
Abstract
Cotton although is an autogamous species could be cross pollinated under favorable climate conditions and/or in the presence of pollinators. The coexistence of cotton with Malvaceae species raises questions on the possibility pollen to be exchanged among Malvaceae species and on the resulted consequences. The present work was undertaken to evaluate the in situ response of cotton flowers (G.hirsutum L, G. barbadense L.) and their F1 interspecific hybrids when are artificially pollinated with Malva sylvestris L., Hibiscus syriacus L. and Abelmoschus esculentus Moench. Furthermore, an in vitro protocol was attempted to support embryos’ growth in order to produce viable progenies originating from crosses between cotton and the aforementioned Malvaceae species. The obtained results gave evidence that pollen from the above Malvaceae species stimulated cotton ovaries without successful hybridization. The interaction between pollen and cotton’s stigmas was higher at early stages when M. sylvestris was used as pollinator but in crosses with H. syriacus and A. esculentus more ovules were activated as revealed by the percentage of carpodesis and life-time of bolls onto maternal plants. Only crosses between cotton with okra produced cotton seeds under in situ conditions. The in vitro embryo-ovule culture protocol, used, increased the number of regenerated cotton plants, especially in crosses among F1 interspecific cotton hybrids and A. esculentus. In this case, regenerated plants were recombinant aneuploids, combining traits from both cotton species. This novel cotton germplasm possessing unique chromosome rearrangements, at aneuploid level could be proved useful after cytogenetic, molecular or QTL genetic analysis referring to important agronomic traits.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Malvaceae is a plant family containing important genera like Gossypium, Hibiscus, Abelmoschus, Malva, Abutilon, Althaea, Lavatera and Malope. Several species from these genera are important agricultural crops (cotton, kenaf, orka) and others are ornamentals or weeds (Tate et al. 2005). The coexistence of Malvaceae species in an agricultural environment raises questions on whether it is possible pollen to be exchanged among Malvaceae species and what could be the consequences of such an exchange. Furthermore, the coexistence of conventional and GM cottons and probably other Malvaceae species as weeds in the same or neighboring fields could affect negatively the final product and have detrimental effect to the greater ecosystem.
From another point of view, intergeneric and interspecific hybridization have long been recognized from cotton breeders as an important tool for the introgression of alien germplasm and agronomic important traits (Mehetre 1993). In any case, wide hybridization could be used as an effective, bionomic and economical approach for transferring desirable genes from one species to another. It can be also applied for designing a strategy to avoid the dispersion of deleterious or undesirable gene flow especially in cases of coexistence of conventional cottons with GM ones at the same geographical areas. This problem is well illustrated by the work of a research group from S. Africa. This group investigated the possibility of outcross between Bt cotton and conventional cotton, and estimated the percentage of transgenes from Bt cotton to the conventional cotton (Bourgou et al. 2013a).
In nature, there are different internal and external barriers to prevent gene flow among related taxa (Hadley and Openshaw 1980). Hybrid formation is prevented mainly by prezygotic barriers at pollen–stigma or pistil interaction level and sometimes by postzygotic ones at embryo or endosperm formation level (Swanson et al. 2004). Previous studies related to crosses among cotton and Malvaceae species, indicated that although crossing is rare, some intergeneric hybridization has been successfully attempted. First of all, Brown (1947) reported the production of a hybrid plant between Gossypium hirsutum L. and Abelmoschus esculentum Moench. and she presumed that this plant (Z-118) should be the product of some form of chromosome rearrangement and reduplication in the ovule at early embryo stage. Currently, 70 years later one could theorize that fertilization and chromosome elimination did happen, although this was not feasible to be realized and explained at that time (Roupakias and Mavromatis 2010). Mehetre et al. (1980) reported the production of an intergeneric hybrid (2n = 38) between G. hirsutum L. and Hibiscus panduriformis Burm. with intermediate characteristics and meiotic irregularities. Zhou et al. (1991) however, reported parthenogenetic seed set at a rate of 1.3–2.4% when cotton flowers were pollinated with pollen from another Malvaceae species, and more precisely from kenaf (Hibiscus cannabinus L.). In such a case, the use and application of plant growth regulators or a supporting in vitro culture protocol of stimulated cotton ovules could be an alternative approach to generate hybrid or parthenogenetic cotton plants after pollination with kenaf (Mavromatis et al. 2005) or okra (Vlachostergios et al. 2007; Kantartzi and Roupakias 2008, 2010). Furthermore, a research group from S. Africa (Burkina Faso) investigated the possibility of outcross between Bt and conventional cotton, and estimated a percentage of outcrossing near to 4,63% from transgenes (Bt cotton) to the conventional ones (Bourgou et al. 2013b), giving useful information for outcrossing.
The present work was undertaken, (1) to study the in situ response of cotton flowers when pollinated with Malva sylvestris L., Hibiscus syriacus L. and Abelmoschus esculentus Moench. using phenotypical and cytogenetical approaches and (2) to develop an in vitro – ovule culture protocol for support embryos’ growth in order to produce viable progenies originated from crosses between cotton and the above mentioned Malvaceae species.
Materials and methods
Plant material
The experiments were established in the farm of the School of Agriculture of Aristotle University of Thessaloniki (40°32′N, 22°59′E, 4 m a.m.s.l.) and lasted for two years. Four G. hirsutum (i.e. Coker 310, Korina, Acala Sindos, 4S) and four Gossypium barbadense L. lines (i.e. Carnak, Menoufi, Pima S5, B 403), were grown in the first year of experimentation. All the aforementioned cultivars were cross pollinated with H. syriacus, A. esculentus (cv. Pilea) and M. sylvestris which were grown under the same field conditions (Fig. 1).

The Malvaceae species that were used as cotton pollinators. a Malva sylvestris, b Hibiscus syriacus, c Abelmoschus esculentus
Furthermore, interspecific hybridization between G. hirsutum x G. barbadense cultivars was attempted in all genotype combinations for the production of F1 cotton hybrids, and to observe the effect of cotton genotype as maternal plant or as a pollinator.
In the following growing season, the resulted five interspecific F1 hybrids (4S × Pima, Korina × Carnak, Carnak × 4S, B403 × Acala Sindos and Menoufi × Coker), were used as maternal plants to test the effect of genotype and influence of hybridization in the response of cotton ovules which produced after the stimulation of pollen from H. syriacus, A. esculentus and M. sylvestris (Table 1).
In situ hybridization and study of compatibility between cotton and Malvaceae species
The cotton flowers were emasculated by hand one day before anthesis and covered with paper bags to prevent cross pollination. The following day, most of the stigmas were pollinated with pollen from Malvaceae species whereas a number of the flowers remained five days after anthesis into bags without pollination (unfertilized flowers). Furthermore, self-fertilized flowers of the same age were used as controls to in situ and in vitro evaluation of all cotton genotypes and cross combinations.
A mean number of thirty cotton bolls of each category (self-fertilized, unfertilized and cross pollinated flowers) per year from all cross combinations, were left on maternal plants for further in situ development. This approach was applied to:
-
1.
Determine whether the percentage of carpodesis (%) (number of flowers developed to bolls five days after pollination/total number of pollinated flowers) and critical age for carpoptosis/lifetime (Days After Anthesis, (DAA)) of cotton bolls are depending on the genotype of cotton, the effect of pollinator and their interaction.
-
2.
Estimate the mean response of each cotton genotype, according to the influence of alien pollen derived from different Malvaceae species (H. syriacus, A. esculentus and M. sylvestris) through the estimation of viable ovules per boll under in situ development (Fig. 2).
Fig. 2 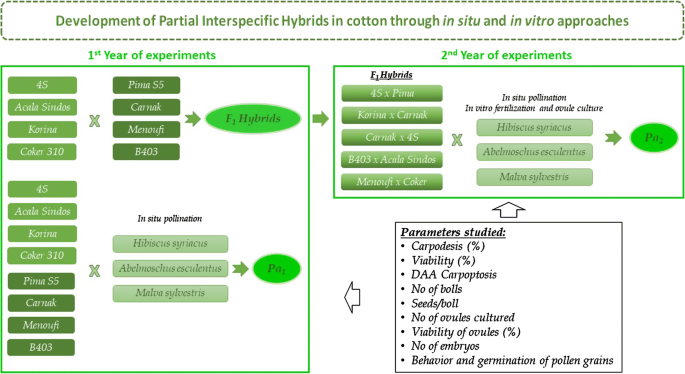
Outline for the development of partial interspecific cotton hybrids through in situ and in vitro approaches

In vitro ovule—embryo culture
The small percentage of bolls produced in situ was attributed to the assumption that a larger number of unfertilized ovules were stimulated by the alien pollen, but were probably unable to reach maturity. In such a case, in vitro culture of stimulated cotton ovules could be suggested as an alternative approach to support the development of young embryos in order to grow a higher number of cotton plants. For this reason, 15 young cotton bolls per cross combination and per year, were collected 5 days after pollination (5 DAP) (Mavromatis et al. 2012). The bolls were surface sterilized by soaking them in ethanol (70%), followed by dropping into CaHClO3 (3%) and finally by immersing them in a commercial chloride solution (Klinex) supplemented with two drops of Tween-20 for five minutes. The bolls were subsequently washed 2–3 times with sterilized distilled water and were dissected under sterile conditions in a laminar flow. The young ovules were ascetically removed and cultured in vitro on SH modified medium (Stewart and Hsu 1978) which had proved to be more suitable for cotton ovules (Mavromatis et al. 2004). Two treatments were applied according to the status of nutrient medium (solid and liquid form) (Fig. 3a, b) to improve the response of cotton ovules in comparison to previous experiments of our research (Vlachostergios et al. 2007; Kantartzi and Roupakias 2008). The treatment for liquid form consisted of 150 tubes with 1 ovule per tube for each genotype combination and the treatment for solid form consisted of 15 petri dishes (6 g/l agar) containing 10 ovules per each for all genotype combinations. In this case four replications were applied at different dates per season. All cultures were maintained under dark conditions for 48–52 days at 28 ± 2 o C. After 18–25 days of culture, most of the ovules were not developed or were depredated. However, in the living ovules when the development continued, young embryos were isolated from the ovules after 58–62 days in culture. These embryos were floated onto SH basal liquid medium. Four to six weeks later many embryos were not survived and only the germinated embryos were transferred to MS medium (Murashige and Skoog 1962) enriched with 1 mg/l Kin and 0.7 mg/l NAA for another two weeks in alternative 16 h photoperiod (4500 lx) at 28/24 o C day/night temperature regime. The survived plantlets were transferred into rooting medium (MS plus 0.5 mg/l NAA) and when the plants acquired 3–4 leaves, were transplanted into pots containing a soil mixture (sand 1: 1 peat moss) under a high relative moisture (80%) conditions in a growth chamber (Fig. 3c).

Main stages in the production of partial interspecific cotton plants. The young ovules cultured in liquid (a) and solid medium (b), the regenerated plantlets (c) and cotton plants survived under field conditions (d)
Cytological studies and ploidy level estimation of regenerated plants
The effect of alien pollen onto cotton’s stigmas, the percentages of germination of pollen grains from Malvaceae species and pollen tube growth through cotton pistils, were studied. For this, twenty cotton pistils were excised from pollinated flowers from all cross combinations at 24 to 72 h after pollination and fixed in FAA solution (1 formaline: 1 glacial acetic acid: 18 ethanol (80%)) v/v/v. After their storage at 4 °C, the pistils were rinsed with tap water, treated with NaOH (8 N) solution for 6 h and stained with 0.1% aniline blue in 0.1 M potassium phosphate solution (Mori et al. 2006). The colored pollen grains were afterwards mounted into a drop of glycerol on a slide, they were squashed after covering with a cover slip and observed by fluorescence microscopy.
Furthermore, the ploidy level of all survived plantlets and of their progenies was determined cytologically by counting the number of chromosomes and by using flow cytometric analysis. Chromosome number counts were carried out on root tips placed in saturated solution of bromonapthalene for 3.5 h and fixed in Carnoy’s solution for at least seven days. The root tips then, were stained in 1.5% acetic orchein for 2 days at 4 °C and the slides were prepared into a drop of 45% acetic acid (Lazaridou et al. 2017).
Flow cytometric analysis was performed with a PA-II flow cytometer (Partec, Germany). The flow cytometric assay involved the use of nuclei isolated from chicken red blood cells (CRBC) or another control (Pisum sativum), which served as an internal reference standard. The relative DNA content of individual plants was expressed as a ratio of DNA content of CRBC or Pea (P. sativum) and tetraploid cotton (DNA index).
In order to estimate the chromosome number using flow cytometry, the relative DNA content of plants with unknown ploidy level was expressed as a percentage of the DNA content of tetraploid cotton plants. In all measurements, the DNA extract sample from pea (P. sativum) (1C: 4.43 pg) was used as a reference standard. The gain of the instrument was adjusted in such a manner that the G1 peak of the pea’s nuclei positioned approximately at channel 150 and the G1 phase of tetraploid cotton’s nuclei at 100 channel so any abbreviation could be due to aneuploid phenomena.
The samples were prepared according to Galbraith et al. (1983) and Dolezel et al. (1998). The young cotton leaves were chopped with a razor blade in a petri dish containing 1 ml of isolation buffer Partec (I) (Mavromatis et al. 2005). Then, the nuclei suspension was filtrated through a nylon disposable filter of 50 mesh (CellTrics) directly into a sample tube. After three minutes of incubation into ice, 3 ml of staining solution DAPI (PARTEC II) was added inside the tube and the analysis was taken place with the UV excitation in Partec CA II—ploidy analyzer.
Results & discussion
A significant criterion for the influence of alien pollen on cotton’s stigma was the percentage of carpodesis. The mean percentage for all cotton genotypes used, was ranged from 5.2% for unfertilized ovules to 82.8% for the self-fertilized ones (3–5 DAP) (Table 2). At the same period the percentages (%) of carpodesis when H. syriacus and A. esculentus were used as pollinators ranged from 41.6 to 39.7%, respectively. These percentages differentiated statistically from self-fertilized and unfertilized ones and also from the percentage recorded when M. sylvestris was used as pollinator (15.8%). Furthermore, the viability of ovules (5 DAP) stimulated by H. syriacus and A. esculentus, was intermediate between the corresponding values for self-fertilized cotton flowers and those pollinated with M. sylvestris. (Table 2).
According to the results of the present study no differences were recorded to the percentage of carpodesis when the cotton plants were fertilized by pollen of M. sylvestris. The influence of pollen from M. sylvestris did not differ statistically from the corresponding values of unfertilized ovules (Table 2). In this case, all young bolls dropped down at 6 to 7 DAP, a period no longer than two or three days with respect to the unfertilized ones (3–5 DAP). Reversely, the percentage (%) of carpodesis was influenced by the genotype of maternal cotton plant and was ranged from 32.2% (Korina × H. syriacus) to 52.6% (Carnak × H. syriacus) when flowers from different cotton genotypes were pollinated with pollen originated from H. syriacus (Table 3). Similar percentages were observed when Korina and Carnak were crossed with A. esculentus (52.3 and 54.2 respectively, Table 3). Analogous high degree of compatibility was also recorded for most of the cotton genotypes, tested.
The percentages of carpodesis recorded for cross combinations, were significantly higher than those observed in unfertilized cotton flowers and significantly lower than self-fertilized ones. It is notable that into crosses with A. esculentus, young cotton bolls remained on the maternal cotton plants for a longer period (12–21 DAP) than the other combinations (7–12 DAP for H. syriacus) and (6–8 DAP for M. sylvestris) (Table 3). This observation clearly indicates that pollen from A. esculentus effectively stimulated cotton ovules. This stimulation could be attributed either to fertilization or only via pollen germination onto cotton’s stigmas. In addition, some of the young bolls developed after pollination with alien pollen, were maintained on the maternal plant for more than 20 days as compared with the unfertilized flowers that were shed at 3 to 5 DAA. The same was recorded and when the cotton plants were crossed with H. syriacus (8–12 DAP) or M. sylvestris (6–7 DAP) (Table 3).
Similar observations were noticed at the 2nd growing season when Malvaceae species were used as pollinators in crosses with F1 interspecific cotton hybrids. In a comparison among different pollinators and self-pollination, a clear influence of the alien pollen effect was observed. Furthermore, the period that young cotton bolls remained onto maternal F1 interspecific plants was influenced by the pollinator and the genotype of the maternal cotton plant. The F1 interspecific hybrids (4S × PIMA), (Carnak × 4S) produced the higher percentage of carpodesis with all pollinators, used (Table 4). Especially, in combinations of the above-mentioned genotypes with A. esculentus, the young cotton bolls remained for a longer period onto the maternal plants and some well-developed seeds reached the maturity stage. The highest percentage of carpodesis (%), the extensive period until carpoptosis (DAA) and the best seed set were observed only in combinations between three of the interspecific hybrids (4S × PIMA, Carnak × 4S and B403 × Acala S.) with A. esculentus (Table 4).
In ovule–embryo culture
In order to increase the number of embryos that could reach maturity an in vitro culture protocol was applied for rescuing young embryos originating from hybridization or alien pollen (A. esculentus, H. syriacus, M. sylvestris) stimulation. In all cross combinations, the number of well developing ovules per boll was ranged from 8 to 12. This number is below the mean number usually observed in self-fertilized bolls which ranges from 20 (G. barbadense) to 28 (G. hirsutum). Furthermore, the mean values of ovule’s viability after 20 days in culture, ranged from 6.4 to 20.8% in combinations with M. sylvestris, 12.8–32% in crosses with H. syriacus and 30.2–79.2% in combinations with A. esculentus (Table 5).
However, it is obvious that the genotype of the female parent had an effect on the ovule’s response. The mean percentage of ovule’s viability when the plants were crossed to A. esculentus ranged from 60.42 to 71.92% when G. barbadense and G. hirsutum were used as maternal plants respectively (Table 5). In the case of F1 interspecific cotton hybrids this percentage ranged from 32.5 to 40.8% and was lower than the respective in cotton genotypes (Table 5). The overall response of all genotypes used, according to percentage of carpodesis, ovule’s viability at 20 days in culture and number of embryos retained within the ovules are given in Table 5. The viability of ovules was lower in crosses with H. syriacus and this was clearly reduced in crosses with M. sylvestris. Also in these cases, G. hirsutum genotypes responded better but no viable embryos were found after 30 days under in vitro culture conditions. On the contrary, in 5.5% of cultured ovules obtained from crosses between F1 interspecific cotton hybrids and A. esculentus, immature embryos were found and they continued to grow until 52 DAP. From all young embryos produced, seven plants were recovered because most of them did not survive after transplantation. This happened because most of the plants were weak, with small chlorotic leaves and were not capable to survive when they transferred from in vitro to field conditions (Fig. 3d).
Cytological studies, fluorescence microscopy and ploidy level estimation of regenerated plants
Despite the production of a limited number of plantlets, hybridization between cotton and okra (A. esculentus) must be theorized impossible. This conclusion was driven from the fluorescence microscopy on pollen grains and after studying the interaction between alien pollen with cotton’s stigmas (Table 6). The pollen grains of okra germinated on cotton’s stigma abnormally, their growth was very slow and nevertheless entered the micropyle (Fig. 4). The majority of cross combinations exhibited an abnormal pollen germination, which means that branching presence of two or more tubes per grain and the appearance of a strong callose layer, were frequent phenomena. The pollen from okra had already germinated 1 DAP but most of the pollen grains remained out of the style. Some pollen grains entered the stigmas (2 DAP) and covered the 1/3 of cotton’s style but the majority of pollen tubes did not grow beyond this area. In case of interspecific cotton hybrids when they were crossed to A. esculentus, it was observed that some pollen tubes from okra continued to grow 72 h after pollination and they approach the area but nevertheless entered inside the micropyle.

The abnormal germination of pollen grains from okra
The interaction of pollen grains from M. sylvestris with cotton stigmas is particularly impressive at the first 24 h after pollination. A large number of pollen grains were germinated (20.8%) and pollen tubes started to grow fast into cotton stigmas. Afterwards a very abrupt stop was observed 48–52 h later, because of the appearance of a strong callose layer. An intermediate behavior (12.4%) was observed in the case of crosses between cotton and H. syriacus. Only a small number of pollen grains germinated 24 h after pollination but a fast growth was observed for all tubes that penetrated the stigma 20 h later. However, 48 h after pollination most of pollen tubes appeared abnormal and stopped growing due to the development of a callose layer.
The mean response for all pollinators used, combined with genotypes from two cotton species (G. hirsutum, G. barbadense) and their F1 interspecific hybrids according to the number of alien pollen grains adhered and penetrated their stigmas as related to the time after pollination (24–72 h), are given in Table 6. Moreover, for a better understanding of the pollen’s effect according to its origin, a comparison among the mean response of all cross combinations (Table 6) and self-fertilization (Table 7) is presented.
Cytological studies applied to in situ produced cotton seeds and also to regenerated cotton plants derived from in vitro culture, indicated that the chromosome number observed in most of the plantlets and seedlings, ranged from 28 to 52 (Fig. 5a–c). The differences between seedlings derived from in situ hybridization (from all cross combinations only seven cotton plants were survived until maturity) and plantlets originated from in vitro culture (four cotton plants regenerated only through crosses with A. esculentus), were focused on the stability in chromosome number. In the first case, most of seedlings (5/7) had quite the same number of chromosomes (32–34) but the young regenerated cotton plantlets were differentiated having from 28 to 38 chromosomes. Also, some plantlets were mixoploid, having simultaneously different number of chromosomes in their roots.

Mitotic metaphase of root meristematic cells of cotton plants showing a range from 46 to 52 chromosomes (a aneuploid, 46 chromosomes, b aneuploid, 48 chromosomes, c normal, 52 chromosomes)
These results were confirmed by flow cytometric analysis (Fig. 6). It is clear that tetraploid cotton used as a control, gave a peak in the climax for G2 phase, very close to 350 nm. In the case of A. esculentus, most of the cells were arrested very close to 50 nm for G1 phase and to 100 nm for G2 phase, respectively. It could be stated that the most of Pa0 cotton plants exhibited their peaks for G2 phase, from 230 to 250 nm which characterized by reduced values for DNA content. These plants could be considered as aneuploids confirming previous analogous reports (Vlachostergios et al. 2007).

Flow cytometric analysis
With the results of the present work, a long-term effort from laboratory of plant genetics and breeding of aristotle university of Thessaloniki (AUTH) was completed. Aim of this effort was to create partial interspecific (PI) hybrids in cotton, incorporating the desired traits from G. hirsutum and G. barbadense into the descendant progenies. Although the first target was the production of PI lines which inherited the desirable cotton characteristics from generation to generation (Roupakias and Mavromatis 2010), a number of aneuploid lines with different number and type of chromosome combinations, was produced.
In our previous research in situ cross experiments and in vitro culture approaches were applied. The unfertilized cotton ovules originating from cultivars and interspecific hybrids from in situ hybridization between F1 interspecific cotton hybrids and H. cannabinus (Mavromatis et al. 2005) or A. esculentus (Kantartzi and Roupakias 2008) and sometimes supported by in vitro ovule or embryo culture protocol (Vlachostergios et al. 2007; Kantartzi and Roupakias 2009), were tested.
In the present research, the experiments were repeated with the same or other genetic material using modified solid and liquid nutrient media and other Malvaceae species (M. sylvestris, H. syriacus) as inducing pollinators. These approaches confirmed our initial hypothesis that hybridization among Malvaceae species could increase the rate and number of regenerated partial interspecific plants and determine the nature of aneuploid PI cotton plants. Most of these plants originated through in situ or in vitro development were viable and capable to participate in fertilization, confirming the assumptions of Myles and Endrizzi (1989) and expectations of many cotton breeders (Tyagi et al. 2014).
Conclusions
The results of this work gave evidence that pollen from Malvaceae species (M. syslvestris, H. syriacus, A. esculentum) stimulated cotton ovaries without successful hybridization. The interaction between pollen and cotton’s stigmas was higher at early stages when M. sylvestris was used as pollinator but in crosses with H. syriacus and A. esculentus more ovules were activated as it was estimated by the percentage of carpodesis and life-time of bolls onto maternal plants. Only crosses of cotton with okra were capable to produce cotton seeds under in situ conditions. Furthermore, by the support of an improved in vitro embryo-ovule culture protocol, a high percentage of regenerated cotton plants could be succeeded. Especially in crosses of F1 interspecific cotton hybrids with A. esculentus the regenerated plants were recombinant aneuploids, combining simultaneously traits from the two cotton species (G. hirsutum and G. barbadense), which were viable and capable to be fertilized. The practical result from this research was the production of novel cotton germplasm possessing unique chromosome rearrangements, at aneuploid level. This germplasm could be proved very useful to cytogenetic and molecular studies or QTL genetic analysis for important traits in cotton.
References
Bourgou L, Sanfo D, Tiemtore CB, Traore O, Sanou J, Traore K (2013a) Assessment of possible hybridization between Bt cotton (Gossypium hirsutum L.) and other Malvaceae species cultivated in Burkina Faso. Afr J Biotechnol 12(14):1609–1616. https://doi.org/10.5897/AJB12.2012
Bourgou L, Sanfo D, Tiemtore CB, Traore O, Sanou J, Traore K (2013b) Assessment of Bollgard II cotton pollen mediated transgenes flow to conventional cotton in the farming conditions of Burkina Faso. Afr J Biotechnol 12(33):5192–5199. https://doi.org/10.5897/AJB12.1931
Brown MS (1947) A case of spontaneous reduction of chromosome number in somatic tissue of cotton. Am J Bot 34:384–388. https://doi.org/10.1002/j.1537-2197.1947.tb13005.x
Dolezel J, Greilhuber J, Lucretti S, Meister A, Lysak MA, Nardi L, Obermayer R (1998) Plant genome size estimation by flow cytometry: inter-laboratory comparison. Ann Bot 82(A):17–26. https://doi.org/10.1093/oxfordjournals.aob.a010312
Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DB, Firoozabady E (1983) Rapid flow cytometric analysis of the cell cycle in the intact plant tissues. Science 220(4601):1049–1051. https://doi.org/10.1126/science.220.4601.1049
Hadley HH, Openshaw SJ (1980) Interspecific and intergeneric hybridization. In: Fehr WR, Hadley HH (eds) Hybridization of crop plants. American Society of Agronomy, Madison
Kantartzi S, Roupakias DG (2008) Production of aneuploids of the cotton hybrid G. barbadense × G. hirsutum L. via intergeneric pollination with Abelmoschus esculentus. Euphytica 161:319–327. https://doi.org/10.1007/s10681-007-9550-z
Kantartzi S, Roupakias DG (2009) In vitro gynogenesis in cotton (Gossypium sp.). Plant Cell Tissue Organ Cult 96:53–57. https://doi.org/10.1007/s11240-008-9459-9
Kantartzi S, Roupakias DG (2010) Study of apomictic seed formation in interspecific, Gossypium barbadense × G. hirsutum, cotton hybrids. Int J Bot 6(2):164–169
Lazaridou TB, Pankou CI, Xynias IN, Roupakias DG (2017) Effect of the 1BL.1RS wheat-rye translocation on the androgenic response in spring bread wheat. Cytol Genet 51(6):485–490. https://doi.org/10.3103/S009545271706007X
Mavromatis AG, Kantartzi SK, Vlachostergios DN, Xynias IN, Skaracis GN, Roupakias DG (2005) Induction of embryo development and fixation of partial interspecific lines after pollination of F 1 cotton interspecific hybrids (Gossypium barbadense × Gossypium hirsutum) with pollen from Hibiscus cannabinus. Aust J Agric Res 56:1101–1109. https://doi.org/10.1071/AR04241
Mavromatis AG, Kantarzi SK, Vlahostergios D, Skaracis G, Roupakias D (2004) Doupled haploids from cotton interspecific hybrids (G. hirsutum × G. barbadense): a new source of genetic variability for cotton breeding. In: COST action meeting of WG 851./WG3, Tulln, Austria Sept. 2004.
Mavromatis AG, Khah EM, Vlahostergios D, Roupakias DG (2012) Wide Hybridization as a scientific tool to understand the genetic background of cotton. In: 1st international conference on science industry and trade of cotton—Oct 2–4, 2012, Gorgan, IRAN.
Mehetre SS (1993) Distant hybridization in cotton breeding-intergeneric hybridization (an overview). J Cotton Res Dev 7(2):179–192
Mehetre SS, Thombre MV, Tyyab MA (1980) Cytomorphological studies in an intergeneric hybrid between Gossypium hirsutum L. (2n = 52) and Hibiscus panduraeformis Burm. Euphytica 29:323–330. https://doi.org/10.1007/BF00025130
Mori T, Kuroiwa H, Higashiyama T, Kuroiwa T (2006) Generative cell specific is essential for angiosperm fertilization. Nat Cell Biol 8:64–71. https://doi.org/10.1038/ncb1345
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497. https://doi.org/10.1111/j.1399-3054.1962.tb08052.x
Myles EL, Endrizzi JE (1989) Aneuploids induced by deficiencies of chromosome 9 and analysis of the time of nondisjunction in cotton. Genome 32(1):12–18. https://doi.org/10.1139/g89-403
Roupakias DG, Mavromatis AG (2010) Cotton Breeding in the 21st Century. In: Wakelyn PJ, Chaudhry MR (eds) Cotton: technology for the 21st Century. International Cotton Advisory Committee, Washington, DC
Stewart JM, Hsu CL (1978) Hybridization of diploid and tetraploid cottons through in-ovule embryo culture. J Hered 69:404–408. https://doi.org/10.1093/oxfordjournals.jhered.a108982
Swanson R, Edlund AF, Preuss D (2004) Species specificity in pollen-pistil interactions. Ann Rev Genet 38:793–818. https://doi.org/10.1146/annurev.genet.38.072902.092356
Tate JA, Aguilar JF, Wagstaff SJ, La Duke J, Bodo TA, Slotta J, Simpson BB (2005) Phylogenetic relationships within the Trible Malveae (Malvaceae) as inferred from ITS sequence data. Am J Bot 92(4):584–602. https://doi.org/10.3732/ajb.92.4.584
Tyagi P, Bowman DT, Bourland FM, Edmisten K, Campbell BT, Fraser DE, Wallace T, Kuraparthy V (2014) Components of hybrid vigor in upland cotton (Gossypium hirsutum L.) and their relationship with environment. Euphytica 195(1):117–127. https://doi.org/10.1007/s10681-013-0987-y
Vlachostergios DN, Mavromatis AG, Kantartzi SK, Roupakias DG (2007) In-vitro development of ovules obtained after pollination of cotton (Gossypium spp) flowers with pollen from okra (Abelmoschus esculentus L. Moench). Plant Cell Tissue Organ Cult 88:109–115. https://doi.org/10.1007/s11240-006-9178-z
Zhou SQ, Qian DQ, Cao XY (1991) Induction of parthenogenesis and chromosome behavior in plants of parthenogenetic origin in cotton (Gossypium hirsutum). Genome 34:255–260. https://doi.org/10.1139/g91-040
Acknowledgements
The authors kindly acknowledge Dr. Anastasios Lithourgidis, Director of the Farm of the Aristotle University of Thessaloniki, for his assistance in field experiments.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
AM and DR conceived and designed experiments. CP and DV conducted experiments and analyzed data. IX and AM conducted the cytogenetic analysis. AM, CP, DV, IX and DR wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Mavromatis, A.G., Pankou, C.I., Vlachostergios, D.N. et al. Hybridization between cotton and Malvaceae species as a tool for production of partial interspecific aneuploid cotton plants. Euphytica 214, 179 (2018). https://doi.org/10.1007/s10681-018-2257-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-018-2257-5