
Abstract
The initial domestication of plants and animals and the subsequent emergence of agricultural economies, which began independently more than 10,000 years ago in a number of different world regions, represent a major evolutionary transition in earth history. It is these domesticates, and the agricultural economies based on them, that have formed the lever with which humans have substantially modified the earth’s terrestrial ecosystems over the past ten millennia. General explanations for this transition from hunting and gathering to food production economies formulated over the past 40 years have been based on standard evolutionary theory (SET) and employ the assumption of unidirectional adaptation—that environments change and species adapt. Here I compare these proposed SET—based externalist explanations for domestication with a recently formulated alternative developed from niche construction theory (NCT). Archaeological and paleoenvironmental records from two independent centers of domestication in the Americas—eastern North America and the Neotropics of northern South America, are found to support the NCT-based explanatory approach but not the SET explanations, underscoring the limitations of externalist SET approaches and the need for broader conceptualization of the processes that direct evolutionary change in order to gain a better general understanding of initial domestication as well as other major evolutionary transitions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Even though Homo sapiens sapiens are acknowledged as being the “ultimate niche constructors” (Odling-Smee et al. 2003; Smith 2007a), analysis of human societies is often considered as having relatively little to contribute to a better understanding of the general role and relative importance of niche construction in shaping ecosystems and evolutionary trajectories. The reason for this apparent paradox is obvious enough—humans have been so successful in modifying their physical and social environments over the past 10,000 years that modern human societies not only largely exist outside of “natural” ecosystems, but also are recognized as being the major cause of their ongoing destruction.
Since the 1930s, however, research worldwide has documented the different ways in which small-scale pre-industrial human societies modify and manage their local ecosystems in a sustainable, long-term manner (e.g. Anderson 2005; Deur and Turner 2005; Lepofsky 2009; Smith 2011). These studies of human niche construction by past and present-day small-scale societies in turn provide the foundation for a new perspective on one of the major evolutionary transitions in earth history—the initial domestication of plants and animals that began about 10,000 years ago (Smith 2012). Over the past ten millennia the farming economies based on these domesticates have provided the lever with which humans have transformed the earth by inexorably expanding agricultural landscapes and gaining ever-increasing control of the earth’s terrestrial ecosystems (Smith and Zeder 2013).
This new niche construction theory (NCT)-based perspective on the initial domestication of plants and animals by human groups offers an alternative approach to a long series of proposed explanations for this evolutionary transition that are derived from standard evolutionary theory (SET). In this article I compare the new NCT-based explanation for domestication with those based on SET, with emphasis on the most recently formulated SET-based explanation, which is linked explicitly to neo-Darwinism and derived from optimal foraging theory (OFT) (Gremillion et al. 2014). This side-by-side consideration of these two alternative general explanatory approaches, one based on standard evolutionary theory and the other based on niche construction theory, shows that the NCT explanatory framework is much better supported by currently available archaeological and paleoenvironmental data sets, and that it redirects research on domestication and broadens the scope of analysis in a manner that promises to advance in our understanding of this major evolutionary transition (Laland and Sterelny 2006: 1760).
This comparison of alternative SET and NCT-based explanations is greatly facilitated by a dramatic increase in available empirical information over the past several decades. Domestication and agricultural origins are no longer viewed as a single monolithic research question, but rather are now recognized as comprising a higher order “research domain” encompassing a substantial expanse of space and time and a wide range of different research questions, data sets, scales of analysis, and analytical approaches (Smith 2015a). The shift from hunter-gatherer subsistence systems to farming economies is now being studied in an increasing number of world areas, including both the eight or more regions that witnessed the independent domestication of plants and animals, as well as other regions into which domesticates and farming economies subsequently diffused (Bar-Yosef and Price 2011). Each of these regions witnessed a distinct evolutionary trajectory from foraging to farming, and taken together they represent a set of constantly updated comparative case studies for anyone interested in looking for variables that may have played a role in initial domestication and agricultural origins across a number of different areas of the world.
It is also increasingly evident that the initial domestication of plants and animals was temporally separated from the subsequent emergence of “agriculture” (i.e. “the near total reliance upon domesticated plants or animals” Winterhalder and Kennett 2006: 3) in different world areas by thousands of years (Smith 2001, 2015a). During the intervening millennia between initial domestication and the development of fully agricultural economies, small-scale societies had “low-level food production” economies that included domesticates but were also strongly reliant on wild species of plants and animals (Smith 2001). Although still often conflated (e.g. Gremillion et al. 2014), initial domestication and the emergence of agriculture actually represent two temporally and developmentally distinct evolutionary transitions. The vast majority of recent and ongoing research focuses on initial domestication, with far fewer studies addressing the subsequent development of agriculture (Smith 2015a).
Research on initial domestication is carried out at a number of different nested scales of analysis and interpretation, ranging from tightly focused to quite broad, including: (1) analysis of particular aspects of domestication of a single species; (2) comprehensive descriptive synthesis of domestication of a single species; (3) descriptive synthesis of the process of domestication of multiple species within particular world regions; and (4) general synthesis frameworks of explanation for initial domestication that are applicable to multiple regions and which attempt to identify and focus on common underlying causal variables involved in domestication worldwide. The vast majority of efforts to account for the shift from hunting and gathering to food production at a level 4, global-scale of analysis that have been proposed over the past half century have conformed to the general principles of standard evolutionary theory.
General explanations for initial domestication based on standard evolutionary theory
In spite of the recent growth of more interactionist subfields in contemporary evolutionary biology (e.g. coevolution), it has been argued that an “externalist stance can still be regarded as the ‘default’ position for many (Godfrey-Smith 1996; Odling-Smee et al. 2003)” (Laland et al. 2012). This default position includes the core assumption that adaptation to change is unidirectional or asymmetrical: “Adaptation is always asymmetrical; organisms adapt to their environment, never vice versa” (Williams 1992: 484). NCT advocates argue that this assumption of unidirectionality, that: “The causal arrow points in one direction only: environments are the source of selection, and they determine the features of living creatures,” has “underpinned evolutionary thought since the Modern Synthesis” (Laland et al. 2009: 197). The extent to which evolutionary biology retains an externalist perspective, however, remains a topic of considerable current debate ((Laland and Sterelny 2006; Laland et al. 2012, 2013; Scott-Phillips et al. 2013; Laland et al. 2014, Wray et al. 2014).
Archaeologists frequently turn to other disciplines for potential new theoretical frameworks, and given the time-lag often associated with such interdisciplinary borrowing, it is not surprising that efforts to employ SET in explaining initial domestication that have been proposed in the half century since Mayr’s classic paper on causation (Mayr 1961) have been strongly influenced by the “default” position in evolutionary biology—that adaptation is unidirectional. In these SET based explanatory frameworks for initial domestication that rely on the assumption of unidirectional adaptation, the shift from hunting and gathering to food production invariably begins with the development of a non-equilibrium condition in the local environment—in this case an imbalance between the energy requirements of the human groups in question, and what is available in their local ecosystem. Within a decade of Mayr’s paper, a prominent archaeologist succinctly articulated this externalist assumption of unidirectional adaptation: “Adaptation is always a local problem, and selective pressures favoring new cultural forms result from non-equilibrium conditions in the local ecosystem” (Binford 1971: 34). These disequilibrium conditions can emerge on either the supply side or the demand side. On the supply side: “A change in the physical environment of a population which brings about a reduction in the biotic mass of the region would decrease the amounts of available food. The previous balance between population and standing crop is upset, and more efficient extractive means would be favored” (i.e. domestication). On the demand side: “Change in the demographic structure of a region…would also upset an established equilibrium system, and might serve to increase the population density of a region beyond the carrying capacity of the natural environment. Under these conditions, manipulation of the natural environment in order to increase its productivity would be highly advantageous”.
Binford (1971: 44–45) combines disequilibrium on both the supply side and demand side in his proposed explanatory framework for initial domestication, with post-Pleistocene human population growth within regions having higher carrying capacity (“natural habitat zones”) leading to the “budding off” or spillover of daughter communities into adjacent regions having lower carrying capacity. Groups moving into these lower carrying capacity “population frontiers” or “adaptive tension zones” would encounter “disequilibrium between population and resources, which in turn, would offer selective advantage to increases in the efficacy of subsistence technology… it is in the context of such situations of stress in environments with plant and animal forms amenable to manipulation that we would expect to find conditions favoring the development of plant and animal domestication.” In Binford’s general scenario, post-Pleistocene human population growth forces expansion into marginal areas, where lower carrying capacity results in the adaptive response of domestication.
In his influential 1977 book The Food Crisis in Prehistory, Cohen, like Binford, adopts the default SET unidirectional adaptation assumption, and argues that a world-wide demographic threshold was crossed during the Early Holocene, resulting in an imbalance between human populations and their subsistence base. In those world regions that both experienced the most acute imbalance between food resources and human population, and which supported species of plants and animals that could be domesticated, agriculture arose as an adaptive response and a means of resolving demographically induced resource pressure (Cohen 1977).
In a similar and more recent application of the asymmetrical adaptation assumption of standard evolutionary theory in attempting to explain the transition from hunting and gathering to food production economies on a global scale, Richerson et al. (2010: 389) employ “Darwinian methods” in arguing that early Holocene stabilization of global climates ushered in rapid growth in human population levels, and when “the carrying capacity set by the environment and the efficiency of the prevailing subsistence system” was reached,
local communities that discover or acquire more intensive subsistence strategies will increase in number and exert competitive pressure on smaller populations with less intensive strategies. Thus, in the Holocene, such inter-group competition generated a competitive ratchet favoring the origin and diffusion of agriculture (Richerson et al. 2010: 389).
Note that in contrast to Winterhalder and Kennett (2006), Richerson et al. (2010: 410) equate initial domestication with agricultural origins: “We mark the origin of agriculture as the first horizon in which plant remains having anatomical markers of domestication are found.” The unidirectional “causal arrow” in the Richerson et al. (2010) explanation of initial domestication begins with early Holocene climatic amelioration setting the stage for rapid human population growth, which eventually and inevitably reaches carrying capacity, in turn causing intergroup competitive pressure and providing a selective advantage to those groups that domesticate species as an adaptive response.
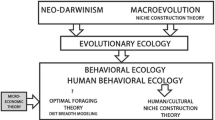
Like the explanatory frameworks proposed over the past half century by Binford, Cohen, Richerson et al., and others, recent efforts to account for initial domestication based on optimal foraging theory (OFT) similarly employ the key assumption of unidirectional adaptation while also explicitly deriving their proposed explanations from higher-level neo-Darwinian theory. Gremillion et al. (2014: 6171) argue that neo-Darwinism has “earned status as an overarching framework for explaining the diversity of life,” that “…evolutionary theory is central to understanding the root causes of human behavior and indeed culture itself;” and that “…evolutionary theory (broadly construed to include cultural as well as biological processes) must play a central role in OA [origins of agriculture] research” (Gremillion et al. 2014: 6171); and that “Such high-level bodies of general theory inform middle- and lower-level theories that in turn generate testable hypotheses” (Gremillion et al. 2014: 6172).Footnote 1 The diet breadth model (DBM), one of the family of OFT models, provides the theoretical framework for proposed explanations of initial domestication, as it “is particularly well suited for studying major directional changes in human subsistence over time because of its ability to make robust, qualitative predictions of prey choice and dietary diversity” (Piperno 2006: 141).
In DBM explanations, domestication is cast as an adaptive response by human societies to an environmental disequilibrium—an imbalance in available energy within the local ecosystem. Such an imbalance, or “resource depression” in DBM terminology (Winterhalder and Kennett 2006), can be caused either on the supply side, by a change in the local biotic community that results in a lowering of the human carrying capacity of a resource catchment area, or on the demand side, due to an increase in human population density resulting in higher harvesting levels of local resources.
Under the rules of the DBM, human societies structure their harvesting of plant and animal resources according to optimization principles that are assumed to increase the fitness of individuals making specific harvesting or “prey choice” decisions, and when taken in aggregate, the fitness of larger social groups. Energy or net caloric return is the “currency” invariably employed in DBM efforts to explain initial domestication. Human groups are modeled as formulating and always following an explicit ranked list of all of the plant and animal components of the local biotic community, with the ranking of any resource based on its energy value. Working down the ranked list, an “optimal diet” boundary is eventually reached where the next lower resource is estimated to provide a lower energy yield that the overall energy yield of the ranked resources above it. When encountered, resources below this “optimal diet” boundary line are always ignored, regardless of their abundance, in favor of continuing to search for those that are ranked above it. When an imbalance between energy supply and demand (resource depression) occurs, the adaptive response by human groups to the associated decline in foraging efficiency takes the form of extending the “optimal diet” boundary farther down the list of ranked resources (resource intensification) (Winterhalder and Kennett 2006).
The boundary that separates optimal diet resources above the line from those that are always passed by because they fall below the line plays the central role in DBM explanations of initial plant domestication. Based on their low energy value and high processing costs, small-seeded plants and those species having underground storage organs, including the wild ancestors of many domesticates, are judged to fall below the optimum diet line of human societies world-wide (Gremillion et al. 2014). As a result, under the rules of the DBM, these low ranking species, including the wild ancestors of many that would become domesticated, will enter the diet of foraging populations only when the optimum diet boundary line has been pushed down the ranked resource list to include them. The unidirectional causal arrow in DBM explanations of initial domestication begin with resource depression—an imbalance between available biotic resources and human energy demand, due either to a reduction in the carrying capacity of resource catchment areas, or to an increase in human population density. The adaptive response by human groups to resource depression in turn involves resource intensification and the downward movement of the “optimal diet” boundary line far enough to include the wild ancestors of eventual domesticates.
The alternative NCT-based explanation for initial domestication
Developed out of macroevolutionary theory in the mid-1980s, niche construction theory presents a major reconceptualization of adaptation and its role in natural selection. In contrast to the default SET unidirectional or asymmetrical definition of adaptation, Lewontin (1983) proposed that organisms do not simply respond to the environment but in fact interact with and modify their surroundings—they actively engineer ecosystems, and they shape their own niches. Odling-Smee et al. (2003) greatly expand on Lewontin’s original proposal, and argue that niche construction is universal and should be regarded, along with natural selection, as a second major participant in evolution: “There are in fact two logically distinct routes to the evolving match between organisms and their environments: either the organism changes to suit the environment, or the environment is changed to suit the organism.” (Odling-Smee et al. 2003: 18). NCT recognizes the capacity of organisms to shape their own evolutionary trajectories through engineered enhancement of their surrounding environment. Adaptation is not always in response to disequilibrium in local environments. Niche construction involves an organism altering the relationship between itself and its ecosystem, and such modification can provide individuals and populations with an evolutionary advantage. Modification of local environments, and associated selective pressures, can increase the survival chances of subsequent generations of their species: “Niche construction by organisms significantly modifies the selection pressures acting on them, on their descendants, and on unrelated populations,” and as a result, “niche constructing organisms frequently influence their own evolution by modifying their own selective environments” (Odling-Smee et al. 2003, p. 2).
Based on this symmetrical and balanced perspective on adaptation, an NCT-derived explanation of initial domestication does not cast small-scale foraging societies as, of necessity, adaptively responding to an imbalance between energy supply and demand, but rather argues that domestication emerged out of broad-scale human efforts at ecosystem enhancement that took place in the absence of any form of disequilibrium or resource depression. Based on the wealth of archaeological, ethnohistorical, and present-day accounts of ecosystem management by small-scale societies world-wide, the NCT theory (Smith 2007b, 2012, 2015a; Zeder and Smith 2009; Zeder 2012) proposes that the foraging societies that first brought plants and animals under domestication had established small central place settlements of a dozen or so household units that were situated in resource-rich ecosystems (e.g., river floodplain corridors and lake and marsh/estuary margins) that emerged during the early and middle Holocene in many regions of the world. Ranging outward from their central settlements, these groups established and maintained resource catchment zones that included a high density and diversity of plant and animal resources. Sophisticated traditional resource management systems were developed, refined, and passed down, generation to generation, through cultural inheritance. Within these stable resource catchment zones and over many generations, a broad range of plant and animal components of biotic communities were comprehensively “auditioned” to assess their potential both for sustained economic utilization and as targets of niche construction. Of the wide range of species that were the subject of varying degrees and forms of trial-and error experimental manipulation and life-cycle intervention, many were identified as low-value candidates for enhancement while others with economic utility responded in ways that encouraged and rewarded additional investment of human capital. The positive feedback loops and contexts of “reciprocal causation” (Laland et al. 2012, 2013) that formed between small-scale societies and some members of this latter species group resulted in important and sustained traditions of management of essentially ‘‘wild’’ populations, while others led to domestication.
Comparing SET and NCT-derived explanations of initial domestication
While higher-level evolutionary theory represents an excellent source for alternative hypotheses regarding domestication, suggestions that such explanations also draw support and explanatory power from the overarching evolutionary theory from which they are derived (e.g. Gremillion et al. 2014) are misplaced. Once formulated, alternative explanations or hypotheses derive no predictive power or explanatory status from their source, but rather are expected to stand on the strength of available evidence alone (Salmon 1963, 1967; Salmon 1975, 1976; Smith 1977, 2006a, 2015a). Determining the relative strength and utility of SET and NCT-derived explanations of initial domestication is based entirely on how well they are supported or contradicted by currently available and relevant archaeological and paleoenvironmental evidence.
The eight or more centers of plant and animal domestication that have been identified world-wide (Bar-Yosef and Price 2011) comprise an excellent set of independent case study opportunities to compare SET and NCT-based explanations for initial domestication in terms of how well they are supported by available data sets. Although formal sets of observational predictions for such alternative explanations been developed and tested (Smith 2015a), it is also possible to compare the relative strength of SET and NCT-based explanations in a more informal fashion, given their quite different perspectives on domestication.
If, for example, following the “default” unidirectional adaptation assumption of SET, domestication was an adaptive response to environmental disequilibrium, then the initial appearance of domesticates in the archaeological record should occur subsequent to and in relatively close temporal proximity to evidence of resource depression—an imbalance between the carrying capacity of local ecosystems and the energy demands of human populations. On the supply side, archaeological markers of resource depression would include paleoenvironmental indications of a reduction in carrying capacity in the local ecosystem—a decline in abundance and energy value of the plant and animal species that could potentially be utilized by human populations. This could be either as a result of human societies occupying marginal environments, or due to ecosystem change. On the demand side, archaeological markers of resource depression can include evidence of increased human population density, as reflected in the number and size of settlements, as well as changes in the composition of plant and animal assemblages recovered from settlements that indicate increased harvesting pressure on higher ranked species (e.g. changes in age profiles of prey species reflecting increased predation pressure). Indications of a simple increase in the number and variety of species utilized by human populations, including those that might be considered to have lower energy value, in the absence of evidence for a concomitant overharvesting of top-ranked species, would not necessarily indicate resource depression, since such an increase in species diversity could reflect a number of other changes in subsistence strategies (Zeder 2012).
In contrast, if domestication was not an adaptive response to resource depression, but rather occurred within a low-stress, broader behavioral context of increased efforts by small-scale human societies generally to modify environments in order to increase the abundance and reliability of those species of importance to them, it should be reflected by the occurrence of a quite different set of archaeological and paleoenvironmental markers. The most obvious markers would reflect the absence of any evidence of resource depression on either the supply side or demand side of the energy equation. The earliest evidence of domesticates should be found in settlements situated in environmental settings having relatively high carrying capacity, rather than in marginal settings, and there should be a lack of evidence of any preceding environmental downturn. Human settlements should be relative small in size and few in number, reflecting an absence of upward population pressure on local carrying capacity. Plant and animal assemblages from settlements should not indicate evidence of increased harvest pressure on high-ranking resources. Markers of the establishment of permanent resource catchment areas should be present, including increasing sedentism and longer time spans of settlement occupation, as well as evidence of corporate “ownership” of landscapes (e.g. cemeteries, ceremonial structures, burial mounds, etc.). In addition, if domestication occurred as part of a broader context of increased management of resources, there should be evidence of modification and enhancement of other, non-domesticate, biotic components within resource catchment areas.
Archaeological and paleoenvironmental markers providing support for alternative SET and NCT-based general explanations for initial domestication, however, are not always easily observed in the archaeological record. Accurately estimating human population levels based on the number and size of identified settlements within a particular region, for example, are complicated by establishing which settlements were actually occupied contemporaneously. Similarly, based on interpretation of archaeobiological assemblages it is often difficult to accurately establish the relative abundance of different components of biotic communities within resource catchment zones, and how heavily they were being harvested by human societies,. At the same time, evidence of human manipulation and enhancement of target resources is often difficult to identify in the archaeological record, since modification of ecosystems by small-scale societies invariably leaves a light footprint on the landscape, and patterns of human niche construction often closely mimic natural processes (Smith 2015b). The quantity and quality of available and relevant empirical data for comparing SET- and NCT-based explanations for initial domestication also varies considerably for different world regions, based on the level of archaeological and paleoenvironmental research that has been carried out. While rapidly accelerating research in many of the less well documented centers of independent domestication world-wide is beginning to provide information regarding the relative strength of SET and NCT explanations, and should allow more detailed comparison in the not two distant future, two regions in the Americas—eastern North America and the Neotropics of northern South America—have been the focus of recent in-depth assessment of these alternative perspectives.
Eastern North America
In eastern North America, Gremillion (1998, 2004) applies diet breadth modeling (DBM), one of the family of optimal foraging theory models characterized as being directly derived from neo-Darwinian evolutionary theory, in an effort to determine if small-scale hunter-gatherer societies situated in the rugged eastern Kentucky uplands of the Cumberland Plateau initially added several eastern North American small-seeded crop plants into their diets at ca. 3000 BP as an adaptive response to resource depression. She concludes, however, that there is no empirical support for the explanation based on standard evolutionary theory. There is no evidence of human population pressure on resources in the upland study area, nor is there any indication of environmental or climatic deterioration (Gremillion 2004: 227) or any indication of resource depression: “There is no independent evidence in eastern Kentucky of the kind of food shortage that would make a broad-based diet including small grains economically advantageous. The patch and diet choice models thus do not explain why small grains, whether naturally available or cultivated, were exploited in the region prehistorically” (Gremillion 2004: 229). At the same time, Gremillion does present clear evidence for human niche construction that is contemporaneous with the initial appearance of eastern crop plants in the diet of Cumberland Plateau hunter-gatherer groups (Delcourt et al. 1998; Gremillion 1998: 140; 2004: 227, 229). Situated 15 miles from Gremillion’s study area, the Cliff Palace Pond pollen record documents “an increase in fire frequency, forest opening, and a shift in forest composition in favor of fire-resistant species (including oaks and chestnut) around 3000 BP,” indicating that “at least some of the major sources of plant calories—including oak and chestnut trees—probably became more abundant rather than less” (Gremillion 2004: 227). Archaeobotanical assemblages from excavated sites provide additional evidence of human niche construction: “macrobotanical data suggest that the shift to a more prominent role for seed crops was accompanied by increased anthropogenic environmental disturbance”; “Ecological analysis of seed data also supports a general increase in anthropogenic habitats such as gardens and clearings near the shelter, or at least in the utilization of such habitats”; “The spatial scale of the disturbance created by agricultural clearing may have been relatively small, but was sufficient to create new habitats for plants that thrive in open, disturbed areas” (Gremillion 1998).
In summary, in the time frame of ca. 3000 BP, when hunter-gatherer societies in the Cumberland Plateau of eastern Kentucky first added domesticated plants into their diet, Gremillion (1998, 2004) finds no evidence of environmental disequilibrium or resource depression, but does offer empirical evidence for the deliberate and sustained human modification of the forest environment in a manner that would have increased the abundance and predictability of food resources. The Cumberland Plateau thus provides no support for an explanation based on the default SET assumption of unidirectional adaptation, while one based on niche construction theory—that domesticates were added to the diet within a broader context of general efforts by upland small-scale societies to enhance local environments and increase the productivity and predictability of food resources, finds some support in the archaeological and paleoenvironmental records of the region: “such forest opening did in fact occur around 3000 BP in eastern Kentucky as a result of burning, perhaps by human populations to increase yields of food resources” (Gremillion 2004: 229).
To the west of Gremillion’s marginally situated Cumberland Plateau study area, in the mid-latitude interior riverine region where indigenous eastern North American seed plants, including squash (Cucucbita pepo ssp. ovifera), sunflower (Healianthus annuus), marshelder (Iva annua), and chenopod (Chenopodium berlandieri) were initially brought under domestication, available empirical evidence also indicates that niche construction theory provides a better explanatory framework for considering the process of domestication than standard evolutionary theory. Here, within broad Middle Holocene Oak-Savanna and Oak-Hickory forest zones (Delcourt and Delcourt 1981) four small settlements in Illinois, Missouri, and Tennessee (Napoleon Hollow, Riverton, Phillips Spring, Hayes) dating to 5000–3800 BP and situated along resource-rich first through third order tributary river valley corridors of the Mississippi River catchment have yielded the earliest evidence of domesticated plants and the transition from hunting and gathering to low-level food production economies (Smith 2006b; Smith and Yarnell 2009).
Located in the Wabash River valley in Illinois and dating to 3800 BP, Riverton is the best documented of these sites, and in addition to providing the earliest evidence of the emergence of a crop complex including at least five domesticated seed plants (Smith and Yarnell 2009), it also represents an excellent society-scale case study example of the general environmental and cultural contexts within which initial plant domestication and the formation of a crop complex occurred in ENA. Judging from prepared clay house floors and associated feature clusters, Riverton was a settlement of perhaps a half dozen extended family units and was occupied on a semi-permanent to permanent basis over several hundred years or more. The subsistence economy of the Riverton settlement was primarily based on wild species, including a wide range of aquatic resources including fish, bivalves and snails, with the white-tailed deer (Odocoileous virginianus) being the most important terrestrial prey species, followed by lesser reliance on a suite of smaller species (e.g. turkey, raccoon, rabbits, raccoon, and squirrels). Nut and mast harvests from oaks, hickory and walnut dominate the plant assemblage, with a lesser reliance on the seeds of wild annual seed-bearing plants (Winters 1969; Smith and Yarnell 2009).
Riverton and other similar Middle Holocene small-scale societies were small in size, exhibited little internal status differentiation, and maintained stable resource-catchment areas and semi-permanent-to-permanent base camp settlements tethered to resource-rich river valley environments of the Oak-Savanna and Oak-Hickory forest zones of the East. Although the number of river valley settlements increases in the late Middle Holocene and corporate cemeteries and deep midden deposits reflect the establishment of long-term human utilization and ownership of sections of river valley corridors and adjacent uplands (Smith 1986; Marquardt and Watson 2005; Charles and Buikstra 1983), there does not appear to be much, if any, evidence that environmental disequilibrium and resource depression played a causal role in either the initial domestication of eastern seed plants or their coalescence into an initial crop complex. In areas where extensive archaeological surveys have documented the size and spacing of Late Archaic river valley settlements with a long history of occupation, resource-catchment zones are often quite substantial. The three river valley settlements identified in the Wabash Valley (Riverton, Swan Island, and Robeson Hills), for example, are spaced at 10-mi intervals, and resource-catchment area estimates for all three settlements is 500 mi2 (Winters 1969).
In summary, current archaeological and paleoenvironmental evidence offers no support for explanations of initial domestication in eastern North America that are derived from standard evolutionary theory and based on the assumption of unidirectional adaptation, environmental disequilibrium and resource depression, including Gremillion’s recent OFT/DBM-based analysis. There is, on the other hand, considerable evidence that initial domestication of plants in the region occurred within a broader context of human niche construction and the stable, long-term control and maintenance of resource catchments within resource-rich environmental settings. In addition, the initial domestication of plants in eastern North America and the associated emergence of low-level food production economies do not appear to have marked an abrupt developmental break; rather, they appear to have represented an integrated additive expansion and enhancement of preexisting hunting and gathering economies (Crawford 1982).
The Neotropics
The most prominent recent effort to employ “default” standard evolutionary theory and the assumption of unidirectional adaptation to explain initial domestication also involves optimal foraging theory and diet breadth modeling, and focuses on the Neotropics of northern South America. Piperno (2006, 2011), identifies two oval areas in northern South America as being potential centers of initial domestication, with one of these (Oval D1) specified as a likely area of domestication for four important lowland crop plants (sweet potato—Ipomoea batatas, squash—Cucurbita moschata, arrowroot—Maranta arundinacea, and achira—Canna edulis), and a possible area of domestication for three more (sieva bean—Phaseolus lunatus, yautia—Xanthosoma saggitfolium, and lerén—Calathea edulis (Piperno 2006: 154; 2011, Figure 1).
Piperno proposes that the late Pleistocene to early Holocene transformation of vegetation communities in northern South America, and an associated environmental disequilibrium, set the stage for initial plant domestication. During the late Pleistocene, from about 20,000 BP to 11,000–10,500 BP, northern South America, including the D1 Oval, is described as being covered by savanna/thorny scrub vegetation communities that “probably contained dense associations of dry-land cacti and legumes (e.g., Opuntia, Prosopis, Agave), which represented an appreciable high-quality and low-cost, edible biomass, with little cost of processing” (Piperno 2006: 151). These open savanna/thorny scrub landscapes also “undoubtedly were homes to many of the more than 30 genera of now extinct, large and medium-sized grazers and browsers… and hunting in drier and more open areas was probably a profitable pursuit” (Piperno 2006: 149).
Between about 11,000 and 10,500 BP, however, at the Pleistocene–Holocene transition, “the climate rapidly turned warmer and wetter, and elements of seasonal tropical forest moved from their glacial locations and began to replace most of the savanna/thorny scrub floristic associations,” and by “about 10,000 to 9000 BP, depending on the region, paleoecological records indicate that where a Pleistocene landscape had supported savanna-like vegetation, species rich, seasonal tropical forests now flourished” (Piperno 2006: 152). Soon after the decline in high-ranking resources, particularly Pleistocene megafauna, associated with the Pleistocene–Holocene transition, a variety of different species of plants are identified as being initially brought under domestication. The shift from a “particularly faunal rich” late Pleistocene savanna/thorny scrub landscape that supported a variety of megafauna prey species, to an early Holocene seasonally dry tropical forest in which “game animals are few in number” and small in size, and “forest plants are poor in calories and widely dispersed in space” (Piperno 2006: 142–144), soon followed by plant domestication, closely follows the default SET concept of unidirectional or asymmetrical adaptation—that initial domestication of plants in northern South America was an adaptive response to environmental disequilibrium and resource depression:
Changes in return rates of a sufficient magnitude likely to elicit new adaptations can be associated with major, natural changes to the environment, as oscillating climate and vegetation bring changes in resource density and distribution and necessitate a series of new options for humans with regard to the availability, exploitation, and procurement of plants and animals (Piperno 2006: 146).
…the single most important factor driving subsistence changes after the close of the Pleistocene probably was the dramatic decline in foraging return rates associated with the demise of glacial-period resources and expansion of forests into regions where open land vegetation had prevailed during glacial times. The removal of many mega- and large- to medium-sized fauna from a resource set and the need to practice foraging full-time in a tropical forest would immediately force subsistence options in the direction of lower-ranked resources and substantially broaden the diet breadth. Following the diet breadth model, people would have started to cultivate some plants as soon as the net return from subsistence strategies involving plant propagation exceeded those resulting from full-time foraging (Piperno 2006: 152).
This explanatory framework has been called into question, however, based on the timing of megafaunal extinctions, the initial colonization of interior northern South America, and the environmental context of the D1 Oval (Smith 2015a). Recent research indicates that in all likelihood Pleistocene megafauna had disappeared from northern South America long before Paleoindian hunter-gatherer groups first arrived. Barnosky and Lindsey’s (2010) analysis of megafaunal extinctions, environmental change, and human arrival throughout South America places the last well-dated occurrence of megafauna (species weighing more than 44 kg) in northern South America at 15,000 BP, a full 3000 to 4000 years earlier than the appearance of Pleistocene hunter-gatherers in the interior of northern South America, and 5000 years before the first evidence of domesticated plants: “the dates analyzed suggest extinction intensity and timing may have varied across the South American continent, starting in the north long before humans ever arrived” (Barnosky and Lindsey 2010: 20, fig. 8a).
These gaps of 3000 and 5000 years that respectively separate the last occurrence of megafauna in northern South America at 15,000 BP from the initial arrival of late Pleistocene hunter-gatherers the interior at 12,000–11,000 BP (Aceituno et al. 2013), and the earliest evidence of plant domestication at ca. 10,000 BP, represent clear empirical evidence that directly contradicts the proposed DBM explanation derived from standard evolutionary theory and based on the assumption of unidirectional adaptation. Late Pleistocene hunter-gatherer societies in northern South America did not rely upon the megafauna and high-quality, low-cost, legumes and cacti of open savanna/thorny scrub landscapes, and the Pleistocene–Holocene transition to seasonally dry tropical forests did not represent resource depression that caused an adaptive response in the form of initial domestication by small-scale human societies in the region.
Piperno’s proposed externalist explanation for initial plant domestication in northern South America is further contradicted by the environmental setting of the hypothesized D1 oval center of domestication. It is not situated within the postulated savanna/dry scrub environmental zone, but to the west of it, where it encompasses several inter-Andean river valleys that supported dry seasonal forests during the late Pleistocene. There are no modern analogs for the Pleistocene/Holocene forests of these inter-Andean river valleys, which contained both low- and high-elevation species, but they are considered to be generally similar to modern dry seasonal tropical forests in terms of animal biomass and relative abundance of plant species of value for human foragers (Gnecco 2003: 14; Gnecco and Aceituno 2006: 91–92; Gnecco and Mora 1997). Based on the similarity in the late Pleistocene and early Holocene forest ecosystems of these inter-Andean river valleys, Aceituno et al. (2013: 31) conclude: “the Pleistocene/Holocene transition was not a dramatic period that required costly adaptive adjustments.”
In their comprehensive assessment of the archaeological evidence for the initial human colonization of northwest South America, Aceituno et al. (2013) conclude that the first settlement of the inter-Andean river valleys encompassed by the D1 Oval occurred at about the same time as the early evidence of forest management by foragers in Panama (ca. 11,000–10,000 BP), based on numerous well-dated sites: “The increase in the archaeological record starting at the Pleistocene/Holocene transition is associated with an expansion of human groups along the river valleys that cross the Cordilleras of the northern Andes” (Aceituno et al. 2013: 31).
On the same time horizon that the inter-Andean river valleys of northern South America were being colonized, hunter-gatherer societies in Panama were actively modifying their tropical forest environments:
Pleistocene hunters and gatherers were not passive actors in their landscape….
Here, an anthropogenic disturbance and fire horizon appears suddenly at ca. 11,050 BP. This horizon is characterized by massive increases in particulate carbon and the appearance of pollen and phytoliths from plants of forest gaps, many of which show signs of direct burning and may indicate cultural maintenance of forest clearings (Piperno et al. 1991a: 213).
The La Yeguada forests were occupied and modified between 11,000 and 10,000 BP, well before any signs of agriculture in the region. The disturbance patterns here, high and sustained levels of charcoal and invasive taxa…point to exploitation of forests for their subsistence resources (Piperno et al. 1991a: 218).
Over a nearly 11,000-year period, habitat modification, apparently accomplished mainly with the use of fire, was pervasive and systematic (Piperno et al. 1991b: 247).
Building on this compelling evidence for pervasive human niche construction in Panama at the Pleistocene–Holocene transition, Gnecco (2003) outlines an explanatory framework for initial domestication of plants in the inter-Andean valley D1 oval area that directly counters the SET-derived resource depression scenario, and incorporates the basic perspective and principles of niche construction theory. Arguing against the concept of unidirectional adaptation, the assumption that “culture is essentially passive, waiting for environmental changes to start working” and “the stereotype of hunting-gathering as an explotative, nontransformative strategy,” he argues instead that initial domestication occurs within a broader context of niche construction: “early hunter-gatherers were already impacting the environment in the Neotropics through forest clearing, burning, and cultural selection of key vegetal resources,” and he cites evidence of “humanly induced forest disturbance and resource manipulation and intervention since the late Pleistocene” (Gnecco 2003: 13–14).
Based on information from inter-Andean river basin sites, Gnecco argues that late Pleistocene and early Holocene foragers in northern South America “not only gathered and hunted but … also altered to their benefit the natural productivity of resources” (Gnecco 2003: 14). Clearing of over-story canopy created “a space open enough for allowing the growth of pioneer species” and allowed, “prior to domestication and fully established agriculture… the artificial concentration of useful, otherwise dispersed plants. The artificial concentration of favored species may have required planting and tending, including forest clearing, and weeding,” and that “such forest clearing and/or tending not only favored useful plant species but also animals” (Gnecco 2003: 14–15). Gnecco concludes that “Evidence from the neotropics indicates early human management of vegetal and, likely, animal resources by 11,000 BP, including forest clearing or utilization and maintenance of natural openings by burning, and the cultural selection of useful species through protection and planting” (Gnecco 2003: 19, see also Gnecco and Aceituno 2006: 92–93).
In summary, there is no current archaeological or paleoenvironmental evidence in support of the SET-derived DBM explanation of initial domestication in the Neotropics based on the assumption of environmental disequilibrium, resource depression, and unidirectional adaptation. At the same time, there is strong support for the NCT-based alternative explanation in the form of abundant evidence and compelling synthesis arguments for the initial domestication of plants having taken place within a larger context of extensive human niche construction and broad-based management and enhancement of tropical forest ecosystems.
Discussion
In the two world regions where SET and NCT-based explanations for initial domestication have been directly compared—eastern north America and the Neotropics, the proposed explanatory frameworks that are derived from standard evolutionary theory and based on the assumption of unidirectional adaptation find no support in currently available archaeological and paleoenvironmental data sets. Explanations incorporating niche construction theory, in contrast, gain considerable support from currently available empirical evidence, while also redirecting research toward relevant variables that are not considered in default SET efforts to explain initial domestication.
Moving beyond the limited externalist focus that is inherent in SET-based explanations—that environments change and humans adaptively respond, an NCT perspective explicitly recognizes initial domestication as a lengthy evolutionary process that involves the complex interplay of a diverse array of environmental and cultural variables and networks or reciprocal causality: “the niche-construction perspective is heuristically valuable: it draws our attention to a range of phenomena that are both important and easily to overlook on standard perspectives.” (Laland and Sterelny 2006: 1760)
A central tenet of the niche construction theory of initial domestication is that human societies first brought plants and animals under domestication in the absence of environmental disequilibrium or “resource depression” on either the supply side or the demand side of the energy capture equation. Initial evidence of domesticates should be found not in marginal environments, or within the context of high human population density, but rather in favorable climatic and environmental situations and low human population levels. More specifically, domestication should take place in settings where short- and long-term climatic fluctuation is limited enough, and biotic communities are rich enough and varied enough, to allow small-scale human societies to establish small semi-permanent to permanent settlements that can be sustained over long periods of time. Essential to the successful creation of such small sedentary settlements that can be occupied over the long-term is their central placement within surrounding resource-rich catchment zones. The resource catchment zones around these settlements should encompass rich and varied biotic communities capable of providing the annual energy and raw material requirements of the human occupants of the zones. The small-scale societies that established and maintained these resource rich catchments zones and occupied centrally placed semi-permanent to permanent settlements, like a wide range of similarly sized and situated present-day and historically described groups (Smith 2011, 2012, 2015a), developed and maintained a large corpus of shared traditional ecological knowledge (TEK) regarding the management and enhancement of those target species that were of economic value to them. This corpus of knowledge was passed down from generation to generation in a variety of ways—stories, myths, rituals, and one-on-one inter-generational real world information transfer. Access to resources within the surrounding catchment zone was corporately held in varying degrees, with this corporate “ownership” of resources within a defined and maintained area periodically confirmed and reinforced both through ceremonies of social integration and through corporate labor projects such as the construction of permanent landscape modifications in the form of group burial mounds or ceremonial structures. Central to the successful development and maintenance of corporate management and enhancement of important biotic resources would have been the restructuring of the social fabric and rules of interaction and cooperation within these small-scale societies, particularly in regard to responsibility for, and access to, different resources.
Closer into the actual process of bringing various species of plants and animals under domestication, these small-scale societies, like the present-day and historically described counterparts, would have been auditioning and experimenting with a wide range of different species of plants and animals over long periods of time, looking for ways of manipulating the life cycles and habitats of economically important species in order to enhance their yield and reliability.
If the niche construction theory of domestication provides a reasonable fit with reality, it is within this emerging social and environmental context that small-scale societies in different world regions first brought plants and animals under domestication. A complex set of inter-related variables were involved—establishing small, long-term settlements and associated corporately managed resource catchment zones in situations of relatively stable and resource-rich biotic communities, developing and passing from generation to generation sophisticated knowledge about maintenance and enhancement of target resources, developing new structures of social integration and interaction, particularly in regard to rules of resource responsibility and access, and continual auditioning of and experimentation with a wide range of different species of plants and animals, some of which developed into domesticates.
I should emphasize that as outlined above, the NCT theory of domestication is not intended as any sort of definitive and comprehensive solution to this complex evolutionary transition. Rather it is an attempt to identify what general form such proposed solutions or explanations should take, and what kinds of information may be relevant to gaining a better understanding of how and why small-scale human societies in many different world areas initially brought plants and animals under domestication. Rather than starting from a default a priori assumption derived from SET that initial domestication represents an adaptive response to environmental disequilibrium, the NCT perspective encourages researchers to redirect their focus toward consideration of how a wide variety of different environmental and cultural variables formed networks of reciprocal causation. Such efforts to gain a better understanding of this major evolutionary transition are best undertaken at a regional scale of analysis (Zeder and Smith 2009), and will involve the recovery and assessment of a wide range of different classes of environmental and archaeological data, and the continuing development, refinement, and testing of innovative new explanatory frameworks.
Notes
The optimal foraging theory (OFT) family of models is often characterized as being nested within a hierarchy of higher-level evolutionary theory (e.g. behavioral ecology, evolutionary ecology, and neo-Darwinism), from which it draws epistemological support and justification (e.g. Gremillion et al. 2014). It is also identified as comprising a robust, well established, and standard set of principles in biology (Piperno 2006, 2011). Rather than being derived from and supported by higher-level evolutionary theory, however, the concept of optimization (and OFT) was first introduced into biology from microeconomics in the mid-1960 s as a still-to-be-tested hypothesis (MacArthur and Pianka 1966). Over the past 50 years it’s value and viability has frequently been questioned in biology and across a wide range of the behavioral and social sciences (see Smith 2015a for an extended discussion).
References
Aceituno FJ, Loaiza N, Delgado-Burbano M, Barrientos G (2013) The initial settlement of Northwest South America during the Pleistocene–Holocene transition: synthesis and perspectives. Quatern Int 30:23–33
Anderson MK (2005) Tending the wild. University of California Press, Berkeley
Baronsky AD, Lindsey E (2010) Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quatern Int 217:10–29
Bar-Yosef O, Price TD (eds) (2011) The origins of agriculture: new data, new ideas. Curr Anthropol 52(Special Supplement 4)
Binford L (1971) Post Pleistocene adaptations. In: Struever S (ed) Prehistoric agriculture. Natural History Press, Garden City, pp 313–341
Charles D, Buikstra J (1983) Archaic mortuary sites in the central Mississippi drainage: distribution, structure, and behavioral implications. In: Phillips J, Brown J (eds) Archaic hunters and gatherers in the American midwest. Academic, Orlando, pp 117–145
Cohen M (1977) The food crisis in prehistory: overpopulation and the origins of agriculture. Yale University Press, New Haven
Crawford G (1982) Late Archaic plant remains from west-central Kentucky: a summary. Midcont J Archaeol 7:205–224
Delcourt P, Delcourt H (1981) Vegetation maps for eastern North America: 40,000 YR B.P. to the present. In: Roman R (ed) Geobotany II. Plenum, New York, pp 123–165
Delcourt P, Delcourt H, Ison C et al (1998) Prehistoric human use of fire, the eastern agricultural complex, and Applachian Oak-Chestnut Forests. Am Antiq 63:263–278
Deur D, Turner N (eds) (2005) Keeping it living: traditions of plant use and cultivation on the Northwest Coast of North America. University of Washington Press, Seattle
Gnecco C (2003) Against ecological reductionism: late Pleistocene hunter-gatherers in the tropical forests of northern South America. Quatern Int 109–110:13–21
Gnecco C, Aceituno F (2006) Early humanized landscapes in Northern South America. In: Morrow J, Gnecco C (eds) Paleoindian archaeology: a hemispheric perspective. University of Florida Press, Gainesville, pp 86–104
Gnecco C, Mora S (1997) Early occupations of the tropical forest of northern South America by hunter-gatherers. Antiquity 71:683–690
Godfrey-Smith P (1996) Complexity and the function of mind in nature. Cambridge University Press, Cambridge
Gremillion K (1998) Changing roles of wild and cultivated plant resources among early farmers of eastern Kentucky. Southeast Archaeology 17:140–157
Gremillion K (2004) Seed processing and the origins of food production in Eastern North America. Am Antiq 69:215–233
Gremillion K, Barton L, Piperno D (2014) Particularism and the retreat from theory in the archaeology of agricultural origins. Proceedings of the National Academy of Sciences USA 111:6171–6177
Laland K, Sterelny K (2006) Perspective: seven reasons (not) to neglect niche construction. Evolution 60:1751–1762
Laland K, Odling-Smee J, Feldman M et al (2009) Conceptual barriers to progress within evolutionary biology. Found Sci 14:195–216
Laland K, Odling-Smee J, Hoppitt W, Uller T (2012) More on how and why: cause and effect in biology revisited. Biol Philos. doi:10.1007/s10539-012-9335-1
Laland K, Odling-Smee J, Hoppitt W, Uller T (2013) More on how and why: a response to commentaries. Biol Philos 28:793–810
Laland K, Uller T, Feldman M, Sterelny K, Muller G, Moczek A, Jablonka E, Odling-Smee J (2014) Does evolutionary theory need a rethink? Point yes, urgently. Nature 514:161–164
Lepofsky D (ed) 2009 Indigenous resource management: past, present, and future. J Ethnobiol 29
Lewontin R (1983) Gene, organism, and environment. In: Bendall DS (ed) Evolution from molecules to men. Cambridge University Press, Cambridge, pp 273–285
MacArthur R, Pianka E (1966) On optimal use of a patchy environment. American Naturalist 100:603–609
Marquardt W, Watson PJ (eds) (2005) Archaeology of the Middle Green River Region, Kentucky. Florida University Press, Gainesville
Mayr E (1961) Cause and effect in biology. Science 134:1501–1506
Odling-Smee J, Laland K, Feldman W (2003) Niche construction. Monographs in population biology, vol 37. Princeton University Press, Princeton
Piperno D (2006) The origins of plant cultivation and domestication in the Neotropics: a behavioral ecological approach. In: Kennett D, Winterhalder B (eds) Behavioral ecology and the transition to agriculture. University of California Press, Berkeley, pp 137–166
Piperno D (2011) The origins of plant cultivation and domestication in the New World tropics. In: Bar-Yosef O, Price TD (eds) The origins of agriculture: new data, new ideas. Curr Anthropol 52(Special Supplement 4):453–470
Piperno D, Bush M, Colinvaux P (1991a) Paleoecological perspectives on human adaptation in Panama I. The Pleistocene. Geoarchaeology 6:201–226
Piperno D, Bush M, Colinvaux P (1991b) Paleoecological perspectives on human adaptation in central Panama II. The Holocene. Geoarchaeology 6:227–250
Richerson P, Boyd R, Bettinger R (2010) Was agriculture impossible during the pleistocene but mandatory during the Holocene? A climate change hypothesis. Am Antiq 66:387–411
Salmon W (1963) Logic. Prentice Hall, Englewood Cliffs
Salmon W (1967) The foundation of scientific inference. University of Pittsburgh Press, Pittsburgh
Salmon M (1975) Confirmation and explanation in archaeology. Am Antiq 40:459–476
Salmon M (1976) “Deductive” vs. “inductive” archaeology. Am Antiq 41:376–380
Scott-Phillips T, Laland K, Shuker M, Dickins T, West S (2013) The niche construction perspective: a critical appraisal. Evolution. doi:10.1111/evo.12332
Smith BD (1977) Archaeological inference and inductive confirmation. Am Anthropol 79:589–617
Smith BD (1986) The archaeology of the southeastern United States: from Dalton to Desoto (10,500 B.P.–500 B.P.). In: Wendorf F, Close A (eds) Advances in world archaeology, vol 5. Academic Press, Orlando, pp 1–92
Smith BD (2001) Low level food production. J Archaeol Res 9:1–43
Smith BD (2006a) Human behavioral ecology and the transition to food production. In: Kennett D, Winterhalder B (eds) Behavioral ecology and the transition to agriculture. University of California Press, Berkeley, pp 289–303
Smith BD (2006b) Eastern North America as an independent center of plant domestication. Proc Nat Acad Sci 103:12223–12228
Smith BD (2007a) The ultimate ecosystem engineers. Science 315:1797–1798
Smith BD (2007b) Niche construction and the behavioral context of plant and animal domestication. Evol Anthropol 16:188–199
Smith BD (2011) General patterns of niche construction and the management of ‘wild’ plant and animal resources by small-scale pre-industrial societies. Phil Trans R Soc B 366:836–848
Smith BD (2012) A cultural niche construction theory of initial domestication. Theoretical Biology 6:260–271
Smith BD (2015a) A comparison of niche construction theory and diet breadth models as explanatory frameworks for the initial domestication of plants and animals. J Archaeol Res. doi:10.1007/s10814-015-9081-4
Smith BD (2015b) Documenting human niche construction in the archaeological record. In: Marsden J, Guedes J, Warinner C (eds) Methond and theory in paleoethnobotany. University Press of Colorado, Boulder, pp 355–370
Smith BD, Yarnell RA (2009) Initial formation of an indigenous crop complex in eastern North America at 3800 B.P. Proc Nat Acad Sci 106:6561–6566
Smith BD, Zeder M (2013) The onset of the Anthropocene. Anthropocene 4:8–14
Williams G (1992) Gaia, nature worship, and biocentric fallacies. Q Rev Biol 67:479–486
Winterhalder B, Kennett D (2006) Behavioral ecology and the transition from hunting and gathering to agriculture. In: Kennett D, Winterhalder B (eds) Behavioral ecology and the transition to agriculture. University of California Press, Berkeley, pp 1–21
Winters H (1969) The Riverton culture. Illinois State Museum, Springfield
Wray G, Hoekstra H, Futuyma D, Lenski T, Mackay T, Schluter D, Strassmann J (2014) Does evolutionary theory need a rethink? Counterpoint no, all is well. Nature 514:161–164
Zeder M (2012) The broad spectrum revolution at 40: resource diversity, intensification, and an alternative to optimal foraging explanations. J Anthropol Archaeol 31:241–264
Zeder M, Smith BD (2009) A conversation on agricultural origins: talking past each other in a crowded room. Curr Anthropol 50:681–691
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Smith, B.D. Neo-Darwinism, niche construction theory, and the initial domestication of plants and animals. Evol Ecol 30, 307–324 (2016). https://doi.org/10.1007/s10682-015-9797-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10682-015-9797-0