
Abstract
Forests, a valuable source provided by nature to living beings, are indispensable for many living organisms; hence, it is important to ensure the sustainability of forests. Determining the factors that exposure threats to the forests, executing protective methods against them and putting these methods into practise are important for the ecological cycle. Bark beetles, which have destructive effects on the ecosystem, are one of the factors that expose a threat to forests. Therefore, monitoring of these species and determination of effective control strategies are increasingly gaining importance in forestry. Conventional pheromone traps, which are being currently used, provide limited information on flight times of target species. Therefore, the technological development of the capture systems of these traps will determine future control trends. Hence, pheromone traps with electronic control unit were prepared in earlier (ѵ1) and new designed (ѵ2) versions. In ѵ2, 97.5% of target species were counted, and instant temperature, humidity and time parameters at the time of capture were recorded at a practiced field work for the system. In addition to the instant parameters recorded in ѵ2, an anemometer used for measuring wind speed, which is considered to have influence on the behaviour of target species, was incorporated into the system. In the trials, the counting success rates under daylight and darkness conditions for Ips sexdentatus adults were 98.1 and 97%, whereas the counting success rates for Pityocteines curvidens adults, which are smaller in size, were 96 and 99%, respectively. In conclusion, data obtained by recording the amount of target species along with the capture moment and parameters related to this will be very useful and provide determinative in the management of target species.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Bark beetles (Coleoptera: Curculionidae: Scolytinae) are one of the most destructive biotic factors that affect the forests (Marini et al. 2017). Although many species of bark beetles prefer dead or dying trees, a few species may kill healthy trees (Lindgren and Raffa 2013). Remarkable changes occur in the forests due to the outbreak of these species (Macek et al. 2017), which are primarily and secondarily harmful (Wood and Bright 1992), resulting in adverse effects, particularly on the ecosystems (Gitau et al. 2013; Hansen 2014). These species, which are considered to have an increasing importance as beetles of conifer forests (Gitau et al. 2013; Lindgren and Raffa 2013), cause considerable economic losses in the forests (Lindgren and Raffa 2013; Sánchez-Martínez and Wagner 2002). Although they cause death of trees in the forests where host species are present (Meddens et al. 2012; Negrón and Popp 2017), the outbreak of these species exposes a major threat to the productivity of forests (Zang et al. 2015). Besides, damages on trees have a significant effect on timber production (Schwab et al. 2009). In order to ensure the sustainability of forest existence, it is indispensable to carry out a control programme of active bark beetles in the forests (Özcan et al. 2016).
The presence of host trees controls the population of bark beetles and limits the growth of their populations (Gómez et al. 2017; Rudinsky 1962). Undoubtedly, the riskiest period for these species is the time of finding a host (Byers 1996). The species have a complex strategy to find their hosts and location in forests (Raffa et al. 1993). Many target species that are monitored using pheromone traps can fly very long distances (Nansen 2004). Long-distance distribution of bark beetles, which causes the finding and placing to host plant and so increase their populations, is a significant feature that influences their population dynamics and distribution (Meurisse and Pawson 2017). Distribution is an important process in the response of beetle’s populations to rapidly changing environmental conditions (Kautz et al. 2016).
The objective in the control of bark beetles must be to minimise and/or reduce the beetle population (Gómez et al. 2017). Silviculture applications may be effective in reducing the population of bark beetles (Gómez et al. 2017). Therefore, sanitation and mass trapping are the principal control methods to determine the populations of these species (Yonker 1990). In addition to being used to study the biology and behaviour of many bark beetle species (Lindgren and Borden 1983), pheromone traps are important in the management of forest beetles (Knodel et al. 1995; Galko et al. 2016). Because pheromones specific to these species are used in these traps, more exact data are obtained (Wainhouse 2005). The use of pheromone traps (Sagitov et al. 2016), one of the basic factor of the Integrated Pest Management, remains important in terms of control strategies (Galko et al. 2016).
Pheromone traps are the most common strategy used to monitor the density and distribution of bark beetle populations (Galko et al. 2013). Pheromone trapping applications are utilised to reduce the risk of outbreak of active beetles and increase the efficiency of control (Zúbrik et al. 2008). Particularly, mass trapping applications are effective in reducing the density of target species population (Özcan et al. 2011). Pheromone traps, which are preferred for monitoring the flight times of target species (Holuša et al. 2012), are important for the early detection of outbreaks, determination of outbreak areas, monitoring of beetle population and selection of a beetle control method (Knodel et al. 1995; Byers 2006; Hayes et al. 2008). One of the most significant advantages of pheromone traps is that they minimise the risk of the host trees located near them being attacked (Lindgren and Borden 1983), which reduces the damage by bark beetles in the rest of the forest (Krieger 1998). In addition, in the study conducted by Sánchez-Martínez and Wagner (2002) in various species, it was stated that the bark beetles can be captured by the traps in a large radius. This is evidence showing that the activity area of traps is considerably large. The traps are biotechnical elements used for both control (Raty et al. 1995; Gillette and Munson 2007; Özcan et al. 2016) and monitoring of target species population (Baker 2008; Bakke 1991; Baker 2008; Faccoli and Stergulc 2006; Lindelow and Schroeder 2001; Meurisse et al. 2008; Özcan et al. 2014; Turchin and Odendall 1996).
Control strategies to be planned against target species, with which the increased population may expose a threat in the forests they are active, always remain in the forefront. Therefore, it is necessary to make additional contributions to the already-existing conventional control strategies in the presence of species. Development of the intended use of pheromone traps, which have been used for target species for a long time, along with the support of various elements and the providing of detailed data will remarkably help in the determination of the future risks. The results from this study and then the successive data to be obtained through common applications will facilitate models to be created by considering parameters including target species population, temperature, humidity, time and wind speed. These results will help in planning control strategies that will enable analytical estimations from monitoring areas to larger forest areas, assist in taking early controls against beetles and reduce damage levels to below the expected. In light of this information, pheromone-baited traps with the integrated electronic control unit as a new methodology was designed and improved ѵ1 which involved laser scanning system and ѵ2 which used a motion sensor to detect large-scale target species, respectively. Also, the improved ѵ2 were made to overcome the explained possible handicaps, although ѵ1 and ѵ2 are successful in determining the target species.
Experimental procedure
New design
The pheromone trap with ECU (version ѵ1) designed by Özcan et al. (2014) aimed to determine various parameters at the time of capture of target species. Hence, temperature and humidity variables at the time of capture of the beetles detected by the laser scanning system were recorded in microSD card with the help of a card ECU. The recorded parameters are stored in the ‘comma-separated CSV’ file within the microSD card by the microcontroller. The designed unit could be applied to all types of traps because it is placed at the entrance of the trap chamber. However, in the fieldworks carried out with ѵ1 system, it was observed that the failure to count beetles due to the location of target species at the time of falling into the chamber through laser scanning system may result in handicaps, i.e. inability to determine small-sized target species. In the first version and in this study, Scandinavian-type® three-funnel pheromone traps that have been commonly used in Turkey were preferred to monitor bark beetles. ECU was created using a PV system equipment (PV module, battery, charge controller, fuse and cabling) and an electronic card equipment (microcontrollers, sensors, microSD card and electronic units) (version ѵ2).
The pheromone trap with ECU (ѵ2) has been developed based generally on the previous design. Some innovations and improvements were made in line with the possible handicaps and their solution proposals. The design of previously used system had been made based on original pheromone traps. However, in light of the possible handicaps during the counting process and the statistical data obtained, it was decided that the traps need to be improved and a new design was developed for the entrance of the reservoir. In this new design, instead of the laser scanning system used in the previous work, a different type of sensor, the motion sensor, was adapted to the system (Fig. 1).

Design view of the system developed using AutoCAD 2016®
Nonetheless, off-grid solar energy systems are still utilised to enable them to operate independently from energy in the forest for long periods of time. This system includes solar panel, charge controller, battery, fuse and cabling. A microcontroller unit is used to control the system. The details of the system have been submitted in previous studies, and the system continues to be used in a similar way.
The parametric data obtained in the first applied system (ѵ1) were expressed as temperature, humidity, date and time. An anemometer was incorporated into the developed system (ѵ2) to determine the wind-speed parameter, in addition to the abovementioned parameters, that is considered to have an effect on the behaviour of beetles. The programme of ѵ2 system was re-updated based on new studies, and the flow diagram of newly designed programme is given below (Fig. 2).

Flow diagram of the newly designed programme
The fieldwork of the ѵ2
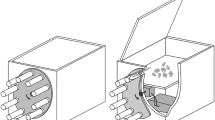
In the scope of the study, a pheromone trap was designed (ѵ1) in order to represent the biology and behaviour of target species along with specific parameters, and the prototype works and system success results were submitted in detail by Özcan et al. (2014, 2016). However, ѵ2 was designed to eliminate certain handicaps observed in ѵ1 in the fieldwork and to successfully record smaller-sized target species (Fig. 3). Besides, every counting group was studied under daylight and darkness conditions in order to evaluate the counting success rate of ѵ2 and determine whether the energy requirement is sufficient, particularly in darkness conditions.

The experimental illustration of the ѵ2 involving microcontroller to determine target species for monitoring
Overall, 649 Ips sexdentatus and 722 Pityokteines curvidens (Germ.) (Coleoptera: Curculionidae: Scolytinae) live adults were used in this study. The sizes of 50 adults for each species were precisely measured. The average size of I. sexdentatus adults that differed in size was found to be between 5.4 and 8.7 mm, whereas that of P. curvidens adults was between 2.1 and 3.1 mm (Fig. 4d, e). The target species were collected from the pheromone traps in the forest and under the bark of the invaded trees in the first days of August in order to evaluate the counting success and parameters recorded during the passage of beetles of different sizes through the trap chamber (Fig. 4a,b,c). Live target species obtained were counted at different hours of the day between 5th and 7th August 2017.

Collecting samples used in the designed system from forest: aDamage by Ips sexdentatus in Anatolian Black Pine Forest (Kastamonu/Turkey, 2017), bIps sexdentatus gallery, c bark beetles collected in pheromone trap, d Ips sexdentatus used in the trials and e Pityokteines curvidens adults
The counting results were obtained by throwing two different target species grouped as mono, double and triple into traps under two different light conditions (day and night). The trials were performed with a total of 1371 adults (649 I. sexdentatus and 722 P. curvidens adults) in a total of 687 throws at random intervals regardless of time frequency (Table 1).
All statistical analyses were performed using IBM SPSS 20.0 for Windows® software. The differences between the counting success rates under daylight and darkness conditions in groups for target species of different sizes were measured using chi-square test.
Results
When I. sexdentatus adults were evaluated regardless of the differences in light conditions in mono, double and triple groups, it was observed in all trials that 97.5% (317) of 325 throws were successfully recorded, whereas 2.5% (8) were not recorded. Similarly, in the evaluations in the same category with P. curvidens adults, it was observed that 97.5% (353) throws were successfully recorded, whereas 2.5% (9) were not recorded. For I. sexdentatus adults, 98.1% of 161 throws in daylight and 97% in darkness were successfully counted and recorded. For P. curvidens adults, the success rate under daylight and darkness conditions were 96 and 98.9%, respectively.
When adults of both target species were separately evaluated, no statistically significant difference was observed between the counting success rates under daylight and darkness conditions (p > 0.05) (Table 2). The absence or presence of light does not affect the counting success for both target species and does not alter the counting success of traps. Moreover, the counting success rate was found to be successful under different light conditions for ѵ2.
In addition, it was observed that the energy required for the operation of ѵ2 can be fully supplied under conditions without daylight. Adults were tested in mono, double and triple groups for the evaluation of two different target species. The counting success rates of each groups in daylight for I. sexdentatus were found to be 96.1, 100 and 98.3%, whereas in darkness, these were 93.5, 98 and 98.1%, respectively (Table 3).
The counting success rates for P. curvidens adults in the same group were 96.3, 94.7 and 98.4% in daylight, and whereas 98.5, 100 and 98.3% in darkness conditions, respectively (Table 4). Rates for the failure to count or the detection of double and triple groups as a single group were ≤ 6.5% for I. sexdentatus adults and ≤ 5.3% for P. curvidens adults. It is highly likely that multiple adults are captured by the pheromone traps based on the natural environment conditions, and it was observed that the margin of error in this regard was considerably reduced with the new sensor design of ѵ2. For the target species, no statistically significant difference in terms of counting success rate was observed among the mono, double and triple groups under both daylight and darkness conditions (p > 0.05) (Tables 3 and 4). Based on this evaluation, the fact that both species are captured by the traps in mono or multi groups does not affect the success rate, and they can be counted and recorded with a high success rate.
In the evaluation of counting success for target species with different sizes, no statistically significant difference was observed among the groups under both daylight and darkness conditions (p > 0.05). The counting success rate for large- and small-sized target species in daylight conditions was between 96.1 and 100%, whereas it was 93.5 and 100% in darkness conditions (Fig. 5). In conclusion, the counting success for species with different sizes is considerably high.

Counting success rates for different target species under both daylight and darkness conditions
Discussion
Galko et al. (2016) emphasise that the development of the capturing systems of traps will be a future trend. In conventional pheromone traps, which are currently used, provide limited information on the flight time and duration of target species (Özcan et al. 2011; Özcan 2017). ECU has been incorporated into these conventional traps to record the capture moment and the temperature and humidity values at that moment (ѵ1) (Özcan et al. 2014). Based on the results of the trials, the designed trap was considered to be successful (Özcan et al. 2016). Effective use of data obtained by these traps will reduce damages and subsequent economic losses caused by the species (Özcan et al. 2016). Although instant temperature, humidity and time parameters were recorded with a high capture success rate of 87.1% in ѵ1 (Özcan et al. 2016), this rate increased to 97.5% in ѵ2. In addition to instant parameters recorded in ѵ1, wind speed was incorporated into the trap. In the trials conducted with I. sexdentatus adults, the counting success rates in ѵ1 under daylight and darkness conditions were 83.7 and 90.2% (Özcan et al. 2016) and the counting success rates in the trials with the same target species were 98.1 and 97%, respectively. The success rate in ѵ2 was increased compared with that in ѵ1 with I. sexdentatus adults (Özcan et al. 2016), whereas the success rates for the same species based on different light conditions were increased by 14.4 and 6.8%, respectively.
In addition, the success rates for P. curvidens adults with smaller sizes were as high as 96 and 99%. It was found that ѵ2 had a higher success rate for the adults of the same species compared with ѵ1, and it was also determined to be considerably successful for small-sized species. ѵ2 was designed so that environmental conditions do not have any effects on the counting success for both target species. The trials with ѵ1 under darkness conditions were more successful than those performed under daylight conditions (Özcan et al. 2016); however, the problems caused by light were resolved and the success rate increased in ѵ2. Additionally, the new design has proved to have a successful counting rate of approximately 100% under different light conditions. In addition, it was observed that the energy required for the operation of ѵ2 can be fully supplied under conditions without daylight. ѵ1 can determine the differences (millisecond) in refraction of light with an average error margin of 15% (Özcan et al. 2016); this rate was decreased to ≤ 6.5% by the use of motion sensor instead of a laser scanning system. Angle of fall for the beetles in the designed ѵ2 cannot be controlled for each throw under different light conditions. Detection of beetles is possible by passing in front of the motion sensor. In darkness and daylight conditions, differences of the success rates, which is negligible, are related to the position of the beetle when it is caught. This difference can be overcome with improvements on the design.
Although mass trapping is not always a precise control method for target species (Panzavolta et al. 2014), it is stated that these applications would be useful for the protection of forests against I. typographus (Faccoli and Stergulc 2008). Using ѵ2 will contribute to the determination of behaviour of target species such as I. typographus. Weslien et al. (1989) suggested that the numbers of I. typographus adults captured by the pheromone traps reflect the damage caused by the bark beetles in the region within the year. However, the data obtained are limited and are expressed by prediction. On the other hand, data obtained from mass traps are important because they can be used to plan the control methods (Yonker 1990). Besides, the flight times of target species adults may vary every year; hence, the population dynamics of target species need to be monitored (Panzavolta et al. 2014). Data obtained by this design, particularly ѵ2, will help to make accurate and reproducible predictions on tree mortality rates in outbreak areas.
It is known that there is a high correlation between the number of beetles captured by the traps and the volume of wood invaded or the number of dead trees (Faccoli and Stergulc 2004). Therefore, the number of beetles captured by the traps can provide information on the population of species and give insight into the possible tree mortality rates. Parameters obtained from the conventional traps during the monitoring and control processes provide detailed information on the behaviour of target species population. The moment and number of species captured are important for making predictions. In addition, data obtained by recording the numbers of target species along with parameters that contribute to the clarification of biology and behaviour of target species will be very useful and provide guidance in the management of target species. On the other hand, elimination of outbreaks of these species is difficult and expensive. Precautions taken to prevent these outbreaks are effective in reducing the losses caused by these beetles (Schowalter 2012). It is indispensable to control programs to reduce the potential future outbreaks of bark beetles or the intensity of outbreaks and to use new methods in this regard in today’s technology.
So far, researchers have tried to determine sensitive areas by taking into account forest conditions and focussing on the use of pheromone traps (Negrón and Popp 2017). However, it is not always possible to see the whole picture in the forest from the limited points. Data obtained by the monitoring of target species through pheromone traps with this design (ѵ2) will help to obtain more information on beetle species and provide an insight into the prediction of possible damages caused by them. In particular, the efficient prediction of early flights (Zang et al. 2015), flight times and durations (Özcan 2017) of bark beetles are important for preventing or minimising large outbreaks (Zang et al. 2015). With the help of the designed system, it will be possible to update the control strategies for damages caused by target species in forests. Bark beetle outbreaks, which are called smokeless fires in the forest ecology, are more hazardous than fires, and it is necessary to use control programme against them. The obtained results from the studies conducted using this system design will help to create models based on the flight of target species, thus ensuring the acquisition of significant records for the prevention of possible damages.
References
Baker, T. C. (2008). Use of pheromones in IPM. In T. Radcliffe & B. Hutchinson (Eds.), Integrated pest management (pp. 273–285). Cambridge: Cambridge University Press.
Bakke, A. (1991). Using pheromones in the management of bark beetle outbreaks. In Baranchikov, Y.N., Mattson, W.J., Hain, F.P. and Payne, T. L. (Eds.), Forest insect guilds: patterns of interaction with host trees (pp. 371–377). Gen. Tech. Rep. NE-153. Forest Service, U.S. Department of Agriculture.
Byers, J. A. (1996). An encounter rate model of bark beetle population searching at random for susceptible host trees. Ecological Modelling, 91, 57–66.
Byers, J. B. (2006). Mountain pine beetle population sampling: inferences from Lingred pheromones trap and tree emergence cages. Canadian Journal of Forest Research, 36, 351–360.
Faccoli, M., & Stergulc, F. (2004). Ips typographus (L.) pheromone trapping in South Alps: spring catches determine damage thresholds. Journal of Applied Entomology, 128, 307–311. https://doi.org/10.1111/j.1439-0418.2004.00848.307.
Faccoli, M., & Stergulc, F. (2006). A practical method for predicting the short-time trend of bivoltine populations of Ips typographus (L.) (Coleoptera, Scolytidae). Journal of Applied Entomology, 130(1), 61–66.
Faccoli, M., & Stergulc, F. (2008). Damage reduction and performance of mass trapping devices for forest protection against the spruce bark beetle, Ips typographus (Coleoptera Curculionidae Scolytinae). Annals of Forest Science, 65(3), 309.
Galko, J., Økland, B., Nikolov, C., Rell, S., & Kunca, A. (2013). Comparison of pheromone traps for monitoring of the European spruce bark beetle. Oppdragsrapport Skog og landskap.
Galko, J., Nikolov, C., Kunca, A., Vakula, J., Gubka, A., Zúbrik, M., Rell, S., & Konôpka, B. (2016). Effectiveness of pheromone traps for the European spruce bark beetle: a comparative study of four commercial products and two new models. Lesnícky časopis-Forestry Journal, 62, 207–215.
Gillette, N.E.; & Munson, S. (2007). Semiochemical sabotage: behavioral chemicals for protection of Western conifers from bark beetles. In J.L. Hayes and J.E. Lundquist (compilers) (Eds.), The Western bark beetle research group: a unique collaboration with forest health protection proceedings of a symposium. Society of American Foresters Conference October 23–28, Portland, Oregon, pp. 85–109.
Gitau, C. W., Bashford, R., Carnegie, A. J., & Gurr, G. M. (2013). A review of semiochemicals associated with bark beetle (Coleoptera: Curculionidae: Scolytinae) pests of coniferous trees: A focus on beetle interactions with other pests and their associates. Forest Ecology and Management, 297, 1–14.
Gómez, D., Hirigoyen, A., Balmelli, G., Viera, C., & Martínez, G. (2017). Patterns in flight phenologies of bark beetles (Coleoptera: Scolytinae) in commercial pine tree plantations in Uruguay. BOSQUE, 38(1), 47–53.
Hansen, E. M. (2014). Forest development and carbon dynamics after mountain pine beetle outbreaks. Forest Science, 60, 476–488.
Hayes, C. J., DeGomez, T. E., Clancy, K. M., Williams, K. K., McMillin, J. D., & Anhold, J. A. (2008). Evaluation of funnel traps for characterizing the bark beetle (Coleoptera: Scolytidae) communities in ponderosa pine forests of north-central Arizona. Journal of Economic Entomology, 101(4), 1253–1265.
Holuša, J., Lukášová, K., & Lubojacký, J. (2012). Comparison of seasonal flight activity of Ips typographus and Ips duplicatus. Scientia Agriculturae Bohemica, 43(3), 109–115.
Kautz, M., Imron, M. I., Dworschak, K., & Schopf, R. (2016). Dispersal variability and associated population-level consequences in tree-killing bark beetles. Movement Ecology, 4(9). https://doi.org/10.1186/s40462-016-0074-9.
Knodel, J., Petzoldt, C. H., & Hoffman, M. P. (1995). Pheromone traps–effective tools form Lepidopterous insect pests of sweet corn. Vegetables.
Krieger, C. (1998). An overview of bark beetle control methodologies. Management Notes, 17.
Lindelow, A., & Schroeder, M. (2001). Spruce bark beetle, Ips typographus (L.), in Sweden: monitoring and risk assessment. Journal of Forest Science, 47, 40–42.
Lindgren, B. S., & Borden, J. H. (1983). Survey and mass trapping of ambrosia beetles (Coloptera: Scolytidae) in timber processing areas on Vancouver Island. Canadian Journal of Forest Research, 13, 481–493.
Lindgren, B. S., & Raffa, K. F. (2013). Evolution of tree killing in bark beetles (Coleoptera: Curculionidae): Trade-offs between the maddening crowds and a sticky situation. Canadian Journal of Forest Research, 145, 471–495.
Macek, M., Wild, J., Martin Kopecký, M., Svoboda, M., & Fischer, A. (2017). Life and death of spruce forest: ecological processes in the first 12 years after bark-beetle outbreak. In: 2nd International Conference on Forests, 26–29 April, Germany. 38 p.
Marini, L., Økland, B., Jönsson, A. M., Bentz, B., Carroll, A., Forster, B., Grégoire, J. C., Hurling, R., Nageleisen, L. M., Netherer, S., Ravn, H. P., Weed, A., & Schroeder, M. (2017). Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography, 40, 001–010. https://doi.org/10.1111/ecog.02769.
Meddens, A. J. H., Hicke, J. A., & Ferguson, C. A. (2012). Spatiotemporal patterns of observed bark beetle-caused tree mortality in British Columbia and the western United States. Ecological Applications, 22(7), 1876–1891.
Meurisse, N., & Pawson, S. (2017). Quantifying dispersal of a non-aggressive saprophytic bark beetle. PLoS One, 12(4), e0174111. https://doi.org/10.1371/journal.Pone.0174111.
Meurisse, N., Couillien, D., & Grégoire, J. C. (2008). Kairomone traps: a tool for monitoring the invasive spruce bark beetle Dendroctonus micans (Coleoptera, Scolytinae) and its specific predator, Rhizophagus grandis (Coleoptera, Monotomidae). Journal of Applied Ecology, 45, 537–548.
Nansen, C. (2004). Technology calculating pheromone trap catches. Pest Control, 36–38.
Negrón, J. F., & Popp, J. B. (2017). Can spruce beetle (Dendroctonus rufipennis Kirky) pheromone trap catches or stand conditions predict Engelmann spruce (Picea engelmannii Parry ex Engelm.) tree mortality in Colorado? Agricultural and Forest Entomology. https://doi.org/10.1111/afe.12239.
Özcan, G. E. (2017). Assessment of Ips sexdentatus population considering the capture in pheromone traps and their damages under non-epidemıc conditions. Journal of the Forestry Society of Croatia, 1–2, 47–56.
Özcan, G. E., Eroğlu, M., & Alkan-Akıncı, H. (2011). Use of pheromone-baited traps for monitoring Ips sexdentatus (Boerner) (Coleoptera: Curculionidae) in oriental spruce stands. African Journal of Biotechnology, 10(72), 16351–16360.
Özcan, G. E., Cicek, O., Enez, K., & Yıldız, M. (2014). A new approach to determine the capture conditions of bark beetles in pheromone-baited traps. Biotechnology & Biotechnological Equipment, 28(6), 1057–1064.
Özcan, G. E., Çiçek, O., Enez, K., & Yıldız, M. (2016). Evaluation of the counting success of pheromone-baited trap with electronic control unit in practice. Current Science, 111(1), 192–197. https://doi.org/10.18520/cs/v111/il/192-197.
Panzavolta, T., Bracalini, M., Bonuomo, L., Croci, F., & Tiber, R. (2014). Field response of non-target beetles to Ips sexdentatus aggregation pheromone and pine volatiles. Journal of Applied Entomology, 138, 586–599.
Raffa, K. F., Phillips, T. W., & Salom, S. M. (1993). Strategies and mechanisms of host colonization by bark beetles. In T. D. Schowalter & G. M. Filip (Eds.), Beetle-pathogen interactions in conifer forests (pp. 103–120). New York: Academic Press.
Raty, L., Drumont, A., De Windt, N., & Grégoire, J. -C. (1995). Mass trapping of the spruce bark beetle Ips typographus L. traps or trap trees? Forest Ecology and Management, 78, 191–205.
Rudinsky, J. A. (1962). Ecology of scolytidae. Annual Review of Entomology, 7, 327–348.
Sagitov, A., Mukhamadiev, N., Ashikbaev, N., Mazarzhanova, K., Bolat, Z., Mendibaeva, G., & Abzhanbayev, D. (2016). Pheromone traps application against bark beetles (Coleoptera: Scolytidae) in Ili-Alatau Mountains. Kastamonu University Journal of Engineering and Sciences, 2(1), 14–20.
Sánchez-Martínez, G., & Wagner, M. R. (2002). Bark beetle community structure under four ponderosa pine forest stand conditions in northern Arizona. Forest Ecology and Management, 170, 145–160.
Schowalter, T. D. (2012). Ecology and management of bark beetles (Coleoptera: Curculionidae: Scolytinae) in southern pine forests. Journal of Integrated Pest Management, 3(2), A1–A7. https://doi.org/10.1603/IPM11025.
Schwab, O., Maness, T., Bull, G., & Roberts, D. (2009). Modeling the effect of changing market conditions on mountain pine beetle salvage harvesting and structural changes in the British Columbia forest products industry. Canadian Journal of Forest Research, 39, 1806–1820.
Turchin, P., & Odendall, F. J. (1996). Measuring the effective sampling area of a pheromone trap for monitoring population density of southern pine beetle. (Coleoptera: Scolytidae). Environmental Entomology, 25(3), 582–588.
Wainhouse, D. (2005). Ecological methods in forest pest management. New York: Oxford University Press.
Weslien, J., Annila, E., Bakke, A., Bejer, B., Eidman, H. H., Narvestad, K., Nikula, A., & Ravn, H. P. (1989). Estimating risks for spruce bark beetle (Ips typographus (L.)) damage using pheromone-baited traps and trees. Scandinavian Journal of Forest Research, 4, 87–98.
Wood, S. L., & Bright, D. E. (1992). A catalog of scolytidae and platypodidae (Coleoptera). Part 2. Taxonomic index. Great Basin Naturalist Memoirs, 13, 1–1553.
Yonker, C. R. (1990). Pheromone trapping applications for monitoring European elm bark beetle populations. Journal of Arboriculture, 16(6), 148–152.
Zang, C., Helm, R., Sparks, T. H., & Menzel, A. (2015). Forecasting bark beetle early flight activity with plant phenology. Climate Research, 66, 161–170.
Zúbrik, M., Raši, R., Vakula, J., Varínsky, J., Nikolov, C., & Novotný, J. (2008). Bark beetle (Ips typographus L., Pityogenes chalcographus L., Col.: Scolytidae) pheromone traps spatial distribution optimisation in central Slovakian mountains. Forestry Journal, 54(3), 235–248.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Özcan, G.E., Çіçek, O., Enez, K. et al. A new design of electronic control unit involving microcontroller to determine important parameters for target species in forest. Environ Monit Assess 190, 600 (2018). https://doi.org/10.1007/s10661-018-6960-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-018-6960-4