
Abstract
Molecular cloning, characterization, and functional analysis of follicle-stimulating hormone receptor (FSHR) in female turbot (Scophthalmus maximus) were evaluated. Results showed that the full-length FSHR cDNA was 3824 bp long and contained a 2202 bp open reading frame that encoded a mature protein of 733 amino acids (aa) and a signal peptide of 18 aa. Multiple sequence analyses showed that turbot FSHR has high homology with the corresponding genes of other teleosts and significant homology with that of Hippoglossus hippoglossus. Turbot FSHR has the typical structural architecture of glycoprotein hormone receptors consisting of a large N-terminal extracellular domain, seven transmembrane domains and short C-terminal intracellular domain. FSHR mRNA was found to be abundant in the ovaries, but deficient in eyes, intestine, brain, muscle, gills, spleen, stomach, heart and kidney. Furthermore, FSHR mRNA was found to increase gradually from pre-vitellogenesis to migratory nucleus stages, with the highest values observed during the late vitellogenesis stage of the reproductive cycle. However, FSHR mRNA was found to decrease dramatically during the atresia stage. Meanwhile, functional analysis with HEK293T cells continual expressing FSHR demonstrated that FSHR was specifically stimulated by ovine FSH, but not ovine LH. These results indicate that turbot FSHR is mainly involved in the stimulation of vitellogenesis, regulation of oocyte maturation as well as promotion of ovarian development via specific ligand binding. These findings open doors to further investigation of physiological functions of FSHR, which will be valuable for fish reproduction and broodstock management.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The development and maturation of ovarian oocytes are key physiological processes in female reproduction. Oogenesis is strictly regulated by numerous endocrine and paracrine factors, particularly gonadotropic hormones (Hurk and Zhao 2005; Lubzens et al. 2010; Chaffin and Vandevoort 2013). In teleosts, like in other vertebrates, gonadotropins mainly act on gonadal tissue by binding to specific receptors, namely follicle-stimulating hormone receptor (FSHR, formerly termed GtHR-I) and luteinizing hormone receptor (LHR, formerly termed GtH-RII) (Swanson et al. 2003; Levavi-Sivan et al. 2010). These receptors are primarily expressed on the surfaces of gonadal somatic cells, and involved in the regulation of gametogenesis via distinct intracellular signaling pathways (Menon and Menon 2012). Numerous studies have reported that fish gonadotropin receptors (GtHRs) exhibit typical characteristics of a glycoprotein hormone receptor and belong to the G protein-coupled receptors (GPCR) superfamily, which are complex transmembrane (TM) rhodopsin-like proteins characterized by seven hydrophobic helices inserted in the plasmalemma as well as intracellular and extracellular domains (Kumar et al. 2001a, b; Oba et al. 2001; Kwok et al. 2005; Rocha et al. 2007; Levavi-Sivan et al. 2010). However, ligand specificity of fish GtHRs is not as well defined as it is in mammalian species and cross-activation may occur under physiological conditions (Bogerd 2007; Levavi-Sivan et al. 2010). Two genetically distinct GtHRs have been documented in amago salmon (Oba et al. 1999a, b), African catfish (Vischer and Bogerd 2003), Atlantic salmon (Maugars and Schmitz 2006), European sea bass (Rocha et al. 2007) Atlantic halibut (Kobayashi and Andersen 2008), Atlantic cod (Mittelholzer et al. 2009), and mummichog (Ohkubo et al. 2013). In addition, molecular biological studies have shown that the two receptors have different expression profiles in repetitive and annual spawning fish during the reproductive cycle (Kumar et al. 2001a, b; Kwok et al. 2005; Kobayashi et al. 2008; Andersson et al. 2013). Thus, detailed information about the characteristics, function and regulation of GtHRs is necessary for sound understanding of potential physiological functions in fish reproductive cycle, which may be useful in controlling fish reproduction and improving broodstock management.
Turbot (Scophthalmus maximus), which has high economic value, delicious meat and rapid growth, is cultured widely in Europe and Asia. Some preliminary studies have been conducted on the nutritional requirements, hormonal induction of spawning, egg quality, and the effect of environmental conditions on female turbot during its reproductive cycle (Jones 1974; Mcevoy 1989; Suquet et al. 1995; Bromley et al. 2000; Jia et al. 2014a). Mature female turbot can spawn 8–12 times at intervals of 3–5 days during the spawning season (Mugnier et al. 2000). The follicles in the mature female turbot ovary represent different developmental stages throughout spawning season. Numerous genes were confirmed to be related to reproduction and involved in sex differentiation in turbot (Ribas et al. 2013). However, the molecular mechanisms that regulate asynchronous development of multiple batches of oocytes in turbot and the functional properties of GtHRs in the ovary during the reproductive cycle remain poorly understood. The related study has been conducted to elucidate the molecular cloning, molecular characteristics and temporal expression profiles of turbot LHR (Jia et al. 2014b). In the present study, we sought to investigate GtHRs in turbot in great detail by cloning and characterizing FSHR from turbot ovarian tissues. In addition, FSHR expression profiles and functions of turbot during the reproductive cycle were evaluated.
Materials and methods
Fish and tissue sampling
Sexually mature female turbots were obtained from Tianyuan Fisheries Co., Ltd. (Shandong Province, China). The turbots were rapidly removed after anesthetized with 100 mg/L tricaine methane sulfonate (MS-222, Sigma, St. Louis, MO, USA). Tissues samples (ovary, eye, intestine, brain, muscle, ovary, gill, spleen, liver, stomach, heart and kidney) were collected from each fish and stored in liquid nitrogen for RNA extraction. The ovaries were placed in Bouin’s solution for hematoxylin and eosin staining to identify the oocyte developmental stages according to our previous study (Jia et al. 2014b). The stages of ovarian development are as follows: pre-vitellogenesis (Prevtg), early vitellogenesis (Evtg), late vitellogenesis (Latvtg), migratory-nucleus (Mig-nucl) and atresia (Atre). Meanwhile, testis were collected from six mature male turbots and stored in liquid nitrogen for RNA extraction.
FSHR gene cloning
Total RNA was extracted using Trizol (Invitrogen) from turbot ovaries according to the manufacturer’s protocol. Quality and quantity of total RNA were analyzed by 1 % agarose gel electrophoresis and quantified by UV spectrophotometry. To obtain full-length 5′- and 3′-termini of the FSHR gene, the SMART™ RACE cDNA Amplification Kit (Clontech, Palo Alto, CA, USA) was used. Approximately 200 ng of total RNA was used to synthesize 5′- and 3′-RACE-Ready cDNA based on the manufacture’s instruction. Two primers (FSHRF0 and FSHRR0) were designed according to conserved sequences of the FSHR gene from other teleost species to obtain a partial fragment of turbot FSHR cDNA. Based on this partial nucleotide sequence of FSHR, two specific primers (FSHRGSP1 and FSHRGSP2) were designed for 5′- and 3′-RACE using the SMART™ RACE cDNA Amplification Kit (Clontech, Palo Alto, CA, USA). The universal primer (UPM) used for 5′- and 3′-RACE was a mixture of long and short primers (supplied with SMART™ RACE cDNA Amplification Kit, Clontech). The primers mentioned above are listed in Table 1.
Sequence analysis
DNA sequence data were edited and analyzed using the open reading frame (ORF) finder available at NCBI (http://www.ncbi.nlm.nih.gov/gorf/gorf.html) to deduce the translation initiation site (ATG) and the primary structure of FSHR. The structural features of FSHR were searched using TMHMM Server 2.0 (http://www.cbs.dtu.dk/services//TMHMM-2.0/). For signal sequence prediction, the SignalP program was used (http://www.cbs.dtu.dk/services/SignalP/). Prediction of potential N-linked glycosylation sites and phosphorylation sites were carried out using NetNGlyc (http://www.cbs.dtu.dk/services/NetNGlyc/) and NetPhos (http://www.cbs.dtu.dk/services/NetPhos/), respectively. Multiple alignments of amino acid sequences were performed using the software ClustalX 1.81. The neighbor-joining (NJ) phylogenetic tree was constructed using MEGA4.0. Sea lamprey (Petromyzon marinus) glycoprotein hormone receptor I and II were used as the out-group. Reliability of the NJ tree was assessed by interior branch test, using 1000 replications. The deduced amino acid sequences of vertebrate FSHR were acquired from the GenBank database.
Functional analysis
For functional analysis of cloned turbot FSHR, we directly amplified the ORF of FSHR using the primers FSHR–EcoRI and FSHR–BamHI (Table 1). The amplified cDNA were then digested by EcoRI and BamHI, and the digested fragments were ligated into the EcoRI/BamHI site of pEGFP-N3 vector (BD Biosciences Clontech, USA). Then, HEK293T cells were transfected with the pEGFP-N3-FSHR plasmid using Lipofectamine® 2000 reagent (Invitrogen, USA) according to the manufacturer’s instructions. HEK293T cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco, USA) supplemented with 10 % fetal bovine serum (FBS, Hyclone, New Zealand), penicillin (100 Units/ml), and streptomycin (100 μg/ml) in a humidified atmosphere of 5 % CO2 at 37 °C. The continual expressing pEGFP-N3-FSHR HEK293 cells were established in DMEM containing 10 % FBS and 100 μg/ml of G-418 (Roche). The transcripts in the transfected cells were confirmed by observing green fluoresce and RT-PCR after transfection for 24 h. Subsequently, cells were seeded in 24-well culture plates (Nunc, Denmark) at a density of 1.4 × 105/well in DMEM supplemented with 0.1 % FBS and stimulated by adding the specified amounts of hormones in the growth medium for the indicated times. Cells were treated with ovine FSH (OVAGENTM, New Zealand) at 0.1–1000 ng/ml and ovine LH (OVAGENTM, New Zealand) at 0.1–1000 ng/ml for 12 h. Cell lysis and measurement of intracellular cAMP was carried out using the cAMP enzyme immunoassay system Kit (Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s protocol. All measurements were taken in triplicate. The chemicals were dissolved in ethanol or dimethylsulfoxide and then diluted with media. The concentration of ethanol or dimethylsulfoxide in the medium was ≤0.1 %. Control cultures received only vehicle.
Real-time quantitative reverse transcription PCR (qRT-PCR)
Real-time qRT-PCR was performed to determine the relative expression of FSHR mRNA in tissues and the different oocyte developmental stages. Briefly, total RNA was extracted from the collected tissues with Trizol (Invitrogen) and measured using NanoDrop 2000 (Thermo fisher scientific, Rockford, IL, USA). Total RNA was reverse transcribed using a Thermo Fish One step RT-PCR kit according to the manufacturer’s instructions. Levels of FSHR and β-actin mRNAs were determined by real-time RT-PCR using TaKaRa RT-PCR Master Mix reagent and ABI StepOne Plus Sequence Detection System (Applied Biosystems, USA). The SYBR Premix Ex Taq™ Kit (Takara Bio., China) was used for amplification, and the reaction mixture composition was as follows: 10 μL of SYBR® Premix Ex Taq™, 0.8 μL of each primer (10 μM), 0.4 μL of ROX Dye (50×), μL of cDNA sample (25 ng/μL) and 6 μL of sterile distilled water. Initial denaturation was conducted at 95 °C for 10 s, followed by 40 cycles at 95 °C for 5 s and at 60 °C for 30 s. The primers FSHRF/R and β-actinF/R were used to amplify the FSHR and β-actin fragments, respectively (Table 1). Two genes displayed the same amplification efficiency via assessing the standard curve by real-time PCR (Supplemental data, S1). All samples were amplified in triplicates. The relative abundance of FSHR mRNA was normalized to β-actin according to the 2−∆∆CT method (Livak and Schmittgen 2001).
Statistical analysis
The experiment was repeated at least three times for each condition. All data were expressed as mean ± SEM and analyzed by ANOVA and Duncan’s multiple range tests using the SAS 8.0 software. P < 0.05 was considered to be statistically different.
Results
Cloning and characteristics of FSHR gene from turbot
The full-length cDNA sequence of FSHR was obtained by 5′- and 3′-RACE. The FSHR cDNA was 3824 bp long with a 2202 bp ORF coding 733 residues of the mature protein, and 661 bp of 5′ and 961 bp of 3′ terminal untranslated regions (UTRs). The 3′-UTR contained one canonical polyadenylation signal (AATAAA), followed by an additional 12 bp in the poly (A) tail (Fig. 1). Structural analyses revealed that turbot FSHR exhibits typical characteristics of a glycoprotein hormone receptor, which is a subfamily of GPCRs. The turbot FSHR has a large extracellular domain with 387 amino acids, including a putative signal peptide of 18 amino acids, three potential N-linked glycosylation sites, flexible hinge region, and seven-transmembrane (TM) domain upstream of the intracellular C-terminus (Fig. 1). The extracellular loops between TM II–III and TM IV–V each contains a cysteine residue (Cys477 and Cys552) that presumably links the extracellular loops via a disulfide bridge (Fig. 1). The intracellular loops between TM V–VI and the intracellular C-terminal domain contain serine and threonine residues that are potential phosphorylation sites (Fig. 1).

Nucleotide (upper line) and deduced amino acid (lower line) sequences of the turbot FSHR. The start codon (ATG) was boxed, and the stop codon (TGA) was marked with an asterisk. The signal peptide sequence and the polyA signal in 3′-UTR are underlined. The position of the seven predicted transmembrane domain is showed as black box. Two conserved cysteins (477C, and 552C), predicted to form an intramolecular disulfide bond, are indicated by dots. Potential N-glycosylation and phosphorylation sites are indicated by open and closed triangles
Phylogenetic analysis and alignment
Comparison of the turbot FSHR amino acid sequence with teleost, mammalian and other vertebrates showed that turbot FSHR has high homologies with FSHRs of other fish species, followed by mouse, chicken and frog FSHRs (Fig. 2). In addition, turbot FSHR was found to contain three specific insertions (aa 162–174, 421–434, 580–592) that are not present in other pleuronectiformes (Fig. 2). Comparison with the paralogous genes LHR and thyroid stimulating hormone receptor (TSHR) demonstrated that the turbot FSHR protein is closely related to Hippoglossus hippoglossus proteins, as depicted in the phylogenetic tree in Fig. 3.

Alignment of the amino acid sequence of the turbot FSHR (SM-FSHR) proteins from various species. Position with >70 % similarity are shaded in light gray, while completely conserved positions are shaded in black. Accessions number: Cynoglossus semilaevis (CS) ACD39387.2, Epinephelus coioides (EC) AEG65826.1, Hippoglossus hippoglossus (HH) ACB13177.1, Solea senegalensis (SS) ADH51678.1, Clarias gariepinus (CG) AJ012647.2, Danio rerio (Z) AAP33512.1, Xenopus laevis (XL) NM_001256260.1, Gallus gallus (GG) NM_205079.1, Mus musculus (M) NM_013523.3, Humo sapiens (H) M65085.1

The neighbor-joining phylogenetic tree of vertebrates based on the amino acid sequences of TSHR, FSHR and LHR. Accession number: FSHR Acanthopagrus schlegelii (ABU49599.1), FSHR Dicentrarchus labrax (AY642113.1), FSHR Epinephelus coioides (AEG65826.1), FSHR Hippoglossus hippoglossus (ACB13177.1), FSHR Solea senegalensis (ADH51678.1), FSHR Cynoglossus semilaevis (ACD39387.2), FSHR Oncorhynchus mykiss (AF439405.1), FSHR Clarias gariepinus (AJ012647.2), FSHR Danio rerio (AAP33512.1), FSHR Xenopus laevis (NM_001256260.1), FSHR Gallus gallus (NM_205079.1), FSHR Humo sapiens (M65085.1), FSHR Mus musculus (NM_013523.3), TSHR Danio rerio (NM_001145763.2), TSHR Homo sapiens (AY429111.1), TSHR Gallus gallus (NM_001193589.1), TSHR Macaca mulatta (NM_001195395.1), TSHR Clarias gariepinus (AY129556.1), TSHR Solea senegalensis (FN677495.1), LHR Xenopus laevis (AB602929), LHR Gallus gallus (AB009283), LHR Homo sapiens (S57793), LHR Mus musculus (M81310), LHR Hippoglossus hippoglossus (EU502845), LHR Danio rerio (AY714133), LHR Ctenopharyngodon idella (EF194761), LHR Clarias gariepinus (AF324540), LHR Oncorhynchus rhodurus (AB030005), LHR Oncorhynchus mykiss (AF439404), LHR Solea senegalensis (GQ472140), LHR Salmo salar (AJ579790), LHR Sparus aurata (AY587261), LHR Dicentrarchus labrax (AY642114), LHR Epinephelus coioides (HQ650770). Sea lamprey (Petromyzo marinus) glycoprotein hormone receptor (GpHR) I (AY750689) and GpHR II (AY750689) were used as the out-groups. Bootstrap values (in %) from 1000 replicates are indicated for each tree node
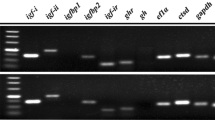
Tissue distribution of the gene expression of FSHR
The tissue distribution of FSHR was analyzed by qRT-PCR. FSHR mRNA was found to be significantly most abundant in the ovary (Fig. 4a, P < 0.05) followed by the liver. Expression of the FSHR was also detected in several extra-gonadal tissues such as the eyes, intestine, brain, muscle, gills, spleen, stomach, heart and kidney at significantly lower levels than the ovary (Fig. 4a, P < 0.05). However, FSHR mRNA was not found to be significantly different among these extra-gonadal organs (Fig. 4a, P > 0.05). FSHR mRNA was also detected in testis, albeit at significantly lower levers compared to the ovary (Fig. 4b, P < 0.05). Furthermore, the tissue distribution of FSHR mRNA was found to be similar to that of LHR mRNA in turbot (supplemental data, S2A). However, FSHR mRNA was significantly higher than LHR mRNA in the ovary and the liver (supplemental data, S2A).

Quantitative analyses of FSHR mRNA expression in different tissues of turbot. Values represent the mean ± SEM (n = 6). Bars with different superscripts are statistically different (P < 0.05)
The expression profile of the FSHR gene during ovarian developmental stages
Gene expression profiles of FSHR at distinct developmental stages of the ovary were examined by qRT-PCR. FSHR mRNA gradually increase from the Prevtg to the Mig-nucl stage, with the highest values observed during the Latvtg stage (Fig. 5, P < 0.05). However, FSHR mRNA decreased dramatically during the Atre stage (Fig. 5, P < 0.05). Meanwhile, the highest values of FSHR mRNA and LHR mRNA were obtained in the Latvtg and Mig-nucl stages, respectively (supplemental data, S2B).

Quantitative analyses of FSHR mRNA expression in ovarian developmental stages of turbot. Values represent the mean ± SEM (n = 6). Bars with different superscripts are statistically different (P < 0.05)
Functional analysis of the FSHR
HEK293T cells constantly expressed turbot FSHR after transfection with the pEGFP-N3-FSHR plasmid (Fig. 6). The HEK293T cells can continually express turbot FSHR after the transfection for 24 h (supplemental data, S3). Then, the transfected HEK293T cells were treated with ovine FSH (0.1–1000 ng/ml) and ovine LH (0.1–1000 ng/ml) for 12 h. After treatment with ovine FSH, intercellular cAMP level of HEK293T cells continual expressing FSHR evidently increased in a dose-dependent manner from 0.1 to 100 ng/ml, but decreased at 1000 ng/ml (Fig. 7a, P < 0.05). However, the intercellular cAMP level of HEK293T cells continual expressing FSHR did not change upon treatment with ovine LH at all concentrations tested (0.1–1000 ng/ml; Fig. 7b, P > 0.05).

FSHR was transfected into HEK293T cells by PEGFP-N3 vector. a The expression of GFP gene in HEK293T cells after transfected 24 h. b PCR-detection of FSHR in transfected HEK293T cells. GFP-labeled HEK293T cells are denoted by arrows. Trans transfect; CTL control

Functional analysis of FSHR. HEK293T cells expressed the FSHR were treated by ovine FSH (a) and ovine LH (b), respectively. Values represent the mean ± SEM (n = 5). Bars with different superscripts are statistically different (P < 0.05)
Discussion
As the most diverse group of vertebrates, fish have various modes of ovarian development and reproductive strategies (Nagahama and Yamashita 2008). The physiological function of gonadotropins and their receptors during the reproductive cycle in teleost is highly controversial (Levavi-sivan et al. 2010). In present study, the FSHR cDNA of turbot was cloned from ovarian tissue and its structural characteristics and expression profiles were evaluated. Analysis of the nucleotide and deduced amino acid sequence of turbot FSHR revealed that this receptor is highly homologous to teleost FSHRs and shows typical structural features of glycoprotein hormone receptors. A relatively long extracellular domain and rhodopsin-like seven TM module were observed. Meanwhile, a highly conserved amino acid sequence (YPSHCCAF) proposed to form a pocket for specific glycoprotein hormone binding and located at the C-terminal exon that forms the extracellular domain of mammalian GtHRs (Lloyd and Griswold 1995) was found to be entirely conserved within the turbot FSHR amino acid sequence (322–329). Cys477 and Cys552 were found to be located in the extracellular loops I and II of the TM domain and were believed to connect the two loops via a disulfide bridge (Levavi-Sivan et al. 2010). Three specific amino acid sequences (162–174, 421–434, 580–592) in the ligand-binding domain and were found to be unique in turbot compared to other pleuronectiformes. Moreover, three potential N-linked glycosylation sites and five phosphorylation sites were identified in the N-terminal and C-terminal regions of turbot FSHRs, respectively. These structural features of FSHR are crucial determinants of ligand binding affinity and signal transduction and play important roles in hormone recognition and receptor activation (Moyle et al. 2004; Vassart et al. 2004; Kene et al. 2005; Ulloa-Aguirre et al. 2007; Jiang et al. 2014). Furthermore, phylogenetic analysis showed that turbot FSHR has significant homology with H. hippoglossus FSHR. These results indicated that the basic structure and characteristics of turbot FSHR are similar to those of other fish and mammalian species and provide valuable insights into the structural and functional evolution of GtHRs in fish and more generally in vertebrates.
FSHRs are mainly expressed on the surface of ovarian cells and involved in mediating ovarian steroidogenesis, vitellogenesis and may affect follicular development via complex signaling pathways in fish and mammals (Clelland and Peng 2009; Luckenbach et al. 2011; Liu and Ge 2013; Lan et al. 2014; Chakraborty and Roy 2015). Meanwhile, FSHRs are also found in extragonadal tissues, including the reproductive tract, placenta, umbilical cord and brain (Ziecik et al. 1992; Liang et al. 2012). In the present study, turbot FSHR was found to be expressed more abundantly in the ovaries than in other tissues, which is in agreement with reports on other fish species (Oba et al. 1999a; Kumar et al. 2001b; Kwok et al. 2005; Maugars and Schmitz 2006; Rocha et al. 2007; Kobayashi et al. 2008; Ohkubo et al. 2013). However, extragonadal expression of FSHR in turbot was found to be different from that in several other fish species. No extragonadal expression of FSHR has been found in Atlantic salmon (Maugars and Schmitz 2006) and sea bass (Rocha et al. 2007), while remarkable expression of FSHR in the pituitary has been reported in the Atlantic halibut (Kobayashi et al. 2008). The difference in FSHR expression levels among extragonadal tissues may be species-specific and should be investigated further along with the in vivo functions of the gene. It is noteworthy in this context that the results of the present study suggest that turbot FSHR is associated with ovarian development and may have a function in female turbot reproductive cycle.
Oocyte development is triggered by gonadotropins, and the subsequent steroidogenic production of estradiol induces vitellogenesis, causing a marked enlargement in oocyte size (Hillier 2001). Meanwhile, gonadotropins are involved in the regulation of oocyte meiotic resumption and oocyte maturation (Fortune 1994). Complex hormonal cross-talk between the developing oocyte and its surrounding follicular cells has been observed (Canipari 2000; Magoffin 2005). Most teleost fish species are oviparous and have three main types of ovarian development based on oocyte development (synchronous, group-synchronic and asynchronous) (Wallace and Selman 1981). Therefore, the regulatory mechanisms of oocyte development and reproductive strategies may differ among different fish species. Turbot is a serial spawner, producing multiple egg batches during reproductive season. Histological observations of the turbot ovary revealed that oocyte maturation involved a vitellogenesis step before the oocyte maturation and ovulation stages in our previous study (Jia et al. 2014b). In the current study, FSHR expression was found to increase from previtellogenic to migratory nucleus stages, and the highest expression level was observed at the oocyte late vitellogenic stage. Meanwhile, we also found the FSHR mainly located on the theca cell in vitellogenic follicles of turbot (Data not shown). Studies on multiple and group-synchronous spawners demonstrated that FSHR is mostly correlated with different phases of follicular development and predominantly associated with vitellogenesis (Hirai et al. 2002; Kwok et al. 2005). Similar results were observed in European sea bass (Rocha et al. 2009). These results suggest that FSHR is involved in the stimulation of vitellogenesis and oocyte maturation, thereby promoting ovarian development of turbot during its reproductive cycle.
In mammals, gonadotropin hormone/receptor interactions are very specific, with no cross-stimulation occurring under physiological conditions. However, promiscuous activation of fish GtHRs has been described in salmonids, catfish and other teleosts (Oba et al. 1999a,b; Kumar et al. 2001a,b; Vischer and Bogerd 2003; Kwok et al. 2005; Bogerd 2007). In the present study, turbot FSHR displayed ligand selectivity as it was only activated by ovine FSH, but not by ovine LH. Strict ligand selectively has also been reported in several other species such as rainbow trout (Sambroni et al. 2007), sea bass (Molés et al. 2011), mummichog (Ohkubo et al. 2013), and chub mackerel (Nyuji et al. 2013). Similar results have been observed in zebrafish and amago salmon (Kwok et al. 2005; Oba et al. 1999b). Nevertheless, mammalian hormones can act in a different way than other fish GtHRs (Bogerd 2007; Levavi-Sivan et al. 2010). This difference in ligand selectivity could suggest that the action of GtHRs in teleosts does not overlap fully with that of their mammalian counterparts, and underscores the need for learning more from turbot and other fish.
In conclusion, full-length cDNAs coding for FSHR were cloned from turbot ovaries. Cloning and structural characterization of FSHR provided new insights into the conservation of glycoprotein hormone receptors among vertebrate. Furthermore, our data on the distribution and expression profiles of FSHR are consistent with its involvement in vitellogenesis and oocyte maturation of turbot. In addition, functional analysis with HEK293T cells continual expressing FSHR demonstrated that FSHR is specifically stimulated by ovine FSH, but not ovine LH. These results strongly suggest that FSHR may have crucial functions in regulating turbot ovarian and follicular development. However, additional investigation using recombinant turbot LH and FSH is essential to determine the spatiotemporal expression patterns and ligand availability of LHR and FSHR which will be helpful in further understanding the functions of GtHRs in regulating the reproductive cycle in fish.
References
Andersson E, Schulz RW, Male R, Bogerd J, Patiña D, Benedet S, Norberg B, Taranger GL (2013) Pituitary gonadotropin and ovarian gonadotropin receptor transcript levels: seasonal and photoperiod-induced changes in the reproductive physiology of female Atlantic salmon (Salmo salar). Gen Comp Endocrinol 191:247–258
Bogerd J (2007) Ligand-selective determinants in gonadotropin receptors. Mol Cell Endocrinol 260(262):144–152
Bromley PJ, Ravier C, Witthames PR (2000) The influence of feeding regime on sexual maturation, fecundity and atresia in first-time spawning turbot. J Fish Biol 56:264–278
Canipari R (2000) Oocyte-granulosa cell interactions. Hum Reprod Update 6:279–289
Chaffin CL, Vandevoort CA (2013) Follicle growth, ovulation, and luteal formation in primates and rodents: a comparative perspective. Exp Biol Med 238:539–548
Chakraborty P, Roy SK (2015) Expression of FSH receptor in the hamster ovary during perinatal development. Mol Cell Endocrinol 400:41–47
Clelland E, Peng C (2009) Endocrine/paracrine control of zebrafish ovarian development. Mol Cell Endocrinol 312:42–52
Fortune JE (1994) Ovarian follicular growth and development in mammals. Biol Reprod 50:225–232
Hillier SG (2001) Gonadotropic control of ovarian follicular growth and development. Mol Cell Endocrinol 179:39–46
Hirai T, Oba Y, Nagahama Y (2002) Fish gonadotropin receptors: molecular characterization and expression during gametogenesis. Fish Sci Suppl 68:675–678
Hurk R, Zhao J (2005) Formation of mammalian oocytes and their growth, differentiation and maturation within ovarian follicles. Theriogenology 63:1717–1751
Jia YD, Meng Z, Liu XF, Lei JL (2014a) Biochemical composition and quality of turbot (Scophthalmus maximus) eggs throughout the reproductive season. Fish Physiol Biochem 40:1093–1104
Jia YD, Meng Z, Niu HX, Peng H, Lei JL (2014b) Molecular cloning, characterization, and expression analysis of luteinizing hormone receptor gene in turbot (Scophthalmus maximus L). Fish Physiol Biochem 40:1639–1650
Jiang XL, Dias JA, He XL (2014) Structural biology of glycoprotein hormone and their receptors: insights to signaling. Mol Cell Encocrinol 382:424–451
Jones BA (1974) Sexual maturity, fecundity and growth of the turbot Scophthalmus maximus L. J Mar Biol Assoc UK 54:109–125
Kene PS, Dighe RR, Mahale SD (2005) Delineation of regions in the extracellular domain of follicle-stimulating hormone receptor involved in hormone binding and signal transduction. Am J Reprod Immunol 54:38–48
Kobayashi T, Andersen Ø (2008) The gonadotropin receptors FSH-R and LH-R of Atlantic halibut (Hippoglossus hippoglossus), 1: isolation of multiple transcripts encoding full-length and truncated variants of FSH-R. Gen Comp Endocrinol 156:584–594
Kobayashi T, Pakarinen P, Torgersen J, Huhtaniemi I, Andersen Ø (2008) The gonadotropin receptors FSH-R and LH-R of Atlantic halibut (Hippoglossus hippoglossus)-2. Differential follicle expression and asynchronous oogenesis. Gen Comp Endocrinol 156:595–602
Kumar RS, Ijiri S, Trant JM (2001a) Molecular biology of channel catfish gonadotropin receptors: 1. Cloning of a functional luteinizing hormone receptor and preovulatory induction of gene expression. Biol Reprod 64:1010–1018
Kumar RS, Ijiri S, Tran JM (2001b) Molecular biology of the channel catfish gonadotropin receptors: 2. Complementary DNA cloning, functional expression, and seasonal gene expression of the follicle-stimulating hormone receptor. Biol Reprod 65:710–717
Kwok HF, So WK, Wang Y, Ge W (2005) Zebrafish gonadotropins and their receptors: I. Cloning and characterization of zebrafish follicle-stimulating hormone and luteinizing hormone receptors-evidence for their distinct functions in follicle development. Biol Reprod 72:1370–1381
Lan RX, Liu F, He ZB, Chen C, Liu SJ, Shi Y, Liu YL, Yoshimura Y, Zhang M (2014) Immunolocalization of GnRHR I, gonadotropin receptors, PGR, and PGRMC I during follicular development in the rabbit ovary. Theriogenology 81:1139–1147
Levavi-Sivan B, Bogerd J, Mañanós EL, Gómez A, Lareyre JJ (2010) Perspectives on fish gonadotropins and their receptors. Gen Comp Endocrinol 165:412–437
Liang H, Chen L, Zhou Liu, Zhou X (2012) Expression of functional follicle-stimulating hormone receptor and luteinizing hormone/chorionic gonadotrophin receptor in oviduct and uterus in prepubertal gilts. Live Sci 148:74–80
Liu KC, Ge W (2013) Differential regulation of gonadotropin receptors (fshr and lhcgr) by epidermal growth factor (EGF) in the zebrafish ovary. Gen Comp Endocrinol 181:288–294
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real time quantitative PCR and the 2−∆∆CT method. Methods 25:402–408
Lloyd TL, Griswold MD (1995) Sequence analysis of glycoprotein hormone receptors for follicle-stimulating hormone, luteinizing hormone, thyroid-stimulating hormone. Biol Reprod 52(suppl 1):102
Lubzens E, Young G, Bobe J, Cerdà J (2010) Oogenesis in teleosts: how fish eggs are formed. Gen Comp Endocrinol 165:367–389
Luckenbach JA, Dickey JT, Swanson P (2011) Follicle-stimulating hormone regulation of ovarian transcripts for steroidogenesis-related proteins and cell survival, growth and differentiation factors in vitro during early secondary oocyte growth in coho salmon. Gen Comp Endocrinol 171:52–63
Magoffin DA (2005) Ovarian theca cell. Int J Biochem Cell Biol 37:1344–1349
Maugars G, Schmitz M (2006) Molecular cloning and characterization of FSH and LH receptors in Atlantic salmon (Salmo salar L.). Gen Comp Endocrinol 149:108–117
McEvoy LA (1989) Reproduction of turbot Scophthalmus maximus L in captivity. Cuard Aera Cienc Mar Semin Estud Galegos 3:9–28
Menon KMJ, Menon B (2012) Structure, function and regulation of gonadotropin receptors-a perspective. Mol Cell Endocrinol 356:88–97
Mittelholzer C, Andersson E, Taranger GL, Consten D, Hirai T, Senthilkumaran B, Nagahama Y, Norberg B (2009) Molecular characterization and quantification of the gonadotropin receptors FSH-R and LH-R from Atlantic cod (Gadus morhua). Gen Comp Endocrinol 160:47–58
Molés G, Gómez A, Carrillo M, Rocha A, Mylonas CC, Zanuy S (2011) Determination of Fish quantity and bioactivity during sex differentiation and oogenesis in European sea bass. Biol Reprod 85:848–857
Moyle WR, Xing YN, Lin W, Cao DH, Myers RV, Kerrigan JE, Bernard MP (2004) Model of glycoprotein hormone receptor ligand binding and signaling. J Biol Chem 279:44442–44459
Mugnier C, Guennoc M, Lebegue E, Fostier A, Breton B (2000) Induction and synchronisation of spawning in cultivated turbot (Scophth almus maximus L.) broodstock by implantation of a sustained-release GnRH-a pellet. Aquaculture 181:241–255
Nagahama Y, Yamashita M (2008) Regulation of oocyte maturation in fish. Dev Growth Differ 50:S195–S219
Nyuji M, Kitano H, Shimizu A, Lee JM, Kusakabe T, Yamaguchi A, Matsuyama M (2013) Characterization, localization, and stage-dependent gene expression of gonadotropin receptors in chub mackerel (Scomber japonicus) ovarian follicles. Biol Reprod 88(148):1–14
Oba Y, Hirai T, Yoshiura Y, Yoshikuni M, Kawauchi H, Nagahama Y (1999a) Cloning, functional characterization, and expression of a gonadotropin receptor cDNA in the ovary and testis of amago Salmon (Oncorhynchus rhodurus). Biochem Biophys Res Commun 263:584–590
Oba Y, Hirai T, Yoshiura Y, Yoshikuni M, Kawauchi H, Nagahama Y (1999b) The duality of fish gonadotropin receptors: cloning and functional characterization of a second gonadotropin receptor cDNA expressed in the ovary and testis of amago Salmon (Oncorhynchus rhodurus). Biochem Biophys Res Commun 265:366–371
Oba Y, Hira T, Yoshiura Y, Kobayashi T, Nagahama Y (2001) Fish gonadotropin and thyrotropin receptors: the evolution of glycoprotein hormone receptors in vertebrates. Comp Biochem Physiol B 129:441–448
Ohkubo M, Yabu T, Yamashita M, Shimizu A (2013) Molecular cloning of two gonadotropin receptors in mummichog Fundulus heteroclitus and their gene expression during follicular development and maturation. Gen Comp Endocrinol 184:75–86
Ribas L, Pardo BG, Fernández C, Álvarez-Diós JA, Gómez-Tato A, Quiroga MI, Planas JV, Sitjà-Bobadilla A, Martínez P, Piferrer F (2013) A combined strategy involving Sanger and 454 pyrosequencing increases genomic resources to aid in the management of reproduction, disease control and genetic selection in the turbot (Scophthalmus maximus). BMC Genom 14:180
Rocha A, Gómez A, Zanuy S, Cerdá-Reverter JM, Carrillo M (2007) Molecular characterization of two sea bass gonadotropin receptors: cDNA cloning, expression analysis, and functional activity. Mol Cell Endocrinol 272:63–76
Rocha A, Zanuy S, Carrillo M, Gómez A (2009) Seasonal changes in gonadal expression of gonadotropin receptors, steroidogenic acute regulatory protein and steroidogenic enzymes in the European sea bass. Gen Comp Endocrinol 162:265–275
Sambroni E, Le Gac F, Breton B, Lareyre JJ (2007) Functional specificity of the rainbow trout (Oncorhynchus mykiss) gonadotropin receptors as assayed in a mammalian cell line. J Endocrinol 195:213–228
Suquet M, Billard R, Cosson J, Normant Y, Fauvel C (1995) Artificial insemination in turbot (Scophthalmus maximus): determination of the optimal sperm to egg ratio and time of gamete contact. Aquaculture 133:83–90
Swanson P, Dickey JT, Campbell B (2003) Biochemistry and physiology of fish gonadotropins. Fish Physiol Biochem 28:53–59
Ulloa-Aguirre A, Zariñán T, Pasapera A, Casas-González P, Dias J (2007) Multiple facets of follicle-stimulating hormone receptor function. Endocrine 32:251–263
Vassart G, Pardo L, Costagliola S (2004) A molecular dissection of the glycoprotein hormone receptors. Trends Biochem Sci 29:119–126
Vischer HF, Bogerd J (2003) Cloning and functional characterization of a gonadal luteinizing hormone receptor complementary DNA from the African Catfish (Clarias gariepinus). Biol Reprod 68:262–271
Wallace RA, Selman K (1981) Cellular and dynamic aspects of oocyte growth in teleosts. Am Zool 21:325–343
Ziecik AJ, Derecka-Reszka K, Rzucidło SJ (1992) Extragonadal gonadotropin receptors, their distribution and function. Physiol Pharmacol 43:33–49
Acknowledgments
This study was supported by China Agriculture Research System (CRAS-50), National Natural Science Foundation of China (31302205, 31402315 and 31402284), Natural Science Foundation of Shandong Province (ZR2012CQ024 and BS2013SW004) and the China Postdoctoral Science Foundation (2012M511559 and 2013T60690). We thank Chunren Gao and Xinfu Liu (Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences) for help in the experiment.
Author information
Authors and Affiliations
Corresponding author
Additional information
Yudong Jia and Ai Sun have equal contribution to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Jia, Y., Sun, A., Meng, Z. et al. Molecular characterization and quantification of the follicle-stimulating hormone receptor in turbot (Scophthalmus maximus). Fish Physiol Biochem 42, 179–191 (2016). https://doi.org/10.1007/s10695-015-0128-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-015-0128-8