
Abstract
Strychnos spinosa Lam. is an important wild edible fruit tree (WEFT) that is increasingly threatened due to anthropogenic pressure. Despite its remarkable socio-economic potential, commercial plantations for the species are rare. Characterization of the genetic diversity and potential of WEFT is a prerequisite for domestication and genetic improvement. This study assessed the morphological diversity and differentiation among populations of S. spinosa across a climatic gradient (Sudano-Guinean vs Sudanian zones) in Benin, West Africa. Morphological data were collected on 81 trees and 810 fruits of S. spinosa from 7 populations in the two climatic zones using nine phenotypic descriptors. Descriptive statistics and multivariate analyses were used to describe and partition differences among trees and study populations. Results showed highly significant differences (P ≤ 0.001) among populations for all measured traits. The within-population variation accounted for the highest proportion (53–90%) of the total variation. Strong and positive correlations (r = 0.91–0.99; P < 0.05) were observed among trunk and fruit traits except for tree height and fruits’ seed weight suggesting that fruit traits (fruit mass, pulp mass, seed number, ratio) can be predicted from trunk traits (dbh). Cluster analysis distinguished three distinct groups of S. spinosa in Benin. Population from Bassila phytodistrict in the Sudano-Guinean zone showed superior phenotypic traits (e.g. tree diameter, fruit mass, and pulp mass) indicating a high potential for selection for domestication purposes. Our study revealed marked phenotypic diversity of S. spinosa in Benin and provides relevant information for domestication and harnessing of S. spinosa genetic resources.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Given the lack of effective control on the adverse effects of changing climate, adaptation strategies are increasingly searched for and developed to improve the resilience of local communities and contribute to food security and nutrition. In such a context, the traditional communities which are often vulnerable, and with low adaptation strategies, deserve more attention (UNFCCC 2018). Several efforts are being made to reduce the high dependence of their agricultural practices upon climate conditions. To this extent, introduction of important agroforestry species into traditional cropping systems is increasingly recommended as evidence exists on their contribution to locals’ subsistence and income generation (Schippmann et al. 2002). These initiatives are justified by the fact that with increasing food shortages caused by global climate change, it is expected that rural people will depend on wild edible fruit trees (WEFT) to meet their food and nutritional needs (Nebel et al. 2006; Bruschi et al. 2014; Berihun and Molla 2017).
However, the success of WEFT introduction in traditional agroforestry systems requires knowledge on the morphological variations within their populations and identification of desirable traits for genetic improvement. This is an important step towards domestication because, eventually, the morphological variations can reveal the best ideotypes suitable for different environments. So far, several studies have been carried out to reveal the morphological variation within WEFT across Africa (e.g. Leakey et al. 2000, 2005a; Abasse et al. 2011; Assogbadjo et al. 2011; Hounkpèvi et al. 2016; Padonou et al. 2017). Most of these studies have documented a continuous variation between and within studied populations for the investigated traits and have highlighted the potential to derive improved ideotypes from wild populations for the domestication purposes (Gouwakinnou et al. 2011). Although, domestication does not necessarily assure conservation and sustainable use of biodiversity, it has been shown that at least, while there is a substantial wild resource, domestication can increase intraspecific diversity (Leakey et al. 2004). In addition to providing important knowledge to sustainably manage useful plant species, studies of morphological variability can add insights into the adaptation capacity of the species in facing effects of unpredictable hazards like climate change (Hounkpèvi et al. 2016).
The monkey orange, Strychnos spinosa Lam. (Loganiaceae) is an indigenous WEFT to tropical and subtropical Africa, and widely spread from Senegal to South Africa (Sitrit et al. 2003; Isa et al. 2014). It naturally occurs in the savannah areas in tropical Africa and dwell in open woodlands including riverine fringes (Orwa et al. 2009). So far, researches undertaken on the species have underscored its importance for the local population welfare. It is among the topmost important edible tree species in the wild (Madzimure et al. 2013) which serves as an important food resource to poor farmers during periods of food shortage (Thiombiano et al. 2013; Bruschi et al. 2014). It is an important source of medicine for a wide range of diseases ( Arbonnier 2002; Isa et al. 2014; Plantz Africa 2017; World Agroforestry Centre 2018). It is also used to increase milk and crop production (Hoet et al. 2006, 2007; Bero et al. 2011; Salifou et al. 2017). In Benin, many parts of the species such as fruits, leaves, roots, seeds, etc. are used to fulfill these different functions (Avakoudjo et al. 2020). From the literature, knowledge on tree and fruit phenotypic diversity and differentiation across its distribution range in sub-Saharan Africa are rarely documented. This prevents from actions towards sustainable management of the species and exploitation of its potential in reaching food security with the local communities (Avakoudjo 2020).
Consistently with the ecological gradient hypothesis, climatic conditions have been shown to significantly determine natural variation of phenotypic traits at a relatively larger scale (climatic zones) in many WEFT (Fandohan et al. 2010; Hounkpèvi et al. 2016; Okello et al. 2018) and worth to consider in understanding differentiation in phenotypic variation. In this study, nine quantitative morphological descriptors were used to investigate the morphological variability and differentiation in S. spinosa populations in its natural range in two contrasting climatic zones where it is found in Benin: the drier Sudanian zone and the relatively more humid Sudano-Guinean zone. The ultimate goal was to identify elite individuals which can serve as basis for the successful domestication. Specifically, the study aimed to describe the relationship between dendrometric parameters and fruits descriptors and determine the extent of variation of the traits between and within populations of the two climatic zones. In particular, we tested the following hypotheses:
-
1.
Individuals from the Sudanian zone have lower values for studied morphological traits compared to the ones in the Sudano-Guinean zone. We expected so, because drought triggers a wide variety of plant responses ranging from cellular metabolism to changes in growth rates and crop yields which often get lower in drier conditions (Shakeel et al. 2011);
-
2.
As natural selection acts directly on phenotypes and important genetic variation within a population usually enables more phenotypic variation (Lumen Boundless Biology 2020), a higher morphological variation is expected for S. spinosa within populations rather than between populations. Indeed, the variation within a population allows some individuals to adapt to the changing environment by increasing their ability to survive and reproduce, which then ensures the survival of the allele in the population (Lumen Boundless Biology 2020). Furthermore, this expectation is also based on the reports of previous studies on WEFT in Benin along the same gradient (Fandohan et al. 2011; Gouwakinnou et al. 2011; Hounkpèvi et al. 2016).
Materials and methods
Study area
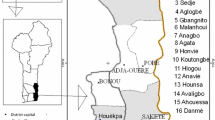
The study was carried out in the Sudanian and the Sudano-Guinean climatic zones which represent the distribution area of S. spinosa in Benin (Akoegninou et al. 2006; Fig. 1). The Sudanian zone (7° 30′–9° 45′ N) is characterised by a truly tropical climate with a unimodal rainfall pattern. The rainy season lasts for 7 months on average and covers the period from April to October with the maximum around August/September. The mean annual rainfall in is often less than 1000 mm. The relative humidity varies from 18 to 99% and temperature ranges from 24 to 31 °C. In the transition zone, which is the Sudano-Guinean zone (7° 30′–9° 45′ N), the climate is sub-humid with a tendency towards a unimodal rainfall regime. The annual rainfall varies between 900 and 1110 mm, whereas temperature and relative humidity ranges between 25 and 29 °C and 31 and 98%, respectively (Adomou et al. 2006; Assogbadjo et al. 2012; Gnanglè et al. 2012). Each of these climatic zones are further divided into phytodistricts which are meso-scale subdivision based on differences in natural vegetation, soil, and to some extent climatic conditions. So, the Sudano-Guinean zone is divided into three phytodistricts namely Zou, South-Borgou, and Bassila. The Sudanian zone also encompasses three phytodistricts including North-Borgou, Mékrou-Pendjari and Atacora chain (Adomou et al. 2006).

Map of Benin showing the location of the studied populations of S. spinosa across phytodistricts
Sampling
Strychnos spinosa has already been subject to cultivation in some areas like Israel (Sitrit et al. 2003) and Australia. Though, no evidence on its cultivation exists for Benin, we focused only on natural populations in protected areas in order to depict natural variation of traits. This was done to limit bias on the studied traits assuming that individuals in the unprotected areas are more prone to human actions. Thus, individuals of the same phytodistrict were considered as a population (a sub-population level was considered when the population individuals’ come from different protected areas in the phytodistrict) and populations of the same climatic zone have been considered as provenance. Therefore, populations of Mékrou-Pendjari (Pendjari national park) and North-Borgou (W national park and Trois-rivières forest) were considered in the Sudanian zone (Supplementary Fig. 1) while populations of Bassila (Penessoulou forest), South-Borgou (Wari-Maro and Toui forests) and Zou (Agoua forest) were considered in the Sudano-Guinean zone (Supplementary Fig. 1), making a total of five populations at least 75 km apart. Tree inventory was done using linear transects of 3 km oriented North–South and East–West from a chosen random starting point based on the protected area map. Identification of trees of S. spinosa was made 50 m apart from both side of the transect and all individuals of the species encountered in the transect were considered and georeferenced. The first individual encountered in the transect was considered as the center of a 1000 m2 plot (50 m × 20 m) and from this first plot, other plots were installed at each 100 m till the end of the transect.
Data collection
At least 30 adult individuals (from the different populations) of the species were randomly selected in each climatic zone (provenance). A distance of at least 100 m was left between fruiting trees in order to avoid genetically close individuals. A total of 81 fruiting individuals of S. spinosa were assessed (30 and 51 respectively in the Sudano-Guinean and Sudanian zone). For each selected tree, the trunk diameter at breast height (Dbh at 1.3 m) and total height (htree, m) were recorded. Then a sample of 10 fruits was collected from each selected tree for further measurement following the protocol described by Leakey et al. (2000) and used by Fandohan et al. (2010) and Hounkpèvi et al. (2016). Seven morphological descriptors were measured on each fruit (Circ: fruit circumference; Thick: fruit thickness; DryM: fruits’ dry mass; SeedsN: seeds number; SeedsW: seeds weight; PulpM: pulp mass; and the Ratio: PulpM/DryM). These parameters were measured with an electronic digital calliper (0.01 mm resolution) and fruit weights were recorded using a 0.01 g-precision electronic balance. To improve accuracy, fruit’s circumference and thickness were measured at the first, the second and the third quarter of each fruit and the arithmetic means considered as the fruit’s circumference and thickness. Similar descriptors were used for other WEFTs (see Leakey et al. 2000, 2005a, b; El-Siddig et al. 2006; Fandohan et al. 2010).
Data analysis
Mean values and coefficient of variation were computed for dendrometric parameters per provenance and per population. The same statistics were calculated for each morphological parameter of fruits and seeds per population and provenance. The skewness was also calculated for each morphological trait to determine the symmetry of their distribution (Idohou et al. 2016).
For each trait, phenotypic classes were defined (Supplementary Table 1) following FAO (2018), Mengistu et al. (2015), and Mille and Louppe (2015). These classes were used to estimate the Shannon–Weaver diversity index (H′) which was used to characterize the phenotypic frequencies of the different traits. This index was computed using the following equation:
where n represent the number of phenotypic classes for a given trait and Pi the proportion of the total number of entries in the ith class. H was estimated for each trait, and per phytodistrict, and climatic zone. H was divided by Hmax = ln(n) in order to standardize H. The standardized phenotypic diversity index (H′) is in the range of 0–1. The diversity index was classified as high (H′ ≥ 0.60), intermediate (0.40 ≤ H′ ≤ 0.60), or low (0.10 ≤ H′ ≤ 0.40), as used by Mengistu et al. (2015).
One-way analysis of variance (ANOVA) followed by Student Newman Keul (SNK) test was performed to compare morphological traits among populations and between provenances. Linear mixed effect models were used to determine the relative importance of different sources of variability namely tree, phytodistrict, and climatic zone for each fruit trait. In this model, tree was considered nested in phytodistrict, and phytodistrict was considered nested in climatic zone. Canonical Discriminant Analysis (CDA) was performed to identify the most discriminating morphological traits and describe each population. This analysis also sought to highlight differences among populations with regards to the discriminant morphological characteristics. Hierarchical clustering analysis was performed to group trees in classes with homogenous characteristics based on both tree and fruit traits. The reliability of the cluster analysis was checked and validated through linear discriminant analysis. Morphological classes were then described using a Principal Component Analysis (PCA) and the results were graphically visualized in a biplot that displayed the groups and the association between individuals and morphological traits. Phylogenetic constellation plot was generated from the hierarchical clustering analysis to depict the genetic relationships among individual trees. The cluster analysis and phylogenetic constellation plots were performed in JMP pro 14.10 (JMP 2019). All remaining statistical analyses were performed using R 3.5.3 (R Core Team 2019).
Results
Morphological variability among S. spinosa individuals
Dendrometric traits of S. spinosa individuals varied significantly between climatic zones (provenances) and populations (phytodistricts) with high amplitude (CV > 25%) for total height and diameter, except for the diameter of North-Borgou population. However, the highest variation was observed in the Sudano-Guinean zone for both diameter and height. The diameter was more variable in South-Borgou and Zou populations whereas the highest variation in tree height was noticed in Zou and Mekrou-Pendjari populations (Table 1).
The coefficient of skewness varied between 0.2 and 1.72 for provenances (Table 1), indicating a right skewed distribution. The trend was almost the same at the population level except for Bassila where the coefficients were negative for the diameter and the height suggesting a left skewed distribution. The largest individuals were encountered in the Sudano-Guinean zone in Bassila population while the tallest ones were found in the Sudanian zone in the Mekrou-Pendjari population.
Morphological variability of S. spinosa fruits
Measured parameters on fruits were significantly higher in the Sudano-Guinean zone than in Sudanian zone (Table 2; P < 0.001). Considering populations, Bassila had the highest value for fruit thickness, dry mass for fruits, seeds number and pulp mass; South-Borgou and Zou populations had the highest value for fruit circumference and seeds weight respectively (Table 2; P < 0.001).
The fruits descriptors were dispersed around mean values in all provenances and populations except for fruit circumference (Sudanian provenance and South-Borgou population) and fruit thickness (Sudanian provenance) which were relatively less dispersed (CV < 15%, Table 2). In most cases, the descriptors showed the highest amplitude of variation in the Sudano-Guinean provenance (Circ; Thick; DryM; SeedsW and PulpM), Zou population (Circ; DryM and PulpM) and Mekrou-Pendjari population (SeedsN; SeedsW and Ratio: PulpM /DryM).
Distribution of the descriptors was largely skewed (moderately or highly) across the provenances except for the circumference in both provenances, and thickness in the Sudanian provenance which were approximately symmetric (skewness = 0.19). The Skewness coefficient was close to zero (approximately normal distribution) for: circumference in all the populations except Zou; thickness in North-Borgou and Mekrou-Pendjari; dry mass, seed number and seed weight in the populations of Bassila, North-Borgou and South-Borgou. There was also a high variation in fruit characteristics within and between populations (Table 3). The within populations variation accounted for the greatest part (53–90%) of the total variation.
Phenotypic diversity among S. spinosa individuals between populations and provenances
For all traits, there was a large natural variation as illustrated in Table 4. The phenotypic diversity estimation (H′) for individual traits ranged from 0.38 for the ratio (pulp mass over dry mass of the fruits) to 0.96 for the pulp mass. The overall mean values of the diversity for all the traits pool together was 0.81. All the traits showed high levels of polymorphism (H′ > 0.60) except the ratio which has the lowest level of diversity (H′ = 0.38).
On the provenance basis, high diversity indices (H′ ≥ 0.60) pooled over descriptors were obtained for individuals from Sudano-Guinean zone (Bassila, H′ = 0.78; Zou, H′ = 0.75; and South-Borgou, H′ = 0.62). The individuals of the Sudanian zone showed intermediate diversity, 0.51 and 0.57 respectively for North-Borgou and Mekrou-Pendjari. The within population H′ depended on the indices of the descriptors. When H’ was considered for each trait, a monomorphism (H′ = 0) was found for ratio in South-Borgou and for diameter in North-Borgou. However, low divergence was recorded for ratio in Bassila (H′ = 0.09), for seed weight in North-Borgou (H′ = 0.35) and in Mekrou-Pendjari (H′ = 0.39), for fruits dry mass in North-Borgou (H′ = 0.2) and in Mekrou-Pendjari (H′ = 0.15).
Relationships between discriminant morphological descriptors
Results of the CDA showed that 93.82% of the information was represented on the first two canonical axes explaining respectively 69.2% and 24.62% of the total variability (Fig. 2; Table S2). Overall, the two axes were highly significant (P < 0.001) in discriminating the populations. The first axis was negatively correlated with the plant’s diameter, fruit circumference, dry mass, seeds number, seeds weight and pulp mass (correlation > 0.5). The second axis was positively correlated with plants height and diameter. All fruit traits were negatively correlated with plant height and positively correlated with plant diameter on the first axis.

Canonical discriminant analysis diagram of the 81 selected individuals showing association of the populations and the morphological traits of S. spinosa
Overall, the population of Bassila had the highest values for all morphological characteristics except for the plant’s height (Fig. 2). The populations of South-Borgou and Zou showed the highest value for seed weight but medium values for the other parameters (Fig. 2). The populations of North-Borgou and Mekrou-Pendjari showed high value for height but low values for other parameters.
Cluster analysis and group characterization
The hierarchical clustering analysis revealed three morphological classes of individuals of S. spinosa with a genetic similarity index > 80%. Individuals in different clusters were 88% similar while the different clusters were just 12% similar. The scatterplot from the linear discriminant analysis (Figure S2) showed that the first discriminant function (86.22%) well separated the individuals of class 2 from the ones of classes 1 and 3. The second discriminant function (13.78%) achieves a fairly good separation of classes 1 and 3, and classes 2 and 3, although it is not totally perfect. The first class comprised 29 individuals (35.8%), whereas the second and third classes consisted of 7 (8.6%) and 45 individuals (55.6%), respectively (Fig. 3). The individuals of class 1 were mostly (87.67%) of Sudanian provenance. The seven individuals of class 2 were all (100%) of Sudano-Guinean provenance. The individuals of class 3 comprised 12 individuals (26.67%) from North-Borgou population in the Sudanian zone and 17 individuals (37.78%) from the three populations of Sudano-Guinean zone.

Phylogenetic constellation plots displaying the relationships between 81 individuals of S. spinosa trees. Blue stands for individuals of class 1; green stands for individuals of class 2; red for individuals of class 3
The first two principal components accounted for the total variability (100 %) among the individuals of Strychnos spinosa studied. The first principal component (Dim1) explained 86.52% of the overall variability and was positively correlated with all morphological descriptors (correlation > 0.9) except for the plant height (Table 5). Plant height was positively correlated with the second principal component (correlation = 0.970) which explained 13.48% of the overall variability. Fruit-related traits were discriminated by the first component and sorted the three morphological classes so that for these traits, class 2 < class 3 < class 1 (Fig. 4). In addition, the second component showed that class 1 has the highest value for plant height. This class is related to North-Borgou and Mekrou-Pendjari individuals while class 3 and class 2 were respectively linked to Bassila and South-Borgou, in one hand and Zou in the second hand.

Biplot showing the relationships between clusters and morphological traits of S. spinosa. Tree diameter; Height: tree height; Circ: fruit circumference; Thick: fruit thickness; DryM: fruits’ dry mass; SeedsN: seeds number; SeedsW: seeds weight; PulpM: pulp mass; Ratio: PulpM /DryM
Correlation among fruits and trees traits
Overall, the correlations among traits were positive and statistically significant (Fig. 5). There was no significant correlation between tree height and fruits’ seed weight and other descriptors. The ratio of pulp mass over fruits’ dry mass was highly correlated with all the descriptors except for plant height, and seed weight. The tree diameter was highly correlated with the fruit mass, the pulp mass and the number of seeds. The dry mass of the fruit was correlated with all the descriptors except seed weight and tree height while the pulp mass was correlated with all the traits except tree height and seed weight. The fruit circumference was correlated with the fruit thickness, fruit mass and pulp mass. The fruit thickness and seed number were both correlated with the fruit’s pulp mass, dry mass and the ratio, and respectively correlated with fruit circumference and tree diameter.

Correlogram showing the pairwise correlation between tree and fruit descriptors. P value significance: ‘***’ significant at 0.001, ‘**’ significant at 0.01, ‘*’ significant at 0.05, ‘–’ significant at 0.1; DBH: Tree diameter; Height: tree height; Circ: fruit circumference; Thick: fruit thickness; DryM: fruits’ dry mass; SeedsN: seeds number; SeedsW: seeds weight; PulpM: pulp mass; Ratio: PulpM/DryM
Identification of superior/desirable morphotypes of S. spinosa
Overall, individuals of class 2 showed good performances for the morphological traits (Table 6). This group was characterized by bigger individuals with the largest fruit, the highest amount of pulp and heavy seeds. The individuals of class 3, while being the smallest, scored well in terms of other parameters, which ranked them behind class 2. Individuals of class 1 were very tall, and had the lowest mean values for the different fruit descriptors. At population level, South-Borgou scored better for the assessed traits followed by the populations from Bassila and Zou (Table 7). At individual level, the elite individuals were found inside Bassila population. They showed morphological superiority in most of the traits assessed indicating a high potential for selection in the domestication process and can be considered for future research on the species’ improvement.
Discussion
Variation of morphological descriptors of S. spinosa
This study showed existence of heterogeneity in S. spinosa populations. This is most likely the result of the species’ fitness in various habitats which could be useful to survive and thrive under climate change (Visser 2008; Hounkpèvi et al. 2016) which impacts are already visible in Benin (e.g. flooding, irregular distribution of rainfall, etc.) (see Gnanglè et al. 2011). Considerable morphological variation exists in the measured traits. Our results on variance components of fruit traits support the hypothesis of higher morphological variation within populations than between populations and in accordance with several previous studies on other tree/shrub species in sub-Saharan Africa (Abasse et al. 2011; Cuni Sanchez et al. 2011; Korbo et al. 2012) in general and particularly in Benin (Fandohan et al. 2011; Gouwakinnou et al. 2011; Hounkpèvi et al. 2016).
All fruit features contributing to the fruit weight such as number of seeds, seeds’ weight, and pulp weight varied significantly between climatic zones and among provenances within climatic zones. The parameters related to the weight had relatively higher variability (pulp mass, seed weight, fruit dry mass) than those related to the volume (circumference and thickness). The high variation in the fruit dry mass and pulp mass is similar to the findings of Mkonda et al. (2003) on Strychnos cocculoides Baker fruits from Zambia, and indicated the potential of S. spinosa to be improved from the wild. Therefore as proposed by Mkonda et al. (2003), genetic selection and methods inducing pathenocarpy might help to increase these traits of great interest. In a study on Tamarindus indica L., Fandohan et al. (2011) reported large variations in pulp/fruit ratio among climatic zones and among individuals of the Sudano-Guinean zone in Benin. It can thus be said from this result that fruits having a greater size and mass have a higher pulp/fruit ratio. This may indicate that for superior morphotypes, increase in pulp mass is higher than that of the remaining part of the fruits. Indeed, for tree genetic resources conservation, intraspecific genetic variation is needed to ensure the future adaptability of the species (Allendorf and Luikart 2008). It comes out that S. spinosa is suitable for domestication because selection for improvement would be possible (Zobel and Talbert 1984; Msukwa et al. 2016).
Correlation between fruit traits and tree size
Fruit traits except seed weight were highly correlated among each other, but were not correlated with tree characteristics (height and in some case tree diameter). Thus, tree height cannot be used as an indicator of fruit production. Msukwa et al. (2016) also reported that height is a weak predictor for fruit production hence, cannot be used in indirect selection. However, the tree diameter can be a good indicator for fruit and pulp mass and ratio of pulp/fruit mass. There was also a strong relationship between fruit mass and pulp mass showing that for future selection for pulp trait, the fruit mass can be a good indicator (Leakey et al. 2005a; Fandohan et al. 2011). For this latter, the variability of the relationship between fruit mass and pulp mass confirms the differences between morphotypes and may have been driven by both ecological and genetic variation. Futhermore, the weight and volume related parameters were highly correlated. This strong relationship can be exploited in the selection process and may eliminate the need for multiple selection criteria (Mkonda et al. 2003). Since correlated quantitative fruit characters are of a major interest in any improvement programme, the search for a desired character may cause simultaneous changes in the other characters (Maghembe et al. 1998).
Estimation and analysis of phenotypic diversity
Previous studies on native fruit tree species in Benin such as that of Assogbadjo et al. (2011) on Adansonia digitata L., Gouwakinnou et al. (2011) on Sclerocarya birrea (A. Rich) Hochst. subsp. birrea, Fandohan et al. (2011) on T. indica and Hounkpèvi et al. (2016) on Vitex doniana Sweet, revealed a high level of diversity, indicating the high genetic potential of these fruit trees for species improvement in domestication program. This study also found large natural variation in monkey orange population collected from different climatic zones. From the estimated diversity indices presented in Table 4, it can be said that all the traits presented diversity indices beyond 0.40, showing intermediate to large genetic diversity among populations for each assessed trait. It is noteworthy that the individuals in sudanian zone are less diversified (H′ range from 0.51 to 0.57) than those of the Sudano-Guinean zone (H′ = 0.62–078). From this observation, the populations of North-Borgou and Bassila can be respectively considered as the less and the most diversified. This situation can be explained by the fact that the climate in the sudanian is more stable than the one of the Sudano-Guinean zone (transition zone). Thus, species in the transition zone will be more subject to adapting continuously to the climate fluctuations; thus causing different phenotypic variations in them.
Implications for the species conservation and domestication
Domestication and improvement of wild edible fruit trees hold great promise in agroforestry development (Mkonda et al. 2003). For this, the large variation found in fruit parameters of the species indicates considerable opportunities for phenotypic selection and quantitative improvement of the traits (Leakey et al. 2005a; Hounkpèvi et al. 2016). The high correlation of the tree diameter with most of the fruit descriptors is interesting since the trait for vigorous vegetative growth (dbh) is good for selection in the domestication process (Shackleton et al. 2002). The assessed traits showed an important variability between and within climatic zones. This is essential for adaptive evolutionary change and selection for domestication purpose since it could offer some possibilities to identify and select new cultivars through an oriented selection scheme (Gouwakinnou et al. 2011). The higher variation within provenances also indicates that rapid improvement progress could be gained through clonal propagation of elite trees with superior phenotypic traits such as higher fruit pulp or big fruit size (Maghembe et al. 1998). The cluster analysis showed three groups of trees with different traits’ performances. It came out that “plus tree” can be found in Sudano-Guinean zone and especially in the phytodistrict of Bassila. This suggests that best individuals are found where the climate is more clement (warm and humid). According to Shakeel et al. (2011) drought induces changes in morphological, physiological and biochemical changes in plants. It progressively decreases CO2 assimilation rates due to reduced stomatal conductance, reduces leaf size, stems extension and root proliferation, disturbs plant water relations and reduces water-use efficiency. This can explain why the individuals of Sudanian provenance did not have a good performance for the assessed descriptors and that the elite individuals were found in the Sudano-Guinean zone. Additionally, the best individuals revealed by this study bear heavy seeds which mean that their seed have enough reserve to germinate and facilitate the survival of seedlings in their early age. This parameter can be considered during the selection process for ideotypes of Sudanian zone. As reported by Gouwakinnou et al. (2011), natural selection in dry areas should favor bigger seeds to allow quick germination and seedling establishment at the onset of rainy season. Altogether, the high variability in phenotypic traits observed in this study indicated significant potential for improvement through the development of cultivars from elite trees using horticultural techniques (Leakey et al. 2008). According to Leakey et al. (2005a), cultivar selection would increase uniformity in the product, increase productivity and provide an incentive for farmers to plant monkey orange trees in their farming systems and home garden. For the species introduction in agroforestry system, rapid progress can be done by selecting superior morphotypes and propagating them as clones (El-Siddig et al. 2006).
Conclusion
The present study assessed variability of the morphological traits of S. spinosa individuals across zones in Benin and provided useful information for selection of appropriate ideotypes for a sustainable management of the species populations. The findings revealed existence of significant variations among individuals from different climatic zones for the studied descriptors and showed that elite trees can be found in the Sudano-Guinean zone. Moreover, it showed that tree diameter can be used to predict fruit descriptors. Therefore, this study paves the way for future investigations leading to setting up appropriate strategies for the species’ domestication and its introduction in traditional agroforestry systems in Benin. Nonetheless, the study was limited by repetition of the data with the time (only one season of data collection). Thus, future investigations are necessary on a larger group of individuals well spread across the country and should include other descriptors in order to reveal all the phenotypic and genetic variation within the species. This will enable a proposition of a definitive management strategies of the species. Further studies should concentrate on the reproductive biology in order to assess the heritability of the interesting characters of the elite individuals (big tree diameter, large fruit, high amount of pulp, heavy seed) found in this study.
Availability of data and materials
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
References
Abasse T, Weber JC, Katkore B, Boureima M, Larwanou M, Kalinganire A (2011) Morphological variation in Balanites aegyptiaca fruits and seeds within and among parkland agroforests in eastern Niger. Agrofor Syst 81(1):57–66. https://doi.org/10.1007/s10457-010-9323-x
Adomou AC, Sinsin B, Van der Maesen LJG (2006) Phytosociological and chorological approaches to phytogeography: a meso-scale study in Benin. SGP 76(2):155–178
Akoegninou A, van der Burg WJ, van der Maesen LJG et al (2006) Flore Analytique du Bénin. Backhuys Publishers, Cotonou
Allendorf FW, Luikart G (2008) Conserving global biodiversity? conservation and the genetics of populations. Conserv Genet 9(2):437–438. https://doi.org/10.1007/s10592-007-9330-2
Arbonnier M (2002) Arbres, arbustes et lianes des zones sèches d’Afrique de l’Ouest. CIRAD - Museum d’histoire naturel, Montpellier-Paris
Assogbadjo AE, Glèlè Kakaï R, Edon S, Kyndt T, Sinsin B (2011) Natural variation in fruit characteristics, seed germination and seedling growth of Adansonia digitata L. in Benin. New For 41(1):113–125. https://doi.org/10.1007/s11056-010-9214-z
Assogbadjo AE, Chadaré FJ, Glèlè Kakaï R, Fandohan B, Baidu-Forson JJ (2012) Variation in biochemical composition of baobab (Adansonia digitata) pulp, leaves and seeds in relation to soil types and tree provenances. Agric Ecosyst Environ 157:94–99. https://doi.org/10.1016/j.agee.2012.01.021
Avakoudjo HGG (2020) Ethnobotany and impacts of climate change on Strychnos spinosa Lam (Loganiaceae) distribution and tree growth in Benin. Ph.D. thesis, Graduate Research Program on Climate Change and Biodiversity of University Félix Houphouët-Boigny, Bingerville, Côte d’Ivoire
Avakoudjo HGG, Hounkpèvi A, Idohou R, Koné MW, Assogbadjo AE (2020) Local knowledge, uses, and factors determining the use of Strychnos spinosa organs in Benin (West Africa). Econ Bot 74(1):15–31. https://doi.org/10.1007/s12231-019-09481-0
Berihun T, Molla E (2017) Study on the diversity and use of wild edible plants in Bullen district northwest Ethiopia. J Bot. https://doi.org/10.1155/2017/8383468
Bero J, Hannaert V, Chataigné G, Hérent M-F, Quetin-Leclercq J (2011) In vitro Antitrypanosomal and antileishmanial activity of plant used in Benin in traditional medicine and bio-guided fractionation of the most active extract. J Ethnopharmacol 137(2):998–1002. https://doi.org/10.1016/j.jep.2011.07.022
Bruschi P, Mancini M, Mattioli E, Morganti M, Signorini M (2014) Traditional uses of plants in rural community of Mozambique and possible links with Miombo degradation and harvesting sustainability. J Ethnobiol Ethnomed 10:52. https://doi.org/10.1186/1746-4269-10-59
Cuni Sanchez A, De Smedt S, Haq N, Samson R (2011) Comparative study on baobab fruit morphological variation between western and south-eastern Africa: opportunities for domestication. Genet Resour Crop Evol 58(8):1143–1156. https://doi.org/10.1007/s10722-010-9647-4
El-Siddig K, Gunasena HPM, Prasad BA, Pushpakumara DKNG, Ramana KVR, Vijayanand P, Williams JT (2006) Tamarind, Tamarindus indica. Southampton Centre for Underutilised Crops, Southampton
Fandohan AB, Assogbadjo AE, Glèlè RL, Sinsin B, Van Damme P (2010) Impact of habitat type on the conservation status of tamarind (Tamarindus indica L.) populations in the W National Park of Benin. Fruits 65(1):11–19. https://doi.org/10.1051/fruits/2009037
Fandohan B, Assogbadjo AE, Glèlè Kakaï R, Kyndt T, Sinsin B (2011) Quantitative morphological descriptors confirm traditionally classified morphotypes of Tamarindus indica L. fruits. Genet Resour Crop Evol 58(2):299–309. https://doi.org/10.1007/s10722-010-9575-3
FAO (2018) Annexure-3: list of descriptors mulberry genetic resources at glance. http://www.fao.org/3/ad107e/ad107e0u.htm. Accessed 30 Aug 2018
Gnanglè CP, Glèlè Kakaï R, Assogbadjo AE, Vodounnon S, Afouda Yabi J, Sokpon N (2011) Tendances climatiques passées, modélisation, perceptions et adaptations locales au Bénin. Climatologie. https://doi.org/10.4267/climatologie.259
Gnanglè PC, Egah J, Baco MN, Gbèmavo CDSJ, Glèlè Kakaï R, Sokpon N (2012) Perceptions locales du changement climatique et mesures d’adaptation dans la gestion des parcs à karité au Nord-Bénin. IJBCS 6(1):136–149. https://doi.org/10.4314/ijbcs.v6i1.13
Gouwakinnou GN, Assogbadjo AE, Lykke AM, Sinsin B (2011) Phenotypic variations in fruits and selection potential in Sclerocarya birrea subsp. birrea. Sci Hortic 129(4):777–783. https://doi.org/10.1016/j.scienta.2011.05.041
Hoet S, Stévigny C, Hérent M-F, Quetin-Leclercq J (2006) Antitrypanosomal compounds from the leaf essential oil of Strychnos spinosa. Planta Med 72(05):480–482. https://doi.org/10.1055/s-2005-916255
Hoet S, Pieters L, Muccioli GG, Habib-jiwan J-L, Opperdoes FR, Quetin-Leclercq J (2007) Antitrypanosomal activity of triterpenoids and sterols from the leaves of Strychnos spinosa and related compounds. J Nat Prod 70(8):1360–1363. https://doi.org/10.1021/np070038q
Hounkpèvi A, Azihou AF, Kouassi ÉK, Porembski S, Glèlè Kakaï R (2016) Climate-induced morphological variation of black plum (Vitex doniana Sw.) in Benin, West Africa. Genet Resour Crop Evol 63:1073. https://doi.org/10.1007/s10722-016-0409-9
Idohou R, Assogbadjo AE, Azihou F, Glèlè Kakaï R, Adomou A (2016) Influence of the landscape context on stand structure and spatial patterns of the doum palm (Hyphaene thebaica Mart.) in the Republic of Benin (West Africa). Agrofor Syst 90(4):591–605. https://doi.org/10.1007/s10457-016-9920-4
Isa AI, Awouafack MD, Dzoyem JP, Aliyu M, Magaji RA, Ayo JO, Eloff JN (2014) Some Strychnos spinosa (Loganiaceae) leaf extracts and fractions have good antimicrobial activities and low cytotoxicities. BMC Complement Altern Med 14:456. https://doi.org/10.1186/1472-6882-14-456
JMP®, Version pro-14.10 (2019) SAS Institute Inc., Cary, NC
Korbo A, Sanou H, Ræbild A, Jensen JS, Hansen JK, Kjær ED (2012) Comparison of East and West African populations of baobab (Adansonia digitata L.). Agrofor Syst 85:505–518. https://doi.org/10.1007/s10457-011-9464-6
Leakey R, Fuller S, Treloar T, Stevenson L, Hunter D, Nevenimo T, Binifa J, Moxon J (2008) Characterization of tree-to-tree variation in morphological, nutritional and medicinal properties of Canarium indicum nuts. Agrofor Syst 73(1):77–87. https://doi.org/10.1007/s10457-007-9103-4
Leakey RRB, Fondoun JM, Atangana A, Tchoundjeu Z (2000) Quantitative descriptors of variation in the fruits and seeds of Irvingia gabonensis. Agrofor Syst 50(1):47–58. https://doi.org/10.1023/A:1006434902691
Leakey RRB, Tchoundjeu Z, Smith RI, Munro RC, Fondoun JM, Kengue J, Anegbeh PO, Atangana AR, Waruhiu AN, Asaah E, Usoro C, Ukafor V (2004) Evidence that subsistence farmers have domesticated indigenous fruits (Dacryodes edulis and Irvingia gabonensis) in Cameroon and Nigeria. Agrofor Syst 60(2):101–111. https://doi.org/10.1023/B:AGFO.0000013259.95628.22
Leakey RRB, Shackleton S, du Plessis P (2005a) Domestication potential of marula (Sclerocarya birrea subsp. caffra) in South Africa and Namibia: 1. Phenotypic variation in fruit traits. Agrofor Syst 64(1):25–35. https://doi.org/10.1007/s10457-005-2419-z
Leakey RRB, Pate K, Lombard C (2005b) Domestication potential of marula (Sclerocarya birrea subsp. caffra) in South Africa and Namibia: 2. Phenotypic variation in nut and kernel traits. Agrofor Syst 64(1):37–49. https://doi.org/10.1007/s10457-005-2420-6
Lumen Boundless Biology (2020) Population genetics. https://courses.lumenlearning.com/boundless-biology/chapter/population-genetics/#:~:text=Because%20natural%20selection%20acts%20directly,the%20allele%20in%20the%20population. Accessed 03 Nov 2020
Madzimure J, Nyahangare ET, Hamudikuwanda H, Hove T, Belmain SR, Stevenson PC, Mvumi BM (2013) Efficacy of Strychnos spinosa (Lam.) and Solanum incanum L. aqueous fruit extracts against cattle ticks. Trop Anim Health Prod 45(6):1341–1347. https://doi.org/10.1007/s11250-013-0367-6
Maghembe AJ, Simons AJ, Kwesiga F (1998) Selecting indigenous fruits trees for domestication in Southern Africa: priority setting with farmers in Malawi, Tanzania, Zambia and Zimbabwe. International Centre for Research in Agroforestry, Nairobi
Mengistu DK, Kirosa AY, Pè ME (2015) Phenotypic diversity in Ethiopian durum wheat (Triticum turgidum var. durum) landraces. Crop J 3(2015):190–199. https://doi.org/10.1016/j.cj.2015.04.003
Mille G, Louppe D (2015) Mémento du forestier tropical. Editions Quae, Versailles
Mkonda A, Lungu S, Maghembe JA, Mafongoya PL (2003) Fruit and seed germination characteristics of Strychnos cocculoides an indigenous fruit tree from natural populations in Zambia. Agrofor Syst 58(1):25–31. https://doi.org/10.1023/A:1025454231002
Msukwa V, Munthali CRY, Missanjo E, Chilima CZ, Mng’omba SA, Sagona W, Mkwezalamba I (2016) Growth performance and fruit production of Sclerocarya birrea (A. Rich.) Hochst. provenances in Malawi. IJRAS 3(2):42–49. https://doi.org/10.12983/ijsras-2016-p0042-0049
Nebel S, Pieroni A, Heinrich M (2006) Ta chòrta: wild edible greens used in the Graecanic area in Calabria, southern Italy. Appetite 47:333–342. https://doi.org/10.1016/j.appet.2006.05.010
Okello J, Okullo JBL, Eilu G, Nyeko P, Obua J (2018) Morphological Variations in Tamarindus indica LINN. Fruits and seed traits in the different agroecological zones of Uganda. I. J Ecol. https://doi.org/10.1155/2018/8469156
Orwa C, Mutua A, Kindt R, Jamnadass R, Simons A (2009) Agroforestree Database: a tree reference and selection guide version 4.0. World Agroforestry Centre, Kenya
Padonou EA, Tovissodé FC, Idohou R, Salako VK, Fantondji L, Vihotogbé R, Fandohan B, Assogbadjo AE (2017) Pilot assessment of locally acknowledged morphotypes of Irvingia gabonensis (Aubry-Lecomte) Baill. in southwestern Benin (West Africa). Fruits 72(5):306–316. https://doi.org/10.17660/th2017/72.5.6
Plantz Africa. Strychnos spinosa Lam. http://pza.sanbi.org/strychnos-spinosa. Accessed 11 Apr 2017
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Salifou CFA, Kassa KS, Ahounou SG, Moussa H, Dotché IO, Agbozo JM, Issifou MT, Youssao IAK (2017) Plantes lactogènes des bovins et leurs modes de préparation dans les élevages traditionnels au Bénin. Livest Res Rural Dev 29(2). https://www.lrrd.cipav.org.co/lrrd29/2/sali29025.html. Accessed 19 June 2017
Schippmann U, Leaman DJ, Cunningham AB (2002) Impact of cultivation and gathering of medicinal plants on biodiversity: global trends and issues. In: FAO Inter Departmental Working Group on Biological Diversity for Food and Agriculture (ed) Biodiversity and the ecosystem approach in agriculture, forestry and fisheries. Satellite event on the occasion of the Ninth Regular Session of the Commission on Genetic Resources for Food and Agriculture. FAO, Rome
Shackleton SE, Shackleton CM, Cunningham T, Lombard C, Sullivan CA, Netshiluvhi TR (2002) Knowledge on Sclerocarya birrea subsp caffra with emphasis on it as a non-timber forests product in South and southern Africa: a summary. Part1: taxonomy, ecology and role in rural livelihoods. S Afr For J 194(1):27–41. https://doi.org/10.1080/20702620.2002.10434589
Shakeel AA, Xiao-yu X, Long-chang W, Muhammad FS, Chen M, Wang L (2011) Morphological, physiological and biochemical responses of plants to drought stress. Afr J Agric Res 6(9):2026–2032. https://doi.org/10.5897/AJAR10.027
Sitrit Y, Loison S, Ninio R, Dishon E, Bar E, Lewinsohn E, Mizrahi Y (2003) Characterization of monkey orange (Strychnos spinosa Lam.), a potential new crop for arid regions. J Agric Food Chem 51(21):6256–6260. https://doi.org/10.1021/jf030289e
Thiombiano DNE, Lamien N, Castro-Euler AM, Vinceti B, Agundez D, Boussim IJ (2013) Local communities demand for food tree species and the potentialities of their landscapes in two ecological zones of Burkina Faso. OJF 3(3):79–87. https://doi.org/10.4236/ojf.2013.33014
UNFCCC (2018) Considerations regarding vulnerable groups, communities and ecosystems in the context of the national adaptation plans. ISBN: 978-92-9219-182-5. https://unfccc.int/sites/default/files/resource/Considerations%20regarding%20vulnerable.pdf. Accessed 25 Feb 2020
Visser ME (2008) Keeping up with a warming world; assessing the rate of adaptation to climate change. Proc R Soc B 275(1635):649–659. https://doi.org/10.1098/rspb.2007.0997
World Agroforestry Centre (2018) Strychnos spinosa. http://www.worldagroforestry.org/treedb/AFTPPDFS/Strychnos_spinosa.PDF. Accessed 25 May 2018
Zobel BJ, Talbert J (1984) Applied forest tree improvement. Wiley, New York
Acknowledgements
This research was supported by the German Federal Ministry of Education, through the West African Science Service Centre on Climate Change and Adapted Land-use (WASCAL). Salako Kolawolé Valère acknowledges the support of the Wallonie-Bruxelles International Post-doctoral Fellowship for Excellence, Belgium (Fellowship # SUB/2019/443681). We are very grateful to the anonymous reviewers whose comments and remarks contributed to the improvement of the quality of the paper. Charles Nelimor’s help for data analysis and language correction is acknowledged. We are also grateful to Edmond Sacla for his help in data analysis. We are indebted to local farmers and field guides for their substantial contribution to this work especially during field work.
Funding
This research was financially supported by the German Federal Ministry of Education and Research (BMBF) through West African Science Centre on Climate Change and Adapted Land Use (WASCAL) Ph.D. scholarship and a research budget allowance.
Author information
Authors and Affiliations
Contributions
AHGG: funding acquisition, investigation, formal analysis, Writing—Original Draft. IR: methodology, resources, writing—review & editing. SV: methodology, formal analysis, writing—review & editing. HA: methodology, formal analysis, writing—review & editing. KMW: supervision, writing—review & editing. AAE: conceptualization, supervision, resources, writing—review & editing.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Avakoudjo, H.G.G., Idohou, R., Salako, K.V. et al. Diversity in tree and fruit traits of Strychnos spinosa Lam. along a climatic gradient in Benin: a step towards domestication. Genet Resour Crop Evol 68, 2423–2440 (2021). https://doi.org/10.1007/s10722-021-01140-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-021-01140-5