
Abstract
Young tropical forests have higher relative plant densities of potentially nodulating Leguminosae species than old forests, but comparisons of their proportions of effective N-fixing plants and amounts of fixed N are scarce. We determined leaf N stocks derived from symbiotic biological N fixation in woody legumes of young and old forests under the following five edaphoclimatic conditions, in Northeastern Brazil: (1) humid lowland coastal; (2) humid montane; (3) subhumid high fertility soil; (4) semiarid high fertility soil; and (5) semiarid montane low fertility soil. N amounts derived from fixation and stocked in the leaves were estimated using the 15N natural abundance technique. Fixed N stocks in the leaves were low in both young and old forests of the nutrient-poor semiarid montane forests (< 0.3 kg ha−1) due to the low relative densities of nodulating legume plants (15 and 10 pl ha−1, respectively), despite most individuals of the nodulating species fixing N. The relative density of nodulating species in the four other sites was higher in the young (148 to 635) than in the old forests (42 to 327 pl ha−1), as was the density of effectively fixing plants (96 to 265 vs. 9 to 140 pl ha−1). Leaf N stocks were also higher in the young than in the old forests, especially in the coastal humid forest (15.6 vs. 3.2 kg ha−1), except in the humid montane site (2.4 vs. 6.2 kg ha−1), due to the large N-fixing legume leaf biomass of the old forest. N fixation seems to provide an ecological advantage over non N-fixing plants in young tropical forests, usually growing on N poor soils, but not in semiarid forests growing on soils which are poor in other nutrients, like P, in which fixation is very low. However, the advantage of N-fixing legumes may also be related to other environmental variables and fixation continues to play a role in old forests.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen (N) is one of the most limiting nutrients for plant growth but can be provided by diazotrophic bacteria capable of biological N fixation (BNF) in symbiosis with vascular plants. Symbiotic N fixation has been considered the most important natural N input in terrestrial ecosystems (Boddey et al. 2000), but other sources (biological or not) may contribute, and this contribution may be smaller (Davies-Barnard and Friedlingstein 2020; Moreira et al. 2021; Sullivan et al. 2014), similar, or even higher (Reis et al. 2020). There is a large diversity of potentially nodulating or potentially N-fixing Leguminosae species in neotropical forests, indicating that the contribution of N fixation is an important ecological strategy (Gei et al. 2018). Interest in understanding BNF has increased in recent years, both in dry (Freitas et al. 2015; Silva et al. 2017) and humid tropical forests (Nardotto et al. 2014; Taylor et al. 2019; Wong et al. 2020). However, the number of studies is still small, particularly for forests in different successional stages (Batterman et al. 2013; Lai et al. 2018; Sullivan et al. 2014; Taylor et al. 2019; Winbourne et al. 2018), and for those growing in nutrient-poor soils, where other nutrients may be more limiting than N (Gei et al. 2018; Vitousek et al. 2013).
The abundance of Leguminosae populations and especially of potentially nodulating species tend to be higher during early forest regeneration stages than in old neotropical forests (Batterman et al. 2013; Brookshire et al. 2019; Costa et al. 2021; Sullivan et al. 2014). It has been hypothesized that BNF can supply the demand of N while biomass is accumulating (Gehring et al. 2005), and that the occurrence of high BNF is generally associated with a low N supply relative to the availability of other resources, such as phosphorus, molybdenum, and potassium (Crews 2016; Vitousek et al. 2013). Nodulating species in old successional stages may lose their ecological advantage if the soil–plant system has already accumulated enough N. In this case, non-nodulating species may dominate the vegetation, resulting in little N input through BNF (Sullivan et al. 2014). The nodulating species may also maintain high proportions of the vegetation biomass and plant density but limit photosynthate transfer to the N-fixing bacteria in the nodules which, in turn, reduce N fixation (Faye et al. 2007; Hedin et al. 2009). Thus, an individual belonging to a potentially nodulating species may not necessarily be an effectively N fixing plant. The higher enrichment in the 15N isotope in the soil–plant system indicates that there are higher relative N losses in dry than in humid tropical forests, losses preferentially involving the lighter 14N isotope (Aranibar et al. 2004; Craine et al. 2009, 2015; Freitas et al. 2015; Swap et al. 2004; Viani et al. 2011). Therefore, it would be expected that N fixation would be proportionally higher in dry than in humid forests and especially in younger than in older stands before enough N accumulates in the soil–plant system. However, humid forests have higher biomass than dry forests and may also have higher Leguminosae biomass (Costa et al. 2021). Higher biomass may compensate for the lower Leguminosae proportions and absolute N fixation could be as high or even higher than in dry forests.
Northeastern Brazil has great variability of rainfall, soil, and geomorphological conditions, which create a large diversity of forest types (Sampaio 2010). Following an east-to-west transect, there are humid low-land forests in the coastal area (Atlantic Forest), followed by subhumid deciduous thorny forests (Agreste caatinga), semiarid deciduous thorny forests (Sertão caatinga), montane humid forests in crystalline inselbergs within the semiarid area (montane forest) and semiarid deciduous non-thorny montane forests (carrasco) in the sedimentary plateau of the far west. The first four forests grow in medium to high fertility soils, while the semiarid montane forest grows in nutrient-poor, phosphorus-deficient soils (Salcedo et al. 1997). Leguminosae species, mainly nodulating ones, are dominant among families in the four first forests and have higher aboveground biomass and plant density in young than in old stands (Costa et al. 2021). The number of nodulating plants in the semiarid montane forest growing in nutrient-poor, phosphorus-deficient soils are low in both young and old stands, indicating that BNF would be low. However, a few plants of nodulating species are present in both young and old stands, and they could have different fixation proportions, since plants in the old stand could have already accumulated enough N and could tend to transfer less carbohydrates to the symbiotic bacteria, by deriving less benefit from the fixation (Faye et al. 2007; Hedin et al. 2009).
The soil and plant 15N signals of these Brazilian forests indicate that they differ in the relative losses and internal cycling of the N in their soil–plant systems, with higher losses in the drier areas and especially high in the semiarid low-fertility montane forest (Freitas et al. 2015). Thus, fixation may also differ among these forest types, but data on amount of fixed N in Brazilian Northeast forests is only available for those in the dry areas where fixation was higher in old than in young stands (Silva et al. 2017; Souza et al. 2012), contrary to the pattern assumed to be more common in other areas (Gei et al. 2018).
Considering the current interest in BNF and the possibility of studying nearby young and old forest stands under different environmental conditions, we estimated the proportions of N stocks derived from BNF in the leaves of woody species of potentially nodulating Leguminosae in forest stands with different regeneration ages. Our objectives were: (1) to elucidate if the amounts of fixed N in the leaves calculated in a land area base vary across forests under five contrasting edaphoclimatic conditions in the Brazilian Northeastern region; and (2) to determine if the amounts of N stocks in the leaves derived from fixation are higher in the young than in the old forests under these conditions. Our first hypothesis is that the proportion of individuals of nodulating species fixing N is higher in the dry forests, except in the dry forest growing on low fertility soil, but that the fixed N stocks may be higher in the humid forests because of their higher leaf biomass of nodulating species. Our second hypothesis is that the proportions of N-fixing Leguminosae plants and leaf N stocks derived from fixation are higher in the young than in the old stands of these forests.
Material and methods
Characteristics of the study areas
A total of 10 native vegetation fragments distributed in five sites were selected for the study, each one with two forest stands, one old and one young, comprising forests with different land-use histories, and following an east–west transect of about 560 km of extension in the Brazilian Northeastern region (Supplementary Fig. S1). The area ranged from 7° 42′ 51.7″ S and 34° 59′ 35.4 W to 7° 19′ 25.5″ S and 40° 05′ 00.1″ W. The vegetation of the selected sites (Table 1) corresponds to: (1) humid coastal forest, henceforth designated as coastal forest; (2) subhumid deciduous forest, part of the hypoxerophytic caatinga, growing on medium to high fertility soils, henceforth designated as subhumid forest; (3) semiarid deciduous forest, part of the hyperxerophytic caatinga, growing on medium to high fertility soils, designated as semiarid high fertility forest; (4) humid montane forest, growing in high fertility soils, designated as humid montane forest; and (5) semiarid deciduous forest, growing in low fertility soils, henceforth called semiarid low fertility forest. The two forest stands in each site were at least 0.3 and at most 17 km apart. The ages and previous land uses were based on information obtained from local residents, property appointments and satellite image sequences. The native forest in all sites had been clearcut and crops had been planted for some years, with the cultivation being abandoned for a longer period in the old stands. A few small trees in the humid montane forest had been cut for firewood in the old stand 30 years before the study was conducted. The soils of the sites and the average annual temperatures were described in previous articles (Costa et al. 2021; Freitas et al. 2015).
Leaf and soil collection and estimate of leaf biomass
First, ten 20 × 20 m plots were established along one transect in each forest stand with 10 m distance between each consecutive plot (Costa et al. 2021). All living plants with stem diameter at 1.3 m from the soil (DBH) equal to or higher than 0.03 m were identified and had their stem diameters measured in each plot of the subhumid and semiarid forests. DBH measurement and identification were also performed in the humid coastal and montane forests, but the inclusion criterion was DBH ≥ 0.05 m. Maximum DBH in the old stands were: 0.62 m in the coastal forest; 0.47 m in the subhumid forest; 0.53 m in the semiarid high fertility forest; 0.86 m in the humid montane forest; and 0.2 m in the semiarid low fertility forest. Densities of the plants per unit area in the same forest stands had been determined before, counting all the plants in the same plots and extrapolating to the standard area of one hectare by multiplying the density in each plot by 25 (Costa et al. 2021).
The leaf biomass of each plant (B, kg) in all plots of all sites was estimated by allometric equations using DBH as the independent variable. The biomass equations for the subhumid and semiarid forests were (Silva and Sampaio 2008): B = 0.0681 DBH1.5829 for species with large stem diameters and B = 0.0180 DBH1.845 for species with small stem diameter. The classification of large (≥ 0.2 m DBH) and small stem diameters (< 0.20 m DBH) were based on the potential size of each species (Sampaio and Silva 2005). Considering the absence of specific allometric equations for leaf biomass in the humid forests of the region, the total aboveground biomass of the tree was estimated using the equation developed by Chave et al. (2001), in which B = EXP (− 2.19 + 2.54 LN (DBH)), and adopting the proportions of 10% of leaf biomass for trees with DBH < 0.1 m and 5% of leaf biomass for trees with DBH ≥ 0.1 m in relation to the total aboveground biomass (Burger and Delitti 1999). The sum of the leaf biomasses of all plants of each species and of the total for all plants in the plots of each stand (4000 m2) was extrapolated to 1 ha (10,000 m2).
Leaf samples were collected in each plot from up to five plants of each Leguminosae species indicated in the literature as a potentially nodulating species or from all plants when less than five plants of the species occurred in the plot. Leaves of non-nodulating Leguminosae species and species of other families were also collected to serve as reference species in the calculations of biological N fixation by the 15N natural abundance technique (Supplementary Table S1). The leaf material was composed of green, fully expanded and apparently healthy leaves (blades without holes, cuts or bruises and with green color typical of the species), being cut from various canopy positions to form a mixed sample. All collections were done when the leaf canopies were fully developed and before the leaves started to senesce and dry, approximately past the middle of the usual rainy season (Supplementary Fig. S2). These samples were used to determine the leaf N and 15N concentrations of each potentially nodulating species and to calculate their N and 15N contents based on the leaf biomass stock of each of these species (described below).
Botanical material of each species was collected for identification by specialists by comparison with information from the specialized literature and with specimens deposited in the Federal Rural University of Pernambuco herbarium (PEUFR). The scientific names of the species were confirmed consulting the W3 Tropics database (www.tropicos.org). The current classification of the Leguminosae subfamilies followed The Legume Phylogeny Working Group (LPWG 2017).
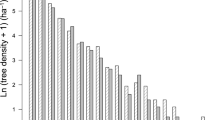
In addition, 10 samples were randomly collected from the 0.2 m superficial soil layer from each plot and mixed into a single composite sample per plot. Then, five from these 10 composite samples were randomly selected to be sieved, air-dried and analyzed for their carbon (C) and N concentrations (Table 2) and their 15N abundances (Fig. 1) to estimate the magnitude of the N losses and to serve as a control for the 15N signals of the reference species.

Relation between leaf δ15N of reference plants and mean soil δ15N of each stand of different forest types in the Brazilian Northeastern region
Estimates of symbiotic biological N fixation
The plant and soil samples were dried at 65 °C for 72 h and ground to fine powder. Their C and N concentrations and N isotope ratios were determined by mass spectrometry. A ThermoQuest-Finnigan Delta Plus (Finnigan-MAT; CA, USA) spectrometer, coupled to an element analyzer (Carlo Erba model 1110; Milan, Italy), belonging to the Ecology Laboratory of CENA—(USP), was used. Reference materials (athropine, yeast and standard soil 502,308 (LECO Corporation) were included in each analytical run. The 15N abundances were expressed in δ units (‰) in relation to the international standard:
in which: Rsample and Rreference are the ratios of the sample and the international standard (N2), respectively.
Effective atmospheric N fixation by potentially nodulating legume plants was assumed to have had occurred when the δ15N signal of each nodulating legume plant was lower than the average value of the reference species in the area, minus the standard deviation of the reference species signal multiplied by 1.96 to account for the 95% confidence interval of the normal distribution. Alternatively, effective N fixation was also assumed to have occurred when the δ15N signal of each nodulating legume plant was lower than 2 ‰ of the average value of the reference species in the plot (Silva et al. 2017). Both are conservative ways of estimating biologically fixed N, since plants with 15N signals above these thresholds may have some N derived from the atmosphere, although in small proportions. This may result in underestimating the fixed N stocks in the whole leaf biomass of all Leguminosae. The proportion of effectively nodulating plants in relation to the total plants of each species in the young and old stands of each site were calculated considering the 10 plots, since the numbers of plants of each species in each plot were often too low to enable a confident evaluation. Therefore, the data of these proportions had no replication and no statistical analysis. These proportions were multiplied by the leaf biomass of each species in each plot of the young and old stands of each site and were then used to calculate the leaf biomass of the effectively fixing plants species.
Next, the proportion of plant N derived from the atmosphere (%Ndfa) in the plants identified as having effectively fixed N, was estimated using the formula described by Shearer and Kohl (1989):
in which: δ15N(reference) is the abundance of 15N of the non-fixing reference species in the area, δ15N(target) is the abundance of 15N of the N-fixing plant in the field of a certain Leguminosae species, and δ15N (no N) is the value of δ15N for fixing plants of this same species grown in pots in the absence of N. The δ15N (no N) value was assumed to be − 1.24 ‰ (Silva et al. 2017).
We estimated two different amounts of fixed N (kg ha−1): (1) the amounts in the leaves of the individuals which were randomly selected to be analyzed for their 15N signals. This was estimated by multiplying the leaves biomass by the leaves N concentrations and the leaves %Ndfa; and (2) the amounts in the leaves of the set of the effectively fixing individuals of each species in each stand, which was estimated multiplying the amount of N in the leaves of all individuals of the species in the plot by the proportion of the effectively fixing N individuals in relation to the total number of individuals of each species in the stand and by the average proportion of plant N derived from the atmosphere (%Ndfa) of each species in the stand of each site.
Statistical analyses
Data of total, Leguminosae and potentially nodulating Leguminosae leaf biomass (kg ha−1) and of Leguminosae, potentially nodulating Leguminosae and effectively fixing plants’ leaf N contents (kg ha−1) were submitted to analysis of variance considering a split-plot (nested) design in which the main plots corresponded to the five sites and the split-plots to the young and old stands in each site. The proportions (%) of the number of effectively fixing plants in relation to the number of nodulating, Leguminosae and total plants (densities) were analyzed considering the same design. The 10 plots of each stand were considered as replicates. Data on soil C and N concentrations (%) and isotope abundance (‰) were also analyzed as a split-plot design, considering the five composite samples in each stand of each forest as replicates. The data were log-transformed to attend the normality criteria. The means were compared by the Tukey’s test at the 0.05 significance level. Analyzes were carried out using the split-plot function of the agricolae package of the R program (R Core Team 2020).
Results
Differences in soil and plant 15N signals
The soil δ15N isotope signals (Fig. 1) significantly increased westward from the humid coastal forest (4.3 and 6.8‰, in the old and young stands, respectively) to the semiarid low-fertility forest where they were very high (13.7 and 14.7‰). Lower values occurred in the humid montane forest (5.1 and 7.0‰) than in the geographically close semiarid forest (10.8 and 10.2‰). Total carbon and nitrogen concentrations (Table 2) in the soil surface layer (0–0.0 m) were higher in the humid montane forest than in the coastal forest and in the other three deciduous forests, and they were lowest in the young semiarid high-fertility forest stand.
The pattern of soil δ15N signals was reflected in higher leaf δ15N values of reference species in the subhumid and semiarid forests, intermediate values in the humid montane forests and lower in the humid coastal forests (Fig. 1 and Supplementary Table S2). The signals of the reference species were slightly lower than those of the soils across all forests. The variability of the δ15N averages of the reference plants within each site was relatively low and the calculations of %Ndfa using the standard deviation of the signals of the reference species or the fixed value of 2‰ (Boddey et al. 2000; Freitas et al. 2010, 2015) to establish the threshold of N fixation were not much different. Therefore, the calculations using the 2‰ fixed value are not presented.
Potentially nodulating species had a wide range of leaf δ15N values in all forests, which corresponded to high variations in %Ndfa on the same site, with an absence of or low symbiotically fixed N in the leaves of some plants, but proportions of fixed N greater than 60% in the leaves of other plants (Supplementary Table S2). The average %Ndfa of the effectively N-fixing Leguminosae was higher in the young than in the old stands of the subhumid forest and the humid montane forests, but the %Ndfa was not significantly different between young and old stands of the other three forests. Thus, any difference between the amounts of fixed N in the leaves of young and old stands in the humid coastal and semiarid forests was not due to higher proportions of fixed N in the leaf tissues, but to differences in plant density per area, leaf biomass stock and/or leaf N content. These differences could be influenced by the different intensity of drivers in the young and old forests, including lower nutrient but higher light availability (Batterman et al. 2013).
Plant density, leaf biomass and N content in the young and old forests
Total, Leguminosae and potentially nodulating Leguminosae plant densities were presented in a previous article (Costa et al. 2021). In summary: (1) total plant densities were similar between the young and the old stands of all forests, except in the montane forest in which the plant density was significantly higher in the young than in the old stand; (2) plant densities of Leguminosae species were higher in the young stands of three of the forests (humid coastal, humid montane and semiarid high fertility) and similar in the other two forests; and (3) plant densities of potentially nodulating species were higher in the young than in the old stands of all forests, except in the semiarid low-fertility forest, where young and old stands had similar plant densities (Supplementary Table S3). The density of the plants with leaf 15N signals indicating that they had symbiotically fixed N (designated as effectively fixing plants), measured in the present article, followed the same trend, being significantly higher in the young than in the old stands of all forests, except the semiarid low-fertility forest, which had low and the same density in both stands (Table 3). The proportions of effectively fixing plants in relation to the total number of plants of the same potentially nodulating species were higher in the young stands of three forests (50 to 65% vs. 21 to 43%) and similar in the semiarid high fertility (32 vs. 28%) and semiarid low fertility forest (67 vs. 100%), These proportions were high and not significantly different in the few nodulating species of both young and old stands of the semiarid low-fertility forest (67 and 100%).
All five old forest stands had higher leaf biomass than the young stands considering the set of all species (Fig. 2). However, considering the set of all legume species, only two of the old stands (the subhumid deciduous forest and the humid montane forest) had higher leaf biomass than the young stands. This indicates that legume leaves comprised a smaller proportion of the total leaf biomass in the old than in the young stands, but were at least 25% in all forests except in the old coastal forest stand (12%).Considering the set of the potentially nodulating species, the young stands of the humid coastal forest and of the semiarid high-fertility forest had higher leaf biomass than the old stands (Fig. 2). The opposite occurred with the humid montane forest and the subhumid high fertility forest, namely that the old stands had higher biomass. However, in calculating them as proportions to the leaf biomass of all legume trees, those of the nodulating legumes were higher in the young than in the old stands in all forests, except in the semiarid low-fertility forest where the proportions were low in both stands. These results are similar to those of aboveground biomass (Costa et al. 2021), since both were calculated based on the stem diameters.

Leaf biomass of the plants which had 15N signals indicating that they had N originated from symbiotic fixed N (A) and belonging to potentially nodulating Leguminosae species (B) and of the plants belonging to Leguminosae species (C) and plants of all families in the stand (D), in young and old stands of different forest types in the Brazilian Northeastern region. Standard errors of ten replicates. Bars with the same small letters comparing young and old stands of the same forest type and the same capital letter comparing either all young or all old forest stands are not significantly different by the Tukey test (p = 0.05)
The leaf biomass of the plants which were effectively fixing N followed a similar trend to that of the potentially nodulating plants, except that not only the young stands of the coastal and semiarid high-fertility forests, but also the young stand of the subhumid forest had higher leaf biomass than the old stands. The leaf biomass proportions of the effectively fixing plants in relation to the total legume leaf biomass were higher in the young than in the old stands, except for the semiarid low-fertility forests, which were very low (2%). The ranges in the young (23 to 39%) and in the old stands (8–12%) of the first four forests were relatively narrow considering the large variations in their biomass. Despite the low density of legume trees (Supplementary Table S3), the very high legume leaf biomass of the montane forest (Fig. 2) means that these trees had high leaf biomass per plant.
Mean leaf N concentrations of all reference species in the young and old stands of the five sites varied from 1.4 to 2.8%, while those of the effectively fixing legumes varied from 2.7 to 4.1% (Supplementary Table S2). In most cases, the N concentrations were higher in the effectively fixing legumes than in the reference species of each site. The concentrations in the potentially N fixing legumes were higher in the old than in the young stands of all forests, except for the humid coastal forest, while the concentrations in the reference species were higher in the old stands of only two forests (subhumid high fertility and humid montane).
Since biomass differences among forests were much larger than those in leaf N concentrations, the patterns of differences in N contents were similar to the biomass patterns (Figs. 2 and 3): the leaf N content of the Leguminosae species was higher in the young than in the old stands of all forests, except the montane forest, which was the only forest in which the leaf N content of the potentially nodulating species and of the effectively fixing plants was also higher in the old than in the young stand.

Actual amount of fixed N in the leaves (A) and leaf N stocks of the plants which had 15N signals indicating that they had N originated from symbiotic fixed N (B), leaf N stocks of the plants belonging to potentially nodulating Leguminosae species (C) and of the plants belonging to Leguminosae species (D) in young and old stands of different forest types in the Brazilian Northeastern region. Standard errors of ten replicates. Bars with the same small letter comparing young and old stands of the same forest type and the same capital letter comparing either all young or all old forest stands are not significantly different by the Tukey test (p = 0.05)
Fixed N stocks in the leaves of the young and old forests
The amounts of N derived from N fixation in the leaf biomass per land area of the effectively fixing plants in the subhumid and semiarid high fertility forests (Fig. 3) were higher in the young (3.1 and 5.0 kg ha−1, respectively) than in the old stands (1.0 and 1.6 kg ha−1). The lower amount of N fixed in the old stand of the subhumid forest reflected the low proportion of effectively N-fixing A. colubrina plants (13% of the A. colubrina plants), which was the species with highest leaf biomass. The proportions of N derived from fixation (%Ndfa) were higher in the young stands (46 and 58% in the subhumid and semiarid high fertility) than in the old stands (23 and 27%, respectively), in both sites. Of the two humid montane stands, the old stand had more leaf fixed N than the young one (6.2 and 2.4 kg ha−1), despite the absence of N fixation of A. colubrina. The low number of nodulating species in the semiarid low fertility forest and their low leaf biomass resulted in low amounts of fixed N in both young and old stands. The young stand in the humid coastal forest had the highest absolute amount of fixed N (15.6 kg ha−1) among all forests due to the much higher leaf biomass among all young forests (6.7 Mg ha−1 vs. 1.1 to 1.8 Mg ha−1). The superiority of the fixed N stock in the coastal forest in relation to the stocks in the subhumid and semiarid forests could be even greater considering that the inclusion criteria in the dry forests (diameter ≥ 3 cm) resulted in relatively more plants being measured than in the humid forests (diameter ≥ 5 cm).
In addition to A. colubrina, only four other species occurred in more than one site, reflecting the different edaphoclimatic conditions of the sites. Of these four species, only one (Piptadenia cf. stipulacea) occurred in both the young and the old stand of two sites (the subhumid and semiarid high fertility sites, like A. colubrina), and also in the young stand of the montane forest. The proportion of N derived from fixation of this species was higher in the montane forest (67%) than in the other forests (32 to 42%). Albizia cf. polycephala and Inga thibaudiana occurred in both the coastal and the montane forest, but the former occurred in both stands of the montane forest and only in the young stand of the coastal forest, while the second species only in the young stand of the montane and in the old stands of the coastal forest. A. polycephala had high proportions of fixed N in both forests (60 to 78%), while the single sampled plant of I. thibaudiana in the coastal forest was not fixing, and the three plants in the montane forest seemed to derive all their N from fixation. Chloroleucon dumosum occurred in both the high (only in the old stand) and the low fertility semiarid forests, with higher proportions of fixed N in the low fertility semiarid forest (37 and 64% versus 24%).
Discussion
High soil 15N signals indicate high N losses in the soil–plant systems
The high 15N signals in the soil and reference plants (Fig. 1) indicate high accumulation of 15N which results from the preferential losses of the lighter 14N isotope from the soil–plant system (Handley et al. 1999). The higher signals in the semiarid than in the humid forests can be interpreted as higher N losses in the system under the seasonally dry climatic conditions, contrasting with higher N recycling in the humid forests where water availability is higher (Aranibar et al. 2004; Coletta et al. 2009; Craine et al. 2009, 2015; Eshetu and Högberg 2000; Freitas et al. 2010b, 2015; Swap et al. 2004; Viani et al. 2011). The fact that the 15N signals of the leaves of the reference species were slightly lower in all forests than those of the soil reflect the absorption of mineralized forms of N which are depleted in the heavier isotope (Amundson et al. 2003; Freitas et al. 2015). The low variability of their 15N signals indicates that all the plants of the reference species absorbed N from the same soil sources and it is very likely that the plants of the target (Leguminosae) species also absorbed N from the same soil sources. This is a basic assumption in the use of the isotope technique to estimate N fixation (Boddey et al. 2000), and the low variability of the averages give more confidence that the technique was correctly used. The 15N signals of the N fixing target species are still lower, but much more variable due to the influx of atmospheric N2 mediated by the symbiotic bacteria (Boddey et al. 2000), which may differ depending on the conditions of each individual.
Higher fixed N stocks in young than old forests, with exceptions
The higher leaf N fixed stocks in the young than in the old stands in three of the five forest types (coastal forest and subhumid and semiarid high-fertility forests) (Fig. 3) seems to confirm the hypothesis that N fixation confers an ecological advantage during the initial successional stages of neotropical forests (Batterman et al. 2013; Gei et al. 2018; Sullivan et al. 2014). However, the absence of a significant difference in the semiarid low-fertility forest and the higher N fixed stock in the old than in the young stand of the humid montane forest indicate that this advantage may be overrun by other factors. These exceptions deserve further attention and are discussed below. It also must be highlighted that the advantage of N-fixing of some legume species in the initial successional stages may occur concomitant with other adaptations to the ecological conditions of these stages, such as facilitated seed arrival and seedling establishment and growth with high temperature and light penetration (Oliveira et al. 2023).The low N fixation in both young and old stands of the semiarid low-fertility forest due to the low plant density of potentially nodulating species (Costa et al. 2021) indicates that the contribution of N fixation to the N cycle of the soil–plant system is low and confers little ecological advantage in such environments. However, the relatively low soil N concentration (Table 2) and the high 15N signals of both stands, which indicate high N losses in the soil system (Aranibar et al. 2004; Craine et al. 2009, 2015; Freitas et al. 2015), suggest that N is not an abundant factor in the system. It is likely that N fixation is limited by one or more soil factors. These limiting factors may not include the absence of diazotrophic bacteria, since the few plants of the three potentially nodulating species derived more than half of their N from the atmosphere (Supplementary Table S2). Soils in the Araripe plateau have low available P, Ca and Mg (Salcedo et al. 1997; Sampaio 2010) and these low availabilities may limit N fixation. A connection of low P and low N fixation has been observed in a caatinga forest growing on a low-fertility sandy soil in the same Brazilian Northeastern region (Silva et al. 2017), as well as in forests of other tropical and temperate regions (Binkley et al. 2003; Vitousek et al. 2013). The apparent stronger limitation of P or other nutrient than N limitation reduces the possibility of any ecological advantage of potentially nodulating species, with the advantage probably being attained by species which are efficient in P absorption and recycling.
Soils in the humid montane forest, especially in the old stand, had the highest N concentrations among all forests, and lower 15N signals than the surrounding semiarid, high-fertility forest, although still positive (Table 2 and Fig. 1). These results indicate that there are net N losses in the soil of the humid montane forest but lower than those of the semiarid forest. In this case, N fixation may not confer an ecological advantage. In fact, the plant densities of potentially fixing species in relation to the total Leguminosae plant density were lower in the montane forest (29% in the young and 24% in the old stand) than in the semiarid, high-fertility forests (86 and 40%, respectively). The density proportions of the effectively fixing plants in relation to the total Leguminosae density were also lower in the humid montane forest (19 and 5%) than in the semiarid, high-fertility forests (27 and 11%). However, N fixation in both stands were of the same order as those of the semiarid, high-fertility forest and higher in the old (6.2 kg ha−1) than in the young stand (2.4 kg ha−1). When N is not limited in the system, some plants may reduce or even stop N fixation to avoid the energy costs of symbiosis (Hedin et al. 2009; Sullivan et al. 2014). The larger amount of fixed N in the old stand of the montane forest resulted from its large leaf biomass, since the proportion of effectively fixed leaf N in relation to the nodulating species’ leaf N was almost four-fold higher in the young than in the old stand (19 vs. 5%).
The estimates of the N stocks in the leaves (total, Leguminosae, nodulating and effectively fixed) are certainly underestimations of the total fixed N in the plants for two reasons. First, some fixed N was also directed to the other plant parts and the aggregate biomass of these other parts (roots, trunks, and branches) is larger than that of leaves, but they usually have lower N concentrations and their biomass accumulates over a longer period (Dickson 1986; Millard and Grelet 2010). Second, the leaf biomass and N stocks can be considered estimates of the annual production, considering that the leaf biomass is turned over every year in Brazilian (Amorim et al. 2009) and other dry deciduous forests (Chaturvedi and Raghubanshi 2016). The leaves of most of the species in both forest types have high specific leaf area (Amorim et al. 2009; Monteiro et al. 2017), which indicates short life spans (Monteiro et al. 2017; Selaya et al. 2008) and some of them may have fallen before or developed after the biomass peak when the measurements were done. Leaves are the most easily measured part of the plant and most of the published estimates of symbiotic N fixation using the 15N isotope technique are based on leaf N stocks and 15N abundances (Bodey et al. 2000; Coletta et al. 2009; Craine et al. 2009; Crews et al. 2016; Freitas et al. 2010, 2015; Gehring et al. 2005; Nardoto et al. 2014; Roggy et al. 1999; Silva et al. 2017; Souza et al. 2012; Viani et al. 2011). We could have estimated the fixed N stocks in the other plant parts if we had assumed that their 15N signals were equal to those of the leaves. However, comparisons of 15N abundances in different plant parts of tropical forests are scarce, with indication that roots and shoots have different abundances (Gathumbi et al. 2002). Data on these comparisons are badly needed because they are critical to estimate the total N fixation of the forests.
Higher difference in fixed N between young than old stands in the coastal forest
The coastal forest had the largest difference between the young and the old stand among the three forests in which the young stands had significantly higher leaf N stocks than the old stands (Fig. 3). The young coastal forest stand had the highest absolute fixed N stock (15.6 kg ha−1) among all stands of all forests and the differences for the dry forests could be even higher because the measurements in the humid forests were made with less inclusive criteria (5 vs. 3 cm of stem diameter) than the dry forests. This large N stock in the young stand resulted from the highest leaf biomass among all young stands, and the fact that a large part of the Leguminosae leaf biomass belonged to effectively fixing plants (Figs. 2 and 3). All nodulating species present in the young stand of the coastal forest were absent in the old stand (Supplementary Table S2), and the low 15N signal in both stands indicates that the soil–plant system may have a relatively high internal recycling, suggesting that N fixation may provide an ecological advantage in the initial successional stages, but then disappears in the later stages. Old stands of other tropical and subtropical humid forests also had low 15N signals and BNF fixation (Gehring et al. 2005).
Lower N fixation in old than younger stands has been estimated in a humid forest in Panama (Batterman et al. 2013), but not in another humid coastal forest in Brazil (Winbourne et al. 2018), both using the acetylene reduction assay. No evidence of facilitative or competitive effect of N2 fixers was found over the first three decades of growth of a humid forest in Panama (Lai et al. 2018). However, there may be higher free-living than symbiotic N2 fixation in some of these forest (Gehring et al. 2005; Reis et al. 2020).
The amount of fixed N in the young stand (15.6 kg ha−1), if assumed to represent the production over one year as discussed before, is higher than the latest estimations of fixation in humid forests in general, which had varied from 5 (Reis et al. 2020) to 8 kg N ha−1 y−1 (Davies-Barnard and Friedlingstern 2020). Considering that the amounts of fixed N in the present work only refer to the stocks in the leaves, much higher figures would be expected if the amounts of fixed N in the rest of the plant were included. Certainly, more data and improved methodologies of obtention are needed (Winbourne et al. 2018) to refine the present estimations of symbiotic N2 fixation in tropical forests.
Higher N fixation in young stands but also important in old stands of the dry forests
The young stands of the subhumid and semiarid high-fertility forests also had higher N stocks than the old stands, but the fixed amounts were not as high as in the coastal forest because of the low leaf biomass (Figs. 2 and 3), probably caused by water limitation (Freitas et al. 2015; Oliveira et al. 2023). The 15N signals indicate systems with high N losses and the soil N concentrations are the lowest (Table 2), which indicates that N fixation may provide an ecological advantage (Aranibar et al. 2004). The advantage seems to be higher in the young stands but apparently persists, although to a lower degree, in the old stands. Both young and old stands had high proportions of their leaf biomass and N content belonging to nodulating species, and higher in the young forest stands. Other reports also point to the dominance of potentially nodulating species in regenerating caatinga forests, but the plants in the young forests were not effectively fixing in some of these forests (Freitas et al. 2010; Silva et al. 2017; Souza et al. 2012).
Absence of N fixation in many plants of most of the potentially fixing species
Many plants of potentially nodulating species were not effectively fixing N in all forests, even in the young stand of the semiarid high-fertility forest. This pattern has been observed in other areas in the same region (Silva et al. 2017) and in other vegetation types elsewhere (Reis et al. 2020; Sylla et al. 2002). The causes are not yet known, but are likely related to the physiology of each specific individual, since diazotrophic bacteria are present and establish effective symbiosis with nearby plants. Previous N accumulation above demand or low photosynthate production due to shading are possible causes (Dickson, 1989; Millard and Grelet 2010).
Conclusions
The ecological advantage of fixation of some legume species in the initial forest successional stages is indicated by the higher amounts of fixed N in leaf biomass in the young than in the old stands in three of the five forest types (humid coastal, subhumid and semiarid high fertility forests). This advantage was higher in the young stands in the subhumid and semiarid high fertility forests, but seemed to persist to a lower degree in the old stands. The advantage of N fixation is lost when the forest grows on nutrient-poor soils, such as those with low P availability. The ecological advantage and the favorable growth conditions may result in high absolute accumulation of fixed N in the leaves of young tropical forests, as in the studied Brazilian coastal forest (15.6 kg ha−1). This amount of N, only measured in the leaves, indicates high total symbiotic N fixation in these humid tropical forests.
Considering that almost all determinations of fixed N in tropical forests using the isotope technique were done sampling only the leaves, which are only a small proportion of the plant biomass, determinations in flowers, roots and mainly stems are badly needed. The causes of fixation absence in some plants of almost all potentially fixing species must continue to be investigated.
References
Amorim IL, Sampaio EVSB, Araujo EL (2009) Fenologia de espécies lenhosas da caatinga do Seridó. RN Árvore 33:491–499. https://doi.org/10.1590/S0100-67622009000300011
Amundson R, Austin AT, Schuur EAG, Yoo K, Matzek V, Kendall C, Uebersax A, Brenner D, Baisden WT (2003) Global patterns of the isotopic composition of soil and plant nitrogen. Glob Biogeochem Cycles 17:1031. https://doi.org/10.1029/2002GB001903
Aranibar JN, Otter L, Macko SA, Feral CJW, Epstein HE, Dowty PR, Eckardt F, Shugart HH, Swap RJ (2004) Nitrogen cycling in the soil-plant system along a precipitation gradient in the Kalahari sands. Glob Chang Biol 10:359–373. https://doi.org/10.1046/j.1529-8817.2003.00698.x
Batterman AS, Hedin LO, van Breugel M, Ransijn J, Craven DJ, Hall JS (2013) Key role of symbiotic dinitrogen fixation in tropical forest secondary succession. Nature 502:224–229. https://doi.org/10.1038/nature12525
Binkley D, Senock R, Cromack K Jr (2003) Phosphorus limitation on nitrogen fixation by Facaltaria seedlings. For Ecol Manag 186:171–176. https://doi.org/10.1016/S0378-1127(03)00240-8
Boddey RM, Peoples MB, Palmer B, Dart P (2000) Use of the 15N natural abundance technique to quantify biological nitrogen fixation by woody perennials. Nutr Cycling Agroecosyst 57:235–270. https://doi.org/10.1071/PP01058
Brookshire ENJ, Wurzburger N, Currey B, Menge DNL, Oatham MP, Roberts C (2019) Symbiotic N fixation is sufficient to support net aboveground biomass accumulation in a humid tropical forest. Sci Rep 9: 7571. https://doi.org/10.1038/s41598-019-43962-5
Burger DM, Delitti WBC (1999) Fitomassa epigéa da mata ciliar do rio Mogi-Guaçu, Itapira – SP. Rev Bras Bot 22:429–435. https://doi.org/10.1590/S0100-84041999000300011
Chaturvedi RK, Raghubanshi AS (2016) Leaf life-span dynamics of woody species in tropical dry forests of India. Trop Plant Res 3:199–212
Chave J, Riéra B, Dubois M (2001) Estimation of biomass in a neotropical forest of French Guiana: spatial and temporal variability. J Trop Ecol 17:79–96. https://doi.org/10.1017/S0266467401001055
Coletta LD, Nardoto GB, Latansio-Aidar SR, Rocha HR, Aidar MPM, Ometto JPHB (2009) Isotopic view of vegetation and carbon and nitrogen cycles in a Cerrado ecosystem, Southeastern Brazil. Sci Agric 66:467–475. https://doi.org/10.1590/S0103-90162009000400006
Costa TL, Sampaio EVSB, Araújo EL, Silva AF, Freitas ADS (2021) Contributions of Leguminosae to young and old stands of neotropical forests under different environmental conditions. Ann for Sci 78:48. https://doi.org/10.1007/s13595-021-01065-0
Craine JM, Elmore AJ, Aidar MPM, Bustamante M, Dawson TE, Hobbie EA, Kahmen A, Mack MC, McLauchlan KK, Michelsen A, Nardoto GB, Pardo LH, Peñuelas J, Reich PB, Schuur EAG, Stock WD, Templer PH, Virginia RA, Welker JM, Wright IJ (2009) Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol 183:980–992. https://doi.org/10.1111/j.1469-8137.2009.02917.x
Craine JM, Brookshire ENJ, Cramer MD, Hasselquist NJ, Koba K, Marin-Spiotta E, Wang L (2015) Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 396:1–26. https://doi.org/10.1007/s11104-015-2542-1
Crews TE (2016) Nitrogen limitation disappears with succession in many lowland tropical rainforests – as expected. Why the persistence in temperate latitudes? New Phytol 209:891–893. https://doi.org/10.1111/nph.13818
Davies Barnard T, Friedlingstein P (2020) The global distribution of biological nitrogen fixation in terrestrial natural ecosystems. Glob Biogeochem Cycles 34. https://doi.org/10.1029/2019GB006387
Dickson RE (1986) Carbon and nitrogen allocation in trees. Ann Sci for 46:631–647
Eshetu Z, Högberg P (2000) Effects of land use on 15N natural abundance of soils in Ethiopian highlands. Plant Soil 222:109–117
Faye A, Sall S, Chotte JL, Lesueur D (2007) Soil bio-functioning under Acacia nilotica var. tomentosa protected forest along the Senegal River. Nutr Cycling Agroecosyst 79:35–44
Freitas ADS, Sampaio EVSB, Santos CERS, Fernandes AR (2010) Biological nitrogen fixation in tree legumes of the Brazilian semi-arid caatinga. J Arid Environ 74:344–349. https://doi.org/10.1016/j.jaridenv.2009.09.018
Freitas ADS, Sampaio EVSB, Ramos APS, Barbosa MRV, Lyra RP, Araújo EL (2015) Nitrogen isotopic patterns in tropical forests along a rainfall gradient in Northeast Brazil. Plant Soil 391:109–122. https://doi.org/10.1007/s11104-015-2417-5
Gathumbi SM, Cadisch G, Giller KE (2002) 15N natural abundance as a tool for assessing N2-fixation of herbaceous, shrub and tree legumes in improved fallows. Soil Biol Biochem 34:1059-1071. https://doi.org/10.1016/S0038-0717(02)00038-X
Gehring C, Vlek PLG, Souza LAG, Denich M (2005) Biological nitrogen fixation in secondary regrowth and mature rainforest of central Amazonia. Agric Ecosyst Environ 111:237–252. https://doi.org/10.1016/j.agee.2005.06.009
Gei M, Rozendaal DMA, Poorter L et al (2018) Legume abundance along successional and rainfall gradients in Neotropical forests. Nat Ecol Evol 2:1104–1111. https://doi.org/10.1038/s41559-018-0559-6
Handley LL, Austin AT, Robinson D, Scrimgeour CM, Raven JA, Heaton THE, Schmidt S, Stewart GR (1999) The 15N natural abundance (d15N) of ecosystem samples reflects measures of water availability. Aust J Plant Physiol 26:185–199. https://doi.org/10.1071/PP98146
Hedin LO, Brookshire ENJ, Menge DNL, Barron AR (2009) The nitrogen paradox in tropical forest ecosystems. Annu Rev Ecol Syst 40:613–635. https://doi.org/10.1146/annurev.ecolsys.37.091305.110246
LPWG The Legume Phylogeny Working Group (2017) A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny. Taxon 66:44–77. https://doi.org/10.12705/661.3
Lai HR, Hall JS, Batterman SA, Turner BL, van Breugel M (2018) Nitrogen fixer abundance has no effect on biomass recovery during tropical secondary forest succession. J Ecol 106:1415–1427. https://doi.org/10.1111/1365-2745.12979
Millard P, Grelet GA (2010) Nitrogen storage and remobilization by trees: ecophysiological relevance in a changing world. Tree Physiol 30:1083–1095. https://doi.org/10.1093/treephys/tpq042
Monteiro HA, Lima ALA, Lins-e-Silva ACB, Silva MAM, Rodal MJN (2017) Influence of functional traits on the demographic rates of 47 arboreal species in a tropical forest in northeastern Brazil. J Environ Anal Progr 2:23–35. https://doi.org/10.24221/jeap.2.1.2017.1023.23-35
Moreira JCF, Brum M, Almeida LC, Barrera-Berdugo S, Souza AA, Camargo PB, Oliveira RS, Alves LF, Rosado BHP, Lambais MR (2021) Asymbiotic nitrogen fixation in the phyllosphere of the Amazon forest: changing nitrogen cycle paradigms. Sci Total Environ 773:145066. https://doi.org/10.1016/j.scitotenv.2021.145066
Nardoto GB, Quesada CA, Patiño S et al (2014) Basin-wide variations in Amazon forest nitrogen-cycling characteristics as inferred from plant and soil 15N:14N measurements. Plant Ecol Divers. https://doi.org/10.1080/17550874.2013.807524
Oliveira CC, Martins FR, Souza BC, Sampaio EVSB, Loiola MIB, Soares MAA (2023) Resilience and successional trends of woody vegetation in seasonally dry tropical forests. Forestry 96:1–14. https://doi.org/10.1093/forestry/cpad010
R Core Team (2020) A language and environment for statistical computing. R Foundation for Statistical Computing Vienna, Austria. http://www.R-project.org
Reis CRG, Pacheco FS, Reed SC, Tejada G, Nardoto GB, Forti MC, Ometto JP (2020) Biological nitrogen fixation across major biomes in Latin America: patterns and global change effects. Sci Total Environ. https://doi.org/10.1016/j.scitotenv.2020.140998
Roggy JC, Prévost MF, Garbaye J, Domenach AM (1999) Nitrogen cycling in the tropical rain forest of French Guiana: comparison of two sites with contrasting soil types using δ15N. J Trop Ecol 15:1–22. https://doi.org/10.1017/S0266467499000644
Salcedo IH, Tiessen H, Sampaio EVSB (1997) Nutrient availability in soil samples from shifting cultivation sites in semiarid NE Brazil. Agr Ecosyst Environ 65:177–186. https://doi.org/10.1016/S0167-8809(97)00073-X
Sampaio EVSB, Silva GC (2005) Biomass equations for Brazilian semiarid caatinga plants. Acta Bot Brasil 19:935–943. https://doi.org/10.1590/S0102-33062005000400028
Sampaio EVSB (2010) Caracterização do bioma caatinga: características e potencialidades. In: Gariglio MA, Gariglio MA, Sampaio EVSB, Cestaro LA, Kageyama PY (orgs) Uso sustentável e conservação dos recursos florestais da caatinga. Serviço florestal brasileiro, Brasília. pp 29–48
Selaya GN, Oomen RJ, Netten JJC, Werger MJA, Anten NPR (2008) Biomass allocation and leaf life span in relation to light interception by tropical forest plants during the first years of secondary succession. J Ecol 96:1211–1221. https://doi.org/10.1111/j.1365-2745.2008.01441.x
Shearer G, Kohl DH (1989) Estimates of N2 fixation in ecosystems: the need and basis of the 15N abundance method. In: Rundel PW, Ehleringer JR, Nagy KA (eds) Stable isotopes in ecological research. Springer-Verlag, New York, pp 342–347
Silva GC, Sampaio EVSB (2008) Biomassas de partes aéreas em plantas da caatinga. Rev Árvore 32:567–575. https://doi.org/10.1590/S0100-67622008000300017
Silva AF, Freitas ADS, Costa TL, Fernandes Júnior PI, Martins LMV, Santos CERS, Menezes KAS, Sampaio EVSB (2017) Biological nitrogen fixation in tropical dry forests with different legume diversity and abundance. Nutr Cycling Agroecosyst 107:321–334. https://doi.org/10.1007/s10705-017-9834-1
Souza LQ, Freitas ADS, Sampaio EVSB, Moura PM, Menezes RSC (2012) How much nitrogen is fixed by biological symbiosis in tropical dry forest? 1. Trees and shrubs. Nutr Cycling Agroecosyst 94:171–179. https://doi.org/10.1007/s10705-012-9531-z
Sullivan BW, Smitha WK, Townsend AR, Nasto MK, Reed SC, Chazdon RL, Cleveland CC (2014) Patially robust estimates of biological nitrogen (N) fixation imply substantial human alteration of the tropical N cycle. PNAS 111:8101–8106. https://doi.org/10.1073/pnas.1320646111
Swap RJ, Aranibar JN, Dowty PR, Gilhooly WP III, Macko SA (2004) Natural abundance of 13C and 15N in C3 and C4 vegetation of southern Africa: patterns and implications. Glob Chang Biol 10:350–358. https://doi.org/10.1046/j.1529-8817.2003.00702.x
Sylla SN, Ndoye I, Gueye M, Ba AT, Dreyfus B (2002) Estimates of biological nitrogen fixation by Pterocarpus lucens in a semi-arid natural forest park in Senegal using 15N natural abundance method. Afr J Biotechnol 1:50–56. https://doi.org/10.5897/AJB2002.000-009
Taylor BN, Chazdon RL, Menge DNL (2019) Successional dynamics of nitrogen fixation and forest growth in regenerating Costa Rican rainforests. Ecology. https://doi.org/10.1002/ecy.2637
Viani RAG, Rodrigues RR, Dawson TE, Oliveira RS (2011) Functional differences between woodland savannas and seasonally dry forests from south-eastern Brazil: evidence from 15N natural abundance studies. Austral Ecol 36:974–982. https://doi.org/10.1111/j.1442-9993.2010.02233.x
Vitousek PM, Menge DNL, Reed SC, Cleveland CC (2013) Biological nitrogen fixation: rates, patterns and ecological controls in terrestrial ecosystems. Philos Trans R Soc 368:1–9. https://doi.org/10.1098/rstb.2013.0119
Winbourne JB, Feng A, Reynolds L, Piotto D, Hastings MG, Porder S (2018) Nitrogen cycling during secondary succession in Atlantic Forest of Bahia, Brazil. Nature 8:1377. https://doi.org/10.1038/s41598-018-19403-0
Wong MY, Neill C, Marino R, Silverio DV, Brando PM, Howarth RW (2020) Biological nitrogen fixation does not replace nitrogen losses after forest fires in the southeastern Amazon. Ecosystems 23:1037–1055. https://doi.org/10.1007/s10021-019-00453-y
Acknowledgements
The authors thank the Brazilian Council for Scientific and Technological Development (CNPq; Projetos Universal 2018, Grant Number 426655/2018-4 and 409519/2018-9), This work is part of the National Observatory of Water and Carbon Dynamics in the Caatinga Biome—NOWCDCB, supported by the Fundação de Amparo à Ciência e Tecnologia de Pernambuco (FACEPE; APQ-0498-3.07/17 ONDACBC), CNPq (grants: 441305/2017-2; 465764/2014-2; 304947/2022-9), and CAPES (grants: 88887.136369/2017-00).
Funding
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the scholarship of the first author and the Brazilian Council for Scientific and Technological Development (CNPq; Projetos Universal 2018, Grant Number 426655/2018-4 and 409519/2018-9). This work is part of the National Observatory of Water and Carbon Dynamics in the Caatinga Biome—NOWCDCB, supported by the Fundação de Amparo à Ciência e Tecnologia de Pernambuco (FACEPE; APQ-0498-3.07/17 ONDACBC), CNPq (Grants: 441305/2017-2; 465764/2014-2), and CAPES (Grants: 88887.136369/2017-00).
Author information
Authors and Affiliations
Contributions
TLC (doctorate student) did the field sampling, analyzed and interpreted the data and wrote the article. EVSBS and ADSF (advisers) designed the project, chose the sampling sites and helped with data analysis and writing of the article. PBC helped with data analysis of 15N. ELA (member of the advisory committee) participated in discussing and writing the article. RMBS helped with the statistical analyses.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Costa, T.L., Sampaio, E.V.S.B., Freitas, A.D.S. et al. Biological nitrogen fixation in young and old tropical forests under five contrasting edaphoclimatic conditions. Nutr Cycl Agroecosyst 128, 183–198 (2024). https://doi.org/10.1007/s10705-024-10345-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-024-10345-w