its ionic form (K+) or in weak complexes from which it can be easily exchanged (Marschner 2012). 2.1–2.3% of the earth’s crust accounts for potassium; nonetheless, the fraction of soil K+ available to plants is only 0.1–0.2% (Britzke et al. 2012; Wedepohl 1995), and the availability of K+ from soil depends on edaphic and some other factors with the capability of plants to take up K+ (Chérel et al. 2014; Hasanuzzaman et al. 2018; Sardans and Peñuelas 2021). K+ uptake from root cells and its mobilization into distinct cellular compartments and organs is facilitated by transporters and channels (Ve’ry et al. 2014; Wang and Wu 2015; Raddatz et al. 2020; Lhamo et al. 2021). K+ in plant cells is carried out by a variety of transporter proteins classified into several families with different structures and transport mechanisms, including channels families such as voltage-dependent shaker-like channels, the tandem-pore (TPK) and two-pore channels (TPC) (Hedrich 2012), and the carrier-like families KT/HAK/KUP (Nieves- Cordones et al. 2014a; Li et al. 2018) HKT uniporters and symporters (Hamamoto et al. 2015), and cation-proton antiporters (CPA).
K+ participates in many physiological processes, viz., photosynthesis and respiration, enzyme activities, water relations, electrical neutralization, assimilate transport, osmoregulation, turgor pressure, protein synthesis, ion homeostasis, anion-cation balance, energy transfer, and utilization (Weng et al. 2007; Maathuis 2009; Zou et al. 2011; Houmani and Corpas 2016; Wang and Wu 2017; Ragel et al. 2019; Srivastava et al. 2020; Cui and Tcherkez 2021; Sardans and Peñuelas 2021; Houmani et al. 2022). Furthermore, K+ indirectly affects plant growth and development (Lu 2016c). All these events can be disrupted by K+ deficiency which is a type of plant abiotic stress that induces a wide range of responses, including oxidative stress, and results in limited crop growth and productivity (Hafsi et al. 2014; Waqas et al. 2021; Houmani et al. 2022; Johnson et al. 2022). K+ starvation makes various responses at different stages, viz., morphological, physiological, biochemical, and molecular (Hafsi et al. 2014). K+ deficiency, also called potash deficiency, leads to decreased chlorophyll content, reducing photosynthetic activity and thus minimizing overall agricultural productivity (Hartt 1969; Pier and Berkowitz 1987; Zhao et al. 2001). Visible symptoms of K+ deficiency take a long time for expression (Kanai et al. 2011); plant growth is reduced and ceases (Mengel and Kirkby 1987). The first noticeable symptom of K+ deficiency is mottled or marginal chlorosis which later develops into necrosis primarily at the leaf tips, margins, and between veins. Still, in several monocotyledonous plants, these necrotic lesions first form at the leaf tips and margins and then extend towards the leaf base. These symptoms first appear on mature leaves at the bottom of a plant due to mobilization of K+ to younger leaves. Stems may be slender, weak, and abnormal short internodal regions of K+ deficient plants (Taiz et al. 2015). Photosynthesis is influenced by K+ deficiency in several ways and affects fruit quality and yield (Kumar et al. 2006; Shen et al. 2017; Zhang et al. 2017a, b).
K+ plays a prominent role in alleviating abiotic stresses, which may have associated with oxidative stress (Trono et al. 2015; Hasanuzzaman et al. 2018; Kumari et al. 2021; Houmani et al. 2022). Plants have evolved strategies to increase K+ uptake while maintaining ROS homeostasis in response to low K+ availability. Nicotinamide adenine dinucleotide phosphate oxidase RHD2/RbohC produces H2O2, a ROS that regulates K+ transporters, particularly the high-affinity potassium transporter HAK5, which is induced under these conditions. Thus, ROS plays an essential role in generating K+ starvation responses at the root level by regulating the expression of the AtHAK5 gene, which encodes a high-affinity K+ transporter that is activated in response to low K+ supply (Kim et al. 2010). During the inadequate K+ response, the member of the type III peroxidase family RCI3 (rare cold-inducible gene 3) also contributes to ROS production (Kim et al. 2010). Furthermore, ROS directly activates the K+ channels AtGORK and AtSKOR, which are involved in K+ efflux from cells during stress (Demidchik et al. 2010; GarciaMata et al. 2010; Demidchik 2018). Potassium starvation causes oxidative stress that increases antioxidant and NADPH-generating systems, thus allowing the halophytes to survive (Houmani et al. 2022). Thus, K+ emerged as a potential anti-stressor molecule due to its protective functions under different plant abiotic stress, viz., salt, drought, temperature, and heavy metal.
The present review focuses on the efficient role of K+ in improving chlorophyll content, net photosynthetic rate, chloroplast morphology, and ultrastructure. An attempt has been made to cover the contribution of K+ in increasing enzymatic antioxidant machinery such as catalase (CAT), peroxidase (POX), superoxide dismutase (SOD), ascorbate peroxidase (APX), and glutathione reductase (GR) in reducing ROS and MDA content and maintaining osmotic content. The current review also highlights the influential role of K+ in mitigating environmental stresses like salinity, drought, waterlogging, temperature, and heavy metal stress with the help of several signaling molecules. Considering the recent reviews focused on K+ transporters and channels (Santa-María et al. 2018; Jegla et al. 2018; Ali et al. 2021; Lhamo et al. 2021; Dave et al. 2022), the present assessment will be focused on K+ deficiency and its effect on photosynthesis and ROS metabolism, and highlighting its possible connections with other signaling molecules such as NO and H2S.
Potassium in the rhizosphere
In the lithosphere, K+ is the fourth most abundant element; nonetheless, a low proportion is available for plants (Mengel and Kirkby 1982). Four distinct pools of K+ in soil: (i) ionic form in soil solution (0.1–0.2%); (ii) exchangeable or readily available (1–2%); (iii) non-exchangeable, slowly available or fixed (fixed in 2:1 clays 1–10%); and, (iv) unavailable or structural (96–99%) (Fig. 1) (Sparks 1987). The first two pools of K+ are reversible and known to meet the instantaneous demand of growing plants, while the last two are irreversible and known for the long-term supply of K+ to plants (Askegaard et al. 2003). When the availability of K+ is low in the soil, the ionic form is the source used by the roots to provide the needs of K+ for plant growth as well as its replenishment when the soil is deficient in K+ that is in equilibrium between the interchangeable and non-interchangeable (Hinsinger et al. 2021). Leaching is one of the factors responsible for the K+ depletion from the reversible pool; therefore, the application of K+ fertilizer will raise the K+ availability for plants (Sparks and Huang 1985; Mouhamad et al. 2016). A major portion of K+ in the soil is in mineral pools such as muscovite, biotite, or feldspars, which are released over a long time due to weathering or breaking down that refills the exchangeable and fixed pool (Römheld and Kirkby 2010; Mouhamad et al. 2016). The exchangeable or readily available K+ is the fraction of the K+ that is surface-adsorbed on negatively charged sites of clay minerals and organic matter and in a rapid equilibrium to soil solution (Sardans and Peñuelas 2015). A minor amount of K+ is found in organic matter because it is not a component of biomolecules, thus rapidly and simply leached from leaves because of its high solubility. Hence low fraction of K+ is directly available to plants (Wang et al. 2010a, b; Britzke et al. 2012). Non-exchangeable pool slowly released K+ from the wedge zones of weathered micas and vermiculites when the level of soil solution and readily available K+ decrease by crop removal and leaching (Sparks et al. 1980; Sparks 2000; Mengel and Kirkby 2001). The sequence of availability of K+ for plant uptake and microorganisms is soil solution > readily available (exchangeable) > slowly available (fixed or non-exchangeable) > unavailable (minerals) (Sparks and Huang 1985; Sparks 1987, 2000) (Fig. 1).

Interrelationship of various forms of soil K+ (modified from Sparks 1987) which includes (i) ionic form in soil solution (0.1–0.2%); (ii) exchangeable or readily available (1–2%); (iii) non-exchangeable, slowly available or fixed (fixed in 2:1 clays 1–10%); and, (iv) unavailable or structural (96–99%)
K+ uptake and transportation within the plant
To facilitate the uptake and transportation of K+ to different cellular compartments, multiple K+ channels and transporters are found mainly in the cell membrane of plant root cells (Ashley et al. 2006; Coskun et al. 2014). Two discrete kinetic systems accomplished the task of K+ uptake via roots depending on affinity for K+ that switch according to the availability of external K+ i.e. Soil (Epstein et al. 1963): (i) High-affinity system (transporters) activated at the lower level of K+ concentration (< 0.2 mmol/L); and (ii) Low-affinity system (channels) activated at a higher level of external K+ concentration (> 0.3 mmol/L).
K+ channels comprise three different classes based on their structural characteristics: (i) Shaker-type channels (voltage-dependent); (ii) Two pore channels (TPC); and (iii) Cyclic nucleotide-gated channels (reviewed by Ashley et al. 2006). Shaker-type K+ channels are also classified as (i) Inward rectifying K+ channels, (ii) Outward-rectifying K+ channels, and (iii) Weakly rectifying channels (Lefoulon 2021; Wang and Wu 2013).
K+ transporters include three distinct classes: (i) K+ uptake permease (KT/HAK/KUP); (ii) High-affinity K+ transporter (HKT); (iii) Cation-proton antiporter (CPA) families (Gierth and Mӓser 2007; Nieves-Cordones et al. 2014; Ve’ry et al. 2014; Li et al. 2018). The first functionally characterized K+ channels in plants belongings to the Shaker family were the Arabidopsis K+ transporter 1 (AtAKT1) and the K+ channel 1 (AtKC1). They interact to form a heteromeric K+ channel that regulates the AtAKT1 activity, thus preventing K+ leakage under low K+ concentration in root cells (Reintanz et al. 2002; Duby et al. 2008; Geiger et al. 2009; Honsbein et al. 2009; Wang et al. 2010a, b, 2016). Under low K+ concentration, two high K+ affinity root proteins (AKT1and HAK5) mediate the sufficient K+ uptake to support plant growth (Pyo et al. 2010). The transcription level of the HAK gene was upregulated in low K+ content (Santa-Maria et al. 1997; Armengaud et al. 2004; Shin and Schachtman 2004; Gierth et al. 2005; Jung et al. 2009; Li et al. 2018), while AtKUP7 participated in K+ uptake in low to moderate external K+ content (~ 0.1 mmol/L) with the effect in translocation from root to shoot (Han et al. 2016).
For long-distance transport, after K+ absorption into the root cells, K+ is transported to root stele tissues and then transported to shoots through xylem vessels. The Stelar K+ Outward Rectifier (SKOR) member of Shaker type outward rectifying K+ channel in stele tissues mediates K+ translocation from root parenchyma cells to xylem sap for K+ transport to shoots (Gaymard et al. 1998; Liu et al. 2006). On the other hand, stomatal movement depends on the K+ content in the guard cells; thus, the guard cells outward rectifying K+ channel (GORK) regulates K+ efflux via membrane depolarization to close the stomata (Ache et al. 2000; Becker et al. 2003; Adem et al. 2020), whereas KAT1 and KAT2 inward channels allow the K+ influx via membrane hyperpolarization into the guard cell to open the stomata (Schachtman et al. 1992; Pilot et al. 2001; Hosey et al. 2003). The vacuole act as a reservoir of K+ ions in plant cells, and under K+ starvation, the ions translocate to the cytoplasm to sustain the K+ content of the cytoplasm (Walker et al. 1996). To maintain homeostasis, K+ translocates between the vacuole and cytoplasm; the AtTPK1 channel found in the tonoplast facilitates the K+ transport necessary for several physiological processes like stomatal movement and seed germination (Czempinski et al. 2002; Gobert et al. 2007). Moreover, tonoplast NHX transporters (AtNHX1 and AtNHX2) act as Na+(K+)/H+ antiporters and facilitate K+ uptake into the vacuole, thus maintaining the K+ vacuolar pool (Venema et al. 2002; Apse et al. 2003; Bassil et al. 2011; Barragan et al. 2012; Andres et al. 2014). Owing to its high mobile nature, K+ recirculates between roots and shoots through the xylem and phloem (Ahmad and Maathuis 2014). Plant tissues load the K+ to phloem with the availability of a sufficient amount of ATP to reach a notable concentration, and K+ comes to the requiring organs with the flow of sap while it can also return to roots (Jeschke et al. 1985; Dreyer and Blatt 2009). AKT2 inward rectifying channel, another member of the shaker family mainly found in the phloem of both leaves and roots (Deeken et al. 2000; Lacombe et al. 2000b) plays two significant roles in loading of K+ in source tissues and sink organs, respectively (Gajdanowicz et al. 2011). At the subcellular level, the K+ Efflux Antiporter (KEA) protein group participates in its homeostasis. In Arabidopsis, two KEA subgroups, KEA1-3, are associated with plastid, and KEA4-6 seems to be connected with the endomembrane system (Tsujii et al. 2019; Wang et al. 2019; DeTar et al. 2021). The main K+ channels and transporters are given in Table 1.
Impact of potassium on photosynthesis
K+ affects photosynthesis at various levels and influences various physiological processes, plant metabolism, and osmotic adjustments (Severtson et al. 2016). Under different levels of K+ photosynthetic capability (Bednarz et al. 1998; Zhao et al. 2001; Basile et al. 2003; Weng et al. 2007; Gerardeaux et al. 2009), physiological and biochemical characteristics of the plants are different (Chartzoulakis et al. 2006). Effects of K+ starvation on the plant’s photosynthetic features are described in Fig. 2.

Summary of the main effects on photosynthesis trigger by K+ starvation. ↑increase and ↓decrease
Chlorophyll
Chlorophyll (Chl) is a light-absorbing green pigment in the chloroplast that converts light to chemical energy in plants (Kalaji et al. 2017). Under K+ starvation, chlorophyll synthesis is inhibited in Eucalyptus grandis (Battie-Laclau et al. 2014). Leaf chlorosis is an early symptom of chlorophyll degradation due to oxidative stress of excessive production of ROS in K+ deficiency (Cakmak 2005; Lu et al. 2019). Lower chlorophyll content and Chl. a/b ratio is the stress symptoms of chloroplast (Jia et al. 2008), changes in morphology and anatomy of chloroplast were associated with chlorophyll content and photosynthesis capability (Tian et al. 2008; Lu et al. 2019), chlorophyll composition change in low K+ stress affects the photosynthesis (Jia et al. 2008). Chlorophyll fluorescence gives accurate information about the condition of photosynthetic machinery and mainly of PSII (Sun et al. 2015). Potential quantum efficiency (Fv/Fm), quantum yield of photosystem II (ΦPSII), photochemical quenching (qP), and electron transport rate (ETR) were markedly reduced under K+ deficiency in Zea mays L. (Qi et al. 2019). It has been reported in several studies that total chlorophyll concentration is reduced under K+ starvation (Bednarz and Oosterhuis 1999; Zhao et al. 2001; Lu et al. 2016b), and the chlorophyll concentration are directly related to photosynthesis (Battie-Laclau et al. 2014). The effect of K+ starvation on chlorophyll is also presented in Table 2.
Chloroplast morphology and ultrastructure
Stabilized morphology and structure of chloroplast contribute resistance to severe impacts of K+ deficiency stress (Jia et al. 2008; Shao et al. 2016), while changes in morphology and structure are notably associated with chlorophyll content and photosynthetic capacity (Tian et al. 2008; Lu et al. 2019). K+ facilities the correct organization of grana and stroma lamellae; however, under K+ deficiency, the ultrastructure is disorganized, and the chloroplast lamella is lopsided, limp, and partly dispersed in the cytoplasm that, triggers a reduction in the number of grana and lamellae (Zhao et al. 2001; Jia et al. 2008). In maize, K deficiency causes a reduction in the number of chloroplasts, which take on an irregular shape, as the outer surface changes from long and oval to elliptical or almost round, thus reducing the length/width ratio. In the corn inbred line 90-21-3, the outer membrane and thylakoids remain the same; the plastoglobule content increases and is swollen. On the other hand, in the inbred line D937, the ultrastructure of chloroplasts was severely damaged with irregular shape and significantly aggregated. At the same time, the swollen plastoglobules and lamellae structure were not typical and mainly scattered in the cytoplasm under K starvation (Qi et al. 2019). In the genotype of Ipomoea batatas, leaves showed big starch granules, broken and indistinguishable chloroplast membrane, grana lamellae limped and distorted, and acute damage in chloroplast ultrastructure under K+ stress (Tang et al. 2015). In another similar study, the chloroplast blade swelled and acquired a ball shape within lots of starch granules under K+ deficiency (Wang et al. 2018). However, a slight difference in chloroplast morphology has been observed in Brassica napus (Hu et al. 2020). In Gossypium hirsutum leaves, poor chloroplast ultrastructure with large starch granules, more and expanded plastoglobuli, little grana, and the thylakoids propels towards periphery likely be the reason for low leaf PN under low K+ stress (Zhao et al. 2001).
Net photosynthetic rate
Net photosynthetic rate (PN) and photosynthetic plant area are the two vital features on which the photosynthetic plant capacity depends. For the enhancement of PN, researchers have been working to increase the potential of yield (Zhu et al. 2004; von Caemmerer et al. 2012; South et al. 2019) and higher PN related to higher biomass (Wang et al. 2012). According to several reports, a decline in the rate of photosynthesis is due to a reduction of chlorophyll content, hindrance in PSII activity, and electron transport (non-stomatal) in K+ deficient plants such as Chinese hickory, cotton, almond tree, and lettuce (Basile et al. 2003; Jin et al. 2011; Wang et al. 2012; Zhang et al. 2017a, b). Table 3 also includes the effects of potassium starvation on the PN.
Limitations in CO2 diffusion during photosynthesis
Three major resistances attributed to decrease net CO2 during photosynthesis under K+ starvation are (i) reduced stomatal conductance, (ii) reduced mesophyll conductance, and (iii) biochemical limitations.
Stomatal conductance (gs)
In the stomatal opening, K+ influx in guard cells is a crucial light-dependent step (Shabala 2003; Inoue et al. 2020), influencing stomatal conductance. It is well documented that the stomatal limitation is associated with K+ concentrations and the stomatal closure under K+ starvation and is often considered the main reason behind the reduction of net PN (Thiel and Wolf 1997; Römheld and Kirkby 2010). Restricted CO2 diffusion through the stomata and mesophyll cells (stomatal conductance and mesophyll conductance) decreased the carboxylation ability, chlorophyll biosynthesis, and assimilate transport (Singh and Reddy 2017). Compared to mesophyll, the photosynthetic rate is generally limited due to stomatal conductance (gs) (Jin et al. 2011). Under K+ starvation, the decrease of gs coincided with the decline in T and Ci in soybean (Singh and Reddy 2017). Still, in Ipomoea batatas (Tang et al. 2015) and Eustoma grandiflorum (Chen et al. 2018), the reduction of gs, PN, and Tr is accompanied by no significant change in Ci. However, a decrease in gs was accompanied by the increase of Ci in maize (Qi et al. 2019), cotton (Zhao et al. 2001) constant or enhanced Ci in hickory seedlings which might be caused by high mesophyll limitations or low capability of the CO2 fixation cycle, rather than to stomatal resistance (Jin et al. 2011). Similar findings had been seen in Prunus dulcis, Gossypium hirsutum L. where stomatal conductance was not the main reason for low PN, and it was reduced via biochemical limitations, little Chl content, poor chloroplast ultrastructure, and restricted saccharide translocation under potassium starvation (Zhao et al. 2001; Basile et al. 2003). The Ji22 genotype of Ipomoea batatas L. plants under K+ deficiency showed a low level of PN, E, gs, and terrible damage to chloroplast ultrastructure in leaf, suggesting that this genotype faced both photoinhibition and stomatal limitations (Wang et al. 2018). In contrast, an enhancement in the stomatal conductance under moderate K+ deficiency in a well-irrigated and water-stressed condition may be attributable to dehydration mainly in water-stressed plants, which ultimately leads to impairment of stomatal closure regulation in Olea europaea L. (Arquero et al. 2006). In mild K+ deficiency, stomatal conductance was the main factor contributing to net photosynthesis change. As the K + deficit became more severe biochemical limitations also contributed to stomatal regulations in the cotton (Gossypium hirsutum L., cv. Deltapine) plant (Bednarz et al. 1998). There is no significant change in transpiration rate and stomatal conductance with a decrease in chlorophyll content and enhanced anthocyanin pigment under K+ deprivation in maize plants. Mg deficiency strongly limits the photosynthetic and transpiration rates, stomatal conductance, photosystem II (PSII) performance, chlorophyll, and flavonol content with concomitantly enhanced anthocyanin content. Furthermore, Ca2+ deficiency also alleviated the rate of photosynthesis and transpiration, stomatal conductance, and PSII activity, whereas the pigment content was not considerably different compared with the control (Sitko et al. 2019). The primary cause of lower photosynthesis in K+ starved plants is a subject of debate due to contrasting observations between the plant species.
Mesophyll conductance (gm)
CO2 dissolves in the cell walls apoplastic water after diffusion via mesophyll cells’ leaf internal air space. Once the CO2 enters the liquid phase, it is found either as dissolved CO2 or HCO3 and further diffuses via the pores of the cell wall, the plasma membrane, the cytosol, the chloroplast envelope and finally reaches the chloroplast stroma, where enzymatic CO2 fixation took place (Tränkner et al. 2018). Moreover, it is reported that under low K+ liquid-phase pathways, conductance decreased by reducing the chloroplast exposed surface area per unit leaf area (Sc/S) and enhancing the resistance of the cytoplasm, thus enhancing the distance between the chloroplast and cell wall and between the adjacent chloroplasts (Lu et al. 2016a, b, c).
Mesophyll diffusion conductance to CO2 is the main photosynthetic trait. The limitation to photosynthesis impeded by gm is significant, and under certain conditions like salinity, temperature, nutrients, light, water and tree length and size, leaf structure and aging, cell wall thickness, and chloroplast distribution can be the most significant photosynthetic limitation (Niinemets et al. 2009a; Han 2011; Whitehead et al. 2011; Zhu et al. 2011; Flexas et al. 2012). Quick response to a change of environmental factors might be regulated by another factor like aquaporin conductance (Flexas et al. 2012). In K+ starved plants, besides gs, reduced leaf mesophyll cell conductance (gm) obstructed the diffusion of CO2 to the chloroplasts (Jin et al. 2011; Battie-Laclau et al. 2014). An alleviated gm (elevated Lm) might be an outcome of the decline of intercellular air space due to the accumulation of photosynthate, decreased carbonic anhydrase activity, and aquaporin channel conductance due to limit CO2 diffusion through mesophyll cells (Cakmak et al. 1994; Zhao et al. 2001; Battie-Laclau et al. 2014; Lu et al. 2016a). It was noted that during photosynthesis, inhibition of net CO2 assimilation (AN) resulted due to reduction in CO2 mesophyll conductance (gm) neither by maximum quantum use efficiency (Fv/Fm) nor by in vivo Rubisco activity under K+ starvation. The reduced gm might be due to variations in the leaf anatomy of Helianthus annuus L. under K+ starvation (Jakli et al. 2017). Leaf area decreased before reduced photosynthetic rates under K+ starvation, and changes in S/W value before Sc/S and Dchl−chl value were the significant reason for the uneven decline in leaf area and photosynthetic rate in Brassica napus plant (Hu et al. 2020). Due to photosynthesis resistance, cucumber leaves were less affected under K+ starvation than rice, wheat, and rapeseed leaves. All species averaged 50.5% and 53.4% of mesophyll conductance accounted for photosynthetic limiting factors under moderate K+ and low K+ conditions, respectively. An increase in leaf hydraulic resistance and RL (ΔRx and ΔRox, the relative growth of xylem and outside-xylem hydraulic resistance, respectively) simultaneously with the rise in K+ deficiency were found to be a significant contributor to photosynthetic limitations, and K+ maintained the leaf photosynthesis and hydraulic conductance through modifications of leaf anatomy (Lu et al. 2019).
Biochemical limitations
The biochemical limitations significantly limit photosynthesis besides stomatal conductance and mesophyll conductance. However, there is considerable discussion on either K+ starvation inhibits photosynthesis directly by reducing leaf chlorophyll content, obstructing photochemical energy conversion or biochemical processes, or inhibiting CO2 diffusion from the atmosphere to the site of carbon assimilation in chloroplasts (Jakli et al. 2017). Rubisco is the primary limiting factor of photosynthesis in rice and tobacco leaves (Yang et al. 2004: Weng et al. 2007; Hu et al. 2021). In Carya cathayensis, the maximum carboxylation rate of Rubisco (Vc, max) and maximum rate of electron transport (Jmax) were the most sensitive factors (Jin et al. 2011). In Raphanus sativa, K+ starvation reduces the photosynthetic rate by disturbing the balance of H+ influx into the thylakoid lumen, thus increasing the acidification that leads to triggering the downregulation mechanism, which detaches the LHCII to Photosystem II (PSII) and an increase in energy dissipation, yet without preventing the PQH2 oxidation at the Cyt b6f complex (Kusaka et al. 2021). Similar results were also found in Glycine max under K+ starvation, the maximal fluorescence (Fm), the maximum quantum efficiency of PSII photochemistry (Fv/Fm), actual photochemical efficiency of PSII (ΦPSII), photochemical quenching (qP), and electron transport rate of PSII (ETR) were considerably decreased with Rubisco with minimal fluorescence (Wang et al. 2015).
The relative contribution of all these three limiting factors under K+ starvation has been explained in the photosynthetic process. It is concluded that the complex interaction of all these factors contributed to limiting photosynthesis. Table 4 illustrates some of these limiting factors under K+ starvation.
H2O2 contents and antioxidant system
In chloroplast thylakoid, PSI and PSII reaction centers are the primary site of reactive oxygen species (ROS) (Asada 2006; Kohli et al. 2019). Several types of stresses trigger the generation of ROS, like nutrient deficiency, high light, high or low temperature, salinity, drought, and pathogen attack (Tripathy and Oelmuller 2012). Due to abiotic stress, stomatal closure limits the CO2 uptake, which induces the production of H2O2 during photorespiration by the peroxisomal glycolate oxidase (Leterrier et al. 2016; Corpas et al. 2020) or singlet oxygen by the reduced photosynthetic electron transport chain (Apel and Hirt 2004; Noctor et al. 2014). If the production of H2O2 is prolonged, this will trigger oxidative stress, which is accompanied by altering the cellular redox homeostasis. ROS accumulation disrupts the functioning of the cell and induces the oxidative damage of membrane, lipids, nucleic acid, proteins, and inhibition of enzymes (Mittler 2002; Shahbaz et al. 2008; Ashraf 2009; Akram et al. 2012; Perveen et al. 2010, 2011, 2012). Some evidence indicates that H2O2 might function as a signaling molecule under K+ deficiency (Hernández et al. 2012). Abiotic and biotic stresses are accompanied by the generation of H2O2 consequence of the dismutation of superoxide radicals (O2•−) generated by the NADPH oxidase, also known as respiratory burst oxidase homolog (RBOH), which is considered to be the most crucial enzyme responsible for this oxidative burst (Mittler et al. 2004; Sagi and Fluhr 2006; Chapman et al. 2019). Deprivation of K+ in Arabidopsis enhanced ROS accumulation at distinct regions of roots just behind the elongation zone that is active in K+ uptake and translocation (Shin and Schachtman 2004). Gene RCI3, a member of the type III peroxidase family, is upregulated under K+ starvation and engaged in ROS production, which affects the regulation of AtHAK5 expression in Arabidopsis roots (Kim et al. 2010). To combat oxidative stress, plants have developed a defense system associated with a noticeable increase in ROS metabolizing enzymes like superoxide dismutase (SOD), catalase (CAT), and enzymes of the ascorbate-glutathione cycles (Mittler 2002; Cakmak 2005; Ashraf 2009; Ahmad et al. 2014; Kohli et al. 2019; Houmani et al. 2022). Many reports suggest that under K+ starvation, the more vital photosynthetic ability is linked to higher antioxidant enzymes (Chen et al. 2008; Jia et al. 2008).
In Gossypium hirsutum L., higher H2O2 and ascorbic acid (ASC) activity were observed, which brought about higher malondialdehyde (MDA) content despite the elevated activity of H2O2-scavenging enzymes like catalase and peroxidase. However, low dehydroascorbate reductase activity and higher ASC content were attributed to lower ascorbate peroxidase (APX) activity in K + starvation, leading to premature leaf senescence marked by chlorophyll degradation and negative chlorophyll fluorescence (Hu et al. 2016). In Brassica juncea plants, initiation of ROS leads to oxidative stress and an increase in antioxidant enzymes like SOD, CAT, APX, and glutathione reductase (GR) under potassium deficiency (Ahmad et al. 2014). Similarly, Houttuynia cordata plant showed H2O2 accumulation inducing oxidative stress with a concomitant increase of the activity of antioxidant enzymes like CAT and peroxidase to palliate the oxidative damages (Xu et al. 2011) under the deficiency of K+.
Involvement of K+ to abiotic stress tolerance and its potential relationship with signal molecules such as NO and H2S
It was noted that K+ plays a vital role as an anti-stress agent against different abiotic stresses (Anschütz et al. 2014; Jia et al. 2018; Perelman et al. 2022), such as drought, temperature, salinity, waterlogging, and heavy metal. Furthermore, new evidence supports the correlation between K content with signaling molecules such as NO and H2S (Zhao et al. 2018; Oliferuk et al. 2020; Lana et al. 2021).
Drought stress
Besides growth and productivity, K+ was also involved in maintaining cell turgidity, osmotic adjustment, stomatal regulation, aquaporin channel conductivity, cell membrane stability, and detoxification of ROS under drought stress (Wang et al. 2013; Hasanuzzaman et al. 2018; Sehar et al. 2021). Low K+ alleviated the plant tolerance to drought stress and K+ absorption (Wang et al. 2013). Reports showed that the K+ application reduces drought stress consequences, specifically in Hibiscus rosa-sinensis (Egilla et al. 2005; Bahrami-Rad and Hajiboland 2017) concluded that applying K+ under water-stressed and K+ starved conditions on Nicotiana rustica L. induced dehydration avoidance and tolerance by augmenting stomatal resistance and also by the accumulation of organic osmolytes and proline mainly in the young leaves.
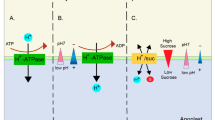
In maize, the overexpression of inwardly rectifying K+ channel 1 (AKT1) facilities the root K+ uptake but, at the same time, mediates the homeostasis of H2O2 and NO, mediating its drought stress tolerance (Feng et al. 2020). More recently, it has proposed a cascade of events associated with the loss of function of chloroplast KEA1 and KEA2 in Arabidopsis, which triggered an increased tolerance to drought stress. Thus, the K+ imbalance in the chloroplasts of this kea1kea2 mutant causes impairment in chloroplast functions, including photosynthesis and photorespiration pathways associated with an imbalance of ROS and NO metabolism affects stomatal movement causing a higher drought stress resilience (Sánchez-McSweeney et al. 2021) unexpectedly. Thus, this new evidence supports the biochemical crosstalk of K+ with NO metabolism and other plant gasotransmitters such as H2S and CO (Zhang et al. 2018; Lana et al. 2021). It should be considered that these signaling molecules participate in a wide range of physiological processes and respond to environmental stresses (Mukherjee and Corpas 2020; Mishra et al. 2021) that would support the benefits exerted by K+ when it is applied as exogenous. Thus, in tomato seedlings, the exogenous application of K+ in coordination with endogenous H2S can regulate H+-ATPase activity, antioxidant system, sugar metabolism, and redox homeostasis that promote drought tolerance stress (Siddiqui et al. 2021).
Waterlogging stress
Waterlogging is one of the significant abiotic stresses that influence plant growth and yield worldwide (Sasidharan et al. 2018). Waterlogging causes inhibition of soil aeration and limits the supply of oxygen (hypoxia or anoxia) to roots (Amin et al. 2015). A lesser amount of oxygen impeded the respiration process in roots and resulted in a shortage of energy required for nutrient uptake (Boru et al. 2003). In waterlogged conditions, a combined supply of K+ (foliar and soil) not only improved growth, photosynthetic pigments, and photosynthetic capacity but also mitigated the negative influence of waterlogging in Gossypium hirsutum L. (Ashraf et al. 2011). Stomatal closure during early senescence is attributed to alleviating PN, and during the late senescence stage, low CO2 assimilation accounted for a low PN. The supply of K+ improved the post-anthesis photosynthesis and yield by maintaining the stomatal openings and CO2 carboxylation capacity by dissipating excess excitation energy, thus reducing the risk of photo-damage in winter wheat at the stem elongation stage under waterlogging conditions (Gao et al. 2021). Recovering root development and yield loss under waterlogged conditions by the combined supply of K+ and N in Vigna radiata L. was observed (Amin et al. 2015).
Temperature stress
Extreme high and low-temperature stress influenced plant growth and development at the whole-plant level, tissue, and cell level, and even at the subcellular level. Direct and indirect injuries due to high and low temperatures reduced the yield (Waraich et al. 2012), inhibited metabolic reactions, and influenced the osmotic and oxidative stresses (Airaki et al. 2012; Wang et al. 2013). Proper plant nutrient management is one of the master strategies for temperature stress tolerance, and K+ plays a significant role in elevating temperature stress tolerance (Hasanuzzaman et al. 2018). Under chilling and freezing stress, the plant faces dehydration due to the loss of apoplastic water. The application of K+ under chilling and freezing stress alleviates the freezing-induced cellular dehydration and adjusts the osmotic potential of plant species (Wang et al. 2013). In low K+ plants, chilling and frost might damage the photosynthetic activities and increase ROS production by reducing the antioxidant activities, thus causing a reduction in plant growth and yield. Exogenous application of high KNO3 enhanced the antioxidant activities and secondary metabolite transcripts, protecting ROS overproduction and increasing chilling stress tolerance in Panax ginseng (Devi et al. 2012). K+ fertilization augmented the frost tolerance as noted by lowering electrolyte leakage mainly in young leaves of Albizia plurijuga, Cedrela dugesii, and Ceiba aesculifolia (Gómez-Ruiz et al. 2016).
High temperature contributed directly to protein denaturation and aggregation, lipid membrane damage, enzymes inactivation of chloroplast and mitochondria, protein synthesis limitation and degradation, and loss of membrane integrity (Waraich et al. 2012). It also enhanced the leaf senescence and negatively influenced the production of photosynthates, thus reducing the yield (Rahman et al. 2005). In wheat, applying potassium orthophosphate contributed to enhanced yield and quality under heat stress conditions (Rahman et al. 2014). In plants, exogenous application of K+ might be helpful to the reduced negative influence of heat stress (Waraich et al. 2012). Upregulation of antioxidant enzymes like SOD, POX, CAT, AsA, phenolics, and MDA enhanced heat stress tolerance in cotton (Sarwar et al. 2019).
Salinity stress
Salinity and K+ starvation are two significant environmental constraints of fields for many crops that generally occur concomitantly and result in loss of plant growth and productivity (Hafsi et al. 2017). Salt stress impedes plant growth, and K+ deficiency further augments the effects of salinity stress on photosynthetic activity. However, it also displays a mechanism of protecting photosynthetic apparatus by non-photochemical quenching and antioxidant activity. Higher accumulation of phenolic compounds under both stresses might be related to antioxidative defense in Sulla carnosa seedlings (Hafsi et al. 2017). In another study, a significant reduction in biomass production under both stresses was noted, and salinity stress impacted more on shoots than roots while potassium starvation the vice versa.
Moreover, the impact of salinity stress on photosynthetic machinery was further enhanced by the deficiency of K+ with the resistance in CO2 photo assimilation. Still, it provided an understanding of protecting PSII photo-damage and dissipation excess energy in Hordeum vulgare (Degl’Innocenti et al. 2009). Oxidative stress is one of the consequences of salinity stress, and the application of K+ under salinity stress alleviated the ROS production by decreasing the NADPH oxidases activity; however, under low K+ supply in bean root cells, the movement of NADPH oxidases was up to eightfold higher in plants compared to K+ sufficient plants (Cakmak 2005). Net uptake rates and translocation of K+ were reduced under salinity stress, while the impact of salinity stress was enhanced with low K+ concentration in Zea mays L. (Botella et al. 1997). Under salinity stress, plants usually reorganize Na+ and K+ contents accompanied by NO accumulation. In Arabidopsis thaliana mutants, it has been demonstrated that NO negatively modulates the K+ channel AKT1, responsible for K+ uptake, by regulating the biosynthesis of vitamin B6 (Xia et al. 2014). Recently, it has been shown that the overexpression of the K+, Na+/H+ antiporter NHX4 in tomato plants improved the salinity tolerance and increased the yield and quality of its fruits (Maach et al. 2020).
Furthermore, in Malus hupehensis seedlings, the pretreatment with H2S palliates salinity and alkaline stress by regulating the Na+/K+ homeostasis because it decreases the root Na+ content, whereas it increases K+ content. This was accompanied by an increase in the activity of antioxidant enzymes, including CAT, POX, and SOD which mitigate oxidative stress (Li et al. 2020). On the other hand, in poplar plants under salinity stress, it has been found that exogenous H2S regulates K+/Na+ homeostasis in roots through the positive control of the H+-ATPases (Zhao et al. 2018). Table 5 provides some representative examples of the beneficial effects triggered by the exogenous application of K+ and how it can mitigate salinity-induced damages (Kumari et al. 2021).
Heavy metal stress
Heavy metal and metalloid stress in the soil is one of the major impediments to the successful establishment and cultivation of crops (Yasin et al. 2018). Metal stress causes a reduction in growth, fruit formation, and development of fruit trees (Worthington 2001; Foyer and Noctor 2005; Lombardi and Sebastiani 2005), and disturbs the biochemical processes, including photosynthetic activity and level of gas exchange, proline production, and K+ nutrition (Lysenko et al. 2015; Song et al. 2015). However, the K+ application benefits plant tolerance to different metal stresses (Dhiman et al. 2022). Prunus persica seedlings treated with exogenous K+ (10 mM) not only improved the status of endogenous K+ but also showed a positive response in alleviating the reduction of zinc-induced photosynthesis and antioxidant defense systems. K+ application was found to upregulate the transcript level of K+ uptake genes (KUP) engaged in K+ acquisition, transport, and homeostasis leading to the protection of photosynthesis and the antioxidative defense system (Song et al. 2015).
Moreover, combining K+ and Ca2+ enhanced the antioxidant enzyme activity in Cd stressed faba bean (Vicia faba L.) seedlings, leading to enhanced plant growth and productivity (Siddiqui et al. 2012). Similarly, in Gladiolus grandiflora L., a combination of K+ and Si mitigate the Cd stress and enhances the protein and proline accumulation and uptake of mineral nutrients (Ca, Mg, Mn, S, and K) and quantity of total phenolics and flavonoids (Zaheer et al. 2018). Similar results were also observed in Gladiolus grandiflora L. under Cd stress by the application of K only (Yasin et al. 2018). An increase in growth, photosynthesis, and nutrient uptake, as well as a reduction in cadmium toxicity, was observed in soybean (Shamsi et al. 2010; Liu et al. 2013) reported that deficiency of K+ enhanced the antioxidant system, which protects the toxicity of Cd in rice seedlings.
Conclusions and future perspectives
K+ is an essential macronutrient involved in both physiological and biochemical processes. K+ starvation is a critical nutritional issue that negatively affects plant growth and yield. This review gives a better understanding of K+ presence in the rhizosphere and its uptake by roots via several kinds of transporter and channels. It is well documented that K+ plays a significant role in photosynthesis, chlorophyll content, and chloroplast ultrastructure. Furthermore, K+ starvation decreased photosynthesis by limiting CO2 diffusion into and through leaves (gs, gm, and biochemical processes). Stomatal resistance, mesophyll resistance, and biochemical limitations are the major factors for the decrease in photosynthesis. Recent investigations also show that K+ promotes antioxidant defense machinery and osmolyte content, which reduces the oxidative stress produced by ROS and MDA, which further helps attenuate abiotic stress.
K+ is an essential macronutrient, and its starvation reduces several physiological and biochemical processes, such as photosynthesis, chlorophyll content, and chloroplast ultrastructure. Furthermore, unraveling the contribution of K+ in modulating genes involved in photosynthesis need to be explored. More studies are required on the genes and signaling molecule that alters plants’ metabolic and cellular responses during stress.
References
Ache P, Becker D, Ivashikina N, Dietrich P, Roelfsema MR, Hedrich R (2000) GORK, a delayed outward rectifier expressed in guard cells of Arabidopsis thaliana, is a K+-selective, K+-sensing ion channel. FEBS Lett 486:93–98. https://doi.org/10.1016/S0014-5793(00)02248-1
Adem GD, Chen G, Shabala L, Chen ZH, Shabala S (2020) GORK channel: a master switch of plant metabolism? Trends Plant Sci 25:434–445
Adhikari B, Dhungana SK, Kim ID, Shin DH (2020) Effect of foliar application of potassium fertilizers on soybean plants under salinity stress. J Saudi Soc Agric Sci 19:261–269. https://doi.org/10.1016/j.jssas.2019.02.001
Ahmad I, Maathuis FJ (2014) Cellular and tissue distribution of potassium: physiological relevance, mechanisms and regulation. J Plant Physiol 171:708–714. doi: https://doi.org/10.1016/j.jplph.2013.10.016
Ahmad P, Ashraf M, Hakeem KR, Azooz MM, Rasool S, Chandna R, Akram NA (2014) Potassium starvation-induced oxidative stress and antioxidant defense responses in Brassica juncea. J Plant Interact 9:1–9. doi: https://doi.org/10.1080/17429145.2012.747629
Airaki M, Leterrier M, Mateos RM, Valderrama R, Chaki M, Barroso JB, Del Río LA, Palma JM, Corpas FJ (2012) Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant Cell Environ 35(2):281–295
Akram MS, Ashraf M (2011) Exogenous application of potassium dihydrogen phosphate can alleviate the adverse effects of salt stress on sunflower. J Plant Nutr 34:1041–1057. https://doi.org/10.1080/01904167.2011.555585
Akram MS, Athar HR, Ashraf M (2007) Improving growth and yield of sunflower (Helianthus annuus L.) by foliar application of potassium hydroxide (KOH) under salt stress. Pak J Bot 39:769–776
Akram NA, Ashraf M, Al-Qurainy F (2012) Aminolevulinic acid-induced regulation in some key physiological attributes and activities of antioxidant enzymes in sunflower (Helianthus annuus L.) under saline regimes. Sci Hort 142:143–148. https://doi.org/10.1016/j.scienta.2012.05.007
Ali A, Raddatz N, Pardo JM, Yun DJ (2021) HKT sodium and potassium transporters in Arabidopsis thaliana and related halophyte species. Physiol Plant 171:546–558
Amin MR, Karim MA, Khaliq QA, Islam MR, Aktar S (2015) Effect of nitrogen and potassium on the root growth, nutrient content and yield of mungbean (Vigna radiata L. Wilczek) under waterlogged condition. Agriculturists 13:67–78. https://doi.org/10.3329/agric.v13i1.26549
Andres Z, Perez-Hormaeche J, Leidi EO, Schlucking K, Steinhorst L, McLachlan DH, Schumacher K, Hetherington AM, Kudla J, Cubero B, Pardo JM (2014) Control of vacuolar dynamics and regulation of stomatal aperture by tonoplast potassium uptake. Proc Natl Acad Sci USA 111:1806–1814. https://doi.org/10.1073/pnas.1320421111
Anschütz U, Becker D, Shabala S (2014) Going beyond nutrition: regulation of potassium homoeostasis as a common denominator of plant adaptive responses to environment. J Plant Physiol 171:670–687
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55:373–399. https://doi.org/10.1146/annurev.arplant.55.031903.141701
Apse MP, Sottosanto JB, Blumwald E (2003) Vacuolar cation/H+ exchange, ion homeostasis, and leaf development are altered in a T-DNA insertional mutant of AtNHX1, the Arabidopsis vacuolar Na+/H+ antiporter. Plant J 36:229–239. https://doi.org/10.1046/j.1365-313X.2003.01871.x
Armengaud P, Breitling R, Amtmann A (2004) The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol 136:2556–2576. https://doi.org/10.1104/pp.104.046482
Arquero O, Barranco D, Benlloch M (2006) Potassium starvation increases stomatal conductance in olive trees. HortScience 41:433–436. https://doi.org/10.21273/HORTSCI.41.2.433
Asada K (2006) Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol 141:391–396. https://doi.org/10.1104/pp.106.082040
Ashley MK, Grant M, Grabov A (2006) Plant responses to potassium deficiencies: a role of potassium transport proteins. J Exp Bot 57:425–436. https://doi.org/10.1093/jxb/erj034
Ashraf M (2009) Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol Adv 27:84–93. https://doi.org/10.1016/j.biotechadv.2008.09.003
Ashraf MA, Ahmad MSA, Ashraf M, Al-Qurainy F, Ashraf MY (2011) Alleviation of waterlogging stress in upland cotton (Gossypium hirsutum L.) by exogenous application of potassium in soil and as a foliar spray. Crop Pasture Sci 62:25–38. https://doi.org/10.1071/CP09225
Askegaard M, Eriksen J, Olesen JE (2003) Exchangeable potassium and potassium balances in organic crop rotations on coarse sand. Soil Use Manag 19:96–103. https://doi.org/10.1111/j.1475-2743.2003.tb00287.x
Bahrami-Rad S, Hajiboland R (2017) Effect of potassium application in drought-stressed tobacco (Nicotiana rustica L.) plants: comparison of root with foliar application. Ann Agric Sci 62:121–130. https://doi.org/10.1016/j.aoas.2017.08.001
Barragan V, Leidi EO, Andres Z, Rubio L, De Luca A, Fernandez JA, Cubero B, Pardo JM (2012) Ion exchangers NHX1 and NHX2 mediate active potassium uptake into vacuoles to regulate cell turgor and stomatal function in Arabidopsis. Plant Cell 24:1127–1142. https://doi.org/10.1105/tpc.111.095273
Basile B, Reidel EJ, Weinbaum SA, DeJong TM (2003) Leaf potassium concentration, CO2 exchange and light interception in almond trees (Prunus dulcis (Mill) D.A .Webb). Sci Hortic 98:185–194. https://doi.org/10.1016/S0304-4238(02)00214-5
Bassil E, Ohto MA, Esumi T, Tajima H, Zhu Z, Cagnac O, Belmonte M, Peleg Z, Yamaguchi T, Blumwald E (2011) The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant Cell 23:224–239. https://doi.org/10.1105/tpc.110.079426
Battie-laclau P, Laclau JP, Beri C, Mietton L, Muniz MRA, Arenque BC, de cassia Piccolo M, Jordan‐meille L, Bouillet JP, Nouvellon Y (2014) Photosynthetic and anatomical responses of Eucalyptus grandis leaves to potassium and sodium supply in a field experiment. Plant Cell Environ 37:70–81. https://doi.org/10.1111/pce.12131
Becker D, Hoth S, Ache P, Wenkel S, Roelfsema MR, Meyerhoff O, Hartung W, Hedrich R (2003) Regulation of the ABA-sensitive Arabidopsis potassium channel gene GORK in response to water stress. FEBS Lett 554:119–126. https://doi.org/10.1016/S0014-5793(03)01118-9
Bednarz CW, Oosterhuis DM (1999) Physiological changes associated with potassium deficiency in cotton. J Plant Nutr 22:303–313. https://doi.org/10.1080/01904169909365628
Bednarz CW, Oosterhuis DM, Evans RD (1998) Leaf photosynthesis and carbon isotope discrimination of cotton in response to potassium deficiency. Environ Exp Bot 39:131–139. https://doi.org/10.1016/S0098-8472(97)00039-7
Boru G, Vantoai T, Alves J, Hua D, Knee M (2003) Responses of soybean to oxygen deficiency and elevated root-zone carbon dioxide concentration. Ann Bot 91:447–453. https://doi.org/10.1093/aob/mcg040
Botella MA, Martinez V, Pardines J, Cerda A (1997) Salinity induced potassium deficiency in maize plants. J Plant Physiol 150:200–205. https://doi.org/10.1016/S0176-1617(97)80203-9
Britzke D, da Silva LS, Moterle DF, dos Santos Rheinheimer D, Bortoluzzi EC (2012) A study of potassium dynamics and mineralogy in soils from subtropical Brazilian lowlands. J Soil Sed 12:185–197. https://doi.org/10.1007/s11368-011-0431-7
Cakmak I (2005) The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J Plant Nutr Soil Sci 168:521–530. https://doi.org/10.1002/jpln.200420485
Cakmak I, Hengeler C, Marschner H (1994) Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency. J Exp Bot 45:1245–1250. https://doi.org/10.1093/jxb/45.9.1245
Chakraborty K, Bhaduri D, Meena HN, Kalariya K (2016) External potassium (K+) application improves salinity tolerance by promoting Na+-exclusion, K+-accumulation and osmotic adjustment in contrasting peanut cultivars. Plant Physiol Biochem 103:143–153. doi: https://doi.org/10.1016/j.plaphy.2016.02.039
Chapman JM, Muhlemann JK, Gayomba SR, Muday GK (2019) RBOH-dependent ROS synthesis and ROS scavenging by plant specialized metabolites to modulate plant development and stress responses. Chem Res Toxicol 32:370–396
Chartzoulakis K, Psarras G, Vemmos S, Loupassaki M, Bertaki M (2006) Response of two olive cultivars to salt stress and potassium supplement. J Plant Nutr 29:2063–2078. https://doi.org/10.1080/01904160600932682
Chen W, Yang X, He Z, Feng Y, Hu F (2008) Differential changes in photosynthetic capacity, 77 K chlorophyll fluorescence and chloroplast ultrastructure between Zn-efficient and Zn-inefficient rice genotypes (Oryza sativa) under low zinc stress. Physiol Plant 132:89–101
Chen Y, Yu M, Zhu Z, Zhang L, Guo Q (2013) Optimisation of potassium chloride nutrition for proper growth, physiological development and bioactive component production in Prunella vulgaris L. PLoS ONE 8:e66259. doi:https://doi.org/10.1371/journal.pone.0066259
Chen CT, Lee CL, Yeh DM (2018) Effects of nitrogen, phosphorus, potassium, calcium, or magnesium deficiency on growth and photosynthesis of Eustoma. HortScience 53:795–798. https://doi.org/10.21273/HORTSCI12947-18
Chérel I, Lefoulon C, Boeglin M, Sentenac H (2014) Molecular mechanisms involved in plant adaptation to low K(+) availability. J Exp Bot 65:833–848
Corpas FJ, González-Gordo S, Palma JM (2020) Plant peroxisomes: a factory of reactive species. Front Plant Sci 11:853
Coskun D, Britto DT, Kronzucker HJ (2014) The physiology of channel-mediated K + acquisition in roots of higher plants. Physiol Plant 151:305–312
Cui J, Tcherkez G (2021) Potassium dependency of enzymes in plant primary metabolism. Plant Physiol Biochem 166:522–530
Czempinski K, Frachisse JM, Maurel C, Barbier-Brygoo H, Mueller‐Roeber B (2002) Vacuolar membrane localization of the Arabidopsis’ two-pore K+ channel KCO1. Plant J 29:809–820. https://doi.org/10.1046/j.1365-313X.2002.01260.x
Dave A, Agarwal P, Agarwal PK (2022) Mechanism of high affinity potassium transporter (HKT) towards improved crop productivity in saline agricultural lands. 3 Biotech 12:51
Deeken R, Sanders C, Ache P, Hedrich R (2000) Developmental and light-dependent regulation of a phloem-localised K+ channel of Arabidopsis thaliana. Plant J 23:285–290. doi: https://doi.org/10.1046/j.1365-313X.2000.00791.x
Degl’Innocenti E, Hafsi C, Guidi L, Navari-Izzo F (2009) The effect of salinity on photosynthetic activity in potassium-deficient barley species. J Plant Physiol 166:1968–1981. https://doi.org/10.1016/j.jplph.2009.06.013
Demidchik V (2018) ROS-activated ion channels in plants: biophysical characteristics, physiological functions and molecular nature. Int J Mol Sci 19:1263. https://doi.org/10.3390/ijms19041263
Demidchik V, Cuin TA, Svistunenko D, Smith SJ, Miller AJ, Shabala S, Sokolik A, Yurin V (2010) Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: single-channel properties, genetic basis and involvement in stress-induced cell death. J Cell Sci 123:1468–1479. https://doi.org/10.1242/jcs.064352
DeTar RA, Barahimipour R, Manavski N, Schwenkert S, Höhner R, Bölter B, Inaba T, Meurer J, Zoschke R, Kunz H-H (2021) Loss of inner-envelope K+/H + exchangers impairs plastid rRNA maturation and gene expression. Plant Cell 33:2479–2504
Devi BSR, Kim YJ, Selvi SK, Gayathri S, Altanzul K, Parvin S, Yang DU, Lee OR, Lee S, Yang DC (2012) Influence of potassium nitrate on antioxidant level and secondary metabolite genes under cold stress in Panax ginseng. Russ J Plant Physiol 59:318–325. https://doi.org/10.1134/S1021443712030041
Dhiman S, Sharma P, Bhardwaj T, Devi K, Khanna K, Kapoor N, Kaur R, Sharma A, Kaur R, Bhardwaj R (2022) Role of potassium in heavy metal stress. In: Iqbal N, Umar S (eds) Role of potassium in abiotic stress. Springer, Singapore, pp 163–182. https://doi.org/10.1007/978-981-16-4461-0_8
Dreyer I, Blatt MR (2009) What makes a gate? The ins and outs of Kv-like K+ channels in plants. Trends Plant Sci 14:383–390. https://doi.org/10.1016/j.tplants.2009.04.001
Duby G, Hosy E, Fizames C, Alcon C, Costa A, Sentenac H, Thibaud JB (2008) AtKC1, a conditionally targeted Shaker-type subunit, regulates the activity of plant K + channels. Plant J 53:115–123. https://doi.org/10.1111/j.1365-313x.2007.03324.x
Egilla JN, Davies FT, Boutton TW (2005) Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 43:35–140. https://doi.org/10.1007/s11099-005-5140-2
Epstein E, Rains DW, Elzam OE (1963) Resolution of dual mechanisms of potassium absorption by barley roots. Proc Natl Acad Sci USA 49:684–692. https://dx.doi.org/10.1073%2Fpnas.49.5.684
Feng X, Liu W, Cao F, Wang Y, Zhang G, Chen ZH, Wu F (2020) Overexpression of HvAKT1 improves drought tolerance in barley by regulating root ion homeostasis and ROS and NO signaling. J Exp Bot 71:6587–6600
Flexas J, Barbour MM, Brendel O, Cabrera HM, Carriquí M, Díaz-Espejo A, Douthe C, Dreyer E, Ferrio JP, Gago J, Gallé A, Galmesa J, Kodamah N, Medranoa H, Niinemetsi U, Peguero-Pinaa JJ, Poua A, Ribas-Carboa M, Tomasa M, Tosensi T, Warrenj CR (2012) Mesophyll diffusion conductance to CO2: an unappreciated central player in photosynthesis. Plant Sci 193:70–84. https://doi.org/10.1016/j.plantsci.2012.05.009
Foyer CH, Noctor G (2005) Oxidant and antioxidant signalling in plants: a re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ 28:1056–1071. https://doi.org/10.1111/j.1365-3040.2005.01327.x
Gajdanowicz P, Michard E, Sandmann M, Rocha M, Correa LGG, Ramirez- Aguilar SJ, Gomez-Porras JL, Gonzalez W, Thibaud JB, van Dongen JT, Dreyer I (2011) Potassium (K+) gradients serve as a mobile energy source in plant vascular tissues. Proc Natl Acad Sci USA 108:864–869. doi: https://doi.org/10.1073/pnas.1009777108
Gao J, Su Y, Yu M, Huang Y, Wang F, Shen A (2021) Potassium alleviates post-anthesis photosynthetic reductions in winter wheat caused by waterlogging at the stem elongation stage. Front Plant Sci 11:607475. doi: https://doi.org/10.3389/fpls.2020.607475
Garcia-Mata C, Wang JW, Gajdanowicz P, Gonzalez W, Hills A, Donald N, Riedelsberger J, Amtmann A, Dreyer I, Blatt MR (2010) A minimal cysteine motif required to activate the SKOR K+ channel of Arabidopsis by the reactive oxygen species H2O2. J Biol Chem 285:29286–29294. https://doi.org/10.1074/jbc.M110.141176
Gaymard F, Pilot G, Lacombe B, Bouchez D, Bruneau D, Boucherez J, Michaux-Ferriere N, Thibaud JB, Sentenac H (1998) Identification and disruption of a plant shaker-like outward channel involved in K+ release into the xylem sap. Cell 94:647–655. https://doi.org/10.1016/S0092-8674(00)81606-2
Geiger D, Becker D, Vosloh D, Gambale F, Palme K, Rehers M, Anschuetz U, Dreyer I, Kudla J, Hedrich R (2009) Heteromeric AtKC1-AKT1 channels in Arabidopsis roots facilitate growth under K+‐limiting conditions. J Biol Chem 284:21288–21295. https://dx.doi.org/10.1074%2Fjbc.M109.017574
Gerardeaux E, Saur E, Constantin J, Porté A, Jordan-Meille L (2009) Effect of carbon assimilation on dry weight production and partitioning during vegetative growth. Plant Soil 324:329–343. https://doi.org/10.1007/s11104-009-9950-z
Gierth M, Maser P (2007) Potassium transporters in plants—involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett 581:2348–2356. https://doi.org/10.1016/j.febslet.2007.03.035
Gierth M, Maser P, Schroeder JI (2005) The potassium transporter AtHAK5 functions in K+ deprivation-induced high‐affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol 137:1105–1114. https://doi.org/10.1104/pp.104.057216
Gobert A, Isayenkov S, Voelker C, Czempinski K, Maathuis FJ (2007) The two-pore channel TPK1 gene encodes the vacuolar K+ conductance and plays a role in K+ homeostasis. Proc Natl Acad Sci USA 104:10726–10731. https://doi.org/10.1073/pnas.0702595104
Gómez-Ruiz PA, Lindig-Cisneros R, de la Barrera E, Martorell C (2016) Potassium enhances frost tolerance in young individuals of three tropical dry forest species from Mexico. Funct Plant Biol 43:461–467. https://doi.org/10.1071/FP15329
Hafsi C, Debez A, Abdelly C (2014) Potassium deficiency in plants: effects and signaling cascades. Acta Physiol Plant 36:1055–1070. doi: https://doi.org/10.1007/s11738-014-1491-2
Hafsi C, Falleh H, Saada M, Ksouri R, Abdelly C (2017) Potassium deficiency alters growth, photosynthetic performance, secondary metabolites content, and related antioxidant capacity in Sulla carnosa grown under moderate salinity. Plant Physiol Biochem 118:609–617. doi:https://doi.org/10.1016/j.plaphy.2017.08.002
Hamamoto S, Horie T, Hauser F, Deinlein U, Schroeder JI, Uozumi N (2015) HKT transporters mediate salt stress resistance in plants: from structure and function to the field. Curr Opin Biotechnol 32:113–120. https://doi.org/10.1016/j.copbio.2014.11.025
Han Q (2011) Height-related decreases in mesophyll conductance, leaf photosynthesis and compensating adjustments associated with leaf nitrogen concentrations in Pinus densiflora. Tree Physiol 31:976–984. https://doi.org/10.1093/treephys/tpr016
Han M, Wu W, Wu WH, Wang Y (2016) Potassium transporter KUP7 Is involved in K(+) acquisition and translocation in Arabidopsis root under K(+)-limited conditions. Mol Plant 9:437–446. https://doi.org/10.1016/j.molp.2016.01.012
Hartt CE (1969) Effect of potassium deficiency upon translocation of C-14 in attached blades and entire plants of sugarcane. Plant Physiol 44:1461–1469. https://doi.org/10.1104/pp.44.10.1461
Hasanuzzaman M, Bhuyan MHMB, Nahar K, Hossain MS, Mahmud JA, Hossen MS, Masud AAC, Moumita, Fujita M (2018) Potassium: a vital regulator of plant responses and tolerance to abiotic stresses. Agron 8:31. doi:https://doi.org/10.20944/preprints201801.0223.v1
Hedrich R (2012) Ion channels in plants. Physiol Rev 92:1777–1811. doi: https://doi.org/10.1152/physrev.00038.2011
Hernandez M, Fernandez-Garcia N, Garcia-Garma J, Rubio-Asensio JS, Rubio F, Olmos E (2012) Potassium starvation induces oxidative stress in Solanum lycopersicum L. roots. J Plant Physiol 169:1366–1374. https://doi.org/10.1016/j.jplph.2012.05.015
Hinsinger P, Bell MJ, Kovar JL, White PJ (2021). Rhizosphere processes and root traits determining the acquisition of soil potassium. In: Murrell TS et al (ed) Improving potassium recommendations for agricultural crops, Springer nature, Switzerland, pp. 99–117
Honsbein A, Sokolovski S, Grefen C, Campanoni P, Pratelli R, Paneque M, Chen Z, Johansson I, Blatt MR (2009) A tripartite SNARE-K+ channel complex mediates in channel-dependent K+ nutrition in Arabidopsis. Plant Cell 21:2859–2877. https://doi.org/10.1105/tpc.109.066118
Hosy E, Vavasseur A, Mouline K, Dreyer I, Gaymard F, Porée F, Boucherez J, Lebaudy A, Bouchez D, Very AA, Simonneau T, Thibaud JB, Sentenac H (2003) The Arabidopsis outward K+ channel GORK is involved in regulation of stomatal movements and plant transpiration. Proc Natl Acad Sci USA 100:5549–5554. https://doi.org/10.1073/pnas.0733970100
Houmani H, Corpas FJ (2016) Differential responses to salt-induced oxidative stress in three phylogenetically related plant species: Arabidopsis thaliana (glycophyte), Thellungiella salsuginea and Cakile maritima (halophytes). Involvement of ROS and NO in the control of K+/Na + homeostasis. AIMS Biophys 3:380–397
Houmani H, Debez A, Freitas-Silva L, Abdelly C, Palma JM, Corpas FJ (2022) Potassium (K+) starvation-induced oxidative stress triggers a general boost of antioxidant and NADPH-generating systems in the halophyte Cakile maritima. Antioxidants (Basel) 11:401. https://doi.org/10.3390/antiox11020401
Hu W, Lv X, Yang J, Chen B, Zhao W, Meng Y, Wang Y, Zhou Z, Oosterhuis DM (2016) Effects of potassium deficiency on antioxidant metabolism related to leaf senescence in cotton (Gossypium hirsutum L.). Field Crops Res 191:139–149
Hu W, Lu Z, Meng F, Li X, Cong R, Ren T, Sharkey TD, Lu J (2020) The reduction in leaf area precedes that in photosynthesis under potassium deficiency: the importance of leaf anatomy. New Phytol 227:1749–1763. doi:https://doi.org/10.1111/nph.16644
Hu W, Di Q, Wei J, Zhang J, Liu J (2021) Grafting tobacco onto nutrient-efficient rootstocks improves photosynthesis. J Am Soc Hort Sci 146:286–293. https://doi.org/10.21273/JASHS05041-21
Inoue SI, Kaiserli E, Zhao X, Waksman T, Takemiya A, Okumura M, Takahashi H, Seki M, Shinozaki K, Endo Y, Sawasaki T, Kinoshita T, Zhang X, Christie JM, Shimazaki KI (2020) CIPK23 regulates blue light-dependent stomatal opening in Arabidopsis thaliana. Plant J 104:679–692
Jákli B, Tavakol E, Tränkner M, Senbayram M, Dittert K (2017) Quantitative limitations to photosynthesis in K deficient sunflower and their implications on water-use efficiency. J Plant Physiol 209:20–30. https://doi.org/10.1016/j.jplph.2016.11.010
Jegla T, Busey G, Assmann SM (2018) Evolution and structural characteristics of plant voltage-gated K + channels. Plant Cell 30:2898–2909
Jeschke DW, Atkins CA, Pate JS (1985) Ion circulation via phloem and xylem between root and shoot of nodulated white lupin. J Plant Physiol 117:319–330. https://doi.org/10.1016/S0176-1617(85)80068-7
Jia Y, Yang X, Islam E, Feng Y (2008) Effects of potassium deficiency on chloroplast ultrastructure and chlorophyll fluorescence in inefficient and efficient genotypes of rice. J Plant Nutr 31:2105–2118. https://doi.org/10.1080/01904160802459625
Jia Q, Zheng C, Sun S, Amjad H, Liang K, Lin W (2018) The role of plant cation/proton antiporter gene family in salt tolerance. Biol Plant 62:617–629
Jin SN, Huang JQ, Li XQ, Zheng BS, Wu JS, Wang ZJ, Liu GH, Chen M (2011) Effects of potassium supply on limitations of photosynthesis by mesophyll diffusion conductance in Carya cathayensis. Tree Physiol 31:1142–1151. https://doi.org/10.1093/treephys/tpr095
Johnson R, Vishwakarma K, Hossen MS, Kumar V, Shackira AM, Puthur JT, Abdi G, Sarraf M, Hasanuzzaman M (2022) Potassium in plants: growth regulation, signaling, and environmental stress tolerance. Plant Physiol Biochem 172:56–69
Jung JY, Shin R, Schachtman DP (2009) Ethylene mediates response and tolerance to potassium deprivation in Arabidopsis. Plant Cell 21:607–621. https://doi.org/10.1105/tpc.108.063099
Kalaji HM, Da browski P, Cetner MD, Samborska IA, Łukasik I, Brestic M, Zivcak M, Tomasz H, Mojski J, Kociel H, Panchal BM (2017) A comparison between different chlorophyll content meters under nutrient deficiency conditions. J Plant Nutr 40:1024–1034. https://doi.org/10.1080/01904167.2016.1263323
Kanai S, Moghaieb RE, El-Shemy HA, Panigrahi R, Mohapatra PK, Ito J, Nguyen NT, Saneoka H, Fujita K (2011) Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in greenhouse tomato prior to its effects on source activity. Plant Sci 180:368–374. https://doi.org/10.1016/j.plantsci.2010.10.011
Kim et MJ, Ciani S, Schachtman DP (2010) A peroxidase contributes to ROS production during Arabidopsis root response to potassium deficiency. Mol Plant 3(2):420–427. https://doi.org/10.1093/mp/ssp121
Kohli SK, Khanna K, Bhardwaj R, Abd Allah EF, Ahmad P, Corpas FJ (2019) Assessment of subcellular ROS and NO metabolism in higher plants: multifunctional signaling molecules. Antioxidants (Basel) 8:641
Kumar AR, Kumar N, Kavino M (2006) Role of potassium in fruit crops—a review. Agric Rev 27:284–291
Kumari S, Chhillar H, Chopra P, Khanna RR, Khan MIR (2021) Potassium: a track to develop salinity tolerant plants. Plant Physiol Biochem 167:1011–1023
Kusaka M, Kalaji HM, Mastalerczuk G, Dąbrowski P, Kowalczyk K (2021) Potassium deficiency impact on the photosynthetic apparatus efficiency of radish. Photosynthetica 59:127–136. https://doi.org/10.32615/ps.2020.077
Lacombe B, Pilot G, Michard E, Gaymard F, Sentenac H, Thibaud JB (2000b) A Shaker-like K(+) channel with weak rectification is expressed in both source and sink phloem tissues of Arabidopsis. Plant Cell 12:837–851. https://doi.org/10.1105/tpc.12.6.837
Lana LG, de Araújo LM, Silva TF, Modolo LV (2021) Interplay between gasotransmitters and potassium is a K+ ey factor during plant response to abiotic stress. Plant Physiol Biochem 169:322–332
Lawanson AO, Otusanya OO, Akomolede DA (1977) Mechanism of potassium deficiency-induced retardation of chlorophyll biosynthesis in Zea mays. Experientica 33:1145–1146. https://doi.org/10.1007/BF01922293
Lefoulon C (2021) The bare necessities of plant K+ channel regulation. Plant Physiol 2187:2092–2109
Leterrier M, Barroso JB, Valderrama R, Begara-Morales JC, Sánchez-Calvo B, Chaki M, Luque F, Viñegla B, Palma JM, Corpas FJ (2016) Peroxisomal NADP-isocitrate dehydrogenase is required for Arabidopsis stomatal movement. Protoplasma 253:403–415
Lhamo D, Wang C, Gao Q, Luan S (2021) Recent advances in genome-wide analyses of plant potassium transporter families. Curr Genomics 22:164–180
Li W, Xu G, Alli A, Yu L (2018) Plant HAK/KUP/KT K+ transporters: function and regulation. Semin Cell Dev Biol 74:133–141. https://doi.org/10.1016/j.semcdb.2017.07.009
Li H, Shi J, Wang Z, Zhang W, Yang H (2020) H2S pretreatment mitigates the alkaline salt stress on Malus hupehensis roots by regulating Na+/K + homeostasis and oxidative stress. Plant Physiol Biochem 156:233–241
Liu K, Li L, Luan S (2006) Intracellular K+ sensing of SKOR, a shaker-type K+ channel from Arabidopsis. Plant J 46:260–268. https://doi.org/10.1111/j.1365-313X.2006.02689.x
Liu CH, Chao YY, Kao CH (2013) Effect of potassium deficiency on antioxidant status and cadmium toxicity in rice seedlings. Bot Stud 54:2. https://doi.org/10.1186/1999-3110-54-2
Liu C, Wang X, Tu B, Li Y, Chen H, Zhang Q, Liu X (2021) Root K affinity drivers and photosynthetic characteristics in response to low potassium stress in K high-efficiency vegetable soybean. Front Plant Sci. https://doi.org/10.21203/rs.3.rs-349341/v1
Lombardi L, Sebastiani L (2005) Copper toxicity in Prunus cerasifera: growth and antioxidant enzymes responses of in vitro grown plants. Plant Sci 168:797–802. https://doi.org/10.1016/j.plantsci.2004.10.012
Lu Z, Lu J, Pan Y, Li X, Cong R, Ren T (2016a) Genotypic variation in photosynthetic limitation responses to K deficiency of Brassica napus is associated with potassium utilisation efficiency. Funct Plant Biol 43:880–891. https://doi.org/10.1071/FP16098
Lu Z, Lu J, Pan Y, Lu P, Li X, Cong R, Ren T (2016b) Anatomical variation of mesophyll conductance under potassium deficiency has a vital role in determining leaf photosynthesis. Plant Cell Environ 39:2428–2439. https://doi.org/10.1111/pce.12795
Lu Z, Ren T, Pan Y, Li X, Cong R, Lu J (2016c) Differences on photosynthetic limitations between leaf margins and leaf centers under potassium deficiency for Brassica napus L. Sci Rep 6:21725. https://doi.org/10.1038/srep21725
Lu Z, Hu W, Ren T, Zhu C, Li X, Cong R, Guo S, Lu J (2018) Impact of K deficiency on leaves and siliques photosynthesis via metabolomics in Brassica napus. Environ Exp Bot. https://doi.org/10.1016/j.envexpbot.2018.11.008
Lu Z, Xie K, Pan Y, Ren T, Lu J, Wang M, Shen Q, Guo S (2019) Potassium mediates coordination of leaf photosynthesis and hydraulic conductance by modifications of leaf anatomy. Plant Cell Environ 42:2231–2244. https://doi.org/10.1111/pce.13553
Lysenko EA, Klaus AA, Pshybytko NL, Kusnetsov VV (2015) Cadmium accumulation in chloroplasts and its impact on chloroplastic processes in barley and maize. Phytosynth Res 125:291–303. https://doi.org/10.1007/s11120-014-0047-z
Maach M, Baghour M, Akodad M, Gálvez FJ, Sánchez ME, Aranda MN, Venema K, Rodríguez-Rosales MP (2020) Overexpression of LeNHX4 improved yield, fruit quality and salt tolerance in tomato plants (Solanum lycopersicum L.). Mol Biol Rep 47:4145–4153
Maathuis FJM (2009) Physiological functions of mineral macronutrients. Curr Opin Plant Biol 12:250–258. https://doi.org/10.1016/j.pbi.2009.04.003
Marschner P (2012) Marschner’s mineral nutrition of higher plants, third edn. Academic Press, London, UK
Mengel K, Kirkby EA (1982) Potassium. In: Mengel K, Kirkby EA (eds) Principles of plant nutrition. International Potash Institute, Worblaufeu-Bern, pp 335–368
Mengel K, Kirkby EA (1987) Principles of plant nutrition. International Potash Institute, Bern
Mengel K, Kirkby EA (2001) Principles of plant nutrition, fifth. Kluwer Acad. Publishers, Dordrecht, The Netherlanad
Mishra V, Singh P, Tripathi DK, Corpas FJ, Singh VP (2021) Nitric oxide and hydrogen sulfide: an indispensable combination for plant functioning. Trends Plant Sci 26:1270–1285
Mittler R (2002) Oxidative stress antioxidants and stress tolerance. Trends Plant Sci 7:405–410. https://doi.org/10.1016/S1360-1385(02)02312-9
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9:490–498. https://doi.org/10.1016/j.tplants.2004.08.009
Mouhamad R, Alsaede A, Iqbal M (2016) Behavior of potassium in soil: a mini review. Chem Int 2:58–69
Mukherjee S, Corpas FJ (2020) Crosstalk among hydrogen sulfide (H2S), nitric oxide (NO) and carbon monoxide (CO) in root-system development and its rhizosphere interactions: a gaseous interactome. Plant Physiol Biochem 155:800–814
Naciri R, Lahrir M, Benadis C, Chtouki M, Oukarroum A (2021) Interactive effect of potassium and cadmium on growth, root morphology and chlorophyll a fluorescence in tomato plant. Sci Rep 11:5384. https://doi.org/10.1038/s41598-021-84990-4
Nieves-Cordones M, Aleman F, Martinez V, Rubio F (2014) K+ uptake in plant roots. The systems involved their regulation and parallels in other organisms. J Plant Physiol 171:688–695. https://doi.org/10.1016/j.jplph.2013.09.021
Niinemets Ü, Diaz-Espijo A, Flexas J, Galmes J, Warren CR (2009a) Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field. J Exp Bot 60:2249–2270. https://doi.org/10.1093/jxb/erp036
Noctor G, Mhamdi A, Foyer CH (2014) The roles of reactive oxygen metabolism in drought: not so cut and dried. Plant Physiol 164:1636–1648. https://doi.org/10.1104/pp.113.233478
Oliferuk S, Simontacchi M, Rubio F, Santa-María GE (2020) Exposure to a natural nitric oxide donor negatively affects the potential influx of rubidium in potassium-starved Arabidopsis plants. Plant Physiol Biochem 150:204–208
Perelman A, Imas P, Bansal SK (2022) Potassium role in plants response to abiotic stresses. In: Iqbal N, Umar S (eds) Role of potassium in abiotic stress. Springer, Singapore, pp 15–39. https://doi.org/10.1007/978-981-16-4461-0_2
Perveen S, Shahbaz M, Ashraf M (2010) Regulation in gas exchange and quantum yield of photosystem II (PSII) in salt-stressed and non-stressed wheat plants raised from seed treated with triacontanol. Pak J Bot 42:3073–3081
Perveen S, Shahbaz M, Ashraf M (2011) Modulation in activities of antioxidant enzymes in salt-stressed and non-stressed wheat (Triticum aestivum L.) plants raised from seed treated with triacontanol. Pak J Bot 43:2463–2468
Perveen S, Shahbaz M, Ashraf M (2012) Is pre-sowing seed treatment with triacontanol effective in improving some physiological and biochemical attributes of wheat (Triticum aestivum L.) under salt stress? J Appl Bot Food Qual 85:41–48
Pettigrew WT (2008) Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol Plant 133:670–681. https://doi.org/10.1111/j.1399-3054.2008.01073.x
Pier PA, Berkowitz GA (1987) Modulation of water-stress effects on photosynthesis by altered leaf K+. Plant Physiol 85:655–661. https://doi.org/10.1104/pp.85.3.655
Pilot G, Lacombe B, Gaymard F, Che´rel I, Boucherez J, Thibaud JB, Sentenac H (2001) Guard cell inward K+ channel activity in Arabidopsis involves expression of the twin channel subunits KAT1 and KAT2. J Biol Chem 276:3215–3221. https://doi.org/10.1074/jbc.M007303200
Pyo YJ, Gierth M, Schroeder JI, Cho MH (2010) High-affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and postgermination growth under low-potassium conditions. Plant Physiol 153:863–875. https://doi.org/10.1104/pp.110.154369
Qi DU, Zhao XH, Le XIA, Jiang CJ, Wang XG, Yi HAN, Jing WANG, Yu HQ (2019) Effects of potassium deficiency on photosynthesis, chloroplast ultrastructure, ROS, and antioxidant activities in maize (Zea mays L.). J Integr Agric 18:395–406. https://doi.org/10.1016/S2095-3119(18)61953-7
Raddatz N, Morales de Los Ríos L, Lindahl M, Quintero FJ, Pardo JM (2020) Coordinated transport of nitrate, potassium, and sodium. Front Plant Sci 11:247
Rady MM, Mohamed GF (2018) Improving salt tolerance in Triticum aestivum (L.) plants irrigated with saline water by exogenously applied proline or potassium. Adv Plants Agric Res 8:193–199. https://doi.org/10.15406/apar.2018.08.00312
Ragel P, Raddatz N, Leidi EO, Quintero FJ, Pardo JM (2019) Regulation of K+ nutrition in plants. Front Plant Sci 10:281
Rahman MA, Chikushi J, Yoshida S, Yahata H, Yasunsga E (2005) Effect of high air temperature on grain growth and yields of wheat genotypes differing in heat tolerance. J Agric Meteorol 60:605–608. https://doi.org/10.2480/agrmet.605
Rahman A, Rahman MM, Hasan MM, Begum F, Sarker MAZ (2014) Effects of foliar application of potassium orthophosphate on grain yield and kernel quality of wheat (Triticum aestivum) under terminal heat stress. Bangladesh J Agric Res 39:67–77. https://doi.org/10.3329/bjar.v39i1.20144
Reintanz B, Szyroki A, Ivashikina N, Ache P, Godde M, Becker D, Palme K, Hedrich R (2002) AtKC1, a silent Arabidopsis potassium channel alpha-subunit modulates root hair K + influx. Proc Natl Acad Sci USA 99:4079–4084. https://doi.org/10.1073/pnas.052677799
Rogiers SY, Greer DH, Moroni FJ, Baby T (2020) Potassium and magnesium mediate the light and CO2 photosynthetic responses of grapevines. Biology 9:144. https://doi.org/10.3390/biology9070144
Römheld V, Kirkby EA (2010) Research on potassium in agriculture: needs and prospects. Plant Soil 335:155–180. https://doi.org/10.1007/s11104-010-0520-1
Sagi M, Fluhr R (2006) Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol 41:336–340
Sánchez-McSweeney A, González-Gordo S, Aranda-Sicilia MN, Rodríguez-Rosales MP, Venema K, Palma JM, Corpas FJ (2021) Loss of function of the chloroplast membrane K+/H + antiporters AtKEA1 and AtKEA2 alters the ROS and NO metabolism but promotes drought stress resilience. Plant Physiol Biochem 160:106–119
Santa-Maria GE, Rubio F, Dubcovsky J, Rodriguez-Navarro A (1997) The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. Plant Cell 9:2281–2289. https://doi.org/10.1105/tpc.9.12.2281
Santa-María GE, Oliferuk S, Moriconi JI (2018) KT-HAK-KUP transporters in major terrestrial photosynthetic organisms: a twenty years tale. J Plant Physiol 226:77–90
Sardans J, Peñuelas J (2015) Potassium: a neglected nutrient in global change. Glob Ecol Biogeogr 24:261–275. https://doi.org/10.1111/geb.12259
Sardans J, Peñuelas J (2021) Potassium control of plant functions: ecological and agricultural implications. Plants (Basel) 10:419
Sarwar M, Saleem MF, Ullah N, Ali S, Rizwan M, Shahid MR, Alyemeni MN, Alamri SA, Ahmad P (2019) Role of mineral nutrition in alleviation of heat stress in cotton plants grown in glasshouse and field conditions. Sci Rep 9:13022. https://doi.org/10.1038/s41598-019-49404-6
Sasidharan R, Hartman S, Liu Z, Martopawiro S, Sajeev N, Veen HV, Yeung E, Voesenek LACJ (2018) Signal dynamics and interactions during flooding stress. Plant Physiol 176:1106–1117. https://doi.org/10.1104/pp.17.01232
Schachtman DP, Schroeder JI, Lucas WJ, Anderson JA, Gaber RF (1992) Expression of an inward rectifying potassium channel by the Arabidopsis KAT1 cDNA. Science 258:1654–1658. https://doi.org/10.1126/science.8966547
Sehar S, Adil MF, Zeeshan M, Holford P, Cao F, Wu F, Wang Y (2021) Mechanistic insights into potassium-conferred drought stress tolerance in cultivated and tibetan wild barley: differential osmoregulation, nutrient retention, secondary metabolism and antioxidative defense capacity. Int J Mol Sci 22:13100
Severtson D, Callow N, Flower K, Neuhaus A, Olejnik M, Nansen C (2016) Unmanned aerial vehicle canopy reflectance data detects potassium deficiency and green peach aphid susceptibility in canola. Precision Agric 17:659–677. https://doi.org/10.1007/s11119-016-9442-0
Shabala S (2003) Regulation of potassium transport in leaves: from molecular to tissue level. Ann Bot 92:627–634. https://doi.org/10.1093/aob/mcg191
Shahbaz M, Ashraf M, Athar HR (2008) Does exogenous application of 24-epibrassinolide ameliorate salt-induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul 55:51–64. https://doi.org/10.1007/s10725-008-9262-y
Shamsi IH, Jiang L, Wei K, Jilani G, Hua S, Zhang GP (2010) Alleviation of cadmium toxicity in soybean by potassium supplementation. J Plant Nutr 33:1926–1938. https://doi.org/10.1080/01904167.2010.512052
Shao RX, Xin LF, Zheng HF, Li LL, Ran WL, Mao J, Yang QH (2016) Changes in chloroplast ultrastructure in leaves of drought-stressed maize inbred lines. Photosynthetica 54:74–80. https://doi.org/10.1007/s11099-015-0158-6
Shen C, Wang J, Jin X, Liu N, Fan X, Dong C, Qirong S, Xu Y (2017) Potassium enhances the sugar assimilation in leaves and fruit by regulating the expression of key genes involved in sugar metabolism of Asian pears. Plant Growth Regul 83:287–300. https://doi.org/10.1007/s10725-017-0294-z
Shen C, Huang R, Tang Y, Wang Z (2018) Potassium nutrition recover impacts on stomatal, mesophyll and biochemical limitations to photosynthesis in Carya cathayensis and Hickory illinoensis. BioRxiv. https://doi.org/10.1101/425629
Shin R, Schachtman DP (2004) Hydrogen peroxide mediates plant root cell response to nutrient deprivation. Proc Natl Acad Sci USA 101:8827–8832. https://doi.org/10.1073/pnas.0401707101
Siddiqui MH, Al-Whaibi MH, Sakran AM, Basalah MO, Ali HM (2012) Effect of calcium and potassium on antioxidant system of Vicia faba L. under cadmium stress. Int J Mol Sci 13:6604–6619. https://doi.org/10.3390/ijms13066604
Siddiqui MH, Khan MN, Mukherjee S, Alamri S, Basahi RA, Al-Amri AA, Alsubaie QD, Al-Munqedhi BMA, Ali HM, Almohisen IAA (2021) Hydrogen sulfide (H2S) and potassium (K+) synergistically induce drought stress tolerance through regulation of H+-ATPase activity, sugar metabolism, and antioxidative defense in tomato seedlings. Plant Cell Rep 40:1543–1564
Singh SK, Reddy VR (2017) Potassium starvation limits soybean growth more than the photosynthetic processes across CO2 levels. Front Plant Sci 8:991. https://doi.org/10.3389/fpls.2017.00991
Singh SK, Reddy VR (2018) Co-regulation of photosynthetic processes under potassium deficiency across CO2 levels in soybean: mechanisms of limitations and adaptations. Photosynth Res 137:183–200. https://doi.org/10.1007/s11120-018-0490-3
Sitko K, Gieroń Ż, Szopiński M, Zieleźnik-Rusinowska P, Rusinowski S, Pogrzeba M, Daszkowska-Golec A, Kalaji HM, Małkowski E (2019) Influence of short-term macronutrient deprivation in maize on photosynthetic characteristics, transpiration and pigment content. Sci Rep 9:14181. https://doi.org/10.1038/s41598-019-50579-1
Song ZZ, Duan CL, Guo SL, Yang Y, Feng YF, Ma RJ, Yu ML (2015) Potassium contributes to zinc stress tolerance in peach (Prunus persica) seedlings by enhancing photosynthesis and the antioxidant defense system. Genet Mol Res 14:8338–8351. https://doi.org/10.4238/2015.July.27.22
South PF, Cavanagh AP, Liu HW, Ort DR (2019) Synthetic glycolate metabolism pathways stimulate crop growth and productivity in the field. Science 363:eaat9077. https://doi.org/10.1126/science.aat9077
Sparks DL (1987) Potassium dynamics in soil. In: Stewart BA (ed) Advances in soil science, vol 6. Springer, New York, pp 1–63
Sparks DL (2000) Bioavailability of soil potassium, D-38-D-52. In: Sumner ME (ed) Handbook of soil science. CRC Press, Boca Raton
Sparks DL, Huang PM (1985) Physical chemistry of soil potassium. In: Munson RD (ed) Potassium in agriculture. ASA, CSSA and SSSA, Madison, pp 201–276. https://doi.org/10.2134/1985.potassium.c9
Sparks DL, Martens DC, Zelazny LW (1980) Plant uptake and leaching of applied and indigenous potassium in Dothan soils. Agron J 72:551–555. https://doi.org/10.2134/agronj1980.00021962007200030034x
Srivastava AK, Shankar A, Nalini Chandran AK, Sharma M, Jung KH, Suprasanna P, Pandey GK (2020) Emerging concepts of potassium homeostasis in plants. J Exp Bot 71:608–619
Sun Z, Chen Y, Schaefer V, Liang H, Li W, Huang S, Peng C (2015) Responses of the hybrid between Sphagneticola trilobata and Sphagneticola calendulacea to low temperature and weak light characteristic in South China. Sci Rep 5:16906. https://doi.org/10.1038/srep16906
Taiz L, Zeiger E, Møller IM, Murphy A (2015) Plant physiology and development, 6th edn. Sinauer associates inc, publishers, Sunderland
Tang ZH, Zhang AJ, Wei M, Chen XG, Liu ZH, Li HM, Ding YF (2015) Physiological response to potassium deficiency in three sweet potato (Ipomoea batatas [L.] Lam.) genotypes differing in potassium utilization efficiency. Acta Physiol Plant. https://doi.org/10.1007/s11738-015-1901-0
Terry N, Ulrich A (1973) Effects of potassium deficiency on the photosynthesis and respiration of leaves of sugar beet. Plant Physiol 51:783–786. https://doi.org/10.1104/pp.51.4.783
Thiel G, Wolf AH (1997) Operation of K+ channels in stomatal movement. Trends Plant Sci 2:339–345. https://doi.org/10.1016/S1360-1385(97)84621-3
Thornburg TE, Liu J, Li Q, Xue H, Wang G, Li L, Fontana JE, Davis KE, Liu W, Zhang B, Zhang Z, Liu M, Pan X (2020) Potassium deficiency significantly affected plant growth and development as well as microRNA mediated mechanism in wheat (Triticum aestivum L.). Front Plant Sci 11:1219. https://doi.org/10.3389/fpls.2020.01219
Tian XL, Wang GW, Zhu R, Yang PZ, Duan LS, Li ZH (2008) Conditions and indicators for screening cotton (Gossypium hirsutum L.) varieties tolerant to low potassium. Acta Agron Sinica 34:1435–1443. https://doi.org/10.1016/S1875-2780(08)60050-4
Tränkner M, Tavakol E, Jákli B (2018) Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol Plant 163:414–431. https://doi.org/10.1111/ppl.12747
Tripathy BC, Oelmüller R (2012) Reactive oxygen species generation and signaling in plants. Plant Signal Behav 7:1621–1633. https://doi.org/10.4161/psb.22455
Trono D, Laus MN, Soccio M, Alfarano M, Pastore D (2015) Modulation of potassium channel activity in the balance of ROS and ATP production by durum wheat mitochondria-an amazing defense tool against hyperosmotic stress. Front Plant Sci 6:1072
Tsujii M, Kera K, Hamamoto S, Kuromori T, Shikanai T, Uozumi N (2019) Evidence for potassium transport activity of Arabidopsis KEA1-KEA6. Sci Rep 9:10040
Venema K, Quintero FJ, Pardo JM, Donaire JP (2002) The Arabidopsis Na+/H+ exchanger AtNHX1 catalyzes low affinity Na+ and K+ transport in reconstituted liposomes. J Biol Chem 277:2413–2418. https://doi.org/10.1074/jbc.M105043200
Ve’ry AA, Nieves-Cordones M, Daly M, Khan I, Fizames C, Sentenac H (2014) Molecular biology of K+ transport across the plant cell membrane: what do we learn from comparison between plant species? J Plant Physiol 171:748–769. https://doi.org/10.1016/j.jplph.2014.01.011
Von Caemmerer S, Quick WP, Furbank RT (2012) The development of C4 rice: current progress and future challenges. Science 336:1671–1672. https://doi.org/10.1126/science.1220177
Walker DJ, Leigh RA, Miller AJ (1996) Potassium homeostasis in vacuolate plant cells. Proc Natl Acad Sci USA 93:10510–10514. https://doi.org/10.1073/pnas.93.19.10510
Wang Y, Wu WH (2013) Potassium transport and signaling in higher plants. Annu Rev Plant Biol 64:451–476. https://doi.org/10.1146/annurev-arplant-050312-120153
Wang Y, Wu WH (2017) Regulation of potassium transport and signaling in plants. Curr Opin Plant Biol 39:123–128. https://doi.org/10.1016/j.pbi.2017.06.006
Wang HY, Zhou JM, Du CW, Chen XQ (2010a) Potassium fractions in soils as affected by monocalcium phosphate, ammonium sulfate and potassium chloride application. Pedosphere 20:368–377. https://doi.org/10.1016/s1002-0160(10)60026-4
Wang Y, He L, Li HD, Xu J, Wu WH (2010b) Potassium channel alpha-subunit AtKC1 negatively regulates AKT1‐mediated K+ uptake in Arabidopsis roots under low K+ stress. Cell Res 20:826–837. https://doi.org/10.1038/cr.2010.74
Wang N, Hua H, Eneji AE, Li Z, Duan L, Tian X (2012) Genotypic variations in photosynthetic and physiological adjustment to potassium deficiency in cotton (Gossypium hirsutum). J Photochem Photobiol B 110:1–8. https://doi.org/10.1016/j.jphotobiol.2012.02.002
Wang M, Zheng Q, Shen Q, Guo S (2013) The critical role of potassium in plant stress response. Int J Mol Sci 14:7370–7390. https://doi.org/10.3390/ijms14047370
Wang XG, Zhao XH, Jiang CJ, Li CH, Cong S, Wu D, Chen YQ, Yu HQ, Wang CY (2015) Effects of potassium deficiency on photosynthesis and photoprotection mechanisms in soybean (Glycine max (L.) Merr.). J Integr Agric 14:856–863. https://doi.org/10.1016/S2095-3119(14)60848-0
Wang XP, Chen LM, Liu WX, Shen LK, Wang FL, Zhou Y, Zhang ZD, Wu WH, Wang Y (2016) AtKC1 and CIPK23 synergistically modulate AKT1-mediated low-potassium stress responses in Arabidopsis. Plant Physiol 170:2264–2277. https://doi.org/10.1104/pp.15.01493
Wang J, Zhu G, Dong Y, Zhang H, Rengel Z, Ai Y, Zhang Y (2018) Potassium starvation affects biomass partitioning and sink–source responses in three sweet potato genotypes with contrasting potassium-use efficiency. Crop Pasture Sci 69:506–514. https://doi.org/10.1071/CP17328
Wang Y, Tang RJ, Yang X, Zheng X, Shao Q, Tang QL, Fu A, Luan S (2019) Golgi-localized cation/proton exchangers regulate ionic homeostasis and skotomorphogenesis in Arabidopsis. Plant Cell Environ 42:673–687
Wang Y, Wu WH (2015) Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency. Curr Opin Plant Biol 25:46–52
Waqas M, Yaning C, Iqbal H, Shareef M, Rehman HU, Bilal HM (2021) Synergistic consequences of salinity and potassium deficiency in quinoa: linking with stomatal patterning, ionic relations and oxidative metabolism. Plant Physiol Biochem 159:17–27
Waraich EA, Ahmad R, Halim A, Aziz T (2012) Alleviation of temperature stress by nutrient management in crop plants: a review. J Soil Sci Plant Nutr 12:221–244. https://doi.org/10.4067/S0718-95162012000200003
Wedepohl KH (1995) The composition of the continental crust. Geochim Cosmochim Acta 59:1217–1232. https://doi.org/10.1016/0016-7037(95)00038-2
Weng XY, Zheng CJ, Xu HX, Sun JY (2007) Characteristics of photosynthesis and functions of the water-water cycle in rice (Oryza sativa) leaves in response to potassium deficiency. Physiol Plant 131:614–621. https://doi.org/10.1111/j.1399-3054.2007.00978.x
Whitehead D, Barbour MM, Griffin KL, Turnbull MH, Tissue DT (2011) Effects of leaf age and tree size on stomatal and mesophyll limitations to photosynthesis in mountain beech (Nothofagus solandrii var. cliffortiodes). Tree Physiol 31:985–996. https://doi.org/10.1093/treephys/tpr021
Worthington V (2001) Nutritional quality of organic versus conventional fruits, vegetables, and grains. J Altern Complement Med 7:161–173. https://doi.org/10.1089/107555301750164244
Xia J, Kong D, Xue S, Tian W, Li N, Bao F, Hu Y, Du J, Wang Y, Pan X, Wang L, Zhang X, Niu G, Feng X, Li L, He Y (2014) Nitric oxide negatively regulates AKT1-mediated potassium uptake through modulating vitamin B6 homeostasis in Arabidopsis. Proc Natl Acad Sci USA 111:16196–16201
Xu YW, Zou YT, Husaini AM, Zeng JW, Guan LL, Liu Q, Wu W (2011) Optimization of potassium for proper growth and physiological response of Houttuynia cordata Thunb. Environ Exp Bot 71:292–297. https://doi.org/10.1016/j.envexpbot.2010.12.015
Xu HJ, Johkan M, Maruo T, Kagawa N, Tsukagoshi S (2021) New insight on low-K lettuce: from photosynthesis to primary and secondary metabolites. HortScience 56:407–413. https://doi.org/10.21273/HORTSCI15130-20
Yang XE, Liu JX, Wang WM, Ye ZQ, Luo AC (2004) Potassium internal use efficiency relative to growth vigor, potassium distribution, and carbohydrate allocation in rice genotypes. J Plant Nutr 27:837–852. https://doi.org/10.1081/PLN-120030674
Yasin NA, Zaheer MM, Khan WU, Ahmad SR, Ahmad A, Ali A, Akram W (2018) The beneficial role of potassium in Cd-induced stress alleviation and growth improvement in Gladiolus grandiflora L. Int J Phytoremediation 20:274–283. DOI: https://doi.org/10.1080/15226514.2017.1374337
Zaheer MM, Yasin NA, Ahmad SR, Khan WU, Ahmad A, Ali A, Rehman SU (2018) Amelioration of cadmium stress in gladiolus (Gladiolus grandiflora L.) by application of potassium and silicon. J Plant Nutr 41:461–476. https://doi.org/10.1080/01904167.2017.1385808
Zhang G, Johkan M, Hohjo M, Tsukagosh S, Maruo T (2017a) Plant growth and photosynthesis response to low potassium conditions in three lettuce (Lactuca sativa) types. Hortic J 86:229–237. https://doi.org/10.2503/hortj.OKD-008
Zhang W, Zhang NS, Zhao JJ, Guo YP, Zhao ZY, Mei LX (2017b) Potassium fertilization improves apple fruit (Malus domestica Borkh. Cv. Fuji) development by regulating trehalose metabolism. J Hortic Sci Biotechnol 92:539–549. https://doi.org/10.1080/14620316.2017.1304165
Zhang L, Li G, Wang M, Di D, Sun L, Kronzucker HJ, Shi W (2018) Excess iron stress reduces root tip zone growth through nitric oxide-mediated repression of potassium homeostasis in Arabidopsis. New Phytol 219:259–274
Zhao DL, Oosterhuis DM, Bednarz CW (2001) Influences of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultrastructure of cotton plants. Photosynthetica 39:103–199. https://doi.org/10.1023/A:1012404204910
Zhao X, Du Q, Zhao Y, Wang H, Li Y, Wang X, Yu H (2016) Effects of different potassium stress on leaf photosynthesis and chlorophyll fluorescence in maize (Zea mays L.) at seedling stage. Agricul Sci 7:44–753
Zhao N, Zhu H, Zhang H, Sun J, Zhou J, Deng C, Zhang Y, Zhao R, Zhou X, Lu C, Lin S, Chen S (2018) Hydrogen sulfide mediates K + and Na + homeostasis in the roots of salt-resistant and salt-sensitive poplar species subjected to NaCl stress. Front Plant Sci 9:1366
Zhu XG, Portis-Jr AR, Long SP (2004) Would transformation of C3 crop plants with foreign Rubisco increase productivity? A computational analysis extrapolating from kinetic properties to canopy photosynthesis. Plant Cell Environ 27:155–165. https://doi.org/10.1046/j.1365-3040.2004.01142.x
Zhu GF, Li X, Su YH, Lu L, Huang CL (2011) Seasonal fluctuations and temperature dependence in photosynthetic parameters and stomatal conductance at the leaf scale of Populus euphratica Oliv. Tree Physiol 31:178–195. https://doi.org/10.1093/treephys/tpr005
Zou J, Lu JW, Li YS, Li XK (2011) Regional evaluation of winter rapeseed response to K fertilization, K use efficiency, and critical level of soil K in the Yangtze River Valley. Sci Agric Sin 10:911–920. https://doi.org/10.1016/S1671-2927(11)60076-X
Funding
Havza Imtiaz gratefully acknowledges the financial assistance rendered by the University Grant Commission, New Delhi, India, in the form of Non-net fellowship. FJC research is supported by a European Regional Development Fund-cofinanced grant from the Ministry of Economy and Competitiveness (PID2019-103924GB-I00) and the Plan Andaluz de Investigación, Desarrollo e Innovación (PAIDI 2020) (P18-FR-1359), Spain.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Additional information
Communicated by Luca Sebastiani.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Imtiaz, H., Mir, A.R., Corpas, F.J. et al. Impact of potassium starvation on the uptake, transportation, photosynthesis, and abiotic stress tolerance. Plant Growth Regul 99, 429–448 (2023). https://doi.org/10.1007/s10725-022-00925-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-022-00925-7