
Abstract
Knowledge of the feeding ecology of fishes is fundamental for understanding the processes that function at the individual, population, and community levels and for the conservation of their populations and habitats. Spirlins are widely distributed and often abundant in fast-flowing waters throughout Europe. However, data on their diets are insufficient and inconsistent. To improve knowledge of the trophic ecology of this common fish species, we studied the diet of Russian spirlin in the rhithral and potamal of rivers located in the Volga River basin. The food niches of spirlin in the rhithral and potamal differed significantly. Fishes consumed mainly terrestrial prey falling into the water in rhithral but aquatic prey in potamal of watercourses. Among aquatic invertebrates, spirlins positively selected mobile species that foraged on the upper surface of the bottom substratum. Aufwuchs were abundant in stomachs, but, apparently, fish did not assimilate them, because intact cells occurred throughout the intestine. Spirlins in all rivers were predominantly insectivorous, not consuming or rarely consuming invertebrates that dominated zoobenthos, mollusks, oligochaetes, and leeches. This specialization, apparently, contributes to the resource partitioning with other fishes inhabiting hyporhithral and epipotamal of watercourses.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Running waters are among the most impacted of all natural ecosystems (Malmqvist and Rundle 2002). Eutrophication, pollution, acidification, overharvesting, introductions of non-indigenous species, and habitat destruction are the threats to the inhabitants of running waters and the goods and services they provide (Carpenter et al. 1992). Recently, the impact of adverse factors may be exacerbated due to climate change, because it induces changes in hydrological conditions, primary productivity, biogeochemistry, and species composition of biological communities (Golubkov and Golubkov 2020; Golubkov 2021). The result is a radical restructuring of many food webs, and a final effect on fish assemblages may be related to the quality and availability of prey under stress conditions (Pletterbauer et al. 2015). In order to anticipate this effect, detailed knowledge of the food selectivity and ecological niches of different fish species is required.
The European spirlin, Alburnoides bipunctatus (Bloch, 1782), is a small fish inhabiting streams and rivers, usually with fast-flowing waters, often in upland and montane areas. This is a previously widespread and abundant species, whose populations have declined dramatically in response to habitat deterioration, because it is vulnerable to changes in hydrological regime and pollution (Aarts and Nienhuis 2003; Kottelat and Freyhof 2007; Trautwein et al. 2013; Marszal et al. 2018). The Russian spirlin was originally described as an eastern subspecies of the European spirlin: Alburnoides bipunctatus rossicus Berg, 1932. Recently, due to high morphological and genetic differences, it began to be considered an independent species, Alburnoides rossicus Berg (Bogutskaya and Coad 2009; Stierandová et al. 2016).
Diet data for European spirlin are inconsistent. Insect larvae and imagoes prevailed in the diet of European spirlin in the tributary of the Vistula River (Poland) (Marszal et al. 2018), while, in Sava River, Croatia, the dominant food item was algae, and invertebrates were a secondary or an accidental prey (Treer et al. 2006). Data of the diet and feeding habitats of the Russian spirlin are very scare (Kotelnikova 2016).
The information on distribution patterns and ecological guilds of macroinvertebrates and fish can be used to assess and manage the ecological integrity of rivers and their biomonitoring (Aarts and Nienhuis 2003). Longitudinal zonation concepts describe the downstream changes in chemico-physical and biological properties of rivers. One of the most popular concepts distinguishes three main zones from the source to the mouth of the river: the crenal zone (close to the river source), the rhithral, and the potamal zones (Illies 1961; Aarts and Nienhuis 2003; Ficsór and Csabai 2021). Rhithral usually refers to the upper parts of the watercourse, located in the foothills, with rocky or gravel-pebble bottom, high flow rate and oxygenated water. The potamal belongs to the lower part of the watercourse adjacent to the rhithral, with a sandy, silted, or silty bottom, with a relatively low discharge and frequent bottom hypoxia. Taking into account this zonation system, spirlins are common in lower rhithral and upper potamal zones (Aarts and Nienhuis 2003).
Spirlins have no commercial value, but due to their large numbers in some rivers, they can be a significant competitor to other more valuable fish species. They also can be an important food item for predatory fish and thus play an important role in the food webs of the river ecosystem. In this context, the aim of our work was to improve knowledge of the trophic ecology of this fish species. We tested the hypothesis that Russian spirlin is a polyphagous opportunist with poor food selectivity, and its diet mainly depends on the specific composition of aquatic communities. To achieve this aim, we studied both the food tracts of fish and the abundance and composition of aquatic invertebrates in the river stretches that can be attributed to rhithral or potamal.
Material and methods
Study sites
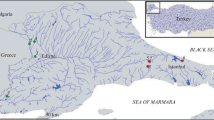
The study was performed in 2010–2017 on seven stretches of watercourses located in the basin of the Volga River (Fig. 1). The climate of the region is humid with cool summers (Kottek et al. 2006). It has a long, cold winter with a stable snow cover.

Map of the study area. Dots indicate sampling stations
Taking into account the longitudinal zonal classification of river stretches, the studied river sections belong to three types: rhithral, potamal of small and medium watercourses, and potamal of large watercourses. Their hydrological and hydrobiological characteristics are given in Table 1. The studied stretch of the Bui River can be classified as hyporhithral. It has a high flow rate, a hard gravel and pebble bottom substratum, and dense woody vegetation on steep banks. The rest of the studied river sections can be attributed to the transition zone from rhithral to potamal, to epipotamal. The current velocities are lower there; sand with gravel and pebble with a large amount of attached algae and higher aquatic vegetation predominated in the grounds. The water temperature in rivers during the sampling periods varied from 12.5 to 16.7 °C. There was no anthropogenic impact on the studied river stretches.
Field survey
The fish were collected using pulsed backpack electrofishing equipment ELLOR-2 and two fry seine nets. The first net had a length of 5.0 m, a height of 1.5 m, and a mesh diameter of 4 mm. It was used on river sections, the width of which did not exceed 5–9 m. The second net had a length of 10 m, a height of 1.8 m, and a mesh diameter of 3 mm. This net was used on stretches of wider rivers. Captured fish was preserved in 8% formaldehyde. A total of 1004 individuals of spirlin with a total length (TL) of 17 to 118 mm were caught for the analysis of their diet (Table 2). Most of the fish were caught in the morning. Since almost all spirlins were caught in the riffles, samples of zoobenthos were collected there.
To match spirlin diet with prey availability, samples of potential prey species were collected. They included 76 samples of zooplankton and 78 samples of zoobenthos. Zoobenthos and zooplankton samples were taken in all rivers except the Lasva River. Zoobenthos was sampled in 4–6 replicates from randomly selected locations using a Surber sampler with a capture area of 625 cm2 and a mesh size of 0.2 mm at depths of up to 0.5 m. At deeper sites (up to 1.2 m), a hydrobiological scraper with a blade length of 0.2 m was used. Samples of invertebrates were sieved in a 0.25 mm mesh.
Laboratory analysis
In the laboratory, invertebrates were sorted from bottom sediments and preserved in 4% formaldehyde. They were identified, counted, and weighed (wet weight, shells of mollusks included).
Total length (TL, to the nearest 1 mm) and wet weight (W, to the nearest 0.01 g) were recorded for each specimen of spirlin. Whenever possible, all macroinvertebrate components in the gut of spirlin were identified to genus or species, and the remains of prey were counted and weighed. The weight of the eaten organisms was reconstructed using the average body weight obtained by weighing and counting macroinvertebrates from hydrobiological samples taken simultaneously with the fish diet samples (Borutskiy 1974).
The gut fullness index (FI, 0/000) was estimated as (Hyslop 1980):
We estimated the consumption index (CI, 0/000) to characterize the relative intensity of fish feeding in various watercourses (Borutskiy 1974):
The assessment of the composition of the diet was based on the frequency of occurrence (P, %), numerical frequency (N, %) of the various diet components, and the percentage of wet weight of food items in fish intestinal tracts (B, %):
where fi is the number of stomachs containing each prey items and ∑f is the total number of intestinal tracts;
where ni is the total number of certain food item and ∑n is the total number of food items consumed by the fish;
where bi is the wet weight of food category in all intestinal tracts of a certain fish species and ∑b is the total wet weight of all food categories in their intestinal tracts (Hyslop 1980).
The contribution of each prey category to the diet was estimated with the index of relative importance (IRI, %; Cortés 1997):
where Fi is the number of stomachs containing each prey items and Bi is the wet weight of food category in all intestinal tracts of a certain fish species.
To calculate dietary preferences, Ivlevs’ electivity index (E; Ivlev, 1961) was used:
where ri is the proportion of the certain resource in the stomach contents (as a percentage of the total recovered weight of food in intestinal tracts) and pi is the relative content of the same resource in the environment.
To compare the overall diet composition of fish in various watercourses, the Morisita-Horn index of food niche similarity (Horn 1966) was calculated:
where xi is the proportion of i-food in species xi and yi is the proportion of i-food in species yi. cλ = 0 means complete dissimilarity in food niches, and cλ = 1 means complete coincidence. An index value > 0.6 was regarded as a biologically similarity of food niches.
Statistical analyses
Statistical analyses were performed using R software version 3.4.3 (R Core Team 2021a). The one-way ANOVA of “aov” function of the “stats” package (R Core Team 2021a) was used to assess the statistical significance of the differences between the parameters of fish feeding and the values of the recovered mass of their food components in different watercourses. The post-hoc “TukeyHSD” function of the “stats” package was used for pairwise comparison.
Analysis of similarity has been used to compare the overall diet composition of different rivers’ fishes using “anosim” function of vegan R package based on assessment of Bray–Curtis distances. Non-metric multidimensional scaling (nMDS) with the 95% ellipsoids was used to visualize similarity in the spirlin diets. Ordinations were performed using relative biomass (%) of prey components identified in the gut of each individual. Permutational multivariate analysis of variance (PERMANOVA R package) (Anderson, 2008) was used with the same data followed by pairwise comparisons (Tukey’s HSD post hoc test) to test whether there were significant differences in spirlin diet among rivers types. Similarity percentage analysis (“SIMPER”) function of vegan R package using Bray–Curtis distances has been used to identify which prey taxa were most likely responsible for the patterns detected by “permanova.” It provided average dissimilarities and identified which prey components made the greatest contribution to any dissimilarity between spirlin diets in different rivers. The value of the standard deviation of the mean is given under the ± sign.
Results
Species composition of aufwuchs and macroinvertebrates
Green algae Ulothrix and Cladophora were abundant in most of the studied rivers. Zooplankton included a small number of species that were found mainly near the riverbanks.
Insect larvae, oligochaetes, leeches, bivalves, and gastropods were found in zoobenthos (Supplemental Table S1). Detritus feeders from oligochaetes of the family Tubificidae or bivalve mollusks from Unionidae and Pisidiidae were predominant in the biomass of zoobenthos in all rivers (Fig. 2). Insect larvae were the most abundant and had the greatest species richness. Significant part of them included rheophilic forms inhabiting hard substratum and the mats of attached algae. Larvae of mayfly Caenis macrura, caddisflies Hydropsyche contubernalis and Psychomyia pusilla, bug Aphelocheirus aestivalis (Heteroptera), swamp mosquito Hexatoma bicolor (Limoniidae), and chironomids Polypedilum scalaenum were common. The species richness and diversity of macroinvertebrates were low, and the importance of species inhabiting the silty substratum (oligochaetes and marsh mosquitoes) was high in the Ilet River, where sandy and silty substrata predominated. In the Bui, Mesha, Cheptsa, and Uruzan rivers with gravel-pebble substrate (Table 1), the species richness and diversity of benthic invertebrates, as well as the proportion of large caddisflies and mayflies, were high (Fig. 2). Periphyton mats, which silted up during the growing season, were colonized by many species of scrapers from Gastropoda, selective algophages from Chironomidae and phyto-detritivorous species from Baetidae (Ephemeroptera) (Supplemental Table S1).

The share of the main components in the total biomass of zoobenthos in various watercourses
Diet composition and food electivity of spirlin
The content of the intestinal tracts of spirlin in the studied biotopes was diverse and consisted of aquatic and terrestrial invertebrates and plants. Aquatic invertebrates included insects, arachnids, and oligochaetes. No zooplankton species were found in their stomachs.
Aquatic insects were the most diverse food component of spirlins (Supplemental Table S2). Spirlins showed high-positive food electivity for many of them. They strongly selected the mayfly larvae of Baetis spp. in most streams and Heptagenia coerulans and Serratella ignita in the Uruzan River (Table 3). However, spirlins had negative electivity for larvae of Caenis macrura or Ephemera lineata.
Spirlin had a high positive selectivity for larvae of caddisflies Hydropsyche contubernalis and Psychomyia pusilla in the Bui River (Table 3), where they were numerous in the benthic communities. In addition, many imagoes of Hydropsyche spp. were found in the stomachs of spirlin in this river. The fish apparently consumed them during insect emergence. Larvae of H. contubernalis were also a significant item in the diet of spirlin in the potamal of the Mesha River, where it positively selected them (Table 3). Caddisflies formed up to 31% of the recovered weight of food and had a high value of IRI in the fish diet in this watercourse (Table 4). The H. contubernalis and P. pusilla were also numerous and had high frequency of occurrence in the intestines and made up a significant part of spirlin diet in the potamal of many other watercourses. Nevertheless, spirlins had negative or neutral selection for these and other caddisfly larvae in most rivers (Table 3).
Diptera were the most diverse component in the spirlin diet (Supplemental Table S2), although fish showed negative selectivity for most species (Table 3). Their proportion in intestinal tract contents and values of the IRI were relatively small, with the exception of the Ilet River, where these indices were high (Table 4). Orthocladius rhyacobius had the highest frequency of occurrence (P) in the diet of spirlin.
Spirlin showed positive selection for the swamp mosquito Hexatoma bicolor in the Bui River, where this species was common. However, in the potamal, fish negatively selected this species. Midge larvae Simulium ornatum had high values of P and IRI in the diet of the spirlin in some watercourses (Table 4). However, they were not found in the benthos of the studied stretches.
Terrestrial insects, ants Myrmica laevinodis, larvae of ground beetle Plagiodera versicolora, and imagoes of ground bugs from Aphididae were abundant in the diet of spirlin in the Bui River. P of Hymenoptera and Heteroptera in its diet were 27 and 23%, respectively (Table 4).
Diverse plant components were found in the intestinal tracts of spirlins (Supplemental Table S2). Filamentous algae Ulothrix spp. and Cladophora spp. had the highest occurrence in their stomachs, especially in the Bui River (Table 4). However, the algae throughout the intestine had intact cells with chlorophyll.
Gut fullness and dissimilarity of spirlins’ dietary niches in various watercourses
Spirlins had the highest values of the FI and CI indices in the rhithral of the Bui River, where their values were significantly higher than in other watercourses (Table 5, FI: F = 16.46, p < 0.001; CI: F = 24.63, p < 0.001). In the potamal of most other watercourses, there were no significant differences in these parameters. The largest proportion of feeding spirlins and the number of food components was found in the Bui River (Table 5). Of the animal prey, most were terrestrial invertebrates (Fig. 3). The largest number of consumed invertebrates per individual and reconstructed weight of the intestinal tract contents were recorded in fish from the potamal of the Vala River (Table 5, F = 55.27, p < 0.001).

The share of the main components in the weight content of the intestinal tracts of spirlin in various watercourses
According the values the Morisita-Horn index, the similarity of spirlin food niches in the rhithral of the Bui River and in the potamal of other studied watercourse was very low. Spirlins had high food niche similarity in the potamal of small and medium watercourses: Lasva, Vala, and Mesha rivers (Table 6).
One-way “permanova” showed the dissimilarity of between spirlin diets in different rivers (pseudo-F = 32.38, P = 0.001). Pairwise comparisons showed that the diet of spirlins differed significantly in three types of watercourses: in hyporhithral of the Bui River, in the epipotamal of small and medium watercourses (Vala, Mesha, Lasva rivers), and in the epipotamal of large watercourses (Cheptsa and Uruzan rivers). Non-metric multidimensional scaling showed significant differences in spirlin diets between these groups of watercourses in the ordination plot (Fig. 4). The “anosim” function of vegan R package indicated dissimilarity (0.367) of spirlin diets in different rivers.

Non-metric multidimensional scaling ordination plot showing spirlin diet overlap in investigated rivers. Stress function value is 0.367
Differences in spirlin diet between these groups of rivers were confirmed by “SIMPER.” It showed the importance of 15 components, which contributed nearly 70% of overall dissimilarity between the diet of Alburnoides rossicus in studied rivers (Table 7). These components may be grouped in three types of prey categories: filamentous green algae (Ulothrix, Cladophora), adult terrestrial insect (bugs, ants, aphids), and larvae of aquatic insects (chironomids, blackflies, mayflies, caddisflies).
Discussion
Dams are prevalent impacts on hydromorphology in rivers across the world, fragmenting river ecosystems and reducing catchment scale connectivity (Nilsson et al. 2005). Reduced water flow may affect fish in fast-flowing rivers at both individual and assemblage levels not only directly, but also via reduced resource availability (Elosegi et al. 2010). Frequent droughts also reduced resource availability for fish (Elosegi et al. 2010).
In Western Europe, spirlin populations have declined dramatically in recent decades resulted mainly from damming and deterioration of species’ fast-flowing water habitat (Marszal et al. 2018). However, in the rivers studied by us, Russian spirlin was among the dominant fish species (Kotelnikova 2016). This is apparently explained by the absence of a significant anthropogenic impact on these rivers. Therefore, the conditions in these rivers can be considered pristine for Europe, which may be important for the development of methods for the restoration of disturbed habitats in them.
The key role of in the structure of the studied macroinvertebrate communities of detritovores (scrapers, collectors, filterers) indicates the great importance in their feeding of fine particulate organic matter carried out by the flow from the headwater stretches of the rivers, as predicted by the River Continuum Concept (Allan and Castillo 2007).
Diet composition and food electivity of spirlin
Influence of prey traits on predator selectivity is a prerequisite for the understanding of community processes (Worischka et al. 2015). In our study, spirlin positively selected mayfly larvae of Baetis spp., apparently due to their high mobility, small size, and soft integument, which makes them easily accessible and digestible food item. Spirlin also positively selected Heptagenia coerulans и Serratella ignita. The larvae of these species are grazers and feed on epiphytes on the upper surface of the substrate, where fish easily consume them. This confirms that macroinvertebrate feeding type is one of the most important traits influencing the prey selectivity of the riverine benthivorous fish, which generally prefer macroinvertebrate grazers and sediment feeders (Worischka et al. 2015). On the other hand, spirlin was negatively selective for the larvae of Caenis macrura and Ephemera lineata (Table 3). The first mayfly species are common in packets of leaf litter, while the second builds burrows in sandy substrates. In these microbiotopes, they are poorly accessible to fish.
Caddisflies were a significant component in the spirlin diet in most of the rivers studied. However, the selectivity of spirlin for them ranged from strongly positive to negative (Table 3). Fish positively selected larvae of Hydropsyche contubernalis in the Bui and Mesha rivers and negatively in most other streams (Table 3). This may be due to the different microdistribution of the larvae in various watercourses. The larvae of Hydropsyche forage sometimes as collectors but are also able to feed on animal food or graze on periphyton (Ficsór and Csabai 2021). The omnivorous larvae of these caddisfly may occupy the bottom, lateral sides, or top of rocks depending on environmental conditions (Voelz and Ward 1996). As predators, they rely primarily on macroinvertebrate drift, being located on the upper surface of rocks (Wallace and Webster 1996). Hydropsychid larvae often depend on animal diet in headwaters (Bing et al. 2015) similar to the rhithral of the Bui River. In this case, they should be more vulnerable to fish predation as compared to downstream stretches of rivers.
Chironomid larvae were abundant in the rivers we studied and were a significant component in the spirlin diet. However, in most cases, fish selected them negatively (Table 3). The reason, apparently, was that these invertebrates mainly inhabited biotopes with epiphytes, which served them as a refuge from fish predation, because physical complexity of microhabitats reduced foraging efficiency of fish (Kornijów 1997; Nunn et al. 2012).
Earlier the importance of Ephemeroptera, Trichoptera, and Chironomidae as food for Alburnoides spp. was shown in streams of Poland, Croatia, European Russia, and Iran (Piria et al. 2005; Abbasi et al. 2013; Kotelnikova 2016; Marszal et al. 2018).
Although Simuliidae were not found in the benthos of most watercourses we studied, they were a significant component of the diet of spirlins in many of them. We believe that spirlin consumed these invertebrates from the water column as they drifted from upstream river stretches, because some studies indicated that this fish species effectively fed on drifting animals (Kotelnikova 2016; Marszal et al. 2018).
Filamentous algae were often found in spirlin stomachs in the rivers studied by us and some other rivers (Piria et al. 2005; Treer et al. 2006; Kotelnikova 2016; Abbasi et al. 2013; Marszal et al. 2018; Treer et al. 2006). However, the nutritional value of this component seems dubious, because we found that the algae throughout the intestine had intact cells with chlorophyll. This means that the fish seem to be poorly assimilating their contents. Aufwuchs is often considered a poor food resource because of its low digestibility and nutritive value (Nunn et al. 2012) . The extensive consumption of aufwuchs by fishes is probably linked to a low availability of suitable animal prey (Nunn et al. 2008). In our case, spirlins apparently consumed filamentous algae by catching macroinvertebrates that lived among them. In other words, these algae were probably a concomitant non-targeted component in their diet.
In the studied rivers, spirlins used a smaller part of zoobenthos for food. Most of the benthos biomass consisted of mollusks, oligochaetes, and leeches, which were not consumed or poorly consumed by spirlin. At the same time, it is known that other fish species with high biomasses in the studied river stretches, Squalius cephalus, Phoxinus phoxinus, Perca fluviatilis, Thymallus thymallus, and Gobio gobio, are capable of consuming these invertebrates to varying degrees (e.g., Hellawell 1971; Kennedy and Fitzmaurice 1972; Rask, 1986; Copp 2008; Balestrieri et al. 2006; Worischka et al. 2012, 2015; Mustamäki et al. 2014; Smoliński and Glazaczow 2019). Therefore, there is a resource partitioning between spirlin and other abundant fish species, which decrease competition with them. Resource partitioning facilitates the co-existence of ecologically similar species and community stability (Nunn et al., 2020). On the other hand, underutilization of autochthonous invertebrates by spirlins can create a lack of food for them, especially after the period of mass emergence of aquatic insects. As a result, these fish had to replenish their diet with allochthonous terrestrial insects that fall to the surface of the water from the riverbanks.
Food webs in lower stream orders, as a rule, have high contribution of allochthonous sources due to dense canopy cover, which generally induces a strong linkage to terrestrial subsidies (Allan and Castillo 2007; Doi 2009). The hyporhithral of the Bui River had the dense riparian vegetation along the banks. As a result, terrestrial insects, which fell from trees into the water, played an important role in the spirlin diet in this river. In addition, high flow rate and erosion of the riverbanks led to the flushing into the river of large numbers of ants, which also played a significant role in the feeding of spirlins in the Bui River. As a result, allochthonous insects predominated in the diet of spirlin in hyporhithral, while in epipotamal, this fish species predominantly consumed autochthonous aquatic insects (Fig. 3), which led to a significant difference in food niches of spirlin in the potamal and rhithral of the studied rivers (Table 6).
Aquatic and terrestrial insects are important items in fish diet in rhithral habitats. For instance, terrestrial invertebrates provided an important energy subsidy for brown trout (Rincón and Lobón-Cerviá 1999; Dineen et al. 2007). In contrast, in the potamal, benthivorous fish feed mainly on zoobenthos, including species burrowing into bottom sediments (Lik et al. 2017). Spirlins are usually most abundant in the transition zones between the rhithral and potamal of rivers (Aarts and Nienhuis 2003) where, as was shown in our study and others (Piria et al. 2005; Kotelnikova 2016; Abbasi et al. 2013; Marszal et al. 2018), they are widely used for food aquatic and terrestrial insects that are common to rhithral. However, their use of invertebrates from the groups typical for the potamon zone, such as mollusks, oligochaetes, and burrowing insect larvae, is limited. Therefore, spirlins can be considered specialized feeder that feeds on autochthonous and allochthonous aquatic insects, rather than opportunistic polyphages.
Data availability
Datasets supporting the conclusions of this article are included within the article and its supplemental file.
References
Aarts B, Nienhuis P (2003) Fish zonations and guilds as the basis for assessment of ecological integrity of large rivers. Hydrobiologia 500:157–178. https://doi.org/10.1023/A:1024638726162
Abbasi F, Ghorbani R, Yelghi S, Hajimoradloo A, Molaei M (2013) Comparative analyses of the diet of the spirlin (Alburnoides eichwaldii) in the Tilabad, Shirabad and Kaboodval streams Golestan Province. Iran World J Fish Marine Sci 5(1):79–83
Allan JD, Castillo M (2007) Stream ecology: structure and function of running waters, 2nd edn. Springer, Dordrecht, The Netherlands
Anderson MJ (2008) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26(1):32–46. https://doi.org/10.1111/J.1442-9993.2001.01070.PP.X
Balestrieri A, Prigioni C, Remonti L, Sgrosso S, Priore G (2006) Feeding ecology of Leuciscus cephalus and Rutilus rubilio in southern Italy. Ital J Zool 73(2):129–135. https://doi.org/10.1080/11250000600679561
Bing T, Müller J, Glaser B, Brandl R, Brändl M (2015) Variation in diet across an elevational gradient in the larvae of two Hydropsyche species (Trichoptera). Limnologica 52:83–88. https://doi.org/10.1016/j.limno.2015.04.001
Bogutskaya NG, Coad BW (2009) A review of vertebral and fin-ray counts in the genus Alburnoides (Teleostei Cyprinidae) with a description of six new species. Zoosystematica Rossica 18(1):126–173. https://doi.org/10.31610/zsr/2009.18.1.126
Borutskiy EV (1974) Methodological guide for the study of nutrition and food relations of fish in natural conditions. Nauka, Moscow (In Russian)
Carpenter SR, Fisher SG, Grimm NB, Kitchell JF (1992) Global change and freshwater ecosystems. Annu Rev Ecol Syst 23:119–139. https://doi.org/10.1146/annurev.es.23.110192.001003
Copp GH (2008) Putting multi-dimensionality back into niche: diel vs day-only niche breadth separation in stream fishes. Fund Appl Limnol 170(4):273–280. https://doi.org/10.1127/1863-9135/2008/0170-0273
Cortés E (1997) A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Can J Fish Aquat Sci 54:726–738. https://doi.org/10.1139/F96-316
Dineen G, Harrison SS, Giller PS (2007) Diet partitioning in sympatric Atlantic salmon and brown trout in streams with contrasting riparian vegetation. J Fish Biol 71:17–38. https://doi.org/10.1111/j.1095-8649.2007.01441.x
Doi H (2009) Spatial patterns of autochthonous and allochthonous resources in aquatic food webs. Popul Ecol 51:57–64. https://doi.org/10.1007/s10144-008-0127-z
Elosegi A, Díez J, Mutz M (2010) Effects of hydromorphological integrity on biodiversity and functioning of river ecosystems. Hydrobiologia 657:199–215. https://doi.org/10.1007/s10750-009-0083-4
Ficsór M, Csabai Z (2021) Longitudinal zonation of larval Hydropsyche (Trichoptera: Hydropsychidae): abiotic environmental factors and biotic interactions behind the downstream sequence of Central European species. Hydrobiologia 848:3371–3388. https://doi.org/10.1007/s10750-021-04602-0
Golubkov SM (2021) Effect of climatic fluctuations on the structure and functioning of ecosystems of continental water bodies. Contemp Probl Ecol 14(1):1–10. https://doi.org/10.1134/S1995425521010030
Golubkov M, Golubkov S (2020) Eutrophication in the Neva Estuary (Baltic Sea): response to temperature and precipitation patterns. Mar Freshwater Res 71(6):583–595. https://doi.org/10.1071/MF18422
Hellawell JM (1971) The food of the grayling Thymallus thymallus (L.) of the River Lugg. Herefordshire J Fish Biol 3:187–197. https://doi.org/10.1111/j.1095-8649.1971.tb03662.x
Horn HS (1966) Measurement of “overlap” in comparative ecological studies. Am Nat 100:419–424. https://doi.org/10.1086/282436
Hyslop EJ (1980) Stomach content analysis – a review of methods and their application. J Fish Biol 17:411–429. https://doi.org/10.1111/j.1095-8649.1980.tb02775.x
Illies J (1961) Versuch einer allgemeinen biozonotischen gliederung der fliessgewasser. Int Rev Hydrobiol 46(2):205–213
Ivlev IS (1961) Experimental ecology of the feeding of fishes. Yale University Press, New Haven, Connecticut, USA
Kennedy M, Fitzmaurice P (1972) Some aspects of the biology of gudgeon Gobio gobio (L.) in Irish waters. J Fish Biol 4:425–440. https://doi.org/10.1111/j.1095-8649.1972.tb05690.x
Kornijόw R (1997) The impact of predation by perch on the size-structure of Chironomus larvae—the role of vertical distribution of the prey in the bottom sediments, and habitat complexity. Hydrobiologia 342:207–213. https://doi.org/10.1023/A:1017067621668
Kotelnikova VS (2016) Feeding of Russian spirlin Alburnoides rossicus Berg, 1924. Proc Zool Inst RAS 320(3):326–335 (In Russian). https://doi.org/10.31610/trudyzin/2016.320.3.326
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World Map of the Köppen-Geiger climate classification updated. Meteorol Z 15:259–263. https://doi.org/10.1127/0941-2948/2006/0130
Kottelat M, Freyhof J (2007) Handbook of European freshwater fishes. Publications Kottelat, Cornol and Freyhof, Berlin
Lik J, Dukowska M, Grzybkowska M, Leszczyńska J (2017) Summer co-existence of small-sized cyprinid and percid individuals in natural and impounded stretches of a lowland river: food niche partitioning among fishes. J Fish Biol 90:1609–1630. https://doi.org/10.1111/jfb.13256
Malmqvist B, Rundle S (2002) Threats to the running water ecosystems of the world. Environ Conserv 29:134–153. https://doi.org/10.1017/S0376892902000097
Marszal L, Grzybkowska M, Blonska D, Leszczynska J, Przybylski M (2018) Diet of the endangered spirlin (Alburnoides bipunctatus) at the centre of its distribution in Europe. Mar Freshw Res 69(11):1712–1723. https://doi.org/10.1071/MF18013
Mustamäki N, Cederberg T, Mattila J (2014) Diet, stable isotopes and morphology of Eurasian perch (Perca fluviatilis) in littoral and pelagic habitats in the northern Baltic Proper. Environ Biol Fish 97:675–689. https://doi.org/10.1007/s10641-013-0169-8
Nilsson C, Reidi CA, Dynesius M, Revenga C (2005) Fragmentation and flow regulation of the world’s large river systems. Science 308:405–408. https://doi.org/10.1126/science.1107887
Nunn AD, Harvey JP, Cowx IG (2008) Winter diet and condition of two 0+ year cyprinid fish species in the lower River Trent, England. J Fish Biol 72:1529–1533. https://doi.org/10.1111/j.1095-8649.2008.01799.x
Nunn AD, Tewson LH, Cowx IG (2012) The foraging ecology of larval and juvenile fishes. Rev Fish Biol Fisher 22:377–408. https://doi.org/10.1007/s11160-011-9240-8
Nunn AD, Vickers LH, Mazik K, Bolland JD, Peirson G, Axford SN, Henshaw A, Cowx IG (2020) Dynamic competition and resource partitioning during the early life of two widespread, abundant and ecologically similar fishes. Hydrobiologia 847:2211–2224. https://doi.org/10.1007/s10750-020-04247-5
Penczak T, Głowacki Ł, Galicka W, Koszalińsk H (1998) A long-term study (1985–1995) of fish populations in the impounded Warta River, Poland. Hydrobiologia 368:157–173. https://doi.org/10.1023/A:1003246115666
Perkins DM, Reiss J, Yvon-Durocher G, Woodward G (2010) Global change and food webs in running waters. Hydrobiologia 657:181–198. https://doi.org/10.1007/978-94-007-0608-8_13
Piria M, Treer T, Aničić I, Safner R, Odak T (2005) The natural diet of five cyprinid fish species. Agric Conspec Sci 70:21–28
Pletterbauer F, Melcher AH, Ferreira T, Schmutz S (2015) Impact of climate change on the structure of fish assemblages in European rivers. Hydrobiologia 744:235–254. https://doi.org/10.1007/s10750-014-2079-y
R Core Team (2021a) The R project for statistical computing (Version 4.1.0) [Software]. Retrieved from http://www.r-project.org.
R Core Team (2021b) The R stats package retrieved from https://stat.ethz.ch/R-manual/R-devel/library/stats/html/00Index.html
Rask M (1986) The diet and feeding activity of perch, Perca fluviatilis L., in a small lake in southern Finland. Annales Zoologici Fennici 23:49–56. http://urn.fi/URN:NBN:fi-fe201705296927
Rincón PA, Lobón-Cerviá J (1999) Prey-size selection by brown trout (Salmo trutta L.) in a stream in northern Spain. Can J Zool 77:755–765. https://doi.org/10.1139/z99-031
Smoliński S, Glazaczow A (2019) Cascading effects of temperature alterations on trophic ecology of European grayling (Thymallus thymallus). Sci Rep-Uk 9:18358. https://doi.org/10.1038/s41598-019-55000-5
Stierandová S, Vukić J, Vasil’eva ED, Zogaris S, Shumka S, Halačka K, Vetešník L, Švátora M, Nowak M, Stefanov T, Koščo J, Mendel J (2016) A multilocus assessment of nuclear and mitochondrial sequence data elucidates phylogenetic relationships among European spirlins (Alburnoides, Cyprinidae). Mol Phylogenet Evol 94:479–491. https://doi.org/10.1016/j.ympev.2015.10.025
Teutschbein C, Sponseller RA, Grabs T, Blackburn M, Boyer EW, Hytteborn JK, Bishop K (2017) Future riverine inorganic nitrogen load to the Baltic Sea from Sweden: an ensemble approach to assessing climate change effects. Global Biogeochem Cy 31:1674–1701. https://doi.org/10.1002/2016GB005598
Trautwein C, Schinegger R, Schmutz S (2013) Divergent reaction of fish metrics to human pressures in fish assemblage types in Europe. Hydrobiologia 718:207–220. https://doi.org/10.1007/s10750-013-1616-4
Treer T, Piria M, Aničić I, Safner R, Tomljanović T (2006) Diet and growth of spirlin, Alburnoides bipunctatus in the barbel zone of the Sava River. Folia Zool 55(1):97–106
Voelz NJ, Ward JV (1996) Microdistributions, food resources and feeding habits of filter-feeding Trichoptera in the Upper Colorado River. Arch Hydrobiol 137(3):325–348. https://doi.org/10.1127/archiv-hydrobiol/137/1996/325
Wallace BC, Webster JR (1996) The role of macroinvertebrates in stream ecosystem function. Annu Rev Entomol 41:115–139. https://doi.org/10.1146/annurev.en.41.010196.000555
Worischka S, Koebsch C, Hellmann C, Winkelmann C (2012) Habitat overlap between predatory benthic fish and their invertebrate prey in streams: the relative influence of spatial and temporal factors on predation risk. Freshwater Biol 57:2247–2261. https://doi.org/10.1111/j.1365-2427.2012.02868.x
Worischka S, Schmidt SI, Hellmann C, Winkelmann C (2015) Selective predation by benthivorous fish on stream macroinvertebrates – the role of prey traits and prey abundance. Limnologica 52:41–50. https://doi.org/10.1016/j.limno.2015.03.004
Acknowledgements
The authors are grateful to the head of the Perm branch of the “Russian Federal Research Institute of Fisheries and Oceanography” (“VNIRO”) A.G. Melnikova and colleagues—E.Yu. Krainev, S.P. Ogorodov, N.A. Martynenko, and V.V. Bezmaternykh for help in the course of the work. We are grateful to Dr. Margaret Docker and two anonymous reviewers for important comments that have significantly improved an earlier version of the paper.
Funding
The study was supported by the Zoological Institute RAS (project 122031100274–7).
Author information
Authors and Affiliations
Contributions
All authors drafted, contributed to, and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval
All applicable international, national, and/or institutional guidelines for sampling, care, and experimental use of organisms for the study have been followed.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Golubkov, S.M., Kotelnikova, V.S. & Pozdeev, I.V. Abundance and feeding mode of Russian spirlin, Alburnoides rossicus, in the rhithral and potamal of Eastern European Rivers. Environ Biol Fish 105, 851–866 (2022). https://doi.org/10.1007/s10641-022-01292-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-022-01292-y