
Abstract
With the aim of unravelling the role of airborne Fusarium graminearum inoculum in the epidemic of Fusarium head blight (FHB) caused by this species in wheat spikes, a network of Burkard air samplers was set up in five wheat fields distributed in Belgium from 2011 to 2013. Each year from April to July, the daily amounts of F. graminearum inoculum above the wheat canopy were quantified using a newly developed TaqMan qPCR assay. The pattern of spore trapping observed was drastically different per year and per location with a frequency of detection between 9 and 66% and a mean daily concentration between 0.8 and 10.2 conidia-equivalent/m3. In one location, air was sampled for a whole year. Inoculum was frequently detected from the wheat stem elongation stage until the end of the harvesting period, but high inoculum levels were also observed during the fall. Using a window-pane analysis, different periods of time around wheat flowering (varying in length and starting date) were investigated for their importance in the relation between airborne inoculum and FHB parameters (FHB severity, frequency of F. graminearum infection and DON). For almost all the combinations of variables, strong and significant correlations were found for multiple window lengths and starting times. Inoculum quantities trapped around flowering were highly correlated with F. graminearum infection (up to R = 0.84) and DON (up to R = 0.9). Frequencies of detection were also well correlated with both of these parameters. DON concentrations at harvest could even be significantly associated with the F. graminearum inoculum trapped during periods finishing before the beginning of the anthesis (R = 0.77). Overall, these results highlight the key role of the airborne inoculum in F. graminearum epidemics and underline the importance of monitoring it for the development of disease forecasting tools.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fusarium head blight (FHB) of wheat is a disease frequently occurring in Europe and throughout the world. This fungal disease is caused by numerous species belonging to the Fusarium and Microdochium genera. Among these species, F. graminearum (teleomorph: Gibberella zeae [Schwein.] Petch) is often reported as one of the predominant species threatening grain yields and sanitary quality because of its capacity to produce mycotoxins (Osborne and Stein 2007). This species can also infect other small-grain cereals such as barley and oats, as well as maize and various weeds (Leplat et al. 2012; Osborne and Stein 2007; Sutton 1982). Deoxynivalenol (DON), one of the major mycotoxins produced by F. graminearum, is frequently found in harvested grains (Desjardins 2006), and its presence in grain lots in Europe (EC Directive No. 1881/2006) and in other parts of the world is regulated. In order to control the disease and the subsequent DON concentration in grains, fungicides, preferably composed of triazoles, can be applied to the ears, with the best results being obtained by treatments at mid-anthesis (Paul et al. 2008). However, the need for fungicide applications to control FHB is not always obvious, as the severity of the disease and the DON concentration in grains can vary greatly between years and locations (Chandelier et al. 2011; Hellin et al. 2016; Xu et al. 2005).
The inoculum of F. graminearum is primarily composed of macroconidia and ascospores (Osborne and Stein 2007). The former are asexually produced by F. graminearum mycelia present in the fields and can be splash-dispersed onto the ears during rain events (Paul et al. 2004). On the other hand, ascospores are produced in perithecia as a result of sexual crossing and can be forcibly discharged into the air (Trail et al. 2005). Because of this characteristic and their relatively light weight (19–24 × 3–4 μm, two-celled spore) as compared to conidia (25–50 × 3–4 μm, 4–8 cells spore), ascospores can be transported over long distances and therefore originate either within the field or from distantly located sources (Booth 1971; Keller et al. 2014; Maldonado-Ramirez et al. 2005; Prussin et al. 2014a). Nevertheless, both types of spores can constitute an airborne inoculum (Keller et al. 2014). The most important inoculum reservoirs are overwintered crop residues (Osborne and Stein 2007; Sutton 1982). Cereal and maize residues are a highly suitable substrate for the production of perithecia (Leplat et al. 2012), and F. graminearum can survive on these residues for more than two years if they are left on the soil surface (Pereyra et al. 2004). For these reasons, maize-wheat rotation, commonly performed in Europe, has a particularly negative impact when it comes to FHB. Good tillage practices are therefore generally performed in order to bury the residues and facilitate their decomposition (Dill-Macky and Jones 2000). However, the contribution of distant airborne inoculum may be more important than that of the field inoculum, as suggested by numerous authors (reviewed in Keller et al. 2014).
Fungal spores can be transported onto the ear during rain events, either by splashing of local inocula or by a washout of spores in suspension in the air. Airborne inoculum can also be deposited on the ear by gravitational settling (Keller et al. 2014). As soon as the ear starts to emerge from the plant (GS50), fungal spores can land on it and theoretically initiate the infection. Wheat spikes are most susceptible to FHB pathogens around mid-flowering but infections have been shown to be still possible during grain maturation (Del Ponte et al. 2007; Yoshida and Nakajima 2010). The epidemiological cycle of F. graminearum is also highly influenced by weather conditions (Osborne and Stein 2007; David et al. 2016). The production of DON by this species in grain is also favoured by warm and humid conditions after infection (Cowger et al. 2009; Kriss et al. 2012; Ramirez et al. 2006). As a consequence, extensive work has focused on relating the meteorological conditions occurring in fields to the subsequent disease level (De Wolf et al. 2003; Kriss et al. 2010, 2012; Xu 2003). Numerous forecasting tools have been elaborated, mostly using descriptive models based on weather data as well as risk factors associated with cultural practices, to predict FHB and DON (Prandini et al. 2009; Xu 2003). However, one of the possible flaws of currently available models is that they lack knowledge of the actual inoculum presence.
The airborne inoculum of F. graminearum has been previously monitored in several studies through its capture by repetitive exposure of selective media (Del Ponte et al. 2005; Maldonado-Ramirez et al. 2005; Nicolau et al. 2012; Prussin et al. 2014a; Schmale et al. 2005) or microscope slides (Manstretta et al. 2015) to the air, or using volumetric air trapping devices (David et al. 2016; de Luna et al. 2002; Inch et al. 2005; Hofgaard et al. 2016; Markell and Francl 2003; Prussin et al. 2014b; Rossi et al. 2002). Inoculum deposited on wheat ears has also been collected by washing the surface of the spikes and collecting the run-off (Markell and Francl 2003; Paul et al. 2007). The amount of spores can then be estimated by visually counting spores or CFUs (David et al. 2016; de Luna et al. 2002; Del Ponte et al. 2005; Inch et al. 2005; Maldonado-Ramirez et al. 2005; Manstretta et al. 2015; Markell and Francl 2003; Nicolau et al. 2012; Paul et al. 2007; Prussin et al. 2014a, b; Rossi et al. 2002; Schmale et al. 2005), or using molecular quantification methods such as qPCR (Hofgaard et al. 2016). Monitoring of F. graminearum airborne inoculum is usually performed around anthesis, since it is considered as the critical period for ear infection. At a given moment, the inoculum deposition of F. graminearum has been shown to vary between years and locations (Hofgaard et al. 2016; Paul et al. 2007; Schmale et al. 2005) and even to be spatially heterogeneous at the field scale (Schmale et al. 2005). This highly dynamic deposition of spores is influenced by meteorological conditions such as rain events (Del Ponte et al. 2005; Markell and Francl 2003; Paul et al. 2007; Rossi et al. 2002).
The quantity of F. graminearum propagules found on wheat spikes during heading correlates positively with the final concentration of DON (Stein and Osborne 2006). Moreover, in controlled environments, the quantity of spores used to inoculate wheat ears is positively correlated with the subsequent FHB symptoms and concentration of DON in the grain (Stein et al. 2009). Yet the relation between the trapped airborne inoculum and both symptoms and DON concentrations is not obvious in the field (Schaafsma et al. 2005). Due to the relatively high labour- and cost-intensive aspect of airborne inoculum monitoring procedures, the number of observations and surveyed situations (# location-years) is usually low, and most studies have focused on explaining the inoculum occurrence rather than its association with the disease.
Spore traps combined with qPCR have been more and more used with the aim of studying the airborne inoculum of various plant pathogens and improving precision in disease risk management (Almquist and Wallenhammar 2015; Chandelier et al. 2014; Duvivier et al. 2013, 2016; Luchi et al. 2013; Rogers et al. 2009; West et al. 2008; Wieczorek et al. 2014). In the current context of integrated pest management established in Europe (Directive 2009/128/EC), methods need to be developed to monitor plant pathogens, forecast disease incidence and reduce pesticide use. A better knowledge of the importance of airborne F. graminearum inoculum in its epidemiological cycle could strengthen the output of current prediction models and ideally provide better advice to farmers on the relevance of a fungicide treatment. The survey of F. graminearum inoculum was therefore undertaken as part of a larger project on the airborne inoculum of wheat fungal pathogens in Belgium (Duvivier et al. 2013, 2016). In the study reported here, a robust monitoring procedure of airborne F. graminearum inoculum in wheat fields was developed by combining the automation of vacuum spore traps and the specificity of TaqMan qPCR. The method was used in a network of trial fields in Belgium to study the temporal distribution of airborne F. graminearum. The final goal was to determine the level of association of airborne F. graminearum inoculum with several FHB parameters (e.g. FHB severity and DON) over multiple periods of time surrounding wheat anthesis, based on the window-pane analysis methodology developed by Kriss et al. (2010, 2012). The study demonstrates the potential of spore trapping to predict the risk of F. graminearum epidemics and DON content.
Material and method
Field trials
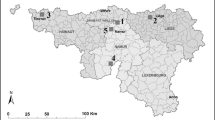
For three years (2011 to 2013), five trial wheat fields distributed in the Belgian Walloon region (Fig. 1a) were surveyed as previously described (Hellin et al. 2016). These fields were located in Perwez, Voroux-Goreux, Tournai and Niverlée, and additional fields were surveyed in Aye (2011), Naast (2012) and Aisemont (2013). These latter fields had maize as the preceding crop, whereas the previous crops in the other fields were minor Fusarium spp. hosts such as sugar beet and oilseed rape. An FHB-susceptible wheat cultivar (cv. “Lexus” in 2010–11 and “Viscount” in both 2011–12 and 2012–13 growing seasons) was sown during the last two weeks of October except in Tournai (2011) and Voroux-Goreux (2013), where it took place on 20 November 2010 and 14 November 2012 respectively. In each field, four untreated wheat plots (1.5 × 10 m) approximately 5 m apart from each other were used in this study. The precise date of the beginning of wheat flowering (GS61; Zadoks et al. 1974) was recorded in each field. At the end of the milk stage (GS77), the percentage of the ear surface showing typical bleached symptoms was assessed in each plot by observing 100 randomly selected ears. The same field network was used by Hellin et al. (2016), who quantified the presence of Fusarium spp. in mature grains (i.e. one week before harvest) by fungal isolation as well as the concentration of DON at harvest in two of the four wheat plots. As well as FHB symptoms, these data were used in this study as parameters to characterize FHB in the fields.
Air sampling
In each field, a Burkard 7-day spore-monitoring trap (Burkard Manufacturing Co. Ltd., Rickmansworth, Hertfordshire, UK) was set up and air trappings were conducted from 15 April to 15–31 July of each year (2011 to 2013), depending on the location (Table S1). In Tournai, a whole-year analysis of the airborne inoculum was performed by collecting the inoculum present from 1 September 2011 to 31 August 2012. The spore trap openings were always placed 1 m above ground level in a 9 m2 wheat-free zone located from 3 to 40 m from the surveyed field plots. Airborne spores vacuumed by the traps were collected onto a slowly rotating Melinex tape covered with Vaseline. The throughput of each spore trap was regulated at 10 l/min, corresponding to 14.4 m3 every 24 h. Each week, the tape was collected from each location and cut into seven equal parts (48 × 20 mm) corresponding to daily fragments (midnight to midnight).
DNA extraction and quantification
Each daily tape segment was placed in a 2 ml microtube for total DNA extraction, using the method developed and validated by Duvivier et al. (2013). A set of primers and probe specific to F. graminearum was designed using Beacon Designer v 7.9 (Biosoft International, Corina Way, CA, US) for a real-time PCR Taqman reaction (Fgram-for: CCCTGTTCGAGGCTATCTTCAA; Fgram-rev: GTCAATCTCCATTTGATCCGTGA; Fgram-probe: Texas Red-CAACGCCCAGCCAACATCGACACAA-BHQ2). The target region consisted of a 119 bp fragment located in the Fg16 gene (accession No. FGRAMPH1_01T03509) coding for a protein of unknown function. This gene has been previously used for the specific detection and quantification of F. graminearum (Nicholson et al. 1998; Waalwijk et al. 2004). Real-time PCR protocol was optimized using variation of product quantities, as well as thermal cycle steps (temperature and duration). The final mixture used for the real-time PCR assay was composed of 12.5 μl of iTaq Universal Probes Supermix (Biorad, Hercules, California), 300 nM of each primer, 500 nM of probe and 2.5 μl of DNA extract in a total volume of 25 μl. The reaction was conducted in a CFX96 Real-Time thermocycler (Bio-Rad, Hercules, CA, USA) as follows: initial denaturation at 95 °C for 3 min, and 40 cycles each, a denaturation step at 95 °C for 5 s and a hybridization and elongation step at 60 °C for 30 s. The assay was evaluated using a fivefold dilution series (starting at 50 ng/μl) of pure F. graminearum DNA (extracted from cultured mycelium); each dilution was tested in triplicates in three separate PCR runs. The coefficient of variation (CV) was used on this set of repeated measurements to determine the overall inter- and intra-plate variability.
The efficiency of each of the subsequent PCR runs was evaluated with a standard curve obtained from four tenfold serial dilutions of pure F. graminearum DNA extract. Each sample was run in two replicates. Across all plates, the concentrations of the standard dilutions were always based on a similar DNA starting quantity, measured with a NanoDrop 1000 spectrophotometer (Thermo Scientific). Data were collected and analyzed using CFX Manager (Bio-Rad).
Specificity and sensitivity of the quantification method
The specificity of the primers and probe was assessed using BLASTN and primer-BLAST (Altschul et al. 1990; Ye et al. 2012). The specificity of the method was also tested using reference strains of F. graminearum (PH1 syn. NRRL 31084, MUCL 43802 and MUCL 53452), F. pseudograminearum (MUCL 6131), F. culmorum (UK99 syn. NRRL 54111, MUCL 43796, MUCL 43797 and MUCL 43798), F. avenaceum (MUCL 55415), F. poae (MUCL 42824, MUCL 42836 and MUCL 42842), Zymoseptoria tritici (MUCL 45550 and MUCL 45549), Staganospora nodorum (MUCL 30165, MUCL 44704 and MUCL 44707), Microdochium nivale (MUCL 15949 and MUCL 31963), Oculimacula yallundae (MUCL 40386 and MUCL 40387), O. acuformis (MUCL 40388, MUCL 40389 and MUCL 40637), and Sclerotinia sclerotiorum (MUCL 11553 and MUCL 30163), as well as isolates of Puccinia striiformis and P. triticina collected on wheat plants in the fields. Furthermore, the specificity of the primer was confirmed by sequencing (Macrogen, Netherlands) the PCR product from 15 positive samples originating from the three-year study (one sample per field and year), and comparing these sequences with the expected DNA sequence. In order to estimate the sensitivity of the real-time PCR to effectively quantify conidia trapped on tape fragments, a macroconidia suspension was prepared from F. graminearum MUCL 43802. The strain was cultivated on synthetic low-nutrient agar (SNA) for two weeks (12 h light/darkness, 22 °C) and macroconidia were collected using 0.1% Nonidet P40. The suspension was filtered through a fine mesh (pore size = 100 μm) and serially diluted five times by a tenfold factor. The actual concentrations of conidia in each dilution were measured using a Thoma haemocytometer. A volume of 50 μl of each conidial suspension was placed in a 2 ml microtube with a piece of clean tape covered with Vaseline. DNA from these artificially spore-coated tapes was extracted and quantified by real-time PCR as described above. The test was replicated twice. The CT values were plotted against the logarithm of the starting quantity of each spore suspension in order to create a calibration curve. Data are always expressed in numbers of “conidia-equivalents” on a daily fragment of tape (conidia/day).
Window-pane analysis and multiple testing
In order to capture the effect of the airborne inoculum (mean inoculum quantity [conidia/day] and frequency of days with inoculum detection [%] during a selected period of time) on the resulting FHB parameters (i.e. symptom severity [%], frequency of F. graminearum isolation from grains [%] and DON concentration [μg/kg]), a window-pane analysis was performed across all years and locations. This type of data mining procedure has already been used by different authors to explain FHB parameters with meteorological variables (Kriss et al. 2010, 2012). In this analysis, inoculum data are summarized over different time intervals of specified length (window) surrounding flowering for each location and year. Multiple window lengths were used in this study, ranging from 10 to 40 days with a 5-day increment. The starting date of each window ranged from −27 to 11 days (1-day increment) from the flowering start date (considered as day “0”). A time window can therefore be considered as a sliding frame of a given size, with two successive time windows sharing all but one day (Kriss et al. 2012). Pearson’s correlations between the summarized inoculum data and FHB parameters (collected in the corresponding fields) are then computed, separately for each combination of window length and starting time. Windows could not start earlier due to missing inoculum data in Aisemont in 2013 before this date. Moreover, windows that extended more than 21 days after the beginning of anthesis were not considered, since no inoculum data were available in Aisemont (2013) after this date. Inoculum data from each year and location were computed separately and were considered as independent. The average daily inoculum quantities during each period were log-transformed [Y = log10(X + 1)] to approximate a normal distribution. Data expressed as percentages (e.g. inoculum frequency and FHB severity) were first arcsine square root-transformed. For each combination of window length and starting time, the normal distribution of each summarized inoculum data across all years and locations was verified using the Shapiro-Wilks test. Multiple hypothesis testing is likely to produce excessive numbers of correlations being wrongly considered as significant. For example, for a 10-day period, 44 correlations are computed for each combination of inoculum and FHB parameters. Adjustments were made to control the proportion of false positives without introducing too many false negatives through a three-step procedure, as previously well described in Kriss et al. (2010, 2012). Firstly, for a given window length, a global significance test across all window starting times was computed for every combination of inoculum and FHB parameters using Simes’s method (Simes 1986). The aim of this procedure was to determine if a particular combination of variable showed at least one significant correlation among the tested starting times. Significance was achieved if the adjusted global P value was smaller than 0.05. Secondly, the significance level of individual correlation was fixed at 0.005 instead of 0.05 to prevent excessive numbers of false positives for the individual correlation results. Finally, in order to be relevant, an individual strong correlation was only declared significant if it was part of a group of at least five successive significant (P < 0.005) correlations, for a given window length and inoculum-disease relationship, since a single strong correlation surrounded by weak correlation was likely to be a false positive. This ad hoc method was first introduced by Pietravalle et al. (2003).
Results
Quantification of FHB importance
The severity of FHB symptoms was scored each year in every field (Fig. 1). According to the ANOVA, significant differences of severity were observed between the years (P < 0.001) and locations (P < 0.001), when considering the four locations that were surveyed every year. Overall, the mean severity of FHB at all sites was similar between 2012 (5.7%) and 2013 (7.7%), whereas it was lower in 2011 (1.9%). The percentage of grain infected by F. graminearum (and other Fusarium spp.), previously estimated in Hellin et al. (2016), was also variable between years and locations (Table S2). F. graminearum was particularly predominant in 2012 and 2013, and its frequency of isolation measured in mature grains was correlated (Spearman’s ρ = 0.77, P < 0.001) with DON concentrations measured in the corresponding grain lots (Table S2; Hellin et al. 2016). Yet in 2011 at Aye, the DON content in grains was extremely high even though a low incidence of F. graminearum was found. This was due to the high frequency (37.2%) in this field of infections caused by F. culmorum, which is also known to produce DON (Hellin et al. 2016). For this reason, these data were ignored in the other analysis concerning DON concentration. In all other fields, the frequency of grains infected by F. culmorum was always lower, with an average of <1%. On the other hand, severity of FHB symptoms was correlated neither to the percentage of grains infected by F. graminearum (P = 0.197) nor to DON concentrations (P = 0.952).

Field network surveyed from 2011 to 2013 in the South of Belgium. a Regions where the surveyed fields were located. Locations are numbered: 1 = Perwez, 2 = Voroux-Goreux, 3 = Tournai, 4 = Niverlée, 5a = Aye (2011), 5b = Naast (2012) and 5c = Aisemont (2013). b Fusarium head blight (FHB) symptom severity in the field network. Error bars correspond to standard error calculated on four replicates
Evaluation of the quantification method
The primers and probe designed for F. graminearum quantification showed high specificity when tested with the BLAST tools. The reaction did not produce any fluorescent signal for any other tested species. Furthermore, the sequences of the PCR fragments obtained after amplification of a subset of positive air samples all matched to the F. graminearum reference sequence with at least 99% nucleotide identity.
The qPCR assay displayed a high linearity of response signals over the F. graminearum DNA concentrations tested, highlighting the broad dynamic range of the assay (125.000 to 1.6 pg; Fig. 2). When the following dilution corresponding to 0.32 pg in the reaction was included, a signal could be obtained for some, but not all reactions. The coefficients of variation of inter- and intra-plate variability calculated on CT values from this data set were both lower than 1% (CV = 0.64 and 0.69% respectively), indicating the high repeatability of the qPCR assay. The efficiency of the reaction was always between 90 and 110% (mean = 99.3%), with R2 = 0.99 for about 50 assays performed.

Linear regression between the log-transformed concentrations of F. graminearum DNA from pure culture and the cycle threshold (CT) values from three qPCR runs performed with triplicates. Error bars represent the standard deviation of CT values between runs
Based on known concentrations of F. graminearum macroconidia deposited on tape fragments from which DNA was extracted and quantified as regular samples, the following equation was obtained and used to approximate the relation between CT (X) and conidia quantity (Y): Y = 10–0.31X + 13.37. Both variables were highly and significantly correlated (R2 = 0.98 and P < 0.001). The DNA of F. graminearum that had been extracted from ~28 conidia on the daily tape fragment (1.9 conidia/m3) was detected in the two extracted samples (one detection out of two replicates in the second sample) (Table 1). This is therefore considered as the detection limit of the method.
Temporal distribution of airborne inoculum
The developed qPCR method was therefore used to quantify the presence of F. graminearum particles in the samples collected in the spore-trap network. The year-long sampling period that took place in Tournai from 1 September 2011 to 31 August 2012 showed that the airborne inoculum of F. graminearum was present in the field during the entire year (Fig. 3). A total of about 55,000 conidia-equivalents were detected in the field during this period (daily average of 10.5 conidia-equivalents/m3). Inoculum peaks with concentrations as high as 7300 conidia-equivalents/day (507 conidia/m3) were recorded in the month of September. High peaks were also recorded in October, but their intensity started to decrease at the end of this month. A cluster of detections also occurred at the end of November. From then to mid-April, the inoculum detections were more sporadic, consisting of about two to three detections a month of around 50 of conidia-equivalents (3.5 conidia/m3) or less. From stem elongation (GS30), the frequency of peaks gradually started to increase again and their intensities reached up to 130 conidia-equivalents/day (9.0 conidia/m3) at the end of June (i.e. end of flowering). During the harvest period, tremendous amounts of inoculum were then recorded again, starting in the middle of July with concentrations up to more than 5000 conidia-equivalents/day (347.2 conidia/m3). High inoculum peaks occurred at both sowing and harvest dates/periods (Fig. 3).

Year-long daily distribution of airborne F. graminearum inoculum (conidia/day) in Tournai from 1 September 2011 to 31 August 2012). Wheat phenology, recorded in the field, is shown at the bottom of the figure following Zadoks’ scale (GS11 = first leaf emerged, GS30 = beginning of stem elongation, GS32 = second node detectable, GS39 = flag leaf fully emerged, GS61 = beginning of flowering and GS69 = end of flowering). Typical sowing and harvest periods in Belgium are indicated by black lines and the asterisks show the exact date of both events in this field. The period during which airborne inoculum was recurrently monitored during the multi-year/location survey is indicated on the graph (15 April to 15–31 July)
Regarding the pattern of detection obtained between 15 April and 15 July for the whole spore-trap network, the quantities of conidia-equivalents collected differed between years and locations (Fig. 4, Table S1). Over this period, the frequency of days with inoculum detections varied between 9 and 66%, and the mean daily concentration between 0.8 and 10.2 conidia-equivalents/m3. The quantities of spores ranged from 0 to 2298 conidia-equivalents/day (159.6 conidia/m3) during the whole survey period. In 2011, F. graminearum inoculum was generally detected more sporadically and later during the season than during the following years. However, the frequencies of inoculum detection intensified in Niverlée and Aye around the second week of June. Detection frequencies calculated for the whole survey period ranged from 8.8% in Perwez to 31.9% in Niverlée. The maximum of cumulated conidia-equivalents that year (5857 conidia-equivalents) was also recorded in Niverlée. In 2012, the frequencies of inoculum peaks rapidly increased around May and the inoculum during the season was more abundant than in 2011, particularly in Voroux-Goreux and Naast. In these two locations, inoculum frequencies for this period reached 66.3 and 64.1% respectively, and the cumulative sum of inoculum was the highest (13,566 and 10,634 conidia-equivalents respectively). In 2013, more detections occurred in the early period of sampling (April), although data on this period were missing for Aisemont. The detection frequencies and the cumulative sums of inoculum were similar between locations (around 35% and 1250 conidia-equivalents respectively). The fields with maize as preceding crop (see “5” in Fig. 4) did not show a specific pattern with a clear difference in terms of number of detections or global quantity trapped.

Temporal distribution of F. graminearum inoculum from 15 April to 15 July in the five fields surveyed from 2011 to 2013. For each year, the upper graph represents the cumulative inoculum concentrations in each field. Under this are shown the daily detections of the inoculum in each field, with each bar representing an inoculum detection for a particular date. Fields are numbered from 1 to 5 as explained in the legend. The fifth field corresponds to Aye, Naast and Aisemont in 2011, 2012 or 2013 respectively. The date of anthesis in each field is indicated by a grey bar. “No Data” indicates periods during which inoculum data are missing
Correlations between F. graminearum inoculum and FHB parameters
Through the window-pane analysis, 203 variables for each inoculum quantity and frequency were generated. Of these 406 inoculum variables, only seven did not follow a strict normal distribution, as shown by significant P-values for the Shapiro-Wilks test, and most of them (n = 5) were obtained for 10-day window lengths. Overall, both inoculum detection frequency and quantity could be significantly correlated to symptom severity [%], frequency of F. graminearum isolation [%] and DON concentration [μg/kg]. Across all window starting times, the global significance level (based on Simes’s method) of the correlation for every possible combination of variables and window length was below the α = 0.05 level, except for the correlation between FHB severity and inoculum frequency for the 10-day windows (P = 0.06). Therefore, for each combination of variables, at least one correlation per window length can be considered as truly significant.
The importance and significance level of correlations between inoculum variables (quantity and frequency) computed for different window times around anthesis and FHB parameters are summarized in Table 2. For each combination of variables and window length, the highest Pearson’s correlation coefficient among all window starting times is given. Higher correlations were generally obtained when using the inoculum quantity rather than the frequency of inoculum detection (Table 2). DON concentration was the disease parameter that overall had the highest correlation with inoculum variables (up to 0.9), followed closely by the frequency of isolation of F. graminearum strain (Fgram). On the other hand, correlations between the inoculum and the disease severity were lower (up to 0.75). The highest coefficients for DON and Fgram were obtained with inoculum data summarized over windows covering most of the flowering period. Conversely, the highest significant relations between FHB severity and inoculum quantity were found with a window covering only a small part of the flowering period. In order to be relevant, a correlation between two variables for a single window starting time should not only be individually significant but also be part a cluster of five successive significant correlations (P < 0.005). For the highest correlations including DON and Fgram, this condition was generally achieved for all window lengths. Clusters of significant correlations were more scarcely found when using FHB severity. The presence of correlations obtained from time windows ending before the anthesis and that were part of such clusters was identified (indicated by asterisks in Table 2). Such correlations could be helpful in order to predict FHB parameters.
Individual Pearson’s correlation coefficients obtained from the window pane analysis for two window lengths (15 days and 30 days) were plotted in the form of bar charts (Fig. 5). These two window lengths were chosen because they gave a good representation of the overall data for the different window lengths and highlighted the most interesting results. Each vertical bar represents a single estimated Pearson’s correlation coefficient between two variables (e.g. DON and inoculum quantity), while the starting time of each window is indicated on the horizontal axis, 0 indicating the beginning of anthesis.

Pearson’s correlation coefficients between F. graminearum inoculum quantity (a) and frequency of days with inoculum detection (b) over selected periods of time around wheat flowering and FHB parameters (DON quantity, F. graminearum isolation frequency and FHB symptom severity). Inoculum variables are computed during periods (windows) of 15 and 30 days (d) starting at different moment (in days) from anthesis (day “0”). The quantity of inoculum is expressed as the log of the average number of conidia-equivalents collected during a period, whereas the frequency is the arcsine square-root transformation of the inoculum detection frequency. In contrast to grey bars, bars in black represent correlations significant at P < 0.005
For the 15-day window length, the succession of Pearson’s correlations calculated between DON concentration and both inoculum parameters (i.e. quantity and frequency) showed a similar pattern. Two main clusters of high and significant coefficients (P < 0.005) were identified for both parameters, the first one starting well before anthesis, the second around anthesis. For a window length of 30 days, only one large cluster of significant correlations was identified for both parameters. Significant correlations with DON occurred at an earlier starting time when the frequency of inoculum detection was used than when its quantity was used.
Concerning the relationship between the frequency of F. graminearum strain isolation at harvest and the airborne inoculum present during the growing season, one main cluster of significant correlations was generally observed for the four combinations of inoculum parameters and window length. Patterns of coefficients were very similar between quantity and frequency, with larger clusters when inoculum quantity was used. Significant correlations were observed at the beginning of anthesis in the case of the 15-day window length, and up to 21 days before when a 30-day length window was used.
Relationships between FHB symptoms and inoculum variables were generally poorer and less significant coefficients were identified regardless of the window length. Only one large cluster could be found with the quantity of inoculum measured in a 30-day period.
Detailed regressions were plotted for two strong and significant relationships of interest obtained between DON and inoculum parameters. In Fig. 6a, the highest relationship, explaining more than 80% of the variation of DON content, is presented; it was obtained with the quantity of inoculum measured on a period of 20 days starting 1 day after the beginning of anthesis as the explanatory factor. Inoculum frequency measured in a window of 15 days that finished 5 days before the end of anthesis explained 60% of the DON concentration (Fig. 6b). At 15 days prior to the beginning of anthesis, wheat was generally at the flag leaf stage (GS 39), as observed in the fields during this survey.

Relationships between DON concentration and F. graminearum inoculum quantity (a) and frequency of days with inoculum detection (b) over selected periods of time. a The inoculum quantity (log-transformed) was measured during a 20-day period starting 1 day after the beginning of wheat flowering. b The inoculum frequency (arcsine root-transformed) was measured during a 15-day period starting 20 days before the beginning of wheat flowering. The numbers refer to the location of the field during each year (2011 to 2013) with 1 = Perwez, 2 = Voroux-Goreux, 3 = Tournai, 4 = Niverlée and 5 = Aye (2011), Naast (2012) or Aisemont (2013)
Discussion
In this study, a method for monitoring airborne F. graminearum inoculum in the field was elaborated by combining the use of Burkard 7-day air samplers and qPCR. The developed TaqMan qPCR assay showed high specificity and sensitivity for F. graminearum DNA and could detect DNA concentration from daily air samples (14.4 m3 of air) corresponding to as little as 1.4 conidia-equivalents per reaction (i.e. 28 conidia/day or 1.9 conidia/m3). This is of the same order of magnitude as the detection threshold of the method developed for F. graminearum by Hofgaard et al. (2016), which combines the use of cyclone samplers with the TaqMan qPCR from Waalwijk et al. (2004). Given that F. graminearum macroconidia contains from 4 to 8 cells as compared to 4-cell ascospores (Booth 1971), this threshold could be increased to 56 ascospores/day (3.9 ascospore/m3), if the airborne inoculum trapped is only composed of ascospores. This method allowed precise daily detection and homogenous quantification of the airborne inoculum of F. graminearum in a total of 15 fields from 2011 to 2013, which had the advantage of presenting contrasting situations in terms of FHB severity, F. graminearum infection frequency and DON contamination. A homogenous data set of daily quantities of airborne F. graminearum inoculum of 1304 inoculum measurements was obtained. This may be valuable for subsequent epidemiological analysis: recent studies combining real-time PCR and spore trapping have shown that these tools can be useful in gaining a better understanding of the epidemiology of arable crop pathogens (Duvivier et al. 2013, 2016; Rogers et al. 2009; Wieczorek et al. 2014).
Thanks to our approach, we established that the airborne inoculum of F. graminearum around wheat flowering was highly variable between years and locations scattered in Belgium. However, an annual trend could be observed, with some years characterized by higher frequencies of detection than others (e.g. 2011 vs 2012). No clear differences of inoculum quantities were observed between years. Although the total quantities of spores trapped in the five sites were similar in 2013, this was not the case during other years, with up to 10-fold differences observed between fields in 2012. Also, and as observed in other studies, the amounts of inoculum of F. graminearum in a field evolved during the growing season (Fernando et al. 2000; Nicolau et al. 2012; Paul et al. 2007). However, considering the broad diversity in sampling and quantification methods (Del Ponte et al. 2005; Hofgaard et al. 2016; Paul et al. 2007), comparing our results with other studies in terms of spore counts is hazardous. Interestingly, in some locations sampled in our study, the frequency and quantity of trapped inoculum seemed to increase over time from mid-April. This inoculum increase could come from the gradual building and dispersal of F. graminearum inoculum from its already developed alternative hosts (such as weeds), or from the maturation and release of ascospores from crop residues under the increasingly favourable weather conditions (Osborne and Stein 2007). In a survey performed in Norway in 2011 and 2012 (from May to September), weekly concentrations of F. graminearum DNA also seemed to increase over time (Hofgaard et al. 2016).
Our results showed that the inoculum of F. graminearum was present not only around the flowering period but also throughout the year, as observed in Tournai from 2011 to 2012. What is particularly intriguing is the large amount of F. graminearum DNA detected in September and October 2011 and from July to the end of August 2012. One of the possible explanations for these high peaks of inoculum is the consequence of farmers’ activity in the fields. The wheat harvest in 2012 took place in the last two ten-day periods of August and could have generated large amounts of dust that was likely to contain fungal particles. In the case of maize, another highly susceptible host of F. graminearum, the harvest usually spans the period from September to November. Other agricultural practices, such as soil preparation, are also likely to pulverize and disperse contaminated soil particles. Although these higher quantities of F. graminearum were only recorded in a single location during this period, similar patterns of airborne inoculum distribution were observed in the same period for other wheat pathogens, namely Z. tritici (Duvivier et al. 2013), P. triticina (Duvivier et al. 2016) and P. striiformis (Duvivier et al., in preparation), in the same field network. Using a Burkard 7-day sampler, inoculum peaks of over 14,000 F. graminearum ascospores per day were also detected in mid-July and August in Canada (Inch et al. 2005). In any case, these inoculum peaks of high magnitude occurring after the end of the flowering period might contribute to pathogen dissemination on a large scale.
The airborne inoculum of F. graminearum can originate from different sources, including wheat and maize residues, but also from multiple weeds commonly found in Belgium (Landschoot et al. 2011). At this point, it is still not known what the main source of the measured inoculum in Belgian fields is, or whether it comes from a local (i.e. within field) or more distant origin. Manstretta et al., (2015) have highlighted a vertical gradient of inoculum frequency, moving upwards from the ground to the wheat ears in fields covered with infected maize debris. However, the three fields that had maize as the preceding crop in our study did not show a particularly different level of inoculum from the field previously sown with crops less sensitive to Fusarium spp. infections. Multiple studies investigating the effect of maize residues and ploughing have also concluded that spores from distant origin may constitute the main inoculum for the infection (reviewed in Keller et al. 2014). For example, after analysing the genetic diversity of the F. graminearum population occurring on wheat ears in fields containing maize residues contaminated with characterized F. graminearum clones, Keller et al. (2010) found that more than 80% of ear infections were due to different F. graminearum strains. Comparing the genetic diversity of F. graminearum populations from each possible inoculum reservoir (e.g. field residues, weeds and air) and to the final population occurring on wheat ears could help resolve these issues and improve our strategies of inoculum management.
The inoculum quantity and frequency in 15 different situations surveyed in this study (five fields during three years) were compared, in multiple time periods of different lengths (10–40 days) and starting date surrounding the beginning of wheat flowering through a window pane analysis (Kriss et al. 2010, 2012), to investigate the existence of potential correlations with disease parameters (i.e. FHB severity, F. graminearum infection and DON contamination). Strong correlations could generally be observed for all disease parameters and window lengths. Nevertheless, shorter window lengths are preferred due to their more practical and biologically relevant nature in the context of FHB. For this reason, a 5-day period was also originally investigated but the generated inoculum variables generally failed to follow a normal distribution (Shapiro-Wilk’s test rejected in about 40% of the time for both inoculum frequency and quantity). Correlation of the airborne F. graminearum inoculum with FHB severity was generally low and decreased drastically from the beginning of wheat flowering. This lack of association could be due to the fact that F. graminearum is not the only species generating bleached symptoms on ears. The decrease in correlation with time suggests that primary infections generating the symptoms observed in the fields at GS77 might have been caused by an inoculum that was present in the air before anthesis and that subsequent deposition of inoculum might not further increase the severity of the symptoms.
Interestingly, correlations between airborne inoculum and both DON and F. graminearum infection frequency were particularly high. The association of both variables with the inoculum quantity or frequency followed a similar pattern. This matches previous results, indicating that DON contents are linked to the quantities of F. graminearum present in the same grain lots (Hellin et al. 2016; Waalwijk et al. 2004). For example, for the 15-day window length, two clusters of significant correlations were observed. The first cluster might correspond to the cluster of significant correlations already observed for FHB symptoms. In fact, the quantities of inoculum found on spikes before flowering have already been linked to the final DON concentrations (Stein and Osborne 2006). The second cluster has a more straightforward explanation, as F. graminearum infections are mostly reported to occur from the beginning of the flowering period, and are still possible later in the season (Del Ponte et al. 2007; Xu 2003; Yoshida and Nakajima 2010). More infectious propagules deposited on the ears would theoretically lead to more grains being contaminated at the time of harvest and more DON accumulated in grain lots (Stein and Osborne 2006).
Our study clearly highlights the importance of the timing and duration of the inoculum monitoring period in explaining/predicting the disease parameters. In a previous study, the quantity of F. graminearum inoculum trapped in the air once at anthesis was not related to FHB symptoms or DON concentration (Schaafsma et al. 2005). We observed that the frequency of inoculum detection during a 15-day period starting around the flag leaf stage (GS39, about 20 days before the beginning of flowering) seemed of particularly high potential as a predictive factor for DON concentration in grains at harvest. The fact that the frequency of inoculum detection before the beginning of anthesis correlates well with DON concentration at harvest gives hope for the use of inoculum monitoring as a possible tool to forecast F. graminearum epidemics and give advice on fungicide application. Monitoring of meteorological conditions around wheat flowering has also provided key factors for the prediction of the disease (Kriss et al. 2012; Prandini et al. 2009), and the use of weather-based variables should be integrated in a predictive model. Additional inoculum data, particularly inoculum data in conjunction with high DON concentrations (>1250 μg/kg), which are lacking in our study, are also needed to validate a model. Fusarium culmorum, another FHB pathogen that produces DON, is sometimes reported (Chandelier et al. 2011; Waalwijk et al. 2003; Xu et al. 2005), and the strong presence of this species would make the results harder to interpret. The implication of airborne inoculum in the epidemiology of F. culmorum is uncertain and this species has not been reported to produce ascospores (Osborne and Stein 2007). Nevertheless, as conidia can be part of the airborne inoculum (Hörberg 2002; Keller et al. 2014), its quantification could be undertaken.
Our results demonstrated that F. graminearum airborne inoculum is correlated with F. graminearum epidemics in wheat fields. The knowledge of inoculum pressure around wheat anthesis could be fundamental for the development of precise forecasting tools, in order to adapt the control of the disease. From a more practical point of view, changes could be made to the method to allow for realistic use in the field. Firstly, since frequency of detection of F. graminearum inoculum yielded similar, and sometimes even better, levels of correlation than inoculum quantity, the replacement of a qPCR assay by another cheaper DNA-based assay (e.g. LAMP) could greatly reduce the cost, therefore improving the overall benefit of the obtained data. Secondly, the use of the Burkard sampler may not be appropriate for obtaining effective “real-time” information on the field situation, due to the time needed to bring the samples back to the lab and perform the analysis (Jackson and Bayliss 2011). Fortunately, new methods developed in the near future for the capture and detection of inoculum as well as the integration of the results will probably reduce the delay between the collection time in the field and the reporting of the results (West and Kimber 2015). Finally, these new tools, combined with a better knowledge of the implications of airborne F. graminearum and F. culmorum inoculum and meteorological conditions around flowering, could give rise to an efficient model for DON prediction that allows the farmer to take risk-based actions.
References
Almquist, C., & Wallenhammar, A.-C. (2015). Monitoring of plant and airborne inoculum of Sclerotinia sclerotiorum in spring oilseed rape using real-time PCR. Plant Pathology, 64(1), 109–118.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipman, D. J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215(3), 403–410.
Booth, C. (1971). The Genus Fusarium (p. 237). Kew: Commonwealth Mycologiscal Institute.
Chandelier, A., Nimal, C., André, F., Planchon, V., & Oger, R. (2011). Fusarium species and DON contamination associated with head blight in winter wheat over a 7-year period (2003–2009) in Belgium. European Journal of Plant Pathology, 130(3), 403–414.
Chandelier, A., Helson, M., Dvorak, M., & Gischer, F. (2014). Detection and quantification of airborne inoculum of Hymenoscyphus pseudoalbidus using real-time PCR assays. Plant Pathology, 63(6), 1296–1305.
Cowger, C., Patton-Özkurt, J., Brown-Guedira, G., & Perugini, L. (2009). Post-anthesis moisture increased fusarium head blight and deoxynivalenol levels in North Carolina winter wheat. Phytopathology, 99(4), 320–327.
David, R. F., Bozorgmagham, A., Schmale, D. G., Ross, S. D., & Marr, L. C. (2016). Identification of meteorological predictors of fusarium graminearum ascospore release using correlation and causality analyses. European Journal of Plant Pathology, 145(2), 483–492.
De Wolf, E. D., Madden, L. V., & Lipps, P. E. (2003). Risk assessment models for wheat fusarium head blight epidemics based on within-season weather data. Phytopathology, 93(4), 428–435.
Del Ponte, E. M., Fernandes, J. M. C., & Pierobom, C. R. (2005). Factors affecting density of airborne Gibberella zeae inoculum. Fitopatologia Brasileira, 30(1), 55–60.
Del Ponte, E. P., Fernandes, J. M. C., & Bergstrom, G. C. (2007). Influence of growth stage on fusarium head blight and deoxynivalenol production in wheat. Journal of Phytopathology, 155(10), 577–581.
Desjardins, A. E. (2006). Fusarium mycotoxins: chemistry, genetics and biology. St Paul: APS Press.
Dill-Macky, R., & Jones, R. K. (2000). The effect of previous crop residues and tillage on fusarium head blight of wheat. Plant Disease, 84(1), 71–76.
Duvivier, M., Dedeurwaerder, G., Proft, M., Moreau, J.-M., & Legrève, A. (2013). Real-time PCR quantification and spatio-temporal distribution of airborne inoculum of Mycosphaerella graminicola in Belgium. European Journal of Plant Pathology, 137(2), 325–341.
Duvivier, M., Dedeurwaerder, G., Bataille, C., De Proft, M., & Legrève, A. (2016). Real-time PCR quantification and spatio-temporal distribution of airborne inoculum of Puccinia triticina in Belgium. European Journal of Plant Pathology, 145(2), 405–420.
Fernando, W. G., Miller, J. D., Seaman, W. L., Seifert, K., & Paulitz, T. C. (2000). Daily and seasonal dynamics of airborne spores of Fusarium graminearum and other Fusarium species sampled over wheat plots. Canadian Journal of Botany, 78(4), 497–505.
Hellin, P., Dedeurwaerder, G., Duvivier, M., Scauflaire, J., Huybrechts, B., Callebaut, A., et al. (2016). Relationship between Fusarium spp. diversity and mycotoxin contents of mature grains in southern Belgium. Food Additives and Contaminants Part A, 33(7), 1228–1240.
Hofgaard, I. S., Seehusen, T., Aamot, H. U., Riley, H., Razzaghian, J., Le, V. H., et al. (2016). Inoculum potential of Fusarium spp. relates to tillage and straw management in Norwegian fields of spring oats. Frontiers in Microbiology, 7, 556.
Hörberg, H. M. (2002). Patterns of splash dispersed conidia of Fusarium poae and Fusarium culmorum. European Journal of Plant Pathology, 108(1), 73–80.
Inch, S., Fernando, W. G. D., & Gilbert, J. (2005). Seasonal and daily variation in the airborne concentration of Gibberella zeae (Schw.) Petch spores in Manitoba. Canadian Journal of Plant Pathology, 27(3), 357–363.
Jackson, S. L., & Bayliss, K. L. (2011). Spore traps need improvement to fulfil plant biosecurity requirements. Plant Pathology., 60(5), 801–810.
Keller, M. D., Waxman, K. D., Bergstrom, G. C., & Schmale III, D. G. (2010). Local distance of wheat spike infection by released clones of Gibberella zeae disseminated from infested corn residue. Plant Disease, 94(9), 1151–1155.
Keller, M. D., Bergstrom, G. C., & Shields, E. J. (2014). The aerobiology of Fusarium graminearum. Aerobiologia, 30(2), 123–136.
Kriss, A. B., Paul, P. A., & Madden, L. V. (2010). Relationship between yearly fluctuations in fusarium head blight intensity and environmental variables: A window-pane analysis. Phytopathology, 100(8), 784–797.
Kriss, A. B., Paul, P. A., Xu, X., Nicholson, P., Doohan, F. M., Hornok, L., et al. (2012). Quantification of the relationship between the environment and fusarium head blight, Fusarium pathogen density, and mycotoxins in winter wheat in Europe. European Journal of Plant Pathology, 133(4), 975–993.
Landschoot, S., Audenaert, K., Waegeman, W., Pycke, B., Bekaert, B., De Baets, B., et al. (2011). Connection between primary fusarium inoculum on gramineous weeds, crop residues and soil samples and the final population on wheat ears in Flanders, Belgium. Crop Protection, 30(10), 1297–1305.
Leplat, J., Friberg, H., Abid, M., & Steinberg, C. (2012). Survival of Fusarium graminearum, the causal agent of Fusarium head blight. A review. Agronomy for Sustainable Development, 33(1), 97–111.
Luchi, N., Ghelardini, L., Belbahri, L., Quartier, M., & Santini, A. (2013). Rapid detection of Ceratocystis platani inoculum by quantitative real-time PCR assay. Applied and Environemental Microbiology, 79(17), 5394–5404.
de Luna, L., Paulitz, T. C., Bujold, I., & Carisse, O. (2002). Ascospore gradients of Gibberella zeae from overwintered inoculum in wheat fields. Canadian Journal of Plant Pathology, 24(4), 457–464.
Maldonado-Ramirez, S. L., Schmale, D. G., III, Shields, E. J., & Bergstrom, G. C. (2005). The relative abundance of viable spores of Gibberella zeae in the planetary boundary layer suggests the role of long-distance transport in regional epidemics of fusarium head blight. Agricultural and Forest Meteorology, 132(1–2), 20–27.
Manstretta, V., Gourdain, E., & Rossi, V. (2015). Deposition patterns of Fusarium graminearum ascospores and conidia within a wheat canopy. European Journal of Plant Pathology, 143(4), 873–880.
Markell, S. G., & Francl, L. J. (2003). Fusarium head blight inoculum: Species prevalence and Gibberella zeae spore type. Plant Disease, 87(7), 814–820.
Nicholson, P., Simpson, D. R., Weston, G., Rezanoor, H. N., Lees, A. K., Parry, D. W., et al. (1998). Detection and quantification of Fusarium culmorum and Fusarium graminearum in cereals using PCR assays. Physiological and Molecular Plant Pathology, 53(1), 17–37.
Nicolau, M., Fernandes, J. M. C., Nicolau, M., & Fernandes, J. M. C. (2012). A predictive model for daily inoculum levels of Gibberella zeae in Passo Fundo, Brazil. International Journal of Agronomy. https://doi.org/10.1155/2012/795162.
Osborne, L. E., & Stein, J. M. (2007). Epidemiology of fusarium head blight on small-grain cereals. International Journal of Food Microbiology, 119(1–2), 103–108.
Paul, P. A., El-Allaf, S. M., Lipps, P. E., & Madden, L. V. (2004). Rain splash dispersal of Gibberella zeae within wheat canopies in Ohio. Phytopathology, 94(12), 1342–1349.
Paul, P. A., Lipps, P. E., De Wolf, E., Shaner, G., Buechley, G., Adhikari, T., et al. (2007). A distributed lag analysis of the relationship between Gibberella zeae inoculum density on wheat spikes and weather variables. Phytopathology, 97(12), 1608–1624.
Paul, P. A., Lipps, P. E., Hershman, D. E., McMullen, M. P., Draper, M. A., & Madden, L. V. (2008). Efficacy of triazole-based fungicides for fusarium head blight and deoxynivalenol control in wheat: A multivariate meta-analysis. Phytopathology, 98(9), 999–1011.
Pereyra, S. A., Dill-Macky, R., & Sims, A. L. (2004). Survival and inoculum production of Gibberella zeae in wheat residue. Plant Disease, 88(7), 724–730.
Pietravalle, S., Shaw, M. W., Parker, S. R., & van den Bosch, F. (2003). Modeling of relationships between weather and Septoria tritici epidemics on winter wheat: A critical approach. Phytopathology, 93(10), 1329–1339.
Prandini, A., Sigolo, S., Filippi, L., Battilani, P., & Piva, G. (2009). Review of predictive models for fusarium head blight and related mycotoxin contamination in wheat. Food and Chemical Toxicology, 47(5), 927–931.
Prussin, A. J., Ross, S. D., & Schmale, D. G. (2014a). Monitoring the long distance transport of fusarium graminearum from field-scale sources of inoculum. Plant Disease, 98(4), 504–511.
Prussin, A. J., Szanyi, N. A., Welling, P. I., Ross, S. D., & Schmale, D. G. (2014b). Estimating the production and release of ascospores from a field-scale source of fusarium graminearum inoculum. Plant Disease, 98(4), 497–503.
Ramirez, M. L., Chulze, S., & Magan, N. (2006). Temperature and water activity effects on growth and temporal deoxynivalenol production by two Argentinean strains of Fusarium graminearum on irradiated wheat grain. International Journal of Food Microbiology, 106(3), 291–296.
Rogers, S. L., Atkins, S. D., & West, J. S. (2009). Detection and quantification of airborne inoculum of Sclerotinia sclerotiorum using quantitative PCR. Plant Pathology, 58(2), 324–331.
Rossi, V., Languasco, L., Pattori, E., & Giosuè, S. (2002). Dynamics of airborne Fusarium macroconidia in wheat fields naturally affected by head blight. Journal of Plant Pathology, 84(1), 53–64.
Schaafsma, A. W., Tamburic-Ilincic, L., & Hooker, D. C. (2005). Effect of previous crop, tillage, field size, adjacent crop, and sampling direction on airborne propagules of Gibberella zeae/Fusarium graminearum, fusarium head blight severity, and deoxynivalenol accumulation in winter wheat. Canadian Journal of Plant Pathology, 27(2), 217–224.
Schmale, D. G., III, Shah, D. A., & Bergstrom, G. C. (2005). Spatial patterns of viable spore deposition of Gibberella zeae in wheat fields. Phytopathology, 95(5), 472–479.
Simes, R. J. (1986). An improved Bonferroni procedure for multiple tests of significance. Biometrika, 73(3), 751–754.
Stein, J. M., & Osborne, L. E. (2006). A relationship between the number of Gibberella zeae propagules present during heading and deoxynivalenol (DON) content in wheat grain. Phytopathology, 96(Suppl), S110.
Stein, J. M., Osborne, L. E., Bondalapati, K. D., Glover, K. D., & Nelson, C. A. (2009). Fusarium head blight severity and deoxynivalenol concentration in wheat in response to Gibberella zeae inoculum concentration. Phytopathology, 99(6), 759–764.
Sutton, J. C. (1982). Epidemiology of wheat head blight and maize ear rot caused by Fusarium graminearum. Canadian Journal of Plant Pathology, 4(2), 195–209.
Trail, F., Gaffoor, I., & Vogel, S. (2005). Ejection mechanics and trajectory of the ascospores of Gibberella zeae (anamorph Fusarium graminearum). Fungal Genetics and Biology, 42(6), 528–533.
Waalwijk, C., Kastelein, P., de Vries, I., Kerényi, Z., van der Lee, T., Hesselink, T., et al. (2003). Major changes in Fusarium spp. in wheat in the Netherlands. European Journal of Plant Pathology, 109(7), 743–754.
Waalwijk, C., van der Heide, R., de Vries, I., van der Lee, T., Schoen, C., Costrel-de Corainville, G., et al. (2004). Quantitative detection of fusarium species in wheat using TaqMan. European Journal of Plant Pathology, 110(5–6), 481–494.
West, J. S., & Kimber, R. B. E. (2015). Innovations in air sampling to detect plant pathogens. Annals of Applied Biology, 166(1), 4–17.
West, J. S., Atkins, S. D., Emberlin, J., & Fitt, B. D. L. (2008). PCR to predict risk of airborne disease. Trends in Microbiology, 16(8), 380–387.
Wieczorek, T. M., Jørgensen, L. N., Hansen, A. L., Munk, L., & Justesen, A. F. (2014). Early detection of sugar beet pathogen Ramularia beticola in leaf and air samples using qPCR. European Journal of Plant Pathology, 138(4), 775–785.
Xu, X. (2003). Effects of environmental conditions on the development of Fusarium ear blight. European Journal of Plant Pathology, 109(4), 683–689.
Xu, X., Parry, D. W., Nicholson, P., Thomsett, M. A., Simpson, D., Edwards, S. G., et al. (2005). Predominance and association of pathogenic fungi causing fusarium ear blight in wheat in four European countries. European Journal of Plant Pathology, 112(2), 143–154.
Ye, J., Coulouris, G., Zaretskaya, I., Cutcutache, I., Rozen, S., & Madden, T. L. (2012). Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinformatics, 13, 134.
Yoshida, M., & Nakajima, T. (2010). Deoxynivalenol and nivalenol accumulation in wheat infected with Fusarium graminearum during grain development. Phytopathology, 100(8), 763–773.
Zadoks, J. C., Chang, T. T., & Konzak, C. F. (1974). A decimal code for the growth stages of cereals. Weed Research, 14(6), 415–421.
Acknowledgements
This research was supported by the Service Public de Wallonie, Direction générale opérationnelle Agriculture, Ressources naturelles et Environnement. The authors wish to thank Gérald Marchal, Frédéric Mathieu, Marie-Eve Renard, Viviane Van Hese, Hanne Verhaegen and Charlotte Liénard for technical assistance.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest and ensure that accepted principles of ethical and professional conduct have been followed. This research does not involve human participants nor animals.
Rights and permissions
About this article

Cite this article
Hellin, P., Duvivier, M., Dedeurwaerder, G. et al. Evaluation of the temporal distribution of Fusarium graminearum airborne inoculum above the wheat canopy and its relationship with Fusarium head blight and DON concentration. Eur J Plant Pathol 151, 1049–1064 (2018). https://doi.org/10.1007/s10658-018-1442-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10658-018-1442-7