
Abstract
Mercury (Hg) is a global pollutant that affects songbird populations across a variety of ecosystems following conversion to methylmercury (MeHg)—a form of Hg with high potential for bioaccumulation and bioavailability. The amount of bioavailable MeHg in an ecosystem is a function of the amount of total Hg present as well as Hg methylation rates, which vary across the landscape in space and time, and trophic transfer. Using songbirds as an indicator of MeHg bioavailability in terrestrial ecosystems, we evaluated the role of habitat, climate, and trophic level in dictating MeHg exposure risk across a variety of ecosystems. To achieve this objective, 2243 blood Hg samples were collected from 81 passerine and near-passerine species in New York State, USA, spanning 10 different sampling regions from Long Island to western New York. Using a general linear mixed modeling framework that accounted for regional variation in sampling species composition, we found that wetland habitat area within 100 m of capture location, 50-year average of summer maximum temperatures, and trophic position inferred using stable isotope analysis were all correlated with songbird blood Hg concentrations statewide. Moreover, these patterns had a large degree of spatial variability suggesting that the drivers of MeHg bioavailability differed significantly across the state. Mercury deposition, land cover, and climate are all expected to change throughout the northeastern United States in the coming decades. Terrestrial MeHg bioavailability will likely respond to these changes. Focused research and monitoring efforts will be critical to understand how exposure risk responds to global environmental change across the landscape.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mercury (Hg) is a pollutant that is globally distributed, but locally variable in its availability for biomagnification and bioaccumulation (Evers and Clair 2005; Driscoll et al. 2013). After being emitted to the atmosphere from natural (e.g., volcanoes) and anthropogenic sources (e.g., coal-fired power plants, municipal incinerators), Hg can be transported globally and deposited in habitats far from the original sources (VanArsdale et al. 2005; Driscoll et al. 2007). Additionally, Hg can enter habitats from local sources through atmospheric deposition (e.g., Evers et al. 2007) or via soil and/or water contamination from human activities (e.g., artisanal gold mining Telmer and Veiga 2009; Gibb and O’Leary 2014; industrial sites; Reis et al. 2009; Davis et al. 2012; Amos et al. 2013). Micro-organisms convert inorganic Hg to methylmercury (MeHg)—a form of Hg that has high potential for bioaccumulation and biomagnification (Boening 2000; Ullrich et al. 2001; Podar et al. 2015). In terrestrial habitats Hg methylation can occur in upland soils (Demers et al. 2007; Rodenhouse et al. 2019) and particularly wetland soils (St. Louis et al. 1994; Kramar et al. 2005).
Songbirds are recognized as critical indicators of MeHg in terrestrial ecosystems, where MeHg can biomagnify in food webs to concentrations that can adversely affect bird populations (Cristol et al. 2008; Jackson et al. 2015). In vertebrates, and specifically avian communities, numerous neurological, immunological, and physiological effects have been documented as a result of MeHg exposure (Scheuhammer et al. 2007; Hawley et al. 2009; Wada et al. 2009). These effects can influence population demography (Brasso and Cristol 2008; Evers et al. 2008; Jackson et al. 2011; Whitney and Cristol 2017). In particular, invertivores like many warblers, vireos, wrens, and some sparrows and thrushes, have been utilized in studies to provide a representation of MeHg concentrations in a variety of ecosystems (Rimmer et al. 2005; Lane et al. 2011; Edmonds et al. 2012; Townsend et al. 2014). However, songbirds can use a variety of habitats in a survey area resulting in individual variation in diet and trophic position resulting in pairing stable isotope with songbird MeHg studies to gain clarity on the causes of the results (Kidd et al. 1995; Cizdziel et al. 2002; Becker et al. 2002; Tsui et al. 2017). When species are habitat generalists, information on individual foraging ecology helps maximize accuracy and precision of monitoring efforts.
Global environmental changes are predicted over this century (Schindler 2001; Meehl et al. 2007), many of which will likely affect MeHg bioavailability in terrestrial food chains and songbirds. Methylmercury production has been shown to vary with Hg inputs, meteorological and hydrological conditions, redox status and land cover (Miskimmin et al. 1992; Ramlal et al. 1993; Sellers et al. 1996; Taylor et al. 2019; Eagles-Smith et al. 2018). In addition to the direct effects of climate on Hg methylation, climate and habitat change will likely alter the location and timing of hotspots of Hg methylation by influencing the abundance and distribution of wetland soils and the frequency and duration of wetting and drying cycles (Craft et al. 2009; Kirwan et al. 2010; Kirwan and Megonigal 2013; Mitsch and Hernandez 2013; Schile et al. 2014). Influx of Hg into ecosystems will change with Hg emissions (Zhang et al. 2016) and the influence of weather on Hg deposition (Mao et al. 2017a, 2017b; Ye et al. 2019). Finally, climate changes can remobilize local stores of Hg in soils and ice, which further increases MeHg bioavailability in habitats with high Hg methylation rates (Stern et al. 2012). Ultimately, as global environmental change alters spatial and temporal patterns of Hg deposition and methylation on the landscape, the future of MeHg bioavailability becomes increasingly uncertain.
In the face of anticipated changes in Hg emissions, deposition, and methylation rates, research on MeHg bioaccumulation in New York State, USA has used passerine and near-passerine communities to explain current patterns of terrestrial Hg biomagnification and bioaccumulation. A single indicator species is rarely present in terrestrial habitats statewide. As a result, sampling multiple species from a broader community is necessary to allow for more cost-effective and geographically comprehensive assessments. This research builds on 14 previous years of Hg research in New York State as well as other long-term Hg monitoring studies on songbirds across the northeastern United States (e.g., Sauer et al. 2019; Lane et al. 2011, 2019) to inform future MeHg monitoring efforts.
Songbird blood Hg concentrations collected in New York State from 2013–2017 were used to better understand the environmental variables that affect MeHg bioavailability in terrestrial ecosystems. As both abiotic and biotic conditions are known affect Hg methylation rates, we hypothesize that both climate and habitat at the sampling location combine to influence Hg concentrations in songbird blood. Moreover, we hypothesize that individual foraging patterns and diet influence blood Hg concentrations in songbirds. To describe patterns of Hg concentrations in songbird blood in New York and address these hypotheses we determined: (1) the species with the highest blood Hg concentrations in each sampling region; (2) the effects of habitat and climate on songbird blood Hg concentrations across all the sampling sites; and (3) the importance of individual diet on Hg blood concentrations using stable carbon and nitrogen isotope analysis. Further, we assess regional variation in the importance of climate, habitat, and individual diet to songbird blood Hg.
Methods
Sampling design and study areas
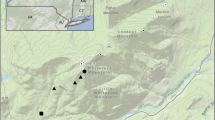
Songbird Hg sampling took place throughout New York State. Sites were selected in a based on the following criteria: (1) previous Hg sampling efforts for songbirds or other biota, (2) habitat sensitivity to MeHg bioaccumulation, and (3) proximity to Hg emission sources. Four ‘core’ sampling sites were selected in each of three regions known to have consistent elevated Hg concentrations capable of causing adverse effects in biota: the Adirondack Mountains, Catskill Mountains, and Long Island (Driscoll et al. 2007; Evers et al. 2007; Fig. 1). Core sites were visited each year to robustly sample each community and estimate interannual variation in Hg exposure (Table 1). Study sites within the Adirondack Park included boreal Sphagnum bog and wetland habitats and a mix of deciduous and coniferous upland forests types. Sites within the Catskills were representative of large wetland complexes, upland deciduous forest, and high-elevation mixed pine-oak forest. Long Island sites were primarily composed of tidal marsh and riparian forest.

Sampling locations for songbird blood Hg monitoring in New York, 2013–2017. Core regions are sites that were sampled in all five years of the study
In addition to these 12 core sites an additional 40 sampling sites were visited once during the five-year study. These sites were selected to increase the spatial scope and habitat diversity of the sampling effort. Previous information on songbird Hg exposure was not a prerequisite for site selection, but the remaining two selection criteria from the core site study design were used. Consequently, five additional regions were identified for statewide sampling: Western New York, Northern New York, Tug Hill Plateau, New York City, the Capital Region and the Finger Lakes (Fig. 1). Sites within the Western New York region were comprised of mixed forest, forested wetlands, and emergent freshwater wetlands. Study sites in Northern New York and the Capital Region included a mix of alvar grasslands, large wetland complexes, and spruce bog/cedar swamp habitats. Tug Hill Plateau included sites representing deciduous upland forests and marsh-beaver meadow. Study sites within the greater New York City region included estuarine emergent marsh and coastal scrub. Finger Lakes sites were represented by several marsh and large wetland complexes interspersed with forest. Within each region, four to six sites were sampled. Generally, all sites in a given region were sampled in the same year, though there were a few exceptions for logistical reasons.
Bird capture and tissue sampling
All bird capture and tissue sampling was conducted from 2013–2017 during periods of peak breeding activity in June and July. Birds were captured by opening 6 m- and 12 m-long, 30–36 mm mesh mist nets for a minimum of 2 h. During this period, capture effort was augmented with conspecific vocalization playback for up to 30 min to attract birds to each net. Vocalizations from species known to be sensitive to Hg bioaccumulation were used in the audio playback to increase capture rate and bias the sample toward these species (Jackson et al. 2015).
Once captured, each bird was banded with a uniquely-numbered USGS aluminum band and sampled. Age, sex, and reproductive status were assessed using plumage and presence of cloacal protuberances/brood patches, and morphometric measures like wing chord length were recorded. Blood samples were collected via venipuncture of the cutaneous ulnar vein with a 27-gauge sterile disposable needle. Fifty to 75 μl of whole blood was collected into heparinized, Mylar-wrapped capillary tubes for Hg and stable isotope analysis. Not all samples were large enough for both Hg and stable isotope analysis, so Hg determination was prioritized. The capillary tubes were sealed with Critocaps® and stored in plastic vacutainers on ice for up to 6 h before freezing at −17° Celsius. All birds were released unharmed within 10–25 min of capture.
Laboratory analysis
Blood samples were analyzed for total Hg at BRI’s Wildlife Mercury Research Laboratory in Portland, Maine. Mercury concentration was determined via thermal decomposition coupled with atomic absorption spectroscopy using a Milestone DMA 80, following Environmental Protection Agency SW-846 Method 7473. Prior to analysis, the equipment was calibrated using NIST-certified standard solutions, and accuracy and precision were evaluated within each analytical batch through continued calibration verifications and the inclusion of certified reference materials, duplicates, blanks, and matrix spikes (approximately ten out of every 40 measurements). Quality control methods, including the use of one of the DOLT-certified reference materials (DOLT 4, DOLT 5) and BCR 463 or CE 464 were used to ensure consistent analytical precision and accuracy. Calibration utilized a blank and two standards, one for each of the two detector cells. Percent recovery of certified reference materials was >90% and relative percent difference (RPD) of duplicates were within 10%. The instrument detection limit was 0.001 µg/g, and all blood Hg concentrations were reported in µg/g wet weight (ww). Methylmercury was not analyzed because approximately 95% of total Hg in songbird blood is in the form of MeHg (Rimmer et al. 2005, Edmonds et al. 2010) and we assume this relationship is consistent across all songbird species.
A total of 1018 songbird blood samples were analyzed at the Boston University Stable Isotope Laboratory in Boston, Massachusetts for stable carbon and nitrogen isotope ratios. Bird blood was analyzed using automated continuous-flow isotope ratio mass spectrometry (Michener and Lajtha 2007). Blood was transferred from capillary tubes into pre-weighed tin capsules. Assuming 70% water content, approximately 1.3 mg of blood was added to the capsules. All capsules were oven dried at 60 °C for 24 h and then reweighed for dry mass. The capsules were then folded and compressed prior to analysis. The samples were combusted in a EuroVector Euro EA elemental analyzer. The combustion gases (N2 and CO2) were separated on a gas chromatography (GC) column, passed through a reference gas box and introduced into a GV Instruments IsoPrime isotope ratio mass spectrometer; water was removed using a magnesium perchlorate water trap.
Climate, habitat, and trophic position data integration
Independent variables were gathered from publicly available data sources to explain songbird blood Hg concentrations. To describe patterns of biomagnification, standardized diet composition data for each species was collected from the Wilman et al. (2014) database. The percentage of year-round diet comprised of invertebrates was extracted for each species, which serves as a relative measure of trophic position for most invertivores and can correlate with MeHg exposure risk (Cristol et al. 2008; Jackson et al. 2015). As stable isotope data was only available for a subset of the samples, the Wilman et al. (2014) data were used to infer species-level dietary differences for all sampled individuals.
Land cover data were gathered from the National Land Cover Database 2011 (Homer et al. 2015). The three categories of forest habitat (Deciduous Forest, Evergreen Forest, and Mixed Forest) were combined into a single category of forested habitats. Similarly, the two wetland habitat categories (Woody Wetlands and Emergent Herbaceous Wetlands) were combined into a single category of wetland habitats. The area of these aggregated categories was summed within a 100 m radius circle around the capture location of each sample (i.e., the capture net) to approximate the foraging habitat of captured birds.
Climate data were gathered from downscaled BCSD-CMIP5 climate projections (https://gdo-dcp.ucllnl.org/downscaled_cmip_projections). Monthly climate analysis results, used to train climate projection models from a common 20C3M simulation, were acquired at the 1/8-degree grid size scale from the Hydrology projection set. These data were used to describe the 50-year averages of maximum temperature and precipitation at the monthly time scale from 1950–1999. These data are not themselves assuming a climate projection scenario, but rather are modeled climate baseline data used to describe the climate of New York. The capture locations of all birds in our database were associated with the climate averages from a 1/8-degree grid cell. Some capture locations were just outside of the closest grid cell (e.g., tidal marsh sites on Long Island) so the climate averages from the nearest available cell were used as a reasonable approximation. Climate variables were averaged across a three-month seasonal window that matches the songbird breeding season for most species (June-August).
Statistical analysis
The mean, standard deviation, and sample size of Hg samples were used to describe overall patterns of Hg exposure for each region. To determine how differing factors influenced Hg bioavailability across sites, however, it was necessary to control for random variation in species and site sampling frequency. Blood Hg concentrations varied considerably across species, primarily due to differences in foraging habitat and diet, and the sampled songbird community varied significantly among sites, which was necessary to achieve unbiased estimates of site-level MeHg bioavailability.
To make comparisons among large numbers of species, this analysis controls for species-level and site-level variance and explains relative differences in species across habitat, climate, and trophic level. To achieve this goal, we parameterized three different general linear mixed models to answer three different questions; all models had a similar overall structure. The response variable used for the models was loge-transformed blood Hg concentrations. Goodness-of-fit was evaluated using R2 (both marginal and conditional), quantile-quantile plots, and fitted versus residual plots. Samples without enough Hg to reach the detection limit were given the value of the analytical detection limit (0.001 Hg ppm ww) to avoid zeroes in the untransformed response variable. All independent covariates were tested for multicollinearity before inclusion. Continuous covariates were scaled by subtracting the mean then dividing by the standard deviation to improve maximum likelihood optimization. Not all individuals sampled were included in the analysis; only breeding birds (i.e., adults) and passerines or near-passerines (i.e., including woodpeckers but excluding incidental captures of raptors, shorebirds, and rails) were included in analysis (n = 2243). Recaptures within years were not included in the analysis, though the small number of recaptures among years were included.
The first model was designed to assess regional variation in species-level blood Hg concentrations and identify species with significant Hg bioaccumulation potential in each region. We used a general linear model with multiple nested random effects; this model parameterization allowed the average Hg blood concentration to be estimated independently for each species/region combination. The species variable was nested within region, and year (a categorical variable) was nested within site to account for spatial and temporal variation in Hg bioavailability. Region was included as an additional standalone random effect. Sex was included in the model as the only fixed effect. Parametric bootstrapping (n = 250) was used to estimate 95% confidence intervals around parameter estimates and predictions from this model (and the following models).
The second model was designed to assess the importance of climate and habitat in influencing terrestrial MeHg bioavailability (as estimated via songbird blood Hg). Nested random effects, fixed effects, and fixed effects with random components were all used to parameterize this model. Random effects were both nested and standalone; species nested within year and region was used to estimate annual means for each species in each region sampled. Site was also included as a random effect with no nesting or interactions to account for spatial variation in sampling within each region. Fixed effects included individual and environmental covariates that were thought to influence blood Hg levels: sex (male/female/unknown), the amount of forest/wetland habitat area around the capture area, the 50-year averages of maximum summer temperatures and total rainfall for the capture area, and the species-level percentage of invertebrates in the diet. For the wetland, temperature, rainfall, and diet covariates we added a random effect that allowed for regional variation in the main fixed effect, as these variables had the potential to have significant spatial variation in their effects.
The third model included the subset of Hg samples for which stable isotope analysis was conducted (n = 1018) and was constructed in a similar manner to the second model. Stable carbon (δ13C) and nitrogen (δ15N) isotopes in blood were used to refine our understanding of the habitat origins of food source (as estimated by δ13C) and trophic level (as estimated by δ15N) in determining blood Hg at the individual scale. Using a similar general linear model structure to that described above, we parameterized a single model that included nested random effects, fixed effects, and fixed and random mixtures. Regional variation the relationship between δ13C and δ15N was allowed to account for differences in isotope composition among biomes (Hobson 1999). Sex was included as a fixed effect, and stable carbon and nitrogen isotopes were included as fixed effects with random components nested within region. As above, parameters that combined fixed and random effects estimated an overall effect of these covariates across all regions and then allowed for variation in Hg availability between regions.
All analyses were conducted using the R statistical software platform (R Core Team 2018). General linear mixed modeling was conducted using package ‘lme4’ (Bates et al. 2015). Data manipulation and figure creation used ‘dplyr’ (Wickham et al. 2018) and ‘ggplot2’ (Wickham 2009), respectively.
Results
Mercury concentrations varied widely among sites and species. The blood Hg concentrations ranged from a low of <0.001 ppm ww (i.e., below the instrument detection limit) in an American Goldfinch at Neversink Preserve (Catskills) to a high of 4.1 ppm ww in a Swamp Sparrow from Northern Montezuma Wildlife Management Area (Finger Lakes). Songbirds on Long Island and New York City had the highest average blood Hg concentrations, followed by the Finger Lakes, Adirondacks and Catskills regions (Table 1). Among the core regions, Long Island showed the highest overall interannual variation in Hg concentrations (Table 1).
Overall model fit
All three general linear mixed models showed strong overall goodness-of-fit (R2 ranged from 0.80 to 0.87) and there appeared to be no signs of heteroscedasticity or other examples of poor fit based on the use of a normal distribution to describe the dependent variable (log-transformed blood Hg). The complex random effects structure used in this effort consistently explained more of the response variance than the fixed effects in all the models. Overall, this shows the importance of controlling for the species and location (site and region) random effects, particularly when using these data to make inference about broader patterns of MeHg availability.
Regional variation in species Hg concentrations
The large number of sites and multi-species sampling approach of this project provided an opportunity to evaluate the blood Hg concentration of many species. A summary of all species/region combinations can be found in supplemental materials (Appendix). Sixty-two percent of the total variation in the blood Hg data was explained by the species nested within region parameters. Changes in songbird Hg within sites and across years represented about 18% of the total variation. The five species with the highest Hg concentrations in each region included a wide range of invertivorous passerines (Fig. 2). Blood Hg concentrations in species varied significantly by region, thus the highest species in some regions were not significantly higher than the overall study average (e.g., the Capital Region) while other regions had many species that were significantly higher than the overall average (e.g., Long Island, the Adirondacks, and the Catskills). Seaside and Saltmarsh Sparrows had much higher average blood Hg concentrations than the other species but were only found in two regions. While Swamp Sparrows were identified as having elevated Hg concentrations relative to the other species sampled in seven different regions. Most species were only observed with elevated blood Hg concentrations in a single region. Sex was not an important predictor of blood Hg concentrations across this sampled community.

Model estimated mean and variance of blood Hg concentrations (ppm ww) for the five highest species in each region. Estimates were obtained from a general linear mixed model that nested species within region and were calculated for males of each species. Error bars represent bootstrapped 95% confidence intervals of the mean. The vertical black line represents the study-wide average blood Hg concentration (0.14 ppm ww)
The effects of climate and habitat on Hg bioavailability
In the generalized linear mixed model with habitat and climate variables, random effects explained approximately 80% of the total variation in songbird blood Hg. Fixed effects explained 8%. Of the random effects, the nested parameters of year, region and species explained 24% of the total variation, and the regional variation in 50-year average summer maximum temperatures explained 44% of the total variation. Site explained 8% of the variation and all other random components explained <5% of the total variation. Of the fixed effects, the amount of wetland habitat in the sampling area was the only parameter that showed a statistically significant effect (β = 0.17, 95% Confidence Interval: 0.03–0.32; Fig. 3). The 50-year average summer rainfall was marginally statistically significant (β = 0.23, 95% CI: −0.04–0.49; Fig. 3). Species-level diet and forest habitat were not important to explaining patterns of Hg concentrations.

Parameter estimates for fixed effects from two separate general linear mixed models that explained patterns in songbird blood Hg concentrations (ppm ww). All covariates were scaled before analysis so covariate magnitude is an accurate reflection of importance. Stable carbon and nitrogen estimates represent a subset of the total database and come from a different model than the rest of the estimates. Points represent maximum likelihood estimates of beta parameters and error bars are bootstrapped 95% confidence intervals
There was significant regional variance in the effects of wetlands and climate on songbird blood Hg concentrations. Overall, songbird blood Hg concentrations increased with increasing amounts of wetland habitat within 100 m of the sampling site; the positive effect of wetlands on blood Hg was particularly important in the New York City and Catskills regions (Fig. 4a). Summer maximum temperature had a more variable effect across regions. Western New York, New York City, and the Finger Lakes regions showed a strong positive relationship between temperature and blood Hg, while Northern New York had a negative relationship (Fig. 4b). Here, even though the effect was not statistically significant for all regions, average temperatures were important to blood Hg concentrations in many regions and was one of the most important variables for explaining songbird blood Hg concentrations overall. Summer rainfall had a borderline important positive effect on blood Hg overall, but there was minimal regional variation in the effect.

Regional variation in the effect of wetland habitat area (a), summer maximum temperatures (b), and trophic position (c) on songbird blood Hg (ppm ww). The x-axis represents the slope of the relationship between the covariate and the response variable from a general linear mixed effects model. Regional means are a combination of the overall fixed effect beta estimate and random regional variation in the effect. The error bar represents two times the standard deviation of the combined estimate and the dotted blue line is at zero. If the error bar overlaps zero, then it is likely that the effect is not strong in that region. The standard deviation is estimated by combining the variance of the fixed and random effects
The effect of trophic position on Hg exposure risk
Using the stable isotope general linear mixed model, we were able to quantify the effects of individual-level δ13C and δ15N on songbird blood Hg levels. In this model, the random effects represented 72% of the total variance in songbird blood Hg and the fixed effects 8%. The most important random effect was region, which explained the vast majority of the variance in the data. Species nested within region and year parameters as well as the site parameters both explained much smaller portions of the total variance. In terms of fixed effects, there was a marginal negative effect of δ13C on blood Hg levels that was consistent among regions (β = −0.04, 95% CI: −0.11–0.04; Fig. 3). Depleted (i.e., more negative) δ13C values would indicate that the animal’s food was coming from more mesic habitats, but we find no evidence of this effect. We lacked the data to understand site to site variation in stable carbon isotope signatures, and this likely reduced our power to assess this effect on Hg bioavailability. The effect of δ15N was strongly positive overall (β = 0.11, 95% CI: 0.04–0.2, Fig. 3), and also variable by region (Fig. 4c). As enriched δ15N values indicate an individual foraging from a higher trophic position, higher trophic level was strongly positively correlated with blood Hg. The effect was strongest in the Adirondacks, Catskills, New York City, Northern New York, and Long Island, and was unimportant only in Western NY, Tug Hill, and the Finger Lakes. The effect of sex on blood Hg concentration was negligible in this model, as it was in the previous analyses.
Discussion
From 2013–2017, blood Hg concentrations across the New York songbird community were affected by habitat, climate, and trophic level. The highest blood Hg concentrations were found in areas of historical Hg monitoring activity: Long Island, the Adirondack Mountains, and the Catskill Mountains. Saltmarsh Sparrows and Seaside Sparrows in the Long Island tidal marshes had elevated blood Hg concentrations that were similar to those found in past studies in the northeastern United States (Warner et al. 2010; Lane et al. 2011, 2019). At inland sites, wetland-dependent species like Palm Warblers also had elevated blood Hg levels that were similar to past studies (Sauer et al. 2019). We found evidence that wetland habitat, climate, and food webs combined to influence MeHg bioavailability in New York songbirds. The amount of nearby wetland habitat and the average maximum summer temperatures explained significant spatial variation in songbird blood Hg across New York. Summer precipitation had a marginally significant effect across all sites, while summer maximum temperature was highly variable in importance and effects across regions.
Variation in Hg bioaccumulation across songbirds
Species with elevated blood Hg concentrations are often useful for detecting spatiotemporal patterns in Hg distributions, as they show consistent accumulation of MeHg at sites and are efficient monitors of changes in MeHg availability (Furness 1993). Some of these species are distributed widely across regions; Swamp Sparrows, for example, were associated with freshwater wetland habitats with elevated Hg methylation rates and were found in multiple regions (e.g., New York City, Northern New York, Western New York, Tug Hill, and the Adirondack Mountains). Other species have narrow habitat requirements; Saltmarsh Sparrows and Seaside Sparrows are tidal marsh obligates (Correll et al. 2017) and while they were only found in two regions (New York City and Long Island) they had some of the highest average blood Hg concentrations found in this study.
Regional differences in Hg exposure results from a combination of many factors. Changes in atmospheric deposition of Hg over New York State (Mao et al. 2017a) and habitat- and soil-specific methylation rates (Ullrich et al. 2001; Podar et al. 2015; Rodenhouse et al. 2019) likely explain much of the observed variation. However, aside from species-level diet composition, we do not explore species-level traits as explanations for interspecies differences in Hg exposure. Annual schedules of nesting, migration, and molt can influence breeding ground blood Hg concentrations (Rimmer et al. 2005, Jackson et al. 2015, Seewagen 2018), and these trends are not accounted for in this study. Female birds eliminate Hg body burden during egg production (Heinz and Hoffman 2009), but we see no evidence of lower female body burden across all species in this study. This could be due to a non-random sampling of females overall (they were only 29% of the total sample), untested species-level variation, or sampling across multiple breeding stages.
Many of the species identified in this study are used as indicators in previous songbird Hg monitoring efforts. Previous studies in the northeastern United States suggest that Saltmarsh Sparrow, Common Yellowthroat, Swamp Sparrow, and Red-eyed Vireo have elevated tissue Hg concentrations (Jackson et al. 2015, Sauer et al. this issue), and several species have been used to assess environmental Hg bioavailability in other studies, including Tree Swallows (Longcore et al. 2007), Catharus thrushes (Townsend et al. 2014) and Northern Waterthrush (Adams et al. 2019). Some of these species, particularly tidal marsh endemics, are experiencing population declines and the role of MeHg in such trends is unclear (Lane et al. 2011, Correll et al. 2017). Monitoring efforts that include means to assess long-term effects of MeHg exposure are needed for species of conservation concern.
The effects of habitat and climate on MeHg exposure
Past research has suggested mechanisms for both habitat and climate to influence Hg deposition and methylation rates. Dry Hg deposition is elevated in forested landscapes while wet Hg deposition is dependent on rainfall (Mao et al. 2017a, 2017b; Risch and Kenski 2018; Ye et al. 2019). Rates of Hg methylation are highest in wetland soils and are increased by higher temperatures and flooding frequencies (Ramlal et al. 1993; St. Louis et al. 2004; Windham-Myers et al. 2014). Causal connections between climate, habitat and songbird MeHg bioaccumulation have yet to be made, though the observations in this study emphasize the value of future work in this area. The role of wetland habitat area in Hg methylation rates seems clear (more wetlands increase MeHg production) but maximum summer temperatures could be associated with both Hg deposition or methylation rates (Meehl et al. 2007; Schindler 2001) and the mechanism of effect is unknown.
Regional variation in the relationship among wetland area, maximum summer temperatures, and songbird blood Hg suggest complex interactions dependent on habitat subtypes and spatial variation in Hg deposition. The regional effect of wetland area ranged from neutral to positive on songbird blood Hg and the mechanism of this effect is unknown. While data on this issue are limited, this result does not appear to be related to wetland subtype. Wetland area is most important to predicting songbird blood Hg concentrations in New York City (where the wetlands are almost entirely tidal marsh) and the Catskills (freshwater wetlands, lakes, and streams) but not extremely important in other similar regions. Given this observation, variation in biomagnification factor or species trophic niches across regions could be a major cause of this pattern. As this study observes the relative effect of habitat after accounting for species-level variation, this result could be dependent upon inconsistency in diet or habitat use within species. The effect of maximum summer temperatures on songbird blood Hg showed the most variation across regions. Northern New York shows a negative correlation between temperature and blood Hg, unlike the other regions where neutral to positive relationships were observed. These patterns are difficult to interpret due to the uncertainty of mechanism between temperature and terrestrial songbird Hg bioaccumulation. Spatial changes in this relationship could be due to variance in atmospheric Hg emissions or prevalence of appropriate Hg methylation conditions (Ullrich et al. 2001; Risch and Kenski 2018). A notable confounding variable could be elevation, which is correlated with both temperature and Hg deposition and would only influence regions with topographic variation (Yu et al. 2013).
Forecasted changes to wetland habitats and climate in North America in the coming decades have the potential to influence songbird MeHg bioavailability. Wetland habitat has declined globally (Zedler and Kercher 2005; Kirwan and Megonigal 2013) but appears stable in the northeastern United States partly due to human-created wetlands (Dahl 2011). Climate change is expected to increase both temperature and precipitation in the northeastern United States (Hayhoe et al. 2007, 2008). Moreover, changes to climate in New York also influences the amount of statewide wetland habitat, in both estuarine (Warren and Niering 1993) and palustrine systems (Hayhoe et al. 2007; Brooks 2009). The present study is based on continental-scale land cover data and 50-year climate averages; future work should focus on understanding the response of MeHg bioavailability to fine-scale changes in climate that will help build model-based forecasts of MeHg bioavailability.
The role of individual foraging niche on MeHg exposure
Individual trophic level is related to tissue Hg concentrations in biota across many ecosystems (Kidd et al. 1995; Cizdziel et al. 2002; Becker et al. 2002; Rodenhouse et al. 2019). While species-level traits can be useful for understanding variation in tissue Hg concentrations (Jackson et al. 2015), they do not explain much of the data observed in this study after species is accounted for. Moreover, regional variation in the relationship between δ15N and blood Hg concentrations—a useful estimate of regional biomagnification factor—is significant in this study. Regional variation in food chain length is a potential explanation for this relationship (Cabana et al. 1994) and would further explain why species-level trophic level estimates are not predictive of the patterns seen in this study.
Given the lack of overall importance of δ13C and significant regional differences in the relationship between δ13C and songbird blood Hg concentrations, we find these data were not useful for explaining statewide patterns of MeHg bioavailability. While δ13C is associated with mesic habitats with Hg methylation potential in past studies (Marra et al. 1998), it could also be associated with marine to freshwater transitions and other changes in C3 and C4 plant abundance (Kelly 2000). This lack of specificity limits the usefulness of these data to the present study, particularly due to the diversity of habitats sampled. Mercury isotopes are useful for identifying local sources of Hg in songbirds (Tsui et al. 2017), and these techniques show promise for understanding individual-level variation in songbird Hg exposure risk. Further work is needed to identify additional tools that accurately describe risk of increased Hg bioavailability across a range of habitats.
Conclusions
Using community sampling techniques, this study was able to estimate blood Hg concentrations across a large number of songbird species and determine how habitat and climate combine to influence relative changes in species Hg exposure across New York State. Wetland habitat and summer maximum temperatures influenced patterns in songbird blood Hg variably across regions. Blood Hg concentrations were also correlated with relative trophic position of individuals. The importance of habitat, climate, and trophic position varied by sampling region, which suggests that unmeasured differences in ecosystems are interacting with these variables to create multiple responses. While single-species study designs could lead to clear results when habitat is similar across study sites (e.g., Evers et al. 2007), this study showed that inference could be made across diverse species and habitats when sample size was sufficiently large and information was shared across species and regions. More research into the impact of sampling scheme on MeHg bioavailability is needed to assess current methods and develop new ones. A clear next step would be to add stable nitrogen sampling to all birds, as well as soils and invertebrates to better describe trophic relationships across regions.
While we have achieved an improved understanding of the scope and origin of songbird MeHg exposure in New York, there are many questions that remain. With some wetland habitats table in the northeast (Dahl 2011) and temperatures increasing (Hayhoe et al. 2007, 2008), future increases in terrestrial MeHg bioavailability appear likely. However, habitat and climate changes can create no-analog communities (Williams and Jackson 2007) that will make forecasting changes to MeHg bioavailability challenging. Recent industrial regulations in the United States appeared to reduce Hg emissions and depositions in the northeast (Driscoll et al. 2015) and reduction in emissions can lead to reductions in bioavailable MeHg (Lee et al. 2016), but these rules are currently in legal flux and their future is unclear. Thus, subsequent monitoring efforts must have the capacity to address multiple objectives: status and trends assessments will need to paired with connections to management actions and meaningful conservation decisions to maximize the knowledge that we gain and the impact of our science (Lyons et al. 2008). In this case, we must design projects that accurately estimate site- and species-level Hg exposure and trends while expanding our knowledge of the effect of habitat and climate and Hg bioaccumulation in terrestrial food webs. Studies that experimentally test mechanisms for climate and habitat interactions on MeHg bioavailability, while gathering detailed and site-specific data on habitat and climate in a variety of terrestrial ecosystems, will be critical to identifying species at risk to future adverse effects of Hg.
References
Adams EM, Williams KA, Olsen BJ, Evers DC (2019) Mercury exposure across the annual cycle in migratory songbirds: implications for migratory behavior. Ecotoxicology (In press)
Amos HM, Jacob DJ, Streets DG, Sunderland EM (2013) Legacy impacts of all-time anthropogenic emissions on the global mercury cycle. Glob Biogeochem Cycles 27(2):410–421
Bates D, Maechler B, Bolker, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48
Becker PH, Gonzalez-Solis J, Behrends B, Croxall J (2002) Feather mercury levels in seabirds at South Georgia: influence of trophic position, sex, and age. Mar Ecol Prog Ser 243:261–269
Boening DW (2000) Ecological effects, transport, and fate of mercury: a general review. Chemosphere 40:1335–1351
Brasso RL, Cristol DA (2008) Effects of mercury exposure on the reproductive success of Tree Swallows (Tachycineta bicolor). Ecotoxicology 17:133–141
Brooks RT (2009) Potential impacts of global climate change on the hydrology and ecology of ephemeral freshwater systems of the forest of the northeastern United States. Clim Chang 95:469–483
Cabana G, Tremblay A, Kalff J, Rasmussen JB (1994) Pelagic food chain structure in Ontario Lakes: a determinant of mercury levels in Lake Trout (Salvelinus namaycush). Can J Fish Aquat Sci 51:381–389
Correll MD, Wiest WA, Hodgman TP, Shriver WG, Elphick CS, McGill BJ, O’Brien K, Olsen BJ (2017) Predictors of specialist avifaunal decline in coastal marshes. Conserv Biol 31:172–182
Craft C, Clough J, Ehman J, Joye S, Park R, Pennings S, Guo H, Machmuller M (2009) Forecasting the effects of accelerated sea-level rise on tidal marsh ecosystem services. Front Ecol Environ 7(2):73–78
Cristol DA, Brasso RL, Condon AM, Fovargue RE, Hallinger KK, Monroe AP, White AE (2008) The movement of aquatic mercury through terrestrial food webs. Science 320(5874):335
Cizdziel JV, Hinners TA, Pollard JE, Heithmar EM, Cross CL (2002) Mercury concentrations in fish from Lake Mead, USA, related to fish size, condition, trophic level, location, and consumption risk. Arch Environ Contamination Toxicol 43:309–317
Dahl TE (2011) Status and trends of wetland in the conterminous United States 2004–2009. U.S. Department of the Interior, Fish and Wildlife Service, Washington, D.C., p 108
Davis JA, Looker RE, Yee D, Marvin-Di Pasquale M, Grenier JL, Austin CM, McKee LJ, Greenfield BK, Brodberg R, Blum JD (2012) Reducing methylmercury accumulation in the food webs of San Francisco Bay and its local watersheds. Environ Res 119:3–26
Demers JD, Driscoll CT, Fahey TJ, Yavitt JB (2007) Mercury cycling in litter and soil in different forest types in the Adirondack region, New York, USA. Ecol Appl 17(5):1341–1351
Driscoll CT, Han Y-J, Chen CY, Evers DC, Lambert KF, Holsen TM, Kamman NC, Munson RK (2007) Mercury contamination in forest and freshwater ecosystems in the Northeastern United States. BioScience 57:17–28
Driscoll CT, Mason RP, Chan HM, Jacob DJ, Pirrone N (2013) Mercury as a global pollutant: sources, pathways, and effects. Environ Sci Technol 47(10):4967–4983
Driscoll CT, Buonocore JJ, Levy JI, Lambert KF, Burtraw D, Reid SB, Fakhraei H, Schwartz J (2015) US power plant carbon standards and clean air and health co-benefits. Nat Clim Change 6:535–540
Eagles-Smith CA, Silbergeld EK, Basu N, Bustamante P, Diaz-Barriga F, Hopkins WA, Nyland JF (2018) Modulators of mercury risk to wildlife and humans in the context of rapid global change. Ambio 47(2):170–197
Edmonds ST, Evers DC, Cristol DA, Mettke-Hofmann C, Powell LL, McGann AJ, Armiger JW, Lane OP, Tessler DF, Newell P, Heyden K, O’Driscoll NJ (2010) Geographic and seasonal variation in mercury exposure of the declining Rusty Blackbird. Condor 112(4):789–799
Edmonds ST, O’Driscoll NJ, Hillier NK, Atwood JL, Evers DC (2012) Factors regulating the bioavailability of methylmercury to breeding rusty blackbirds in northeastern wetlands. Environ Pollut 171:148–154. https://doi.org/10.1016/j.envpol.2012.07.044
Evers DC, Clair TA (2005) Mercury in northeastern North America: a synthesis of existing databases. Ecotoxicology 14:7–14
Evers DC, Han Y-J, Driscoll CT, Kamman NC, Goodale WM, Lambert KF, Holsen TM, Chen CY, Clair TA, Butler TJ (2007) Biological mercury hotspots in the Northeastern United States and Southeastern Canada. BioScience 57:29–43
Evers DC, Savoy L, DeSorbo C, Yates DE, Hanson W, Taylor KM, Siegel L, Cooley JH, Bank MS, Major A, Munney K, Mower B, Vogel HS, Schoch N, Pokras M, Goodale MW, Fair J (2008) Adverse effects from environmental mercury loads on breeding common loons. Ecotoxicology 17(2):69–81
Furness RW (1993) Birds as monitors of pollutants. In: Furness RW, Greenwood JJD (eds) Birds as monitors of environmental change. Springer, Dordrect
Gibb H, O’Leary KG (2014) Mercury exposure and the health impacts among individuals in the artisan and small-scale gold mining community: a comprehensive review. Environ Health Perspect 122(7):667
Hawley DM, Hallinger KA, Cristol DA (2009) Compromised immune competence in free-living tree swallows exposed to mercury. Ecotoxicology 18(5):499–503
Hayhoe K, Wake CP, Huntington TG, Luo L, Schwartz MD, Sheffield J, Wood E, Anderson B, Bradbury J, DeGaetano A, Troy TJ, Wolfe D (2007) Past and future changes in climate and hydrological indicators in the US Northeast. Clim Dyn 28(4):381–407
Hayhoe K, Wake B, Anderson X-Z, Liang E, Maurer J, Zhu J, Bradbury A, DeGaetano A, Hertel, Wuebbles D (2008) Regional climate change projections for the northeast U.S. Mitig Adapt Strateg Glob Change 13(5-6):425–436
Heinz GH, Hoffman DJ (2009) Mercury accumulation and loss in mallard eggs. Environ Toxicol Chem 23(1):222–224
Hobson KA (1999) Stable-carbon and nitrogen isotope ratios of songbird feathers grown in two terrestrial biomes: implications for valuating trophic relationship and breeding origins. Condor 101:799–805
Homer CG, Dewitz JA, Yang L, Jin S, Danielson P, Xian G, Coulston J, Herold ND, Wickham JD, Megown K (2015) Completion of the 2011 National Land Cover Database from the conterminous United States—representing a decade of land cover change information. Photogramm Eng Remote Sens 81(5):345–354
Jackson AK, Evers DC, Etterson MA, Condon AM, Folsom SB, Detweiler J, Schmerfeld J, Cristol DA (2011) Mercury exposure affects the reproductive success of a free-living terrestrial songbird, the Carolina wren (Thryothorus ludovicianus). Auk 128(4):759–769
Jackson A, Evers D, Adams E, Cristol D, Eagles-Smith C, Edmonds S, Gray C, Hoskins B, Lane O, Sauer A, Tear T (2015) Songbirds as sentinels of mercury in terrestrial habitats of eastern North America. Ecotoxicology 24:453–467
Kelly JF (2000) Stable isotopes of carbon and nitrogen in the study of avian and mammalian trophic ecology. Can J Zool 78(1):1–27
Kidd KA, Hesslein RH, Fudge RJP, Hallard KA (1995) The influence of trophic level as measured by δ15N on mercury concentrations in freshwater organisms. Water Air Soil Pollut 80:1011–1015
Kirwan ML, Megonigal JP (2013) Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504:53–60
Kirwan ML, Guntenspergen GR, D’Alpaos A, Morris JT, Mudd SM, Temmerman S (2010) Limits on the adaptability of coastal marshes to rising sea level. Hydrol Land Surf Stud 37:L23401
Kramar D, Goodale WM, Kennedy LM, Carstensen LW, Kaur T (2005) Relating land cover characteristics and common loon mercury levels using geographic information systems. Ecotoxicology 14(1-2):253–262
Lane OP, O’Brien KM, Evers DC, Hodgman TP, Major A, Pau N, Ducey MJ, Taylor R, Perry D (2011) Mercury in breeding Saltmarsh Sparrows (Ammodramus caudacutus caudacutus). Ecotoxicology 20:1984–1991
Lane O, Adams EM, Pau N, O’Brien KM, Reagan K, Farina M, Schneider-Moran T, Zarudsky J. (2019). Long-term monitoring of mercury in adult saltmarsh sparrows breeding in Maine, Massachusetts and New York, USA 2000–2017. Ecotoxicology (In press)
Lee C-S, Lutcavage ME, Chandler E, Madigan DJ, Cerrato RN, Fisher NS (2016) Declining mercury concentrations in Bluefin Tuna reflect reduced emissions to the North Atlantic Ocean. Environ Sci Technol 50(23):12825–12830
Longcore JR, Haines TA, Halteman WA (2007) Mercury in tree swallow food, eggs, bodies, and feathers at Acadia National Park, Maine, and an EPA Superfund site, Ayer, Massachusetts. Environ Monit Assess 126(1-3):129–143
Lyons JE, Runge MC, Laskowski HP, Kendall WL (2008) Monitoring in the context of structured decision-making and adaptive management. J Wildl Manag 72(8):1683–1692
Mao H, Ye Z, Driscoll C (2017a) Meteorological effects on Hg wet deposition in a forested site in the Adirondack region of New York during 2000–2015. Atmos Environ 168:90–100
Mao H, Hall D, Ye Z, Zhou Y, Felton D, Zhang L (2017b) Impacts of large-scale circulation on urban ambient concentrations of gaseous elemental mercury in New York, USA. Atmos Chem Phys 17(18):11655
Marra PP, Hobson KA, Holmes RT (1998) Linking winter and summer events in a migratory bird by using stable-carbon isotopes. Science 282:1884–1886
Meehl GA, Stocker TF, Collins WD, Friedlingstein P, Gaye T, Gregory JM, Kitoh A, Knutti R, Murphy JM, Noda A et al. (2007) Global climate projections. Cambridge University Press, Cambridge, UK
Michener R, Lajtha K (2007) Stable isotopes in ecology and environmental science. Blackwell Publishing, Malden, Massachusetts, USA.
Miskimmin BM, Rudd JWMN, Kelly CA (1992) Influence of dissolved organic carbon, pH, and microbial respiration rates on mercury methylation and demethylation in lake water. Can J Fish Aquat Sci 49(1):17–22
Mitsch WJ, Hernandez ME (2013) Landscape and climate change threats to wetlands of North and Central America. Aquat Sci 75:133–149
Podar M, Gilmour CC, Brandt CC, Soren A, Brown SD, Crable BR, Palumbo AV, Somenahally AC, Elias DA (2015) Global prevalence and distribution of genes and microorganisms involved in mercury methylation. Sci Adv 1(9):e1500675
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ramlal PS, Kelly CA, Rudd JWM, Furutani A (1993) Sites of methyl mercury production in remote Canadian Shield lake. Can J Fish Aquat Sci 50:972–979
Reis AT, Rodrigues SM, Araújo C, Coelho JP, Pereira E, Duarte AC (2009) Mercury contamination in the vicinity of a chlor-alkali plant and potential risks to local population. Sci total Environ 407(8):2689–2700
Rimmer CC, McFarland KP, Evers DC, Miller EK, Aubry Y, Busby D, Taylor RJ (2005) Mercury concentrations in Bicknell’s thrush and other insectivorous passerines in montane forests of northeastern North America. Ecotoxicology 14:223–240
Risch MR, Kenski DM (2018) Spatial patterns and temporal changes in atmospheric-mercury deposition for the Midwestern USA, 2001–2016. Atmosphere 9(1):29
Rodenhouse NL, Lowe WH, Gebauer RLE, McFarland KP, Bank MS (2019) Mercury bioaccumulation in temperature forest food webs associated with headwater streams. Sci Total Environ 665:1125–1134
Sauer AK, Driscoll CT, Evers DC, Adams EM, Yang Y. (2019) Mercury exposure in songbird communities within Sphagnum Bog and upland forest ecosystems in the Adirondack Park (New York, USA). Ecotoxicology (In press)
Scheuhammer AM, Meyer MW, Sandheinrich MB, Murray MW (2007) Effects of environmental mercury on the health of wild birds, mammals, and fish. AMBIO: J Hum Environ 36:12–19
Schile LM, Callway JC, Morris JT, Stralberg D, Parker VT, Kelly M (2014) Modeling tidal marsh distribution with sea-level rise: evaluating the role of vegetation, sediment, and upland habitat in marsh resiliency. PLOS ONE 9(2):e88760
Schindler DW (2001) The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millennium. Can J Fish Aquat Sci 58:18–29
Seewagen CL (2018) The threat of global mercury pollution to bird migration: potential mechanisms and current evidence. Ecotoxicology. https://doi.org/10.1007/s10646-018-1971-z
Sellers P, Kelly CA, Rudd JWM, MacHutchon AR (1996) Photodegradation of methylmercury in lakes. Nature 380:694–697
St. Louis VL, Rudd JWM, Kelly CA, Beaty KG, Bloom NS, Flett RJ (1994) Importance of wetlands as sources of methyl mercury to boreal forest ecosystems. Can J Fish Aquat Sci 51(5):1065–1076
St. Louis VL, Rudd JWM, Kelly CA, Bodaly RA, Paterson MJ, Beaty KG, Hesslein RH, Heyes A, Majewski AR (2004) The rise and fall of mercury methylation in an experimental reservoir. Environ Sci Technol 38:1348–1358
Stern GA, Macdonald RW, Outridge PM, Wilson S, Chetelat J, Cole A, Hintelmann H, Loseto LL, Steffen A, Wang F, Zdanowicz C (2012) How does climate change influence Arctic mercury? Sci Total Environ 414:22–42
Taylor VF, Buckman KL, Seelen EA, Mazrui NM, Balcom PH, Mason RP, Chen CY (2019) Organic carbon content drives methylmercury levels in the water column and estuarine food webs across latitudes in the Northeast United States. Environ Pollut 246:639–649
Telmer KH, Veiga MM (2009) World emissions of mercury from artisanal and small scale gold mining. In: Mercury Fate and Transport in the Global Atmosphere: Measurements, Models, and Policy Implications Interim Report. UNEP Global Mercury Partnership, Chapter 6, p 96–130. Springer, Boston, MA
Townsend JM, Driscoll CT, Rimmer CC, McFarland KP (2014) Avian, salamander, and forest floor mercury concentrations increase with elevation in a terrestrial ecosystem. Environ Toxicol Chem 33:208–215
Tsui MTK, Adams EM, Jackson AK, Evers DC, Blum JD, Balogh SJ (2017) Understanding sources of methylmercury in songbirds with stable mercury isotopes: challenges and future directions. Environ Toxicol Chem 37(1):166–174
Ullrich SM, Tanton TW, Abdrashitova SA (2001) Mercury in the aquatic environment: a review of factors affecting methylation. Crit Rev Environ Sci Technol 31(3):241–293
VanArsdale A, Weiss J, Keeler G, Miller E, Boulet G, Brulotte R, Poissant L (2005) Patterns of mercury deposition and concentration in Northeastern North America (1996–2002). Ecotoxicology 14:37–52
Wada H, Cristol DA, McNabb FMA, Hopkins WA (2009) U.S. Department of Agriculture. 2015. Summary Report: 2012 National Resources Inventory, Natural Resources Conservation Service, Washington DC, and Center for Survey Statistics and Methodology. Iowa State University, Ames, Iowa
Warner SE, Shriver WG, Pepper MA, Taylor RJ (2010) Mercury concentrations in tidal marsh sparrows and their use as bioindicators in Delaware Bay, USA. Environ Monit Assess 171:671–679
Warren RS, Niering WA (1993) Vegetation change on a northeast tidal marsh: interaction of sea-level rise and marsh accretion. Ecology 74(1):96–103
Whitney MC, Cristol DA (2017) Impacts of sublethal mercury exposure on birds: a detailed review. In: Reviews of environmental contamination and toxicology, Springer, Cham, vol 244, p 113–163
Wickham H (2009) ggplot2: elegant graphics for data analysis. Springer-Verlag New York. Suppressed adrenocortical responses and thyroid hormone levels in birds near a mercury-contaminated river. Environ Sci Technol 43:6031–6038
Wickham H, Francois R, Henry L, Muller K (2018) dplyr: a grammar for data manipulation. R package version 0.7.5. https://CRAN.R-project.org/package=dplyr
Williams JW, Jackson ST (2007) Novel climates, no-analog communities, and ecological surprises. Front Ecol Environ 5(9):475–482
Wilman H, Belmaker J, Simpson J, de la Rosa C, Rivadeneira MM, Jetz-Elton W (2014) Traits 1.0: species-level foraging attributes of the world’s birds and mammals. Ecology 95:2027. https://doi.org/10.1890/13-1917.1
Windham-Myers L, Fleck JA, Ackerman JT, Marvin-DiPasquale M, Stricker CA, Heim WA, Bachand PAM, Eagles-Smith CA, Gill G, Stephenson M, Alpers CN (2014) Mercury cycling in agriculture and managed wetlands: a synthesis of methylmercury production, hy6drological export, and bioaccumulation from an integrate field study. Sci Total Environ 484:221–231
Ye Z, Mao H, Driscoll CT (2019) Primary effects of changes in meteorology vs. anthropogenic emissions on mercury wet deposition: a modeling study. Atmos Environ 198:215–225
Yu X, Driscoll CT, Huang J, Holsen TM, Blackwell BD (2013) Modeling and mapping of atmospheric mercury deposition in Adirondack Park, New York. PloS ONE 8(3):e59322
Zedler JB, Kercher S (2005) Wetland resources: status, trends, ecosystem services, and restorability. Annu Rev Environ Resour 30:39–74
Zhang Y, Jacob DJ, Horowitz HM, Chen L, Amos HM, Krabbenhoft DP, Slemr F, St. Louis VL, Sunderland EN (2016) Observed decrease in atmospheric mercury explained by global decline in anthropogenic emissions. PNAS 113(3):526–531
Acknowledgements
Sample collection occurred under all required state (NYS DEC Scientific License to Collect and Possess Permits #1873, 1893; NYS Temporary Revocable Permits #2386, 2262, 8957, 2057/8128, 1979/7493) and federal permits (USGS BBL Permit #22636). Kathryn Williams provided comments on the manuscript and assistance in field sampling. The work of many trained songbird biologists was needed for this large sampling effort; the work of Melissa Duron and Sarah Johnson is specifically acknowledged. We would also like to thank the many field technicians that provided assistance during the course of the project: Katherine Gilbert, Kylie O’Driscoll, Mike Brennan, Paul Josephson, Tom Daniel, Lyneé Sauer, and Bob Sauer. We would like to acknowledge the many individuals and organizations for their generous support and collaboration as part of our research efforts: Adirondack League Club, Elizabeth Ballantine, Dan Josephson, Neil Gifford and the Albany Pine Bush Preserve, Black Rock Forest Consortium, Cornell University, Boston University, Harvard University, Massawepie Scout Camps, NYS Department of Environmental Conservation, New York State Parks, Frost Valley YMCA, SUNY-ESF Huntington Wildlife Forest, Syracuse University, Michael Farina, Rob Longiaru, Tara Schneider-Moran, John Zarudsk and many others with The Town of Hempstead Department of Conservation and Waterways, Mashomack Preserve, The Nature Conservancy (Adirondack Chapter, Central and Western NY Chapter, Eastern NY Chapter, Joe Jansen, Nicole Maher, Derek Rogers and many others from the Long Island Chapter), Alison Kocek and the field crew from SUNY ESF for collecting samples in the New York City region, and the US Geological Survey. This work could not have been done without extensive publicly available online resources. We acknowledge the World Climate Research Programme's Working Group on Coupled Modeling, which is responsible for CMIP, and we thank the climate modeling groups for producing and making available their model output. For CMIP the U.S. Department of Energy's Program for Climate Model Diagnosis and Intercomparison provides coordinating support and led development of software infrastructure in partnership with the Global Organization for Earth System Science Portals.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
This study was funded by the New York State Energy Research and Development Authority (NYSERDA, Award # 34358). All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix. Estimates of methylmercury exposure across all species and all regions
Appendix. Estimates of methylmercury exposure across all species and all regions
Eighty-three species were sampled across all regions, including 214 unique species/region combinations. Here we include a figure that summarizes the average blood Hg concentrations (ppm ww) of all species/region combinations as estimated in the analysis. These averages (and 95% confidence intervals) are based on the generalized linear mixed modeling approach described in the text to identify species with elevated blood Hg concentrations in each region. Here we document the results for all species (Table 2).
Rights and permissions
About this article

Cite this article
Adams, E.M., Sauer, A.K., Lane, O. et al. The effects of climate, habitat, and trophic position on methylmercury bioavailability for breeding New York songbirds. Ecotoxicology 29, 1843–1861 (2020). https://doi.org/10.1007/s10646-019-02151-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-019-02151-w