
Abstract
Background and aims
The interpretation of ecosystem nitrogen isotopic patterns is subject to controversies since there are few surveys of signals along climate gradients within the same region. We determined the effects of a rainfall gradient, under high temperatures, on the soil and plant δ15N signals in Northeast Brazil.
Methods
Nitrogen, carbon and δ15N signals were determined from coastal perennial to subhumid and semiarid deciduous forests to savanna – deciduous forest transition.
Results
Soil C and N concentrations were linearly related and both related quadratically with mean annual rainfall while δ15N concentrations decreased with higher rainfall (16 to 4‰). Plant δ15N was 1–3‰ lower than soil δ15N (Δδ15N) in all areas. Only in the deciduous forests, leaves of target species had significantly lower 15 N signals than those of non-fixing species, allowing estimation of fixation.
Conclusions
In the perennial forests the low signals indicated that the 15 N natural abundance method was not suitable to estimate fixation, while in the savanna – deciduous forest transition, high signals indicated that the legume species were not fixing. The smaller Δδ15N than those reported in cooler areas and the absence of rainfall effect indicates that, when high, temperature overrides the rainfall effect.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The biogeochemical cycle of nitrogen in land ecosystems involves several isotopic fractionation processes whose balance determines the δ15N signals of their soil and plant components (Robinson 2001; Hedin et al. 2009). Inputs from symbiotic and free-living biological fixation have δ15N signals close to 0‰ (Högberg 1997; Robinson 2001), while atmospheric depositions have a wide range, but usually positive, δ15N signals (Agnihotri et al. 2011). Generally biological fixation prevails over deposition (Cleveland et al. 1999). Losses, from volatilization and leaching, have depleted δ15N signals leaving the soil with positive signals (Shearer and Kohl 1986; Högberg 1997; Handley et al. 1999; Robinson 2001). Plants have more negative signals than soil and the difference depends on climatic conditions (Amundson et al. 2003) and on the preferential absorption of NO3 − or NH4 + (Kahmen et al. 2008). Thus, promising applications of the signals are the possibility to determine N fixation and to estimate soil – plant system N dynamics (Craine et al. 2009).
The patterns revealed by several studies are that plant and soil δ15N signals tend to decrease towards zero as the annual rainfall increases (Austin and Vitousek 1988; Handley et al. 1999; Amundson et al. 2003; Nardoto et al. 2008; Craine et al. 2009) and the signals are higher in tropical than in temperate forests (Martinelli et al. 1999; Amundson et al. 2003). The differences of plant and soil signals also decrease with increasing temperature and with decreasing rainfall and latitude (Amundson et al. 2003). This occurs because in more humid places the δ15N results mainly from the interaction of the internal C cycling between dead and live organic compartments, with little N losses. This almost close cycle is interrupted in the tropics and especially in the dry tropics, where proportionally more N flows from the organic to the inorganic compartments, the latter being more subjected to losses, leading to the 15 N enrichment of soil and vegetation components (Handley and Scrimgeour 1997; Austin and Vitousek 1988; Handley et al. 1999; Martinelli et al. 1999; Aranibar et al. 2004; Swap et al. 2004; Freitas et al. 2010b). Therefore, dry places would have higher relative N availability to the plants than humid places, a hypothesis that is supported by the positive correlation between δ15N signals and total N concentrations in plants (Craine et al. 2009; Viani et al. 2011). However, relatively high N losses have also been reported in tropical rainforests which would be compensated by fixation, and fixation would occur in spite of high relative N availability to the fixing plants, creating an apparent paradox to whose solution Hedin et al. (2009) proposed a few mechanisms.
The δ15N signal also allows identifying plants with symbiotic N2 fixation if the soil signal is distinctly different from zero, which occurs in some places (Freitas et al. 2010a; Andrews et al. 2011; Souza et al. 2012) but not in others (Roggy et al. 1999a; Gehring and Vlek 2004; Nardoto et al. 2008; Nardoto et al. 2014). If the plant biomass is known, the isotopic technique also allows determining the amount of N2 biologically fixed (BFN).
In spite of these potential applications, the interpretation of patterns of differences of δ15N among places and between soil and plants is still subject to controversies. Several exceptions to the general trends have been reported (Amundson et al. 2003; Swap et al. 2004; Freitas et al. 2010b) and the relation between plant δ15N and climate is not completely clear (Craine et al. 2009). The isotopic signals of most of the tropical forests are unknown and there are few systematic surveys of signals along rainfall and soil gradients occurring at short distances, mainly at high mean annual temperatures (Craine et al. 2009; Handley et al. 1999). At mean annual temperatures above 23 °C, data on the comparison of soil and plant δ15N were absent until few years ago (Amundson et al. 2003) and no more recent report on the subject was found.
The Northeast region of Brazil, with its diversity of rainfall and soil conditions and similarity of temperatures, offers a good opportunity for to explore the relationship between climatic variables, the isotopic composition of tropical plants and soil, and the implications these relationships hold for ecosystem scale patterns of N cycling. In this article we describe the 15 N signals of plants and soils along a rainfall gradient under similar high mean annual temperature in three parallel transects about 1 latitude degree apart, in Northeast Brazil, to gain insights into the N cycling in the soil – plant system. We also evaluated the possibility to estimate biological N2 fixation from the signals of fixing and non-fixing plants. While we acknowledge the complexity of C, N and N isotope dynamics, we propose the following null hypotheses: 1) soil δ 15 N signals decrease with increasing rainfall but are high enough to allow determination of symbiotic N 2 fixation in legume plants. Plant 15 N signals in Brazilian Northeastern deciduous forests are high (δ15N of 7–9‰; Freitas et al. 2010b, Souza et al. 2012), as those of other dry places in the world (Handley et al. 1999, Aranibar et al. 2004, Viani et al. 2011), possibly because mineralization peaks earlier than plant absorption in the onset of the rainy season and mineral N is subject to losses. Signals in more humid forests of Northeast are not available but some tropical rainforests have signals higher than temperate forests and in the range of 4 to 6 ‰ (Martinelli et al. 1999, Roggy et al. 1999b, Nardoto et al. 2014), sufficiently higher than the signal of biologically fixed N and indicating that the N cycle is not completely close; 2) since the soil signals are more positive with higher relative N losses they become enriched in the heavier isotope with lower total C:N ratios. N losses are higher when soil organic matter has mineralized beyond the point of N immobilization and the C:N ratio decreases towards the limit of composition of its chemical compounds (about 10:1), justifying the negative relationship of 15 N signals and C:N ratios described by Amundson et al. (2003) and by Stevenson et al. (2010); 3) the differences of plant and soil δ 15 N (Δδ 15 N) in the hot and dry areas of Northeast are lower than those in dry and colder regions of the world but these differences increase in Northeast areas of higher rainfall. The patterns of decreasing Δδ15N with higher temperature and higher rainfall, in a global scale, were reviewed by Amundson et al. (2003) and are probably linked to differences in the chemical form of the N absorbed by the plants; and 4) symbiotic fixation is higher in areas of high soil C:N ratio, since fixation can be inhibited by high N availability and under this condition the ecological advantage of fixing plants is lost (Menge and Hedin 2009).
Material and methods
Study sites
The Brazilian Northeast region has an Atlantic coastal line roughly following a north – south direction. Annual rainfall is abundant in the coastal area (>1800 mm) and decreases westward to a semiarid core area, with low and erratic rainfall (about 500 mm). West of this core area, rainfall increases again to 1000–1500 mm, more regularly distributed in a 5–6 months rainy season. Interspersed in the semiarid area there are a few mountains (about 1000 m a.s.l.), where rainfall is higher than in the surrounding area. Soils in the coastal zone are deep, low fertility Latosols in the Brazilian classification (Embrapa 2006) or Ferrasols or Oxisosl in the FAO and American classifications, respectively; in the semiarid zone are shallow, high fertility Luvisols (Luvisols or Alfisols); and in the western portion are similar to those of the coastal zone. The coastal area was covered by tropical rain forest, a few remnants of which still dot the landscape. Half of the semiarid area is covered by a low dense shrub and tree vegetation, locally called caatinga. Few fragments remain of the semideciduous tall forest that covered the mountain tops in the semi-arid area. The western portion vegetation is a mixture of caatinga and savanna (cerrado) vegetation.
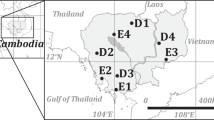
Fourteen areas with native vegetation without any major disturbance were selected following roughly three parallel east – west transects (see Fig. 1 for site locations), along the states of Paraíba (PB), Pernambuco (PE) and Alagoas (AL). The areas cover a large variation of rainfall (from 400 to more than 2000 mm y−1) and water availability, conditioned by different rainfall distribution and evapotranspiration rates along the year, and a short variation of temperatures (23 to 26 °C). This variation translates into six vegetation types (Table 1): 1) coastal perennial rainforest, in João Pessoa (PB), Recife (PE) and Rio Largo (AL) municipalities; 2) coastal semideciduous forest in Aliança (PE) and São Vicente (PE) municipalities; 3) subhumid deciduous forest (Agreste caatinga), in Caruaru (PE) and Remígio (PB); 4) semiarid deciduous forest (Sertão caatinga), in Santa Teresinha (PB), Serra Talhada (PE) and Pão de Açúcar (AL) municipalities; 5) semideciduous montane forest in Areia (PB), Caruaru (PE) and Mata Grande (AL); and 6) savanna – deciduous forest transition in Araripina (PE).

Map of the studied areas in Paraiba (PB), Pernambuco (PE) and Alagoas (AL) states, Brazil
Soil and plant sampling and analysis
In each site, five to six circular plots of about 20 m radius each and at least 50 m apart were marked in native mature vegetation patches. In each plot, one plant of the species previously selected was sampled. The selected species included all the legumes in the plot and also those non-legume species with the most abundant and largest plants, based on previous phytosociological studies. In Araripina, legume plants of species of known nodulating potential were also sampled in an experimental site adjacent to the native vegetation site. One composite sample of 10 to 20 fully expanded healthy leaves was collected from each plant, at about the same canopy height (1.5 to 2 m). Sampling was done in the peak of the rainy season in all the semideciduous and deciduous vegetation but in different seasons in the rainforests, always avoiding the period of leaf flushing. Part of the data for the subhumid deciduous forest, in Caruaru (PE) and Remígio (PB), and for the semiarid deciduous forest, in Santa Teresinha (PB) and Serra Talhada (PE) was already published by Freitas et al. (2010a, b)).
The plants were classified as non-legumes, non-nodulating legumes and target species. Non-nodulation potential was based on Allen and Allen (1981), Faria et al. (1984, 1987, 1989, 1994, 2010), Faria and Lima (1998), Sprent (2009) and Freitas et al. (2010a)) and these non nodulating legumes and the non-legumes were considered as reference species, to estimate symbiotic N2 fixation of the target species. Target species were all those with a known nodulation potential and also all legume species without complete botanical identification or for which the nodulating potential was unknown. These later species could not be safely considered as reference species and their δ15N values were compared to the values of the reference species. In case their δ15N values were significantly and more than 2 ‰ lower (Högberg 1997) they could be considered as potentially fixing species. In case their δ15N values were not significantly different from the values of the reference species, they could either be non-nodulating species or be potentially nodulating but not fixing sufficiently N to lower the δ15N signals.
Soil samples were collected from the top 20 cm layer of each plot, air-dried, sieved and cleaned of any plant debris. Those of the coastal semideciduous forest in Aliança, PE, were lost. Subsamples were ground to finer powder. The plant samples were dried and also ground to fine powder. A sub-sample of leaf or soil was placed in a capsule and loaded into a ThermoQuest-Finnigan Delta Plus isotope ratio mass spectrometer (Finnigan-MAT; CA, USA) interfaced with an Elemental Analyzer (Carlo Erba model 1110; Milan, Italy) at the Laboratory of Isotope Ecology (CENA-USP, Brazil) to obtain the nitrogen isotope ratio and the total nitrogen content of these samples. Stable isotope ratios of nitrogen were measured relative to internationally recognized standards. Internal reference materials (atropine, yeast and soil standard no. 502–308 from LECO Corporation) were included in every analytical run. The concentrations of 15 N were expressed in δ units in relation to the international standard (atmospheric N2), based on the equation δ = (Rsample/Rstandard – 1) × 1000.
The difference between leaf and soil δ15N, designated here as Δδ15N (Amundson et al. 2003), was calculated for each plant. Foliar Δδ15N values represent the 15 N depletion of a plant leaf compared to the soil δ15N background (Kahmen et al. 2008).
Data analysis and statistics
Soil organic C and total N values were regressed against each other and against rainfall and soil δ15N signature were regressed against soil C:N ratio and leaf δ15N signature of reference species. Leaf δ15N signature of reference species were regressed against rainfall and against the coefficient of variation of the δ15N signals of all reference plants in the site. Only the values of reference species were used to avoid the interference of atmospheric N2 fixation of the nodulating legume species. All regressions were tested for homocedasticity.
Soil data (C, N and δ15N) were submitted to analysis of variance and the averages of the different sites were compared using the Tukey test at the 0.05 probability level. Plant data of the different sites did not have homogeneous variation and were not submitted to analysis of variance. Within each site, δ15N values of target legume species were compared with the average δ15N values of reference species, using a t test at the 0.05 probability level. Only the comparison of the reference average value with average value of the target species with the lower δ15N average is presented in the text, to identify if any species in the site was fixing atmospheric N2.
Results
Total carbon and nitrogen concentrations in the superficial soil layer (0 to 20 cm) varied significantly among areas (Table 2) and related quadratically with mean annual rainfall (Fig. 2), the highest values in areas with intermediate rainfall (1000 to 1500 mm). C and N were linearly related (Fig. 3), and the slope of the equation translates into a C:N ratio of 14.9:1. The abundance of the 15 N isotope (δ15N) also varied significantly among areas (Table 2) and it was not significantly related to total C or N concentrations but it was negatively related to the C:N ratio, if the data from the savanna caatinga transition area is excluded (Fig. 4).

Soil organic carbon (SOC) and total nitrogen (STN) concentrations as a function of mean annual rainfall in different vegetation types in Northeast Brazil

Soil N as a function of soil C in different vegetation types in Northeast Brazil

Soil 15 N abundance as a function of soil C:N ratio in different vegetation types in Northeast Brazil (Araripina data were not included in the regression)
Concentrations of 15 N of reference species varied considerably among areas (Table 3), in general areas with higher rainfall (>1000 mm) having lower values than areas with lower rainfall (<1000 mm), with a negative significant relationship (Fig. 5). Leaves of reference species and soil had significantly related δ15N signals (Fig. 6) and leaves were depleted of 15 N in relation to the soil (Δδ15N), except those of one montane forest (Fig. 7). In spite of the good correlation of the averages, the differences of individual plant value to that of the soil in the same area varied considerably, typically in a range of 1 to 2 δ15N‰ (Fig. 7). There were no significant effects of rainfall, soil C, N and C:N ratio, and latitude on Δδ15N values of the different areas.

Nitrogen-15 abundance (δ15N) of leaves of non-N2-fixing species as a function of rainfall in different vegetation types in Northeast Brazil

Nitrogen-15 abundance (δ15N) of leaves of non-fixing species as a function of nitrogen-15 abundance in the soil in different vegetation types in Northeast Brazil

Difference between leaf and soil δ15N (Δδ15N) in different vegetation types in Northeast Brazil. PB Paraiba state (transect 1); PE Pernambuco state (transect 2); AL Alagoas state (transect 3)
Only in the deciduous forests and in one montane forest, leaves of target species had significantly lower 15 N signals than those of reference species (Table 3. Additional data in Table ESM 1, presented as electronic supplementary material). Fixation of atmospheric N in these deciduous forests accounted for percentages of the N in the fixing legume species generally above 50 % (Freitas et al. 2010a). In the montane forest, the signals of two target species were significantly lower than those of the reference species but the difference was lower than 2 ‰. In all other sites, the signals of target species and reference species were not significantly different.
In the coastal rainforests the signals of both species groups were 1 to 3 δ15N‰ units above atmosphere, indicating the natural abundance method was not suitable to estimate fixation. In the savanna caatinga transition, both signals were high, indicating absence of fixation of all target species.
Leaf N concentrations of non-legume species varied within a narrow range among areas and were lower than concentrations of legume species but the differences were only statistically significant in the montane and in two deciduous forests (Table 4).
Discussion
Soil C (7 to 46 g kg−1) and N concentrations (0.7 to 3.3 g kg−1) were relatively low (Table 2) but within the range usually reported for the surface horizons of tropical soils (Marin-Spiotta et al. 2009; Kirkby et al. 2011; Marín-Spiotta and Sharma 2013; Xu et al. 2013). The highest values were found in the coastal semideciduous and montane forests, located in areas with intermediate annual rainfall (1000 to 1500 mm), indicating that the balance of organic matter deposition and mineralization favored accumulation to a higher degree than in the other areas. The size of trees and consequently the aboveground biomass in these coastal semideciduous and montane forests are similar to or somewhat lower than in the coastal rainforests and higher than in the subhumid and semiarid deciduous forest and in the savanna – caatinga transition vegetation (Sampaio 1996). Assuming that the organic deposition is proportional to the aboveground biomass (Batjes 1996; Marín-Spiotta and Sharma 2013), the litter mineralization rates in these drier semideciduous and montane forests must be lower than in the costal rainforests. In the even drier subhumid and semiarid deciduous forest and in the savanna – deciduous forest transition vegetation, the lower soil C and N accumulations probably result from lower litter depositions. The C:N ratios had an average close to 15 and a lower relative variation than the variation of the C and N concentrations. The range and the average are similar to those reported for soils in the world (Batjes 1996) and indicate that the C and N cycles in these soils are approximately equilibrated.
The accumulation of 15 N has been appointed as a measure of the openness of the N cycle (Austin and Vitousek 1988; Handley et al. 1999; Roggy et al. 1999b; Eshetu and Högberg 2000; Aranibar et al. 2004; Swap et al. 2004; Ometto et al. 2006; Coletta et al. 2009; Freitas et al. 2010b; Viani et al. 2011), a more open cycle having higher losses of the lighter isotope (14 N). The negative relation of the C:N ratio and the soil δ15N values (Fig. 4) confirms our second hypothesis and suggests that soils with lower C:N ratios would have more open N cycles, that is, they would have greater N losses. However, considering the lower C:N ratios in these soils, losses of C are even proportionally higher than those of N. Negative relation of total C:N ratio and soil δ15N values was reported in the comprehensive review of Amundson et al. (2003) and by Stevenson et al. (2010). Therefore, the subhumid and semiarid deciduous forests, with their highest δ15N values (excluding the savanna – caatinga transition vegetation), would have the greatest relative N and C losses. Losses have not been determined in any of these areas but temperate forests with higher C:N ratios had lower nitrate losses through leaching (Gundersen et al. 1998). The losses may be related to seasonal variations in litter deposition and decomposition. Deposition in the deciduous forests is concentrated after the end of the rainy period and decomposition may start with photodegradation while the vegetation is denuded of foliage (Parton et al. 2007) and must accelerate with the beginning of the next rainy season, with abundant water availability. In the other less seasonal vegetations, particularly the rainforests, the almost constant litter deposition (Sampaio et al. 1993) with high C:N ratio would result in an equilibrium with higher soil C:N ratio.
The case of the savanna – caatinga transition vegetation, with both high C:N ratio and high δ15N values, requires a different explanation. The extremely low soil P availability (Salcedo et al. 1997) may be implicated in higher relative N losses than C losses. The vegetation is short and quite open with many bare spots and probably has a large root:shoot ratio, as other savanna (Mokany et al. 2006) and caatinga (Costa et al. 2014) areas. Therefore the biomass input to the soil is probably low and composed of hard and leathery leaves, which may take longer to decompose than in other areas, and a large part of underground deposition where the biomass is not subjected to photodegradation (Parton et al. 2007). On the other hand, they have high N concentrations indicating that N is not a limiting nutrient, the mineral pool may not be readily depleted and some mineral N may be lost through leaching. The openness of the system and the absence of symbiotic fixation (discussed below), which would incorporate N with δ15N values close to zero, turn the N balance towards high relative N losses and, therefore, high δ15N signals.
The pattern of leaf 15 N of reference species followed closely that of soil 15 N, as expected considering that the soil is their sole source of N. In all cases except one, the plant signals are impoverished in 15 N in relation to the soil where they grow, with an average Δδ15N signal of −2.34‰, corresponding to the absorption of mineralized forms of N which are depleted in the heavier isotope. The values are lower than all those listed by Amundson et al. (2003) for areas with lower mean annual temperatures, confirming the trend they reported of lower values with increasing temperatures, as also stated in our third hypothesis. However, contrary to their finding and our hypothesis, differences in rainfall had no significant effect on the Δδ15N values. Apparently, when the temperature is high its influence overrides that of the water availability. Even in the colder sites reviewed by Amundson et al. (2003), the influence of temperature was stronger than that of rainfall. There was no effect of latitude on the Δδ15N values (or in any other measured variable) but the 1° difference in our sites is too small and the latitude effect is probably mediated through temperature variation, which was not relevant among our sites. In all Brazilian Northeastern forests more N could be absorbed as NO3 −, with its lower depletion factor, than as NH4 +, as a result of faster soil organic matter turnover (Tiessen et al. 1994) and nitrification due to higher temperatures but the subject is still controversial and may involve enrichment of mineral N due to denitrification and less absorption through mycorrhizal fungi/plant interactions (Amundson et al. 2003; Kahmen et al. 2008). For all these processes there is no available information for forests in Northeast Brazil.
The low δ15N signals in the more humid coastal and montane forests and the high δ15N signals in the subhumid and semiarid vegetations partially confirm our first hypothesis of decreasing signals with increasing rainfall. The pattern of higher δ15N signals as water availability decreases has not been described within many regions but many dry area vegetations in Africa have also high plant δ15N signals (Handley et al. 1999; Aranibar et al. 2004). The savanna – deciduous forest transition δ15N signal is also much higher than those found in savanna areas in Central Brazil (Bustamante et al. 2004; Coletta et al. 2009; Viani et al. 2011) where rainfall is also much higher. In contrast, the signals of the coastal rainforests are lower than those reported for Amazonian forests (Roggy et al. 1999b; Ometto et al. 2006) although even in these last forests low values have also been reported (Nardoto et al. 2014). Therefore, the signals of our rainforest do not support the statement that tropical forests have higher signals than temperate forests (Martinelli et al. 1999) and they were so few units above 0 ‰ that left little margin to detect significant decreases in the signals of fixing species. Therefore, the second part of our first hypothesis was not confirmed and the technique of natural abundance would only be adequate to measure fixation in our rainforests if the plants were acquiring very high percentages of their N through fixation.
Certainly, water and latitude are not the only drivers of the N cycle and plant and soil 15 N signals are not only influenced by lower losses of the heavy isotope but also by fixation either by plant symbiosis or by soil free-living microorganisms, with their influx of 0 ‰ δ15N (Högberg 1997; Robinson 2001), and by atmospheric deposition, usually with positive δ15N values (Agnihotri et al. 2011). Therefore, areas with low δ15N values may have closed cycles with fast immobilization or absorption of mineralized N or, alternatively, high losses compensated by high fixation and low deposition. The uncertainty about fixation in our rainforests prevents determining which of the two patterns they follow. In some of these forests (Sampaio 1996), the low number of potentially fixing legume plants would limit the fixation input even if the plants fixed high proportions of their N but not all of the forests had the floristic composition determined. The C and N cycles are also affected by the availability of other nutrients, especially P (Brenner et al. 2001). The low soil P availability in the savanna – deciduous forest transition area (Salcedo et al. 1997) could result in high N losses and explain part of its high δ15N signal.
The δ15N signals of target species were only significantly lower than averages of reference species and more than 2 ‰ δ15N in the four deciduous forests which were the subject of a previous publication (Freitas et al. 2010b). Since the value of 2 ‰ δ15N was established by Högberg (1997) as a safe limit to estimate biological fixation, we only estimated fixation in these forests. In these forests, symbiotic fixation accounted for high proportions of the plant N but the amounts fixed were low because of the low densities of the populations of the fixing legume species. The proportions of fixed N were usually above 50 % of the total leaf N but the annual contribution of fixed N was below 10 kg ha−1 (Freitas et al. 2010a). In one montane forest the δ15N averages of two species were significantly lower than the reference average (Table ESM 1, presented as electronic supplementary material) but the differences were lower than 2 ‰. These two species, Bowdichia virgilioides Kunth and Albizia polycephala (Benth.) Killip, are known to nodulate and could be fixing small proportions of their N.
In all other areas there was no apparent contribution of symbiotically fixed N to the target species. In the rainforests the lack of significant differences resulted from signals of reference species with low values and high variances. In fact, the variances of signals in all areas were not much different, with standard deviations from 0.98 to 2.30‰ (Table 3), in a way that their coefficient of variation decreased with the increase in the average value (Fig. 8). The consequence is that it is more difficult to detect the decrease in signal caused by fixation in areas with low reference values. It is the case of all coastal perennial forests and one of two of the coastal semideciduous and two of the three montane forests, with signals below 4 ‰, in which the technique of natural abundance could not be used to estimate fixation. Therefore, we cannot determine if their relatively low soil and plant values originate from a large contribution of biological fixation. No data are available for fixation by soil free-living microorganisms in these forests but in other regions (Hedin et al. 2009) this fixation can be of similar magnitude as those we found in the four deciduous forests (Freitas et al. 2010a).

Coefficient of variation of leaf 15 N signals as a function of average leaf 15 N signals in different vegetation types in Northeast Brazil
In the other coastal semideciduous and one montane forests, and especially in the savanna – deciduous forest transition, the signals of the reference species were sufficiently higher than atmosphere N, theoretically allowing estimation of fixation if the signals of the fixing species were significantly lower. However, the differences between the average signals of these reference species and those of target species were too small to indicate any dilution of the 15 N coming from the soil by 15 N from the atmosphere (Table ESM 1, presented as electronic supplementary material). This is particularly clear in the savanna – caatinga transition area, where the signals of the reference species were very high, one of the highest in the literature (Handley et al. 1999; Amundson et al. 2003; Craine et al. 2009). The absence of fixation of the native legume species was confirmed by the analysis of well-known potentially fixing species from the growth experiment in the same site. The causes of this absence are not known but a similar fact was registered for regenerating caatinga in one of the semideciduous forests (Freitas et al. 2012) and has been noticed in other regions (Ndiaye and Ganry 1997; Faye et al. 2007). It is interesting to note that in the coastal semideciduous and montane forests, the signals of the target species had larger variations than those of the reference species and frequently higher absolute averages (Table 3). This could indicate not only absence of fixation but also a preference for absorption of soil NH4 + for NO3 − (Kahmen et al. 2008), but there is not enough information on the subject to reach any firm conclusion.
One consequence of the absence of fixation in the savanna – caatinga transition area and the absence or undefinition of fixation in the montane and rainforests, the areas with higher soil C:N ratios, is that our fourth hypothesis of higher fixation in areas with high C:N ratios was not validated. In fact, fixation was only determined in the deciduous forests, which had the lowest C:N ratios. As discussed above, these areas had great losses of N, reflected in their high δ15N, but even greater losses of C, reflected in the low C:N ratios.
Fixation has an influence in the 15 N signal of the soil, proportional to the amount fixed in relation to the general recycling of N in the area. Soil δ15N in the deciduous forests would be lowered by the contribution of organic matter coming from the fixing species, but the effect would be small because this contribution is small and, in fact, the soil signals in these forests are among the highest of all areas.
Conclusions
Total soil carbon and nitrogen concentrations were higher in areas with intermediate (1000 to 1500 mm) than with lower or higher rainfall, where biomass productions are low and high, respectively, indicating high losses of organic matter in the rainier forests. The abundance of the 15 N isotope (δ15N) was negatively related to the mean annual precipitation and had a large span, from some of the highest values in the world (>10 ‰) to values slightly above atmosphere abundance. Therefore, the dry deciduous forests had higher relative N losses and the rainier forests a more closed N cycling in spite of high organic matter losses. The δ15N values were also negatively correlated to the soil C:N ratio indicating higher N losses where soil organic matter had mineralized to a greater degree, the dry deciduous forests.
In general, plants were depleted of 15 N in relation to the soil (Δδ15N) and the differences were lower than those reported for colder places of the world, confirming the tendency of decreasing values in higher mean annual temperatures. Contrary to the world trend, rainfall had no significant influence on the Δδ15N values, indicating that when the temperature is high its effect overrides that of water availability. The similarity of Δδ15N in all northeastern forests suggests that they are absorbing the same forms of soil N, possibly more NO3 −.
Except in the deciduous forests, the low signals of reference plants and/or the small depletion of 15 N of target species did not allow estimation of fixation. In the rainforests the low signals of reference species indicate that the 15 N natural abundance technique is not suitable to estimate fixation. In the savanna – deciduous forest transition fixation was not occurring.
References
Agnihotri R, Mandal TK, Karapurkar SG, Naja M, Gadi R, Ahammmed YN, Kumar A, Saud T, Saxena M (2011) Stable carbon and nitrogen isotopic composition of bulk aerosols over India and northern Indian Ocean. Atm Environ 45:2828–2835. doi:10.1016/j.atmosenv.2011.03.003
Allen ON, Allen EK (1981) The Leguminosae: a source book of characteristics use and nodulation. University of Wisconsin Press, Wisconsin
Amundson R, Austin AT, Schuur EAG, Yoo K, Matzek V, Kendall C, Uebersax A, Brenner D, Baisden WT (2003) Global patterns of the isotopic composition of soil and plant nitrogen. Glob Biogeochem Cycles 17:1031. -3 1-31. - 10. doi: 10.1029/2002GB001903
Andrews M, James EK, Sprent JI, Boddey RM, Gross E, Reis FB Jr (2011) Nitrogen fixation in legumes and actinorhizal plants in natural ecosystems: values obtained using 15N natural abundance. Plant Ecol Divers 4:131–140. doi:10.1080/17550874.2011.644343
Aranibar JN, Otter L, Macko SA, Feral CJW, Epstein HE, Dowty PR, Eckardt F, Shugart HH, Swap RJ (2004) Nitrogen cycling in the soil-plant system along a precipitation gradient in the Kalahari sands. Glob Chang Biol 10:359–373. doi:10.1046/j.1529-8817.2003.00698.x
Austin AT, Vitousek PM (1988) Nutrients dynamics on a precipitation gradient in Hawai’i. Oecologia 113:519–529
Batjes NH (1996) Total carbon and nitrogen in the soils of the world. Eur J Soil Sci 47:151–163
Brenner DL, Amundson R, Baisden WT, Kendall C, Harden J (2001) Soil N and 15N variation with time in a California annual grassland ecosystem. Geochim Cosmochim Ac 65:4171–4186
Bustamante MMC, Martinelli LA, Silva LA, Camargo PB, Klink CA, Domingues TF, Santos RV (2004) 15N natural abundance in woody plants and soils of central Brazilian savannas (Cerrado). Ecol Appl 14:200–213. doi:10.1890/01-6013
Cleveland CC, Townsend AR, Schimel DS, Fisher H, Howarth RW, Hedin LO, Perakis SS, Latty EF, Von Fischer JC, Elseroad A, Wasson NF (1999) Global patterns of terrestrial biological nitrogen (N2) fixation in natural ecosystems. Global Biogeochem Cy 13:623–645
Coletta LD, Nardoto GB, Latansio-Aidar SR, Rocha HR, Aidar MPM, Ometto JPHB (2009) Isotopic view of vegetation and carbon and nitrogen cycles in a Cerrado ecosystem, Southeastern Brazil. Sci Agricola 66:467–475. doi:10.1590/S0103-90162009000400006
Costa TL, Sampaio EVSB, Sales MF, Accioly LJO, Althoff TD, Pareyn FGC, Albuquerque ERGM, Menezes RSC (2014) Root and shoot biomasses in the tropical dry forest of semi-arid Northeast Brazil. Plant Soil 314:113–123. doi:10.1007/s11104-013-2009-1
Craine JM, Elmore AJ, Aidar MPM, Bustamante M, Dawson TE, Hobbie EA, Kahmen A, Mack MC, McLauchlan KK, Michelsen A, Nardoto GB, Pardo LH, Peñuelas J, Reich PB, Schuur EAG, Stock WD, Templer PH, Virginia RA, Welker JM, Wright IJ (2009) Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol 183:980–992. doi:10.1111/j.1469-8137.2009.02917.x
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA – EMBRAPA (2006) Centro nacional de pesquisa de solos. Sistema brasileiro de classificação de solos, Rio de Janeiro
Eshetu Z, Högberg P (2000) Effects of land use on 15N natural abundance of soils in Ethiopian highlands. Plant Soil 222:109–117
Faria SM, Lima HC (1998) Additional studies of the nodulation status of legume species in Brazil. Plant Soil 200:185–192
Faria SM, Franco AA, Jesus RM, Menandro MS, Baitello JB, Mucci ESF, Dobereiner J, Sprent JI (1984) New nodulating legume trees from southeast Brazil. New Phytol 98:317–328
Faria SM, Lima HC, Franco JB, Mucci ESF, Sprent JI (1987) Nodulation of legume trees from southeast Brazil. Plant Soil 99:347–356
Faria SM, Lima HC, Carvalho AM, Gonçalves VF, Sprent JI (1994) Ocurrence of nodulation in legume species from Bahia, Minas Gerais and Espírito Santo States of Brazil. In: Sprent JI, McMey D (eds) Advances in legume systematics 5. The nitrogen factor. Royal Botanic Gardens, Kew
Faria SM, Diedhiou AG, Lima HC, Ribeiro RS, Galiana A, Castilho AF, Henriques JC (2010) Evaluating the nodulation status of leguminous species from the Amazonian forest of Brazil. J Exp Bot 61:3119–3127. doi:10.1093/jxb/erq142
Faye A, Sall S, Chotte JL, Lesueur D (2007) Soil bio-functioning under Acacia nilotica var. tomentosa protected forest along the Senegal River. Nutr Cycles Agroecosyst 79:35–44. doi:10.1007/s10705-007-9093-7
Freitas ADS, Sampaio EVSB, Menezes RSC, Tiessen H (2010a) 15N natural abundance of non-fixing woody species in the Brazilian dry forest (caatinga). Isot Environ Health Stud 46:210–218. doi:10.1080/10256016.2010.488805
Freitas ADS, Sampaio EVSB, Fernandes AR, Santos CERS (2010b) Biological nitrogen fixation in legume trees of the Brazilian caatinga. J Arid Environ 74:344–349. doi:10.1016/j.jaridenv.2009.09.018
Gehring C, Vlek PLG (2004) Limitations of the 15N natural abundance method for estimating biological nitrogen fixation in Amazonian forest legumes. Basic Appl Ecol 5:567–580. doi:10.1016/j.baae.2004.09.005
Gundersen P, Callesena I, de Vriesb W (1998) Nitrate leaching in forest ecosystems is related to forest floor C/N ratios. Environ Pollut 102:02–407
Handley LL, Scrimgeour CM (1997) Terrestrial plant ecology and 15N natural abundance: the present limits to interpretation for uncultived systems with original data from a Scottish old field. Adv Ecol Res 27:133–212
Handley LL, Austin AT, Robinson D, Scrimgeour CM, Raven JA, Heaton THE, Schmidt S, Stewart GR (1999) The 15N natural abundance (δ15N) of ecosystem samples reflects measures of water availability. Aust J Plant Physiol 26:185–199
Hedin LO, Brookshire ENJ, Menge DNL, Barron AR (2009) The nitrogen paradox in tropical forest ecosystems. Annu Rev Ecol Evol Syst 40:613–635
Högberg P (1997) 15N natural abundance in soil-plant systems. New Phytol 137:179–203
Kahmen A, Wanek W, Buchmann N (2008) Foliar δ15N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient. Oecologia 156:861–870. doi:10.1007/s00442-008-1028-8
Kirkby CA, Kirkegaard JA, Richardson AE, Wade LJ, Blanchard C, Batten G (2011) Stable soil organic matter: a comparison of C:N:P:S ratios in Australian and other world soils. Geoderma 163:197–208. doi:10.1016/j.geoderma.2011.04.010
Marín-Spiotta E, Sharma S (2013) Carbon storage in successional and plantation forest soils: a tropical analysis. Global Ecol Biogeogr 22:105–117. doi:10.1111/j.1466-8238.2012.00788.x
Marin-Spiotta E, Silver WL, Swanston SW, Ostertag R (2009) Soil organic matter dynamics during 80 years of reforestation of tropical pastures. Global Chang Biol 15:1584–1597. doi:10.1111/j.1365-2486.2008.01805.x
Martinelli LA, Piccolo MC, Townsend AR, Vitousek PM, Cuevas E, McDowell W, Robertson GP, Santos OC, Treseder K (1999) Nitrogen stable isotopic composition of leaves and soil: tropical versus temperate forests. Biogeochemistry 46:45–65
Menge DNL, Hedin LO (2009) Nitrogen fixation in different biogeochemical niches along a 120 000-year chronosequence in New Zealand. Ecology 90:2190–2201
Mokany K, Raison RJ, Prokushkin AS (2006) Critical analysis of root: shoot ratios in terrestrial biomes. Global Chang Biol 12:84–96. doi:10.1111/j.1365-2486.2005.001043.x
Nardoto GB, Ometto JPHB, Ehleringer JR, Higuchi N, Bustamante MMC, Martinelli LM (2008) Understanding the influences of spatial patterns on N availability within the Brazilian Amazon Forest. Ecosystems 11:1234–1246. doi:10.1007/s10021-008-9189-1
Nardoto GB, Quesada CA, Patiño S, Saiz G, Baker TR, Schwarz M, Schrodt F, Feldpausch TR, Domingues TF, Marimon BS, Marimon B Jr, Vieira ICG, Silveira M, Bird MI, Phillips OL, Lloyd J, Martinelli L (2014) Basinwide variations in Amazon forest nitrogen-cycling characteristics as inferred from plant and soil 15N:14N measurements. Plant Ecol Divers 7:173–187. doi:10.1080/17550874.2013.807524
Ndiaye M, Ganry F (1997) Variation in the biological N2 fixation by tree legumes in three ecological zones from the North to the south of Senegal. Arid Soil Res Rehab 11:245–254
Ometto JPHB, Ehleringer JR, Domingues TF, Berry JA, Ishida FY, Mazzi E, Higuchi N, Flanagan LB, Nardoto GB, Martinelli LA (2006) The stable carbon and nitrogen isotopic composition of vegetation in tropical forests of the Amazon Basin, Brazil. Biogeochemistry 79:251–274. doi:10.1007/s10533-006-9008-8
Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Curre W, King JY, Adair EC, Brandt LA, Hart SC, Fasth B (2007) Global scale similarities in nitrogen release patterns during long-term decomposition. Science 135:361–364
Robinson D (2001) δ15N as an integrator of the nitrogen cycle. Trends Ecol Evol 16:153–162
Roggy JC, Prévost MF, Gourbiere F, Casabianca H, Garbaye J, Domenach AM (1999a) Leaf natural 15N abundance and total N concentration as potential indicators of plant nutrition in legumes and pioneer species in a rain forest of French Guiana. Oecologia 120:171–182
Roggy JC, Prévost MF, Garbaye J, Domenach AM (1999b) Nitrogen cycling in the tropical rain forest of French Guiana: comparison of two sites with contrasting soil types using δ15N. J Trop Ecol 15:1–22
Salcedo IH, Tiessen H, Sampaio EVSB (1997) Nutrient availability in soil samples from shifting cultivation sites in semiarid NE Brazil. Agr Ecosyst Environ 65:177–186
Sampaio EVSB (1996) Fitossociologia. In: Sampaio EVSB, Mayo SJ, Barbosa MRV (eds) Pesquisa botânica nordestina: progresso e perspectivas. Sociedade Botânica do Brasil, Recife, pp 203–224
Sampaio EVSB, Dall’Olio A, Nunes KS, Lemos EEP (1993) A model of litterfall, litter layer losses and mass transfer in a humid tropical forest at Pernambuco, Brazil. J Trop Ecol 9:291–301
Shearer G, Kohl DH (1986) N2-fixation in field settings: estimations based on natural 15N abundance. Aust J Plant Physiol 13:699–756
Souza LQ, Freitas ADS, Sampaio EVSB, Moura PM, Menezes RSC (2012) How much nitrogen is fixed by biological symbiosis in tropical dry forests? 1. Trees and shrubs. Nutr Cycl Agroecosyst 94:171–179. doi:10.1007/s10705-012-9531-z
Sprent JI (2009) Legume nodulation: a global perspective. Wiley-Blackwell, Chichester
Stevenson BA, Parfitt RL, Schipper LA, Baisden WT, Mudge P (2010) Relationship between soil 15N, C/N and N losses across land uses in New Zealand. Agric Ecosyst Environ 139:736–741
Swap RJ, Aranibar JN, Dowty PR, Gilhooly WP III, Macko SA (2004) Natural abundance of 13C and 15N in C3 and C4 vegetation of southern Africa: patterns and implications. Glob Chang Biol 10:350–358. doi:10.1046/j.1529-8817.2003.00702.x
Tiessen H, Cuevas E, Chacon P (1994) The role of soil organic matter in sustaining soil fertility. Nature 371:783–785
Viani RAG, Rodrigues RR, Dawson TE, Oliveira RS (2011) Functional differences between woodland savannas and seasonally dry forests from south-eastern Brazil: evidence from15N natural abundance studies. Austral Ecol 36:974–982. doi:10.1111/j.1442-9993.2010.02233.x
Xu X, Thornton PE, Post WM (2013) A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Global Ecol Biogeogr 22:737–749. doi:10.1111/geb.12029
Aknowledgments
The authors thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; Projeto Universal 2012, Processo 472997/2012-2) and FACEPE (Processo APQ-0077-5.01/09, edital 05–2010) for the financial support. They also like to thank the technical support from Dr. Plínio B. Camargo (CENA/USP).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Elizabeth M Baggs.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(XLSX 25 kb)
Rights and permissions
About this article

Cite this article
de Freitas, A.D.S., de Sá Barretto Sampaio, E.V., de Souza Ramos, A.P. et al. Nitrogen isotopic patterns in tropical forests along a rainfall gradient in Northeast Brazil. Plant Soil 391, 109–122 (2015). https://doi.org/10.1007/s11104-015-2417-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2417-5