
Abstract
The objective of this work was to evaluate the genetic variability of Cenchrus ciliaris accessions using morpho-agronomic descriptors for the identification of germplasm with phenotypic divergence that can be used as male parents in breeding programmes for the species. Thirty-eight accessions were characterised using six quantitative and 12 qualitative descriptors. The descriptors that most contributed to the divergence were inflorescence length (22.48%), leaf blade width (19.52%) and leaf blade length (18.35%). Based on the mean grouping test, for the character basal tillering intensity, the accessions were distributed into three groups, of which accessions CPATSA79125 and CPATSA79150 had the most tillers. Thus, there was genetic diversity among the evaluated accessions, with an emphasis on the CPATSA79123 and CPATSA90611 genotypes, as they were the most divergent accessions and thus can be used in clonal selections and in the prospection of genes of agronomic interest. The different accessions can be selected for the most varied objectives of the improvement programmes for the species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Brazil’s vast and diverse landscape encompasses a wide range of climatic zones, which greatly influence the distribution of livestock and the importance of forage production. In general, we can divide Brazil into semi-arid zones (Northeast and West Central regions) and humid zones (Amazon, South and Southeast regions) and discuss their significance for livestock and forage production. While humid regions are characterised by a humid tropical or temperate climate with abundant rainfall and well-defined seasons, semi-arid zones of Brazil are characterised by a steppe climate with prolonged dry seasons and irregular and low rainfall. The predominant vegetation includes Caatinga, which consist of drought-resistant shrubs and trees. Livestock, primarily goats and cattle, is raised in this region, and forage production is crucial for their sustenance during dry periods (Sá and Silva 2015).
Research with native and exotic forages for the Brazilian semiarid region is still limited; however, in this region, a promising seed market has been formed mainly by cultivars developed for other regions with different edaphoclimatic conditions (Antônio 2015). Thus, there is a need for initiatives to advance the development of forage cultivars that are more adapted to semi-arid regions. Allied to this, the growth of herds of goats, sheep and cattle in the region is factors that determine the increase in the consumption of forage seeds. According to IBGE (2021), Brazil’s goat population stands at 11,923,630 heads, with the state of Pernambuco housing 3,204,448 of them. This figure surpasses all other states in the Northeast region, except for the state of Bahia, which boasts a slightly larger goat herd. Similarly, in terms of cattle, Pernambuco has 2,173,313 heads, which is the highest figure among the states in the Northeast region. In this way, exploring the potential of species, whether native or exotic forage destined for animal feed in the semiarid region, using this information for breeding programmes for these crops is very pertinent. In this sense, the Active Germplasm Bank (BAG) ex situ of Embrapa Semiarid of the genus Cenchrus with about 120 accessions exists to keep the genetic variability of the species preserved. These characteristics are positive, especially in regions with a semi-arid climate, since the herds in this region have food available in periods of drought.
Buffelgrass is known for its exceptional drought tolerance, making it well adapted to areas with irregular and low rainfall. It can be used to produce hay and silage due to its high nutritional value, persistence, versatile grazing options and adaptability to a range of soil types; however, there are few commercial genotypes (Voltolini et al. 2019). Therefore, the addition of new accessions is necessary to expand the options for future crossings and, consequently, to obtain new cultivars with desirable characteristics for semi-arid conditions. This need for expansion is due to numerous reasons. For example, in the case of feeding goats that are medium-sized animals, accessions with a more prostrate growth habit facilitate access to food, making it a positive feature to select.
The genetic diversity offered by the BAG is essential for the activities of breeding programmes, enabling the selection of more divergent genitors destined for crosses if this potential is identified (Bonato et al. 2006). In a breeding programme, obligate apomictic plants can only be used as male parents in crosses. The recognition of a good material for incorporation in breeding programmes is determined by its characterisation or its description, allowing the observation of different patterns and thus the correct identification of each plant material (Nevada 2011). Therefore, we sought to estimate the genetic variability among buffelgrass accessions to identify promising genotypes for incorporation and exploitation in genetic improvement programmes for the species.
Materials and methods
For the evaluated accessions, assessments were carried out at the recommended time for each descriptor/characteristic, using quantitative and qualitative morpho-agronomic descriptors. The experiment was conducted at the Caatinga Experimental Station, which is affiliated with Embrapa Semiarid, situated in Petrolina-PE. The experimental site is located at coordinates S09° 24′ 38″ and W40° 29′ 56″. The climate is classified as semi-arid with an altitude of 377 m. The soil in the area is classified as red-yellow dystrophic argisol with a sandy loam texture.
Thirty-eight accessions of the genus Cenchrus belonging to the Active Germplasm Bank of Embrapa Semiarid were used (Table 1). The studied accessions were selected because they were the most promising in terms of size for use within the Cenchrus genetic improvement program. Seeds of these accessions were sown in trays containing commercial substrates composed of vermiculite. The seedlings were transplanted to the field at a size of about 10 cm. The experiment was carried out in a randomised block design with three replications, where six plants per plot were evaluated.
For the characterisation of accessions, the descriptors and procedures described by MAPA (2019) were used. Twelve qualitative descriptors and six quantitative descriptors were used, as follows: Qualitative descriptors: type of growth—TG (1-erect, 2-semi-erect and 3-prostrate), rhizome shape—RS (1-linear, 2-globose), leaf blade edge—LBE (1-smooth, 2-serrated), leaf blade colour—LBC (1-light green, 2-green, 3-blue green), leaf blade hairiness—LBH (1-absent or 2-present), intensity of the leaf blade hairiness—ILBH (0-no hair, 3-low, 5-medium and 7-high), sheath pilosity—SP (1-absent or 2-present), inflorescence base pilosity—IBP (1-absent or 2-present), inflorescence colouring—IC (1-cream, 2-brown, 3-purple, 4-black, 5-purple with cream, 6-black with cream), spikelet shell shape—SSS (1-bristles together, 2-free bristles), spikelet grouping—SG (1-isolated or 2-grouped) and spikelet anther colour—SAC (1-white, 2-beige, 3-pink, 4-purple, 5-black, 6-yellow, 7-dark yellow, 8-purple); Quantitative descriptors: basal tillering intensity—BTI, stem diameter—SD, stem internode length—SIL, leaf blade length—LBL, leaf blade width—LBW and inflorescence length—IL.
Qualitative data were presented only in a table of accessions, with the patterns found within the accession considering the large number of classes present within these variables. Data on quantitative characteristics were subjected to an analysis of variance using the following model:
where \({y}_{ij}\) = value observed in the experimental unit that received treatment i, block j; \(m\) = general effect of the mean; \({b}_{j}\) = effect of block j; \({t}_{i}\)= effect of treatment i; \({e}_{ij}\) = random error (residue).
To estimate the genetic divergence among accessions using data from quantitative morpho-agronomic descriptors, a matrix of genetic distances among accessions was obtained using the generalised distance of Mahalanobis (1936).
The statistical analysis model used for calculating the generalised Mahalanobis distance with quantitative data was as follows:

where \({D}_{ii{\prime}}^{2}\) = Mahalanobis distance between genotypes i and i′; \(\Psi^{ - 1}\) = the matrix of variance and residual covariances; \(\delta {\prime}\)= [d1, d2.., dv], where dj = yij − yi′j;  = yi′j: average of the i-nth genotype in relation to the j-nth variable.
= yi′j: average of the i-nth genotype in relation to the j-nth variable.
After obtaining the matrix, the accessions were grouped using mean clustering between the groups (UPGMA) to create the dendrogram. The cutline in the dendrogram for the formation of groups was established accord to the method of Mojena (1977), and the consistency of the groups formed by the hierarchical method was verified using the cophenetic correlation coefficient (CCC). In addition, the relative contribution of the descriptors to divergence was determined using the computational software GENES (Version 2021.1.9) (Cruz 2016). Finally, the average number of accessions for each characteristic was grouped using the Scott and Knott (1974) grouping method.
Results and discussion
There was a significant difference among the accessions for all evaluated traits, which indicates a high level of genotype variation (Table 2). This represents the availability of base materials for exploitation regarding the composition of improved materials. Bruno et al. (2017) evaluated 25 Cenchrus accessions with the aim of characterising and identifying genetic diversity and verifying a wide genetic variation among individuals. These results showed that there was variability within the Cenchrus BAG of Embrapa Semi-Arid to be explored, which may enable the development of new cultivars, an extremely important factor since there are few cultivars of buffelgrass on the market.
Based on the mean grouping test (Table 3), for the characteristic BTI, the accessions were distributed into three groups. This characteristic defines the ability of the plant to regrow in arid and semi-arid regions, being one of the main characteristics of forage grasses, as it ensures that this plant will regrow after grazing or cutting (Mansoor et al. 2002). The regrowth capacity ensures the perennialism of forage grasses, guaranteeing their survival by enabling the development of a new leaf growth (Costa et al. 2004).
With respect to stem diameter (SD), the accessions were grouped into five groups (a, b, c, d, e). The accessions with the smallest SD were CPATSA 79129, CPATSA 79141 and CPATSA 90563 (Table 3). Accessions with smaller diameters are more palatable, while accessions with larger stem diameters have a greater capacity for regrowth (Costa et al. 2004).
Some accessions in the present study presented excellent results in the BTI since the raw material destined to feed the animals is the aerial part, allowing greater food production. In this context, accession CPATSA79125 can be incorporated into breeding for the species, seeking to transfer alleles that confer this pattern to other genotypes. If the accession is apomictic, this gene transfer can be performed via biotechnological methods, such as genetic transformation or gene editing.
Accession CPATSA79125 stands out for its greater leaf blade width (LBW) and inflorescence length (IL), which indicates that this material may present high production of the aerial part that can be used to feed the herds. In addition, this accession is superior to the commercial cultivar Biloela (CPATSA7602), which indicates a certain potential for exploitation within the breeding programmes for the species in the composition of new materials or even in the transfer of these alleles to other genotypes in the final stage of breeding.
Arshad et al. (2007) reported a significant and high correlation between the number of tillers per plant and fresh weight, although the latter trait was not measured in our evaluation. There is evidence that, for this trait, accessions stand out from the rest. The number of tillers per plant is important for ensuring the persistence of buffelgrass pastures, as it shows the capacity for regrowth.
The SIL was another characteristic separated into four groups (a, b, c, d). For the characteristic LBC, four groups were formed (a, b, c, d). LLW and IL were the characteristics with the greatest distribution of accessions, indicative of the dissimilarity between them (a, b, c, d, e, f).
When seeking to obtain a hybrid combination with a greater BTI and LBL, recombination between accessions CPATSA79125 and CPATSA78122 would possibly result in a promising combination in obtaining high productivity materials with greater leaf blade width, with characteristics combined in a potentially vigorous hybrid for biomass production.
IL was the trait that most contributed to estimating genetic diversity (22.48%) (Table 4). When observing the mean clustering test (Table 3), accessions CPATSA 79125, CPATSA79130, CPATSA90558 and CPATSA90570 stood out with lengths ranging from 14.6 to 12.25 cm, which indicates the existence of greater variation in the inflorescence, a behaviour that can be easily explored as it is a characteristic that is easy to see during its evaluation.
In addition, IL, LBW and LBL also stood out with values of 22.48%, 19.52% and 18.35%, respectively (Table 4). These characteristics are related to the aerial part of the grass, one of the main structures, in addition to the stem diameter used for animal feed. In addition to stem diameter, in the present study, the characteristics that most contributed to genetic diversity among accessions were those related to biomass production. In addition, they are characteristics that can be evaluated right away in the initial development of the plant, different from the characteristics related to the inflorescence, for example, which cannot be evaluated until anthesis. Another approach that can be used to facilitate the evaluation of these characteristics is the conversion of quantitative variables into qualitative ones (creating classes from the intervals of quantitative values), seeking to optimise time and effort until reaching the improved genotype.
Based on the qualitative descriptors (Table 5), for the type of growth, 5 accessions were identified as erect, 24 as semi-erect, and 5 as prostrate. The rhizome shape in accessions presented the globose form, and only one had a linear form, which was the commercial cultivar Biloela (CPATSA7602). The rhizomes are a monopolar stem branching system, rich in reserves, as described by Appezzato da Glória (2003). Squires and Myres (1970) highlighted the importance of rhizomes in Paspalum paniculatum L. (Poaceae), as they enable greater tolerance to grazing and drought.
All leaves of the evaluated accessions had a smooth leaf blade edge. For leaf blade colouring, 20 accessions were green, and 18 accessions were light green. Hairiness at the base of the inflorescence was absent for 22 accessions and present for 16 accessions. For inflorescence colouration, 25 were cream-coloured, 8 were purple-and-cream, 2 were black, 1 was black-and-cream, 1 was purple and 1 was brown.
For the spikelet shape, 32 accessions had free bristles, and 6 accessions had united bristles. There was no variation in the grouping of spikelets, as all were isolated. Spikelet anther colour was as follows: 29 were yellow, 6 were beige, 1 was dark yellow (CPATSA7602), 1 was pink (CPATSA79121), and 1 was purple (CPATSA90611).
IC is a characteristic that is easy to identify and differentiate from other genotypes, as it varies among accessions, as well as the colour of the anthers, which also vary in colour. These qualitative descriptors confirm the dissimilarity among the genotypes, which are of great importance, especially when used together with other traits in the choice of genitors intended for crosses. Several authors have already attested to the efficiency of characterisation by means of markers or morphological descriptors, such as in elephant grass (Silva et al. 2009) and Stylosanthes spp. (Karia et al. 2002). These works were useful in identifying characteristics with great variation in genotypes.
The genetic dissimilarity between the genotypes was estimated according to the Mahalanobis distance (1936), where the distance values among the accessions ranged from 1.98 to 301.11, with emphasis on the accessions CPATSA79147 and CPATSA7754 and CPATSA79123 and CPATSA90611 for being the least and most dissimilar genotypes, respectively (Table 6). This indicates that the recombination between the first two accessions will result in hybrids and segregating populations with a narrow genetic base and, consequently, not very interesting for exploration within the breeding programmes for the species, unlike the second pair of genotypes, which can result in hybrid combinations that complement each other by having different characteristics.
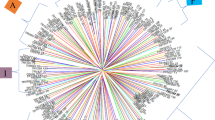
The dendrogram of the graphic representation of the diversity among the accessions, with the cut in the grouping following the method of Mojena (1977), shows the formation of three groups: one composed by accession CPATSA78103, another by accessions CPATSA18621 and CPATSA90611, and the last formed by the other accessions divided into subgroups (Fig. 1). As observed in the dissimilarity measures, the first group presented the most contrasting genotype among the evaluated materials, presenting itself as an excellent genitor for recombination, since this accession “CPATSA78103” has an inflorescence colouration different from the others, black, which identifies it easily for this descriptor. The accessions that also formed a group were CPATSA18621 and CPATSA90611, where they presented pink and purple colouring, respectively, for the descriptor anther colour (spikelet), thus making them easy to identify from these accessions. These results reinforce the existence of genetic divergence among genotypes.

Dendrogram representative of the genetic diversity among accessions of buffelgrass, using mean linkage clustering between groups (UPGMA), with a cophenetic correlation coefficient of 72%
When working with C. ciliaris, Bruno et al. (2017), Marshall et al. (2012), Arshad et al. (2007), Jorge et al. (2008), and Mansoor et al. (2002) found genetic divergence among the genotypes they evaluated, which corroborates the results of the present study. Thus, they reinforced the importance of characterising Cenchrus germplasm. Based on the dendrogram, the CCC presented a value greater than 72%, indicating that the grouping of accessions presents low distortion between the matrix and the dendrogram.
Conclusion
There was genetic diversity among the evaluated accessions.
Characteristics related to biomass production, such as length and width of the leaf blade and basal tillering intensity, were the most informative in estimating the genetic divergence between accessions.
Accessions CPATSA79123 and CPATSA90611 were the most divergent accessions and thus could be potential male parents in the breeding programme.
Depending on the purpose/objective of the breeding programme, the other accessions could also be considered for improving the adaptivity and productivity of the species.
References
Antonio RP (2015) Espécies forrageiras: principais contribuições, estado atual e perspectivas para a pesquisa na Embrapa Semi árido. Documentos, 269 - Embrapa Semi-árido AOAC (2002) Métodos oficiais de análise, 17 edition
Appezzato da Glória B (2003) Morfologia de sistemas subterrâneos. In: Pinto AS (ed). Ribeirão Preto, Brazil
Arshad M, Ashraf M, Ahamad M, Zaman F (2007) Morphogenetic variability potential of Cenchrus ciliaris L., from Cholistan desert, Pakistan. Pak J Bot 39(5):1481–1488
Bonato ALV, Calvo ES, Geraldi IO, Arias CAA (2006) Genetic similarity among soybean (Glycine max (L) Merrill) cultivars released in Brazil using AFLP markers. Genet Mol Biol 29(4):692–704
Bruno LRGP, Antonio RP, Assis JGA, Moreira JN, Lira ICSA (2017) Buffel grass morphoagronomic characterization from Cenchrus germplasm active bank. Rev Caatinga 30(2):487–495
Costa NL, Magalhães JA, Townsend CR, Paulino VT (2004) Fisiologia e manejo de plantas forrageiras. Rondônia, Documentos 85, Embrapa Rondônia https://www.infoteca.cnptia.embrapa.br/bitstream/doc/916005/1/doc85plantasforrageiras.pdf
Cruz CD (2016) Genes software–extended and integrated with the R, Matlab and Selegen. Acta Sci-Agron 38(4):547–552
IBGE Censo agropecuário (2021) resultados preliminares. Rio de Janeiro, 2021. Disponível em: https://www.ibge.gov.br/explica/producao-agropecuaria/. Accessed 12 Sep 2023
Jorge MAB, Wouw MVD, Hanson J, Mohammed J (2008) Characterization of a collection of buffel grass (Cenchrus ciliaris). Trop Grassl 42(1):27–39
Karia CT, Andrade RP, Charchar MJDA, Gomes AC (2002) Caracterização morfológica de acessos do gênero Stylosanthes no banco ativo de germoplasma da Embrapa Cerrados: Coleção 1994/1995. Embrapa Cerrados, Planaltina, DF
Mahalanobis PC (1936) On the generalized distance in statistics. Proc Natl Inst Sci India 2(1):9–55
Mansoor U, Hameed M, Wahid A, Rao AR (2002) Ecotypic variability for drought resistance in Cenchrus ciliaris L. germplasm from Cholistan desert in Pakistan. Int J Agric Biol 4(3):392–397
MAPA Ministério da Agricultura e Pecuária (2019) – Instrução para execução de ensaios de distinguibilidade, homogeneidade e estabilidade de capim buffel (Cenchrus ciliaris; Cenchrus pennisetiformis; Cenchrus setigerus emseus híbridos. Brasília 2019. Disponível em: https://www.gov.br/agricultura/pt-br/assuntos/insumosagropecuarios/insumos-agricolas/protecao-de-cultivar/forrageiras Accessed 29 Sep 2023
Marshall VM, Lewis MM, Ostendorf B (2012) Buffel grass (Cenchrus ciliaris) as an invader and threat to biodiversity in arid environments: a review. J Arid Environ 78:1–12. https://doi.org/10.1016/j.jaridenv.2011.11.005
Mojena R (1977) Hierarchical grouping methods and stopping rules: an evaluation. Comput J 20:359–363
Nevada AF, Vázquez Badillo MG, Borrego Escalante F, Sánchez Aspeytia D (2011) Análisis de la homogeneidad, distribución y estabilidad de tres variedades sobresalientes de tomate. Rev Mex Cienc Agr 2(1):5–16
Sá IB, Silva PCG (2015) Brazilian semi-arid region: research, development, and innovation. Embrapa, Brasília. https://ainfo.cnptia.embrapa.br/digital/bitstream/item/165677/1/E-book-Brazilian-Semi-arid-region-research-development-and-innovation.epub
Scott AJ, Knott MA (1974) Cluster analysis method for grouping means in the analysis of variance. Biometrics 30(2):507–512
Silva SHB, Santos MVF, Lira MA, Dubeux Junior JCB, Freitas EV, Ferreira RLC (2009) Uso de descritores morfológicos e herdabilidade de caracteres em clones de capim-elefante de porte baixo. Rev Bras Zootec 38(8):1451–1459. https://doi.org/10.1590/S1516-35982009000800008
Squires UR, Myres LF (1970) Performance of warm season perennial grasses for irrigated pastures at Deniliquin, southeastern Australia. Trop Grassl 4(1):153–161
Voltolini TV, Santana SRA, Antunes GR, Araújo GGL (2019) Alternativas alimentares para os rebanhos. In: Melo RF, Voltolini TV (eds) Agricultura familiar dependente de chuva no Semiárido. Embrapa, Petrolina, pp 229–261
Acknowledgements
The authors would like to thank the National Council for Scientific and Technological Development (CNPq), the Coordination for the Improvement of Higher Education Personnel (CAPES), the Pernambuco Science and Technology Support Foundation (FACEPE), and Embrapa Tropical Semi-Arid for their support, scholarship and grants received through project numbers: PQ-306505/2022-3, BFP-0087.5.01/22, and CAPES 001.
Funding
The authors would like to thank the National Council for Scientific and Technological Development (CNPq) through project number PQ-306505/2022-3 and Embrapa Tropical Semi-Arid (Project 22.16.04.018.00.04.010) for the financial support. We also acknowledge the Coordination for the Improvement of Higher Education Personnel (CAPES) for the doctoral scholarship Finance Code 001.
Author information
Authors and Affiliations
Contributions
ICNSR, RPA and NFM conceived and planned the study and wrote the manuscript. RPA provided the germplasm accessions, which were multiplied by ICNSR for the experiment. ICNSR and RPA carried out the phenotyping. and ICNSR, NFM and TLN analysed the data. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have not disclosed any competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
dos Santos Ribeiro, I.C.N., do Nascimento, T.L., Antônio, R.P. et al. Estimation of genetic diversity in Cenchrus ciliaris accessions introduced in the Brazilian semi-arid region. Genet Resour Crop Evol 71, 1883–1892 (2024). https://doi.org/10.1007/s10722-023-01746-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-023-01746-x