
Abstract
The breeding stages of a clonally propagated crop entails several steps and can take more than five years from hybridisation till cultivar release. The Accelerated Breeding Scheme (ABS) in sweetpotato relies on the use of multiple locations at the early breeding stages to reduce the years required for field evaluation. The aim of the study was to select the best progenies based on the ABS in sweetpotato towards the development of genotypes that combine high protein content with other important agronomic traits. Botanical seeds were generated from crosses between two diverse sets of parents, crossed using a North Carolina II design. F1-progenies (n = 363) were planted in a lattice design at three distinct locations during the 2017/18 planting season. Parameters recorded included pest and disease infestation, storage root traits and yield components, root protein content and nutrient related traits. Nutrient content was determined by near-infrared spectrometry. Analysis of variance and multiple t-distribution test and best test grouping were performed. There were significant differences (p < 0.001) among the genotypes for total number of roots, total root yield, marketable number of roots and marketable yield. The severity of Alternaria blight was significantly higher at Jozini. Forty progenies were selected (12.8% selection pressure) based on agronomic performance. Using the nutrient content results, four promising orange-fleshed clones (NC12-9, NC53-11, NC55-8, NC55-2) and one cream-fleshed clone (NC51-1) were identified with high storage root protein, iron and zinc contents. NC51-1 and NC55-2 produce promising storage root yields. The ABS approach is resource-demanding in the first phase but advances progenies rapidly to the subsequent breeding phases for cultivar release.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Roots and tubers crops are of major importance to the global food system. In 2019, the global annual production was 846 Mt and consumption has been projected to reach 21.0 kg/year by 2025 (Mourtala et al. 2023). Sweetpotato (Ipomoea batatas (L.) Lam) is a major staple crop in some regions of Africa, especially Malawi (5.3 Mt), Nigeria (2.9 Mt), Uganda (2.08 Mt) and Republic of Tanzania (2.06 Mt) (FAOSTAT, 2021). Although the plant originated in Central America, sweetpotato is a major cash and food security crop in sub-Saharan Africa where farmers produce the crop all year round (Low et al. 2017). Awareness of worldwide malnutrition, and increased knowledge on sweetpotato’s nutritional limitations has placed attention on the nutritional status of sweetpotato, resulting in breeders seeking to breed high yielding cultivars with superior nutritional profiles, which are better suited for farmers and consumers (Tumwegamire et al. 2016; Saltzman et al. 2017; Mbogo et al. 2021). Likewise, in South Africa the main breeding goals now include high storage root yield (40–50 t/ha under favourable conditions), good storage root quality, medium–high storage root dry matter content (RDMC) (22–25%), pleasant taste, a high storage root ß-carotene content (RBCC) (> 90 µg/100 g) and tolerance to drought and Fusarium wilt (Laurie et al. 2017, 2022; Mphela et al. 2022).
Breeding of sweetpotato relies on recurrent cycles of recombination and selection (Grüneberg et al. 2015). The efficiency of breeding is determined by the genetic gain and the time involved in achieving genetic advance. The conventional breeding process of a clonally propagated crop can take 7 to 8 years to release a new variety, while the Accelerated Breeding Scheme (ABS), as proposed by Grüneberg et al. (2009), shortens this period to 4–5 years, achieving faster gains in desired traits by the use of multiple evaluation sites during the early breeding phase (Grüneberg et al. 2015; Mwanga et al. 2021). The ABS's implementation on sweetpotato began in 2005 to reduce lengthy breeding processes and maximize resources.
ABS is based on the general principle of breeding clonally propagated crops, whereby genetic recombination is obtained through sexual seed production followed by asexual clonal propagation (Grüneberg et al. 2015). The authors explained that the population developed from botanical seeds consists of very diverse and heterozygous genotypes, each one of which can potentially be a new variety. Time reduction begins immediately after the multiplication of botanical seeds, by testing the clones in small plots in 2 to 3 contrasting environments with different selection pressures. This differs from the conventional process in which a single-row plot of 3 to 5 plants is planted, with no replications, at a single site in the first season. In ABS, non-performing progenies in marginal or adverse environmental condition, are discarded through visual selection at the early breeding stage. Progenies are selected based on highly heritable traits such as general performance (growth type, storage root size, -shape and -colour), resistance to pests and diseases, harvest index, dry matter and nutritional quality. These selections enter directly into the advanced breeding stages, leading to a shorter breeding cycle. In this breeding scheme, all the relevant agronomic data necessary for cultivar release is available by the end of the fourth year (Grüneberg et al. 2015). However, ABS is only feasible if the genotype-by-environment interaction is relatively low (Andrade et al. 2017).
ABS has been used successfully in Mozambique, enabling the release of 14 varieties in 2011, ‘Irene’ being one of the released varieties that is now grown in several countries in sub-Saharan Africa (Andrade et al. 2017). Four of these novel varieties were recently released in Eswatini, following two seasons of extensive evaluation trials (CARDESA 2023). Several African research institutions have enhanced their sweetpotato breeding strategies since 2009. However, results from the programs are not widely published (Grüneberg et al. 2015). Focussing on the traits beta-carotene, iron, zinc contents and virus resistance, Mwanga et al. (2021) reported the release of 158 varieties resulting from ABS schemes in several sub-Saharan African countries. Of these 27 are drought-tolerant and two have enhanced levels of iron and zinc. This resulted from the emphasis on accelerated development and release of cultivars by 14 participating breeding programs in national agricultural research institutes (Low et al. 2020). South Africa also received improved botanical seed populations from the three sub-regional sweetpotato support platforms that were established in Uganda, Mozambique, and Ghana (Mwanga et al. 2021).
Sweetpotato has lower storage protein content (1.8 g 100–1 kcal) than other staple crops (USDA, 2019). Hence, there is a need to breed new sweetpotato cultivars for enhanced protein content to circumvent global protein-energy malnutrition, notably in developing countries. The present study aimed to select progenies emanating from crosses made between two sets of parents identified by Naidoo et al. (2022) using ABS at three locations in South Africa to develop genotypes which combine high protein content with other relevant agronomic traits.,
Material and methods
Plant material
Genomic DNA extracted from 31 sweetpotato accessions subjected to analysis by eight selected polymorphic SSR markers (Buteler et al. 1999; Veasey et al. 2008; Karuri et al. 2010; Gwandu et al. 2012; Naidoo et al. 2016) enabled identification of two heterotic groups, A and B, each consisting of five female and five male parental clones for hybridization to create F1 progenies. The clonal parents, five females and five males, were crossed using a North Carolina II (NCII) mating design. The parents were pre-selected according to magnitude of storage root protein content (RPC) that varied from high to low, total storage root yield (TRY), storage root dry matter content (RDMC), storage root flesh colour (RFC), their flowering ability as established from previous field evaluation, and genotyping (Laurie et al. 2020; Naidoo et al. 2020). A summary of the characteristics of the 10 parental lines are provided in Table 1. Family (‘Bophelo × 2008-3-1′) did not produce botanical seeds, therefore only 24 families (crosses) were evaluated.
Botanical seeds were obtained by manually crossing the parental clones for two consecutive planting seasons, in 2015/16 and 2016/17, in the field and glasshouse conditions at the ARC-VIMP, Roodeplaat, Pretoria (25.604° S, 28.345° E). Crosses were performed under field conditions and in the greenhouse during the summer and winter, respectively. Parental lines were planted in 5L nursery bags in the glasshouse (30/20 °C day/night) temperature and trellised onto 2 m wooden poles. In the field 10 plants per parents were planted at 1 m spacing, and elongated vines were trellised onto 2.5 m wooden poles, using plastic twine. During the crossing period, individual flowers were closed at 2.00 pm using paper clips. Hand pollinations were performed the following day from 8.00 a.m. till 10.30 a.m.. Pollinated flowers were closed by tying them closed with 2 mm woolen string. Individual pollinated flowers were labelled with a paper tag that recorded the identity of the parents involved in the cross. Successful crosses produced capsules 4 to 6 weeks after hand pollination. Harvested botanical seeds were stored in brown envelopes at room temperature (24 °C).
Scarification of botanical seeds and vine multiplication
Harvested botanical seeds were scarified with 98% sulphuric acid (H2SO4) (Merck, Darmstadt, Germany) as described by Gurmu (2015). Scarified seeds were rinsed under running water for 5 min, then placed in Petri dishes lined with moistened filter paper and placed in the dark for germination. Germinated botanical seeds were planted in seedling trays filled with Hygromix commercial growing medium (Hygrotech, South Africa). Table 2 indicates the parental combinations and their respective number of seedlings.
Seedlings of approximately 10–12 cm height were transplanted to 5L nursery bags filled with an enriched soil mixture (ratio 2:1:1 soil, organic compost and Hygromix) and placed in the glasshouse for the multiplication of vines. The 10 sweetpotato parental lines were similarly multiplied by growing 4-node cuttings from the mother plant in seedling trays, and then transplanted to 5L nursery bags. Sixteen weeks later, six cuttings with 4-nodes each were taken from the parents and F1 plants for field evaluation trial at the first experimental site. The plants in the glasshouse were allowed to regrow for a period of 20 weeks to obtain further cuttings for subsequent plantings at two more sites.
Description of the experimental sites
Field evaluation of sweetpotato progenies for the ABS were conducted in three sites with distinct climatic conditions and management, namely Roodeplaat, Jozini, and Lwamondo. Descriptions of the experimental sites and environmental conditions are presented in Table 3. The first planting site was Roodeplaat, followed by Jozini and Lwamondo. Of the three locations, the Roodeplaat trial was the most intensively monitored because it is the main research site of the institute compared to Lwamondo (a sub-research station) and Jozini (a farmer’s field).
Experimental design
Roodeplaat and Jozini
The F1 hybrid progenies were established in the field using a 14 × 26 balanced lattice design with three replicates, at the two sites. The intra-row spacing was 30 cm, and between rows was 1 m. The randomised field layout was produced by CycDesigN (Whitaker et al. 2002).
Lwamondo
After preliminary observations on the growth habit and thickness of the vines under glasshouse conditions, a number of genotypes with thin, twining vines were discarded, reducing the number of progenies planted in Lwamondo to 327 genotypes. Varieties with thin twining vines have lower field survival rates. A similar plant density was used at the Roodeplaat and Jozini sites.
Trial maintenance
The fields were prepared and fertilized following the guidelines for cultivation of sweetpotato in South Africa, taking into account soil analysis results (Van den Berg and Laurie 2004). NPK (1:0:1; 300 kg/ha) and Superphosphate (500 kg/ha) were broadcasted before planting. At Jozini and Lwamondo, 2:3:4 [(30) + Zn 0.34%; 300 kg/ha] and Superphosphate (500 kg/ha) were applied before planting. Limestone Ammonium Nitrate (LAN, 28%; 120 kg/ha) was used to top dress the plants 21 days after planting. The trial at Roodeplaat and Jozini was conducted under irrigated condition, whereas the trial at Lwamondo depended on rainfall only. All field management practice was observed according to the crop’s cultivation requirements.
Harvesting, selection and data collection
Symptoms of diseases (virus, Alternaria stem, leaf and wilt blight) were scored before harvesting on the stems and leaves by visual estimation of the percentage of leaf area showing symptoms. Incidences of leaf damage due to insects were assessed by visual estimation of the percentage of leaf area affected.
After harvest, the storage roots were graded in categories of marketable roots (MRY) (200–1000 g) and unmarketable roots (URY) (< 200 g or > 1200 g or mechanically damaged) (Laurie et al. 2017). Storage roots that displayed insect damage (IDRY) were recorded separately. Total storage root yield (TRY) was obtained by combining MRY, URY and IDRY. The number of storage roots in each category was also recorded for the marketable number of storage roots (MNR), unmarketable number of storage roots (UNR) and insect damage number of storage roots (IDNR). The total number of storage roots (TNR) was obtained by adding MNR, UNR and IDNR.
Two medium-sized storage roots (150–300 g) per genotype were collected from two replicates for nutrient content determination. The storage roots were washed, peeled and cut longitudinally into quarters. Opposite quarters were pureed using an electric food processor (Philips Food Processor, 300v, France), then 100 g of blended flesh was stored at − 20 °C. The samples were freeze dried in a bench top freeze dryer (Martin Christ LDplus, Germany) at a temperature of − 45 °C for 48 to 72 h or until dried. The freeze-dried samples were manually milled using a porcelain mortar and pestle. Milled samples were sent to the International Potato Center (CIP) in Maputo, Mozambique. Nutrient quality parameters were estimated using near-infrared spectrometry (NIRS) calibrated similarly as Zum Felde et al. (2010), namely storage root ß-carotene content (RBCC), storage root protein content (RPC), iron (Fe), calcium (Ca), magnesium (Mg), zinc (Zn), fructose, sucrose, glucose and starch.
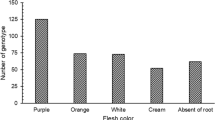
Progenies with the following traits were selected: good yield, few storage root defects (veins, constrictions, cracks), best storage root uniformity, regular shape, acceptable storage root skin and flesh colour (white, cream and orange), little insect damage, little root rot, and a good raw taste (taste scale: not sweet, edible, sweet and very sweet) and root dryness (watery, dry and very dry)].
Data analysis
Due to a large number of missing plots (as explained above for the Lwamondo site), the data was analysed using a randomised block design. Analysis of variance (ANOVA) was performed using the General Linear Models Procedure (PROC GLM) of SAS software (Version 9.4; SAS Institute Inc, Cary, USA) to compare the genotypes for yield components (TRY, TNR, MRY, UMRY, IDRY, MNR, UNR, IDNR), per site. Fischer’s Least Significance Difference (LSD) was calculated at 5% to compare genotype means (Ott and Longnecker 2010). Thereafter, the multiple t distribution test procedure of Gupta and Panchapakesan (1979, 2002) was performed to identify the best group of entries using a macro of SAS software (Version 9.4; SAS Institute Inc., Cary, USA). The principle of this procedure (hereafter called the best test) was to divide the genotypes (entries) into two groups. The best group was composed of progenies that did not differ from each other at p ≥ 0.05. The results of the best test were used to support the visual selection in terms of quantified storage root yield. Yield selection was based on the cut-off point for the lowest acceptable yield indicated by the best test. In addition, progenies were discarded on the basis of best test results for negative traits, e.g., stem blight and insect damage. Progenies were selected based on their performance at all three sites.
Results
Roodeplaat
Results from the ANOVA at Roodeplaat, Gauteng, showed significant differences (p < 0.05) among the progenies for TNR, TRY, MNR, MRY, UNR and URY (Table 4). Mean values among the progenies varied from 0.06 to 5.81 kg/plant for TRY and from 0.00 to 3.48 kg/plot for MRY. The mean MRY was 1.46 kg/plant. The best test group for TNR contained only 2 progenies, 22 progenies for TRY, 175 for MNR, and 168 for MRY (Table 4). At harvest root phenotypic selection was performed and 72 progenies were selected based on general appearance and raw taste. URY and IDRY were negative selection criteria.
The ANOVA results for the nutrient content (RBCC, RPC, Fe, Ca, Mg, Zn, fructose, sucrose, glucose and starch) estimated from storage roots sampled at Roodeplaat, are summarized in Table 5. Significant differences among progenies were obtained for all the nutritional components.
Fusarium wilt disease pressure, identified from leaf yellowing and browning of vascular tissue (Mphela et al. 2022), was low as only two progenies showed infection symptoms, namely 59 (NC14-14) and 301 (NC51-13). Alternaria leaf blight (ALB) and Alternaria stem blight (ASB) symptoms (typical dark grey necrotic lesions on stems (Narayanin et al. 2010) were noted for 75 progenies. Disease prevalence were used as a negative selection criterion. The percentage prevalence of ALB and ASB in sweetpotato progenies evaluated at Roodeplaat is presented in Fig. 1. It was seen that ASB was prevalent in 30 progenies and ALB in 45 progenies. Progeny NC35-6 showed 100% infection, while NC32-5, NC33-11 and NC35-10 had 90% infection. Progenies NC35-13 and NC53-13 had a high level (90%) of infection of both ALB and ASB.

Disease incidence (%) for 70 sweetpotato clones evaluated at Roodeplaat showing Alternaria leaf blight (ALB) and Alternaria stem blight (ASB) (M1 = male parent 1987-2-1; M2 = male parent 2008-3-1)
Jozini
There were significant differences for all variables at Jozini except URY and Alternaria leaf blight (Table 6). The mean values of TNR and TRY were 4.96 roots/plant and 0.54 kg/plant, respectively. The mean values for MNR and MRY were 1.55 roots/plant and 0.31 kg/plant, respectively. One hundred and nineteen progenies grouped with the best group for MRY (cut-off point 0.32 kg/plant) (Table 6). There were significant differences in the expression of ASB among the progenies, confirming the high disease pressure at this site. ALB was also prevalent, infecting 86 progenies of which the 63, as presented in Fig. 2. Sweetpotato clones NC14-6, NC51-8 and NC54-8 were infected with both ALB and ASB. Genotype NC14-6 showed the highest level of ASB at 40%. Progenies NC12-13, NC13-5, NC33-15, NC45-6 and NC54-9 had disease infestation of 50% for ALB.

Sweetpotato breeding clones with high disease incidence (%) of leaf and stem blight at Jozini (ALB = Alternaria leaf blight; ASB = Alternaria stem blight)
Lwamondo
Significant differences (p < 0.05) were found at Lwamondo (Table 7) among the progenies for all the traits except for MRY, MNR, IDRY, and leaf weevil damage (showing small holes in the inner part of leaves). Mean values for TNR and TRY were 7.61 roots/plant and 0.87 kg/plant, respectively.
Disease pressure was low, with only 23 clones showing symptoms of ALB and ASB. Some breeding clones developing ASB infection were: NC55-7 and NC23-10 (80%); NC22-3 (60%); NC41–6, NC25-9 and NC25-7 (50%); 366 (P3 = female parental clone), N43-2, NC35-15, NC34-5 and NC11-10 (40%); NC53-7, NC53-6, NC45-15, NC34-12 and NC34-4 (30%); NC11-11 and NC14-7 (20%). Only five breeding clones showed a high susceptibility to ALB, namely M4 (male parental clone) (80%), NC11-12 (70%), M5 (male parental clone) (50%), NC54-11 and P3 (40%). A few breeding clones were considered in addition to those selected from Roodeplaat and Jozini by using the best test results of Lwamondo for MRY, with prerequisite that the breeding clones had average yield at Jozini and Roodeplaat. These were clones NC23-7 and NC23-11.
Combined results across the three sites
Table 8 presents a summary of the 40 best performing progenies at the three sites (Roodeplaat, Jozini and Lwamondo). The TRY of the selected progenies ranged from 1.67 to 4.23 kg/plant at Roodeplaat, 0.35 to 2.56 kg/plant at Jozini and 0.04 to 2.26 kg/plant at Lwamondo. The highest TRY was recorded at Roodeplaat. The MRY varied from 0.10 to 3.48 kg/plant at Roodeplaat, 0.17 to 3.19 kg/plant at Jozini and 0 to 1.38 kg/plant at Lwamondo for the selected progenies. The highest MRY was obtained at Roodeplaat.
In addition to the agronomic traits, the nutrient content (RBCC, RPC, Fe, Ca, Mg, Zn, fructose, sucrose, glucose and starch) estimated from storage roots sampled at Roodeplaat, are presented in Table 8 for the best performing progenies. High RPC were noted for progenies NC12-9, NC51-1, NC51-6, NC54-2 and NC55-8. Both progenies NC12-9 and NC55-8 exhibited high estimates for RBCC.
For Fe, eight progenies had levels above 2% namely NC12-9 (2.22), NC13-7 (2.04), NC22-1 (2.08), NC24-13 (2.19), NC31-2 and NC32-11 both with 2.24, NC51-1 (2.12), NC51-6 (2.03), NC55-2 (2.04), NC55-4 (2.10) and NC55-8 (2.23). The highest Zn levels were recorded for NC12-9 (1.65) and NC51-1 (1.62) both with Zn levels above the control progenies (Table 8). High Ca values were observed for progenies NC12-9 (286), NC22-1 (280.5), NC24-13 (284.2), NC31-11 (268.6), and NC41-10 (271.9). Progenies NC22-1 (154.3) and NC24-13 (161.7) also had the highest Mg content.
Progenies NC33-12 (74.02), NC41-15 (75.23), NC42-3 (74.31) and NC43-6 (75.48) exhibited high starch content. For fructose and glucose, both traits had high values for progenies NC11-9 (4.11 and 8.25), NC13-11 (6.16 and 10.34), NC24-13 (5.29 and 9.86), NC31-11 (5.07 and 8.70), NC41-10 (4.47 and 8.35), NC51-1 (4.28 and 8.46) and NC53-14 (6.38 and 10.86). Four progenies had high values for sucrose NC22-1 (24.27%), NC24-13 (23.86%), NC55-3 (20.13%) and NC55-4 (21.92%).
Discussion
In the present study, highly significant differences (p < 0.001) were found among genotypes for most of the storage root yield and component traits of economic importance considered in this study across three sites in South Africa (Roodeplaat, Jozini and Lwamondo). These progenies were created by crossing five females with five males comprising of complementary traits. The main principle of breeding clonally propagated crops is to create genetic variation through sexual reproduction and generation of botanical seeds for clonal selection. The F1 families constitute new individuals and a new population that displays genetic variation (Grüneberg et al. 2015). Variability in the population that results from gene recombination in the offspring is fundamental for the success of any crop breeding program, including sweetpotato (Grüneberg et al. 2015). A novel feature of the present study is including selection for high protein content in the ABS approach that allowed speed breeding of sweetpotato. The study further used multiple t-distribution test procedures proposed by Gupta and Panchapakesan (1979, 2002) that allowed the identification of the best clones. Genotypes were selected based on storage root yield response on the cut-off point for the lowest acceptable yield response aided by phenotypic selection and a negative selection against susceptibility to stem blight and insect pest damage.
The present study provided complementary data to advance ABS to reduce the cost and time in sweetpotato breeding progarms. In the ARC-VIMP sweetpotato breeding program, cultivar release typically takes 7 to 8 years after hybridization. This breeding cycle can be accelerated by using the ABS process. This scheme potentially can shorten a breeding timeline to 4 to 5 years and optimise the use of limited breeding resources. The efficiency of a breeding method is determined by the genetic gain and the time required to breed a cultivar (Grüneberg et al. 2009). By using three sites in the agronomic evaluation of the new breeding clones generated by a 5 × 5 NCII design, a total of 72 clones were selected at Roodeplaat (20.1% selection ratio), based on phenotypic assessment. Only clones selected at Roodeplaat were considered for selection at the other sites, therefore 21 clones were selected at Jozini (5.9% selection ratio). Overall, a total of 40 progenies were selected from the three sites. These progenies were the best performing clones in each site. The skin colour of the selected progenies varied from purple to pink and orange to cream. Andrade et al. (2016) evaluated a staggering 198 592 clones followed by selection of superior clones over four sites. Selection criteria to advance to the next testing stage included storage root yield above 10 t/ha, β-carotene content (BC) > 5 mg/100 g DW, dry matter content (DM) > 200 g/kg, vine biomass above 10 t/ha, and host plant resistance to pests (including viruses and weevil). Following a series of clonal (23,839 clones), preliminary yield trials (3318 clones) and advanced yield trials (1258 clones) 23 clones were included in genotype x environment analysis, and finally 15 varieties were released. From seedling to clonal level, the selection pressure was 12% (Andrade et al. 2016). Meanwhile, de Oliveira Silva et al. (2022) applied a 2.5% selection pressure for the traits of suitability for human consumption, ethanol production, and animal feed through separate indices. Low et al. (2020) reported that the utilization of an ABS approach reduced the breeding cycle from 8–10 to 4–5 years.
In ABS, selection is effective in environments which have stressed conditions, e.g., disease infestation, which allows the selection of resistant genotypes with potential to be advanced to further agronomic and quality evaluation (Grüneberg et al. 2009). Progenies that showed high levels of disease symptoms at Roodeplaat, Jozini or Lwamondo, were discarded immediately before further tests (sensory and storability tests) and advanced field trials, even if they were high yielding. The incidence and expression of Alternaria stem and leaf blight was higher at the Jozini and Roodeplaat sites than at Lwamondo. The high disease severity in the two locations allowed clear discrimination among progenies for disease resistance. As reported by Kandolo et al. (2016), wet conditions combined with temperatures of between 20 and 25 °C are favourable for infection development and spread of Alternaria blight. This disease is one of the major constraints in the production of sweetpotato, and can cause yield losses of more than 50% in susceptible genotypes (Sseruwu et al. 2016). In a similar approach, Andrade et al. (2016) used a site, Guruè, for high virus pressure, and two sites, Umbuluzi and Chokwe, for drought tolerance selection.
Five progenies (NC12-9, NC51-1, NC51-6, NC54-2, NC55-8) had high RPC levels compared to the control cultivars, and of these, two (NC12-9 and NC55-8) had high RBCC, Fe and Zn. The selected clones will be advanced to further field testing, and evaluation of quality traits such as storability, palatability, and protein content. Progenies with high level of multiple nutrients have the potential to become important food sources to combat malnutrition in South African and Sub-Saharan Africa (SSA) communities. Due to the volumes of sweetpotato consumed in Africa, the crop can contribute to a greater intake of protein and minerals (Fe, Zn, Ca and Mg) as part of a balanced human diet, which will combat malnutrition (Tumwegamire et al. 2011).
Complimentary work by Naidoo et al. (2021) reported heritability values for the traits presented here. Broad sense heritability was high for all nutrient compositions, excluding Mg, TNR, and TRY, and intermediate for MRY and MNR. Narrow sense heritability was high for RBCC, Fe, Zn, fructose, sucrose and glucose, intermediate for starch and RPC and low for Mg and Ca (Naidoo et al. 2021). This indicates the presence of additive genetic effects playing a role in these traits and further suggests that reliable crop improvement can be achieved through phenotypic selection. Estimates of heritability with genetic advance are more reliable and meaningful than individual consideration of the traits.
Mwanga et al. (2021) reported that the process of using ABS to accelerate exploiting heterosis in vegetatively propagated crops, originated at the International Potato Center in Peru in 2009. The approach has been tested and validated in Mozambique, Uganda, Ghana and other national programs over a period of 10 years. Heterosis exploiting breeding has the potential to increase genetic gains for several traits, including storage root yield, number of commercial storage roots, virus resistance, and iron status. Low et al. (2020) added to this abiotic tolerance, host plant resistance to pests and diseases and early maturity.
Large strides can be made using hybrid populations and one cycle of reciprocal recurrent selection (Grüneberg et al. 2022). Genetic gain in the storage root yield was remarkable for H1 relative to the foundation in a population for wide adaptation and earliness (O-WAE) at 118.8% for early harvest time and 81.5% for normal harvest time, while H1 for high iron (O-HIFE) achieved 97.1%. The storage root yield traits exhibited population average heterosis increments of up to 43.5%. This shows that population hybrid breeding is a plant breeding tool that can achieve large genetic gains in sweetpotato yield because it provides for more efficient population improvement, and consequently the potential for rapid dissemination of botanical seed that is generated from reproducible elite crosses, in contrast to the time-consuming and pricy process of virus elimination in maternal material of elite clones, due to the vegetative propagation of sweetpotato.
Conclusion
ABS for clonal propagation crops such as sweetpotato is a relatively new concept that is rapidly being implemented in Sub-Saharan Africa. This approach has the potential to increase breeding efficiency of sweetpotato with regards to time, human and financial resources due to shorter breeding cycles in multiple environments. Through ABS, 40 progenies were selected for further field trial evaluation and testing of sensory and quality traits. Four progenies were identified that combined high protein content and essential minerals. ABS is resource-demanding in the first year of progeny evaluation. However, it enhances the selection process to identify the best progenies to be included in the advanced breeding phases. The selected progenies will be advanced in further field and sensory evaluations and determination of nutrient content using wet chemistry.
References
Andrade MI, Alvaro A, Menomussanga J, Makunde GS, Ricardo J, Gruneberg WJ, Eyzaguirre R, Low J, Ortiz R (2016) Alisha’, ‘Anamaria’, ‘Bie’, ‘Bita’, “Caelan’, ‘Ivone’, ‘Lawrence’, ‘Margarete’, and ‘Victoria Sweetpotato. HortScience 51(5):597–600
Andrade MI, Ricardo J, Naico A, Alvaro A, Makunde GS, Low J, Ortiz R, Grüneberg WJ (2017) Release of orange-fleshed sweetpotato (Ipomoea batatas [L.] Lam.) cultivars in Mozambique through an accelerated breeding scheme. J Agric Sci 155(6):919–929
Buteler M, Jarret R, Labonte D (1999) Sequence characterization of microsatellites in diploid and polyploid Ipomoea. Theor Appl Genet 99:123–132
CARDESA (2023) Eswatini releases orange fleshed sweet potatoes variety. https://www.ccardesa.org/eswatini-releases-orange-fleshed-sweet-potatoes-variety. Accessed 6 Jan 2023
De Oliveira Silva JC et al (2022) Mixed model-based indices for selection of sweet potato genotypes for different agronomic aptitudes. Euphytica 218:86–103. https://doi.org/10.1007/s10681-022-03033-9
FAOTSTAT. The State of Global Nutrition (2021) Available online: https://globalnutritionreport.org/reports/2021-globalnutrition-report/. Accessed 17 April 2022
Grüneberg W, Mwanga R, Andrade M, Espinoza J (2009) Breeding clonally propagated crops. In: Ceccarelli S, Guimarães EP, Weltzien E (eds) Plant breeding and farmer participation. Food and Agriculture Organization of the United Nations, Rome, p 275
Grüneberg WJ, Ma D, Mwanga ROM, Carey E, Huamani K, Diaz F, Eyzaguirre R, Guaf E, Jusuf M, Karuniawan A, Tjintokohadi K, Song Y-S, Hossain M, Rahaman E, Attalur I, Some K, Afuape S, Adofo K, Lukonge E, Karanja L, Ndirigwe J, Ssemakula G, Agili S, Randrianaivoarivony J-M, Chiona M, Chipungu F, Lurie S, Ricardo J, Andrade M, Fernandes Rausch F, Mello AS, Khan A, Labonte DR, Yencho GC (2015) Chapter 1 Advances in sweetpotato breeding from 1993 to 2012. In: Nyongesa M, Quinn S, Parker M, Low J (eds) Potato and Sweetpotato in Africa: transforming the value chains for food and nutrition security. CABI, Wallingford, pp 1–77
Grüneberg WJ, De Boeck B, Diaz F, Eyzaguirre R, Low JW, Reif JC, Campos H (2022) Heterosis and responses to selection in orange fleshed sweetpotato (Ipomoea batatas L.) improved using reciprocal recurrent selection. Front Plant Sci. https://doi.org/10.3389/fpls.2022.793904
Gupta SS, Panchapakesan S (2002) Multiple decision procedures: theory and methodology of selecting and ranking populations. In: Society for industrial and applied mathematics (SIAM) books
Gupta SS, Panchapakesan S (1979) Multiple decision procedures: theory and methodology of selecting and ranking populations. Wiley, New York
Gurmu FB (2015) Breeding of sweetpotato for improvement of root dry matter and β-carotene contents in Ethiopia. PhD thesis, University of KwaZulu-Natal. https://ukzn-dspace.ukzn.ac.za/bitstream/handle/10413/14073/Balcha_Fekadu_Gurmu_2015.pdf?sequence=1&isAllowed=y
Gwandu C, Tairo F, Mneney E, Kullaya A (2012) Characterization of Tanzanian elite sweet potato genotypes for sweet potato virus disease (SPVD) resistance and high dry matter content using simple sequence repeat (SSR) markers. Afr J Biotechnol 11:9582–9590
Kandolo SD, Aveling TAS, Van Der Waals JE, Truter M, Laurie SM (2016) Effects of wetness duration, inoculum concentration and temperature on the development of Alternaria blight on transplants of sweet potato. Acta Hort 1204:211–217
Karuri H, Ateka E, Amata R, Nyende A, Muigai A, Mwasame E, Gichuki S (2010) Evaluating diversity among Kenyan sweet potato genotypes using morphological and SSR markers. Int J Agric Biol 12:33–38
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World map of the Köppen-Geiger climate classification updated. Meteorol Z 15:259–263
Laurie S, Calitz F, Mtileni M, Mphela W, Tjale S (2017) Performance of informal market sweet potato cultivars in on-farm trials in South Africa. Open Agric 2(1):431–441
Laurie SM, Naidoo SIM, Magwaza L, Shimelis H, Laing MD (2020) Assessment of the genetic diversity of sweetpotato germplasm collections for protein content. SA J Bot 132:132–139. https://doi.org/10.1016/j.sajb.2020.03.041
Laurie SM, Bairu MW, Laurie RN (2022) Analysis of nutritional composition and drought tolerance traits of sweet potato: selection criteria for breeding lines. MDPI Plants 11:1804. https://doi.org/10.3390/plants11141804
Low JW, Mwanga ROM, Andrade M, Carey E, Ball A (2017) Tackling Vitamin A deficiency with biofortified sweetpotato in sub-Saharan Africa. Glob Food Sec 14:23–30
Low JW, Ortiz R, Vandamme E, Andrade M, Biazin B, Grüneberg WJ (2020) Nutrient-dense orange-fleshed sweetpotato: advances in drought tolerance breeding and understanding of management practices for sustainable next-generation cropping systems in Sub-Saharan Africa. Front Sustain Food Syst 4:50. https://doi.org/10.3389/fsufs.2020.00050
Mbogo D, Muzhingi T, Janaswany, (2021) Starch digestibility and β-carotene bioaccessibility in the orange- fleshed sweet potato puree-wheat bread. J Food Sci 86:901–906. https://doi.org/10.1111/1750-3841.15620
Mourtala IZM, Innocent NM, Habibou HO (2023) Recent progress in breeding beta-carotene, dry matter content and sugar in sweet potato [Ipomoea batatas (L.) Lam]—a review. Eur J Agric Food Sci 5:6–13. https://doi.org/10.24018/ejfood.2023.5.1.551
Mphela WM, Laurie SM, Bihon W, Minnaar-Ontong A (2022) Development and screening of Fusarium wilt resistant lines in sweet potato [Ipomoea batatas (L.) Lam]. Euphytica 218:68. https://doi.org/10.1007/s10681-022-03016-w
Mwanga ROM, Swanckaert J, Da Silva PG, Andrade MI, Makunde G, Grüneberg WJ, Kreuze JF, David M, De Beck B, Carey E, Ssali R, Yencho GJ, Utoblo O, Gemenet GC, Otema M, Yada B, Chelangat D, Oloka O, Mtunda K, Chiona M, Koussao S, Laurie SM, Campos H, Low JW (2021) Breeding progress for Vitamin A, iron and zinc biofortification, drought tolerance, and sweetpotato virus disease resistance in sweetpotato. Front Sustain Food Syst 5:616674. https://doi.org/10.3389/fsufs.2021.616674
Naidoo SIM, Laurie SM, Odeny DA, Vorster BJ, Mphela WM, Greyling MM, Crampton BG (2016) Genetic analysis of yield and flesh colour in sweetpotato. Afr Crop Sci J 24:61–73
Naidoo SIM, Laurie SM, Shimelis H, Laing MD (2020) Morpho-agronomical characterisation of local and international sweet potato germplasm from the South African collection. SA J Plant Soil 37:308–320. https://doi.org/10.1080/02571862.2020.1791983
Naidoo SIM, Laurie SM, Booyse M, Mphela W, Makunde GS, Shimelis H, Laing MD (2021) Combining ability, heterosis and heritability of sweet potato root protein, β-carotene, sugars and mineral composition. Euphytica 217:109. https://doi.org/10.1007/s10681-021-02829-5
Naidoo SIM, Laurie SM, Assefa A, Shimelis H, Laing MD (2022) Elucidating the genetic diversity and population structure of sweetpotato accessions using simple sequence repeat markers. MDPI Plants 11:1802. https://doi.org/10.3390/plants11141802
Narayanin CD, Thompson AH, Slabbert MM (2010) First report of Alternaria blight of sweetpotato caused by Alternaria bataticola in South Africa. Afr Plant Prot 16:7–9
Ott R L, Longnecker M (2010) An introduction to statistical methods and data analysis. Brooks/Cole Cengage Learning, Belmont, USA, 463–468 pp
Saltzman A, Birol E, Oparinde A, Andersson MS, Asare-Marfo D, Diressie MT, Gonzalez C, Lividini K, Moursi M, Zeller M (2017) Availability, production, and consumption of crops biofortified by plant breeding: current evidence and future potential. Ann N Y Acad Sci 1390:104–114
Sseruwu G, Shanahan P, Melis R, Ssemakula G (2016) Genetic analysis of resistance to Alternaria leaf petiole and stem blight of sweetpotato in Uganda. Euphytica 210(3):393–404
Tumwegamire S, Rubaihayo P, Labonte D, Diaz F, Kapinga R, Mwanga R, Grüneberg W (2011) Genetic diversity in white- and orange-fleshed sweetpotato farmer varieties from East Africa evaluated by simple sequence repeat markers. J Crop Sci 51:1132–1142
Tumwegamire S, Rubaihayo P, Grüneberg W, Labonte D, Mwanga ROM, Kapinga R (2016) Genotype x Environment interactions for East African orange-fleshed sweetpotato clones evaluated across varying ecogeographic conditions in Uganda. J Crop Sci 56(4):1628–1644
U.S. Department of Agriculture—Agricultural Research Service (USDA-ARS), 2019. USDA National Nutrient Database for Standard Reference, Release 28. https://data.nal.usda.gov/dataset/composition-foods-raw-processed-prepared-usda-national-nutrient-database-standard-reference-release-28-0
Van Den Berg AA, Laurie, SM (2004) Cultivation. In: Niederwieser, JG (ed), Guide to sweetpotato production in South Africa. Agricultural Research Council-Roodeplaat, Pretoria, South Africa, 39–56 pp
Veasey EA, Borges A, Rosa M, Squeiroz-Silva JR, Bressan EDA, Peroni N (2008) Genetic diversity in Brazilian sweet potato [Ipomoea batatas (L.) Lam., Solanales, Convolvulaceae] landraces assessed with microsatellite markers. Genet Mol Biol 31:725–733
Whitaker D, Williams ER, John JA (2002) CycDesigN: a package for the computer generation of experimental designs. In: CSIRO forestry and forest products, CSIRO, Canberra
Zum Felde T, Burgos G, Espinoza J, Eyzaguirre R, Porras E, Gruneberg W (2010) Screening for β-carotene, iron, zinc, starch, individual sugars and protein in sweetpotato germplasm by near-infrared reflectance spectroscopy (NIRS). In: 15th Triennial symposium of the international society for tropical root crops, Lima, Peru. http://www.istrc.org/images/Documents/Symposiums/Fifthteenth/s8_zumfelde.pdf
Funding
Open access funding provided by Agricultural Research Council. This research was funded by the parliamentary grant allocated annually to the Agricultural Research Council by the South African government.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by SN, WM and SL. The first draft of the manuscript was written by SL and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Naidoo, S.I.M., Laurie, S.M., Booyse, M. et al. Early generation clonal selection of sweetpotato (Ipomoea batatas (L.) Lam) using an accelerated breeding scheme. Euphytica 220, 19 (2024). https://doi.org/10.1007/s10681-023-03276-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-023-03276-0