
Abstract
Contemporary patterns of morphological variation among populations reflects the interplay between historic and contemporary processes that result from selection and constraint. Using the pumpkinseed (Lepomis gibbosus), a species native to North America and introduced to Europe, we assessed the shared and unique aspects of morphological divergence in lentic and lotic environments among native and non-native populations. Ten native and thirteen non-native pumpkinseed populations were collected between 2003 and 2010 from lakes, rivers and reservoirs within the Iberian Peninsula and east-central North America. Fifteen linear external measurements among homologous landmarks that pertain to body size, fin position and fin size were taken from all sampled individuals. Eleven of these measurements were used to test for morphological differences among populations. Pumpkinseed found in lotic water bodies exhibited a more anterior placement of pectoral and pelvic fins and a deeper caudal peduncle and body than those found in lentic water bodies from the same geographic region. However, pumpkinseed also showed morphological differences between geographic origins: pumpkinseed from native populations exhibit a more posterior placement of pectoral and pelvic fins, a narrower anterior caudal peduncle and a more slender body than pumpkinseed from non-native populations. In addition, unique responses of populations to waterbodies within geographic origins revealed a shift between water body types that was opposite in direction for native and non-native populations. Native populations exhibited shorter and deeper caudal peduncles and deeper bodies in lotic habitats, whereas non-native populations showed longer and slender caudal peduncles and more slender bodies in the same type of habitat. Our study demonstrates that contemporary patterns of morphological variation among native and non-native pumpkinseed populations can be explained by contemporary selection and/or a common plastic developmental response among water bodies, historical effects related to geographic origin and unique responses of populations to habitats within geographic origin, and that the effects of history and the interaction between history and contemporary habitat were larger than contemporary processes in explaining morphological variation at this large spatial scale.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The existence of habitat-specific morphological differences in a species is a valuable tool for determining the relative magnitude of genotypic and phenotypic effects on body shape due to the unique evolutionary histories of different populations (Langerhans and DeWitt 2004). Unique histories among organisms might produce different results under similar selective forces (i.e., historical contingency; Gould and Woodruff 1990; Price et al. 2000), or natural selection might produce similar outcomes under similar environmental conditions (Schluter 1996a; Losos et al. 1998). Thus, when different groups of organisms face a common environmental gradient, their patterns of divergence might exhibit both shared and unique elements (Langerhans and DeWitt 2004). Experiencing similar environmental gradients is a characteristic common across many different organisms. For example, diversification occurs in fishes across benthic and limnetic habitats (Robinson and Wilson 1994; Schluter 1996b; Smith and Skúlason 1996), across lake and stream habitats (Sharpe et al. 2008; Kaeuffer et al. 2012) and across predator regimes (Reznick 1982; Breden et al. 1987; Johnson 2001; Langerhans and DeWitt 2004; Sharpe et al. 2008). The explicit quantification of both shared and unique aspects of diversification provides a greater understanding of the relative roles of natural selection and historical contingency in evolution (Langerhans and DeWitt 2004).
Fishes have a variety of body shapes and fin configurations that have functional roles in foraging and locomotion (Webb 1984a; Blake 2004; Langerhans 2008, 2009). Species that are adapted for cruising or steady swimming reflect features that maximize thrust and reduce drag. Thrust is maximized by a large anterior body depth and mass to minimize recoil energy losses, a high aspect ratio lunate caudal fin, and a narrow caudal peduncle. These features, together with a streamlined body shape also act to reduce drag. Species which show posterior placement of paired fins, and longer and narrower caudal peduncles are adapted for a ‘fast start’ type of propulsion (accelerating) to escape from predators, and/or to capture evasive prey. Fishes with a deep body and long, anteriorly positioned median/paired fins are well-suited for manoeuvring within complex habitats (e.g. coral reefs, kelp beds, weedy rivers and ponds) (Webb 1984a).
Morphological differentiation is an important mechanism for both range expansion and speciation (Rundell and Price 2009; Yoder et al. 2010). A good example of a species that illustrates both range expansion and speciation is the pumpkinseed Lepomis gibbosus Linnaeus. Polymorphism in sunfishes (including that of the bluegill Lepomis macrochirus Rafinesque) is interesting because it is embedded in the historical adaptive radiation of North American sunfishes (Wainwright and Lauder 1992), and occurs along the same resource and habitat axes as those related to the formation of bluegill and pumpkinseed sunfish taxa (Riopel et al. 2008).
The pumpkinseed is endemic to eastern North America. The species was introduced into European waters in the late nineteenth century (Künstler 1908), and into the north-eastern Iberian Peninsula at the beginning of the twentieth century (García-Berthou and Moreno-Amich 2000), where it has spread rapidly across Spain and Portugal (Godinho and Ferreira 1998; Elvira and Almodóvar 2001). Several studies have demonstrated that morphological differentiation occurs between pumpkinseed populations separated by large geographical distances (Vila-Gispert et al. 2007), those from adjacent lentic and lotic water bodies within the same geographic range (Brinsmead and Fox 2002; Naspleda et al. 2012), and even those within a single water body (Robinson et al. 1993; Robinson and Wilson 1996; Mittelbach et al. 1999; Gillespie and Fox 2003; Bhagat et al. 2006). Morphological differences among pumpkinseed populations are often interpreted as being the result of contemporary processes, such as adaptation to current environmental conditions, and a common plastic developmental response (Robinson et al. 2000; Gillespie and Fox 2003; Jastrebski and Robinson 2004; Riopel et al. 2008), but it seems that historical evolutionary processes also contribute to explain phenotypic variation among native pumpkinseed populations (Weese et al. 2012). Although a previous population genetics study (Detta 2011) has suggested that Iberian populations originated from source populations from eastern North America, where most of the native populations analysed in this study came from, the influence of historical evolutionary processes which have undoubtedly affected Iberian populations, have to be taken into account to explain morphological variation among native and non-native pumpkinseed populations.
Pumpkinseed develop convergent phenotypic characters across similar ecological gradients (i.e. littoral/benthic; pelagic/limnetic) (Robinson et al. 1993; Gillespie and Fox 2003; Jastrebski and Robinson 2004), which could suggest that diversifying selection drives an evolutionary response (Robinson and Wilson 1994; Robinson et al. 1996; Riopel et al. 2008) or could reflect a common plastic developmental response. On the other hand, Weese et al. (2012) pointed out that phenotypic convergence across North American regions was also influenced by the evolutionary history and the interaction between evolutionary history and contemporary habitat. Under the framework proposed by Langerhans and DeWitt (2004), one can quantify the shared and unique responses of native and non-native populations to a common environmental gradient. Shared responses would suggest that common selective pressures generate similar phenotypes across regions with similar biophysical environments, whereas unique aspects of divergence might be more reflective of historical effects related to geographic origin and local responses of populations to novel habitats. Examining the nature and relative magnitude of both shared features and unique aspects of diversification will permit the development of a stronger understanding of the relative roles of natural selection, developmental plasticity, evolutionary history and local environmental pressures in explaining the patterns of morphological variation among native and non-native pumpkinseed populations. We expect that shared responses will be of great magnitude if the common selective forces produced convergent patterns, whereas the unique responses are expected to be high when comparing distantly related populations and large local environmental differences exist.
Methods
Sample sites
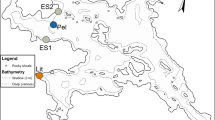
Native and non-native pumpkinseeds were collected between 2003 and 2010 from lakes, rivers and reservoirs within the Iberian Peninsula and east-central North America (Fig. 1, Table 1). Pumpkinseeds at the northern edge of their native range in Ontario, Canada were sampled from six water bodies. Balsam Lake, Rice Lake, Sturgeon Lake and the Otonabee River are part of the Trent-Severn Waterway, which forms a connection between two Laurentian Great Lakes (Lake Huron and Lake Ontario). Lake Simcoe is connected to the Laurentian Great Lakes (Lake Huron); Looncall is an inland lake situated north of Sturgeon Lake. All of these lakes have extensive shallow nearshore areas with abundant aquatic macrophytes. Fish communities within these water bodies are similar to those in other north temperate lakes, and include piscivorous species (Micropterus salmoides Lacepède; Micropterus dolomieu Lacepède; Perca flavescens Mitchill), and cyprinids (e.g., Taillon and Fox 2004). Pumpkinseeds were also collected from a reservoir in the central part of its native range (Ohio, USA), and a river and a lake near the southern end of its native range in North Carolina, USA. All three of these U.S. populations have diverse and structured communities, including piscivorous species (M. salmoides and M. dolomieu), and sunfishes (bluegill and pumpkinseed).

Sampling sites of pumpkinseed populations in North America and Iberian Peninsula. Native pumpkinseed were only collected from the American states of Ohio and North Carolina, and the Canadian province of Ontario. Non-native pumpkinseed were collected throughout Spain. Landmasses are not to scale
Iberian populations were collected from water bodies where the pumpkinseed is known to be abundant. The Ebro, Guadalquivir, Guadiana, Segura, Ter, Muga and Fluvià are Mediterranean rivers, exhibiting a high degree of seasonal water flow variation (Gasith and Resh 1999). Piscivores such as Silurus glanis Linnaeus, M. salmoides, Anguilla anguilla Linnaeus, Ameiurus melas Rafinesque and Sander lucioperca Linnaeus are present at low density in all sites, except in the sampling areas of the Ter and Fluvià rivers where piscivores are absent, and in the Ebro River where there is a high density of piscivores. Vila-Gispert et al. (2007) and Naspleda et al. (2012) provide further site descriptions. In the Guadanuño River (Guadalquivir Basin), the sampling area was located in a small pool with a low density of predators.
Susqueda, Ribaroja and Boadella reservoirs are located in north-eastern Spain in the same catchments as the Ter, Ebro and Muga rivers, respectively. These systems differ with respect to their littoral zones: Ribaroja and Boadella contain shallow littoral zones with an abundance of aquatic vegetation, whereas Susqueda lacks any major littoral areas. Ojós Reservoir, located in south-eastern Spain, is the smallest of the lentic sites, and it contains steep-sided banks and some shallow vegetated areas. Habitat and hydrological differences among these reservoirs have been previously described in Vila-Gispert et al. (2007) and Naspleda et al. (2012). Piscivores are present in all of these water bodies.
Sample collection and morphological assessment
Water conditions dictated the method of capturing pumpkinseeds in the sampling sites. In Iberian water bodies, individuals from lentic systems and the Ebro River were collected by boat electrofishing, whereas individuals from the Muga, Fluvià, Ter, Segura, Guadiana and Guadanuño rivers were collected with a shoreline electrofishing apparatus. In native water bodies, pumpkinseeds from the Otonabee River and Looncall Lake were seined, whereas those from all the other sampling sites were collected by boat electrofishing. Pumpkinseeds were sacrificed in the field and kept on ice until they were transferred to the laboratory. Individuals were then frozen and later thawed. Following the protocol of Gillespie and Fox (2003), samples were subsequently transferred to 10% neutral buffered formalin for several days, and then rinsed with water and placed in 70% ethanol.
Morphological measurements from Susqueda, Boadella, Ribaroja and Ojós reservoirs, Banyoles and Looncall lakes, and from Muga, Fluvià, Ter, Ebro-1 (Flix), Segura and Guadiana rivers, were taken from previous studies (Vila-Gispert et al. 2007; Naspleda et al. 2012). Morphological measurements for all specimens were made using digital callipers (precision 0.01 mm).
Fifteen linear external measurements among homologous landmarks that pertain to body size, fin position and fin size were taken from all sampled individuals. Fourteen of these measurements were used to calculate a centroid to correct for the effects of body size (see below), and 11 of measurements, previously identified as having functional significance for swimming or foraging in pumpkinseed and other fishes (Robinson et al. 1993; Gillespie and Fox 2003; Jastrebski and Robinson 2004), were used to test for morphological differences among populations (Fig. 2).

Modified box truss of external morphological traits, measured on the left side of each fish. Traits consist of (a) predorsal length, (b) prepectoral length, (c) prepelvic length, (d) body depth [anterior dorsal fin to pelvic insertion], (e) maximum pectoral fin length, (f) pelvic insertion to anterior anal fin, (g) anterior dorsal to posterior anal fin, (h) dorsal fin base length, (i) pelvic insertion to posterior dorsal fin, (j) anal fin base length, (k) anterior caudal peduncle depth, (l) dorsal caudal peduncle length, (m) ventral caudal peduncle length, (n) posterior caudal peduncle depth, and (o) caudal peduncle truss. This modified box truss was used to calculate centroid size, and encompasses eleven morphological traits that are identified as functionally significant for locomotion in pumpkinseed (a, b, c, d, e, h, j, k, l, m, n)
Statistical analysis
We examined morphological variation among native and non-native populations between water body types (lotic vs. lentic) using a nested Multivariate Analysis of Covariance (MANCOVA). Dependent variables were 11 external morphological measurements (log10 transformed to meet the assumption of normality), while independent variables included body size with centroid as covariate, geographic origin (native vs. non-native), water body type (lotic vs. lentic), all possible interactions, and populations nested within water body type-geographic origin combinations. All non- predictor variables that were not significant under the full model were removed all at once from the analysis. For all remaining factors in the model, MANCOVA was used to generate canonical variates (CVs) that best explained the variation associated with that factor. The factors of particular interest include water body type (shared morphological divergence among native and non-native populations), geographic origin (specific history effect), and the interaction between water body type and geographic origin (unique aspects of divergence between native and non-native populations). To evaluate the relative importance of these factors in the model, we estimated effect size using Partial eta squared (η2), which is a measure of the partial variance explained by a given factor. Graphical representation of the canonical variates for water body type and the interaction between water body type and origin were made using MorphoJ, geometric morphometrics software (Klingenberg 2011).
Because we included centroid size as a covariate, our analysis provided a test for homogeneity of slopes. Slopes were homogenous between the interaction of geographic origin and water body type; however slopes were not homogenous for either water body type or geographic origin. The centroid size x water body type and centroid size x geographic origin interaction terms were centered (i.e. the covariate was converted to deviation scores from a mean of 0) to provide meaningful hypothesis tests of the water body type and geographic origin terms by avoiding multicollinearity problems between water body type and geographic origin terms with their interaction terms (Tabachnick and Fidell 2001).
To determine whether body size of our pumpkinseed population samples could have influenced our MANCOVA results, we tested for differences in body size (centroid) between geographic regions and waterbody types. We conducted this analysis using a Linear Mixed Effects Model, with geographic origin and water body type as fixed factors and population as a random factor.
We used discriminant function analyses (DFA) to examine the strength of factor effects and to determine the percentage of individuals correctly classified by the discriminate function. Each DFA used morphological variables as the dependent variables and one particular factor as the independent variable. We used the resulting DFA to classify fish by geographic origin irrespective of water body type, water body type irrespective of geographic origin, and geographic origin-water body type combinations. The first DFA evaluated the magnitude of morphological differences between geographic origins, and the second examined the magnitude of shared features of morphological divergence between water body types. The third DFA was done to assess the increase in predictive ability over the second DFA when both shared and unique elements of divergence were used to water body type classification (Langerhans and DeWitt 2004). As a cross-validation technique, we conducted DFA’s using jackknife sampling, which removed one individual from the data set, classified that individual based on a DFA of the remaining data, returned the individual to the data set, and then repeated this process for each individual. Statistical analyses were conducted using RStudio software (version 3.2.3) and SPSS Statistics 23.
Results
The nested MANCOVAs showed strong effects of geographic origin and moderate effects of both water body type and geographic origin x water body type interaction on morphological traits (Table 2). Given the large variation in body size among populations (Table 1), the analysis of body size (centroid) differences using a linear mixed effect model with geographic origin and water body type as fixed effects and population as a random factor showed that the sampled pumpkinseed in North America were significantly larger than those sampled from the Iberian Peninsula (F1,19 = 11.15, P = 0.0035), but there were no significant differences in body size between those taken from lentic and lotic water bodies (F1,19 = 0.646, P = 0.4313) nor between those from North American and Iberian lentic and lotic water bodies (F1,19 = 0.110, P = 0.7436).
The effect of water body type suggests significant morphological divergence between populations living in lotic and lentic water bodies, regardless of geographic origin. Canonical variates derived from this effect indicate a shared axis of diversification (Fig. 3), although a large amount of variation due to geographic origin remains (Table 2). Morphological shifts were evident from the examination of the canonical loadings from the water body type: pumpkinseed found in lotic water bodies exhibited a more anterior placement of pectoral and pelvic fins, a deeper anterior caudal peduncle and a deeper body than those found in lentic water bodies from the same geographic region (Table 3, Fig. 3). However, the shared axis of morphological divergence also identifies differences between geographic origins: pumpkinseed from native populations exhibit a more posterior placement of pectoral and pelvic fins, a narrower anterior caudal peduncle and a more slender body than pumpkinseed from non-native populations (Fig. 3). This divergence pertained to the greater effect size of geographic origin than that of water body type (Table 2).

Results for canonical variates for water body type (CVWBT) and the interaction between water body type and origin (CVWBTxOR). Symbols represent mean scores (± standard errors) of each water body type x origin group mean scores, with group means calculated from pooled individual fish within the group. Traits that correlated most heavily onto each axis are indicated (PREPELVL = prepelvic length, PREPECTL = prepectoral length, ACPD = anterior caudal peduncle depth, ADFAPF = anterior dorsal fin to anterior pelvic fin, VCPL = ventral caudal peduncle length, DCPL = dorsal caudal peduncle length). Red outlines represent the shape of a generic pumpkinseed; each truss connects to a homologous landmark on the body. Black deformation grids provide a visual representation of the shape differences between axes, and are overlaid across the generic shape
Unique responses of populations to waterbodies within geographic origins were also evident (interaction term; Table 2). Combining canonical axes describing shared and unique aspects of morphological divergence, the canonical axis derived from the interaction term described a morphological shift between water body types that was opposite in direction between native and non-native populations. Native populations exhibited shorter and deeper caudal peduncles and deeper bodies in lotic habitats, whereas non-native populations showed longer and slender caudal peduncles and more slender bodies in the same type of habitat (Fig. 3).
In our nested MANCOVA, we also found a moderate effect of populations nested within water body type x origin interaction (Table 2). This effect might be related to unique histories of populations.
Based on morphology, the first DFA classified 90% of the fish to the proper geographic origin (Wilks’s λ = 0.388, P < 0.0001). The second DFA correctly classified 73% of the fish to the proper water body type despite the marked differences in morphology among geographic origins (Wilks’s λ = 0.701, P < 0.0001). The third DFA, which incorporated both shared (contemporary selection and/or shared plastic developmental response among water body types) and unique histories of divergence related to geographic origins, correctly classified 67% of the fish into the proper geographic origin and water body type (Wilks’s λ = 0.717, P < 0.0001).
Discussion
Our analysis revealed that contemporary patterns of morphological variation among native and non-native pumpkinseed populations can be explained by contemporary selection and/or common plastic developmental response among water bodies, historical effects related to geographic origin and unique responses of populations to habitats within geographic origin, and that the effects of evolutionary history and the interaction between history and contemporary habitat were larger than contemporary processes in explaining morphological variation at this large spatial scale. Our results are consistent with previous studies, which found evidence that both contemporary and historical processes contribute to trait variation among species and populations (Taylor and McPhail 2000; Langerhans and DeWitt 2004; Langerhans et al. 2006; Berner et al. 2010) and, specifically for pumpkinseed populations, Weese et al. (2012) provided evidence that the effects of history and the interaction between history and contemporary habitat were larger than contemporary processes in structuring phenotypic variation across North American regions. Each of these effects is discussed below.
Our analysis detected strong effects of geographic origin on morphological variation among pumpkinseed populations (Table 2). But, the analysis of body size differences showed that the sampled pumpkinseed in North America were significantly larger than those sampled from the Iberian Peninsula, but there were no significant differences in body size between those taken from lentic and lotic water bodies nor between water bodies within geographic origin. These results suggest that the morphological differences between North American and Iberian pumpkinseed could be due, in part, to the larger size of fishes in the former, but that body size was not likely a factor in the lentic-lotic differences nor in North American and Iberian lentic-lotic differences found in our study.
Native populations exhibited morphological traits such as more posterior placement of paired fins, narrower caudal peduncles and body depths that were more streamlined relative to non-native populations (Table 3, Fig. 3); these particular patterns reflect characteristics that are well-suited to steady swimming (or cruising), fast-starts, and powered turns (sudden bursts of unsteady swimming activity, also termed accelerating) (Webb 1984a; Blake 2004). In contrast, non-native populations tended to exhibit more anteriorly positioned paired fins, deeper caudal peduncles and deeper bodies, which can facilitate low-speed manoeuvring in littoral areas (Webb 1984b), where prey are cryptic and foraging demands necessitate that individuals move through areas with complex structures (Jastrebski and Robinson 2004).
The development of a ‘cruising’ and ‘accelerating’ morphology in North American populations could be related to the stronger predation pressures experienced by pumpkinseed in that region, because predation is known to influence phenotypic development of several fishes (Webb 1986; Reznick et al. 1997; Johnson and Belk 2001; Langerhans and DeWitt 2004; Johansson and Andersson 2009), including pumpkinseed (Januszkiewicz and Robinson 2007). Pumpkinseed in the Iberian Peninsula experience predation from some obligate piscivores that are also not native to the Iberian Peninsula (e.g. Silurus glanis; Carol et al. 2009); these include species that were introduced from Europe and North America [i.e. Esox lucius Linnaeus and M. salmoides, respectively (Godinho et al. 1997)]. Despite the introduction of these piscivorous species, fish communities in the Iberian Peninsula remain dominated by cyprinids (Doadrio et al. 2011), which are easier for predators to physically consume than the pumpkinseed (Hoyle and Keast 1987). In fact, M. salmoides from the Iberian Peninsula are known to consume fewer pumpkinseed than other fishes, despite the high abundance of pumpkinseed in Iberian water bodies (Rodríguez-Jiménez 1989; García-Berthou 2002; see also Fox and Copp 2014). Pumpkinseed are generally more important prey to obligate piscivores in their native range than in Europe (Persson 2002), and the phenotypic differences we detected may be the result of higher predation risk in North America than in the Iberian Peninsula. If so, the development of manoeuvring traits in Iberian populations could be a direct result of reduced predation risk. However, previous studies have suggested that differences in morphological traits among native and non-native pumpkinseed populations may be related to habitat type and resource use (Vila-Gispert et al. 2007; Yavno et al. 2013), and to climatic differences including the degree of environmental variability (Naspleda et al. 2012; Yavno and Fox 2013). Furthermore, the observed differences between North American and European pumpkinseed could also have a genetic basis due to the influence of the particular source populations that originally became established in Spain, to population bottlenecks or genetic drift occurring in Iberian Peninsula following colonization. The particular reason for these differences cannot be determined without controlled experiments or more detailed population genetic studies than those that currently exist (i.e., Detta 2011).
Our study showed moderate effects of water body type that reflect shared responses across two distinct geographic regions, suggesting the presence of common selective forces across water bodies, which may have helped to produce convergent morphological characters. Alternatively, a shared developmental response which could have a genetic basis or not could also explain similar divergent responses within each geographic region by pumpkinseed from lentic and lotic water bodies (Fig. 3). The shared effects of water body type are evident despite intercontinental differences in the biotic and abiotic characteristics of the water bodies (e.g., climate, predation, competition, prey community). For example, the similar kinematics necessary to forage in littoral habitats, including the execution of sharp turns using short and deep caudal peduncles (Webb 1984a, 1984b; Blake 2004), may reflect the similar development of ‘manoeuvring’ traits exhibit by pumpkinseed from different geographic regions.
We expected that pumpkinseed from lotic waterbodies would exhibit characteristics that enhance swimming, regardless of their geographic origin, because natural selection would favour streamlined body forms in lotic water bodies. Our findings revealed that a proportion of morphological divergence, shared across geographically distant populations, pertained to the position of the paired fins, and body and caudal peduncle depth (Fig. 3). Some of these results conform to previous observations among pumpkinseed populations from lotic systems in their native range (more robust caudal peduncles; Brinsmead and Fox 2002), and also in part with observations made on lotic non-native populations (deeper bodies; Naspleda et al. 2012). While previous experimental work has demonstrated that native and non-native pumpkinseed are capable of developing streamlined body forms in response to water flow (Yavno and Fox 2013), the present study does not explicitly employ experimental work to support this finding. Therefore, we are unable to ascertain if selective pressures that pertain to swimming within these aquatic systems or alternatively, common plastic developmental responses have contributed to the shared morphological divergence. Future work could test the swimming performance of pumpkinseed from lotic and lentic water bodies, to identify whether water flow may represent one of the selective pressures which contributes to the shared morphological divergence.
Finally, we also found moderate effects of the interaction between geographic origin and water body type. These unique divergence patterns may suggest that particular body forms enhance fitness within their respective water body (Langerhans and DeWitt 2004). That is to say, there are probably alternative forms of achieving the hydrodynamic optimum for lotic water bodies. The unique axis of diversification (water body type x origin interaction, Fig. 3) revealed differences between native and non-native populations from lotic water bodies with respect to their body depth, and the length and depth of their caudal peduncle. The size/shape of body and caudal peduncle may align differently in each geographic region to achieve the phenotypic form that is optimal for that specific aquatic system. Divergent phenotypic responses have been identified in previous empirical studies. Yavno and Fox (2013) found that under flowing conditions, Iberian young-of-year pumpkinseed exhibit morphological changes to their caudal region that are the opposite of the changes observed in North American populations. Those findings suggest that morphological divergence related to flow may have a genetic basis, but as with the present study, this interpretation must be taken with caution because of the small number of populations tested.
Evolutionary historic differences between geographic regions influence how pumpkinseed respond to diversifying selection between water body types. In fact, differences in evolutionary histories could result in an evolutionary constraint, in which case any adaptive or developmental response would be biased by differences in the evolutionary history between geographic regions. Alternatively, unique divergence patterns might be reflective of events that occurred over the course of rapid contemporary evolution. These possibilities can be assessed by relating phenotypic differences among populations to genetic variation in mitochondrial DNA and microsatellite loci. On the other hand, Kaeuffer et al. (2012) suggested an ecological explanation for phenotypic divergence among populations which is related to the classification of environments into discrete types (in our study, lotic vs. lentic water bodies) and ignoring selective factors that differ among a given water body type. As they suggested, this explanation can be evaluated by relating trait differences among populations to quantitative differences in important ecological variables such as prey availability, diet, or the type and abundance of predators.
In conclusion, our analysis provides evidence that the patterns of morphological variation among native and non-native pumpkinseed populations can be explained by both shared (natural selection and/or common plastic developmental response among water bodies) and unique features of divergence (historical effects related to geographic origin and local responses of populations to habitats within geographic origin). We found that morphological divergence patterns shared across native and non-native populations are of lesser magnitude than those that are unique, suggesting that natural selection and/or common plastic developmental responses are weak in comparison to the relative roles of evolutionary history and local environmental pressures in explaining morphological divergence patterns among native and non-native pumpkinseed populations. These findings may contribute to increase our understanding about the nature and magnitude of the processes involved in the successful invasion and expansion of the pumpkinseed in the Iberian Peninsula and to highlight the evolutionary implications of invasions.
References
Berner D, Roesti M, Hendry AP, Salzburger W (2010) Constraints on speciation suggested by comparing lake-stream stickleback divergence across two continents. Mol Ecol 19:4963–4978
Bhagat Y, Fox MG, Ferreira MT (2006) Morphological differentiation in introduced pumpkinseed Lepomis gibbosus (L.) occupying different habitat zones in Portuguese reservoirs. J Fish Biol 69:79–94
Blake RW (2004) Fish functional design and swimming performance. J Fish Biol 65:1193–1222
Breden F, Scott A, Michel E (1987) Genetic differentiation for antipredator behaviour in the Trinidad guppy, Poecilia reticulata. Anim Behav 35:618–620
Brinsmead J, Fox MG (2002) Morphological variation between lake- and stream-dwelling rock bass and pumpkinseed populations. J Fish Biol 61:1619–1638
Carol J, Benejam L, Benito J, García-Berthou E (2009) Growth and diet of European catfish (Silurus glanis) in early and late invasion stages. Fundam Appl Limnol 174:317–328
Detta A (2011) The genetic origin and dispersal of introduced pumpkinseed (Lepomis gibbosus) in the Iberian peninsula. Trent University, Ontario, Canada, Master’s thesis
Doadrio I, Perea S, Garzón-Heydt P, González JL (2011) Ictiofauna continental española. Bases para su seguimiento. DG Medio Natural y Política Forestal, Madrid
Elvira B, Almodóvar A (2001) Freshwater fish introductions in Spain. Facts and figures at the beginning of the 21st century. J Fish Biol 59:323–331
Fox MG, Copp GH (2014) Old world versus new world: life-history alterations in a successful invader introduced across Europe. Oecologia 17:435–446
García-Berthou E (2002) Ontogenetic diet shifts and interrupted piscivory in introduced largemouth bass (Micropterus salmoides). Int Rev Hydrobiol 87:353–363
García-Berthou E, Moreno-Amich R (2000) Introduction of exotic fish into a Mediterranean lake over a 90-year period. Arch Hydrobiol 149:271–284
Gasith A, Resh VH (1999) Streams in Mediterranean climate regions: abiotic influences and biotic responses to predictable seasonal events. Annu Rev Ecol Syst 30:51–81
Gillespie GJ, Fox MG (2003) Morphological and life-history differentiation between littoral and pelagic forms of pumpkinseed. J Fish Biol 62:1099–1115
Godinho FN, Ferreira MT (1998) The relative influences of exotic species and environmental factors on an Iberian native fish community. Environ Biol Fish 51:41–51
Godinho FN, Ferreira MT, Cortes RV (1997) The environmental basis of diet variation in pumpkinseed sunfish, Lepomis gibbosus, and largemouth bass, Micropterus salmoides, along an Iberian river basin. Environ Biol Fish 50:105–115
Gould SJ, Woodruff DS (1990) History as a cause of area effects: an illustration from Cerion on great Inagua, Bahamas. Biol J Linn Soc 40:67–98
Hoyle JA, Keast A (1987) Effect of prey morphology and size on handling time in a piscivore, the largemouth bass (Micropterus salmoides). Can J Zool 65:1972–1977
Januszkiewicz AJ, Robinson BW (2007) Divergent walleye (Sander vitreus)-mediated inducible defences in the centrarchid pumpkinseed sunfish (Lepomis gibbosus). Biological Journal of the Linnaean Society 90:25–36
Jastrebski CJ, Robinson BW (2004) Natural selection and the evolution of replicated trophic polymorphisms in pumpkinseed sunfish (Lepomis gibbosus). Evol Ecol Res 6:285–305
Johansson F, Andersson J (2009) Scared fish get lazy, and lazy fish get fat. J Anim Ecol 78:772–777
Johnson JB (2001) Adaptive life-history evolution in the livebearing fish Brachyrhaphis rhabdophora: genetic basis for parallel divergence in age and size at maturity and a test of predator-induced plasticity. Evolution 55:1486–1491
Johnson JB, Belk MC (2001) Predation environment predicts divergent life-history phenotypes among populations of the livebearing fish Brachyrhaphis rhabdophora. Oecologia 126:142–149
Kaeuffer R, Peichel CL, Bolnick DI, Hendry AP (2012) Parallel and nonparallel aspects of ecological, phenotypic, and genetic divergence across replicate population pairs of lake and stream stickleback. Evolution 66:402–418
Klingenberg CP (2011) MorphoJ: an integrated software package for geometric morphometrics. Mol Ecol Resour 11:353–357
Künstler J (1908) Amiurus nebulosus et Eupomotis gibbosus. Bulletin de la Société d'Acclimation 1908:238–244
Langerhans RB (2008) Predictability of phenotypic differentiation across flow regimes in fishes. Integr Comp Biol 48:4265–4271
Langerhans RB (2009) Trade-off between steady and unsteady swimming underlies predator-driven divergence in Gambusia affinis. J Evol Biol 22:1057–1075
Langerhans RB, DeWitt TJ (2004) Shared and unique features of evolutionary diversification. Am Nat 164:335–349
Langerhans RB, Knouft JH, Losos JB (2006) Shared and unique features of diversification in greater Antillean Anolis ectomorphs. Evolution 60:362–369
Losos JB, Jackman TR, Larson A, de Queiroz K, Rodríguez-Schettino L (1998) Contingency and determinism in replicated adaptive radiations of island lizards. Science 279:2115–2118
Mittelbach GC, Osenberg CW, Wainwright PC (1999) Variation in feeding morphology between pumpkinseed populations: phenotypic plasticity or evolution? Evol Ecol Res 1:111–128
Naspleda J, Vila-Gispert A, Fox MG, Zamora L, Ruiz-Navarro A (2012) Morphological variation between lake-and stream dwelling pumpkinseed Lepomis gibbosus in the Iberian peninsula. J Fish Biol 81:1915–1935
Persson L (2002) Community Ecology of Freshwater Fishes. In: Hart JB and Reynolds P (eds) Handbook of Fish Biology and Fisheries, Volume 1: Fish Biology. Blackwell Publishing Ltd, Oxford
Price T, Lovette IJ, Bermingham E, Gibbs HL, Richman AD (2000) The imprint of history on communities of north American and Asian warblers. Am Nat 156:354–367
Reznick DN (1982) The impact of predation on life-history evolution in Trinidadian guppies: genetic basis of observed life history patterns. Evolution 36:1236–1250
Reznick DN, Shaw FH, Rodd FH, Shaw RG (1997) Evaluation of the rate of evolution in natural populations of guppies (Poecilia reticulata). Science 275:1934–1937
Riopel C, Robinson BW, Parsons KJ (2008) Analysing nested variation in the body form of Lepomid sunfishes. Environ Biol Fish 82:409–420
Robinson BW, Wilson DS (1994) Character release and displacement in fishes: a neglected literature. Am Nat 144:596–627
Robinson BW, Wilson DS (1996) Genetic variation and phenotypic plasticity in a trophically polymorphic population of pumpkinseed sunfish (Lepomis gibbosus). Evol Ecol 10:631–652
Robinson BW, Wilson DS, Margosian AS, Lotito PT (1993) Ecological and morphological-differentiation of pumpkinseed sunfish in lakes without bluegill sunfish. Evol Ecol 7:451–464
Robinson BW, Wilson DS, Shea GO (1996) Trade-offs of ecological specialization: an intraspecific comparison of pumpkinseed sunfish phenotypes. Ecology 77:170–178
Robinson BW, Wilson DS, Margosian AS (2000) A pluralistic analysis of character release in pumpkinseed sunfish (Lepomis gibbosus). Ecology 81:2799–2812
Rodríguez-Jiménez AJ (1989) Feeding habits of Micropterus salmoides (Pisces: Centrarchidae), Lepomis gibbosus (Pisces: Centrarchidae) and Gambusia affinis (Pisces: Poeciliidae) in the Proserpina reservoir banks (Extremadura, Spain). Limnética 5:13–20
Rundell RJ, Price TD (2009) Adaptive radiation, non-adaptive radiation, ecological speciation and non-ecological speciation. Trends Ecol Evol 24:394–399
Schluter D (1996a) Ecological causes of adaptive radiation. Am Nat 148:S40–S64
Schluter D (1996b) Ecological speciation in postglacial fishes. Philos Trans R Soc Lond B 351:807–814
Sharpe DMT, Räsänen K, Berner D, Hendry AP (2008) Genetic and environmental contributions to the morphology of lake and stream stickleback: implications for gene flow and reproductive isolation. Evol Ecol Res 10:849–866
Smith TB, Skúlason S (1996) Evolutionary significance of resource polymorphisms in fishes, amphibians, and birds. Annu Rev Ecol Syst 27:111–133
Tabachnick BG, Fidell LS (2001) Computer-assisted research design and analysis. Allyn and Bacon, Boston
Taillon D, Fox MG (2004) The influence of residential and cottage development on littoral zone fish communities in a mesotrophic north temperate lake. Environ Biol Fish 71:275–285
Taylor EB, McPhail JD (2000) Historical contingency and ecological determinism interact to prime speciation in sticklebacks, Gasterosteus. Proc R Soc Lond B 267:2375–2384
Vila-Gispert A, Fox MG, Zamora L, Moreno-Amich R (2007) Morphological variation in pumpkinseed Lepomis gibbosus introduced into Iberian lakes and reservoirs; adaptations to habitat type and diet? J Fish Biol 71:163–181
Wainwright PC, Lauder GV (1992) The evolution of feeding biology in the sunfishes (Centrarchidae). In: Mayden RL (ed) Systematics, historical ecology, and north American freshwater fishes. Stanford University Press, Stanford CA, pp 472–491
Webb PW (1984a) Body form, locomotion and foraging in aquatic vertebrates. Am Zool 24:107–120
Webb PW (1984b) Form and function in fish swimming. Sci Am 25:72–82
Webb PW (1986) Effect of body form and response threshold on the vulnerability of four species of teleost prey attacked by largemouth bass (Micropterus salmoides). Can J Fish Aquat Sci 43:763–771
Weese DJ, Ferguson MM, Robinson BW (2012) Contemporary and historical evolutionary processes interact to shape patterns of within-lake phenotypic divergences in polyphenic pumpkinseed sunfish, Lepomis gibbosus. Ecol Evol 2:574–592
Yavno S, Fox MG (2013) Morphological change and phenotypic plasticity in native and non-native pumpkinseed sunfish in response to sustained water velocities. J Evol Ecol 26:2383–2395
Yavno S, Fox MG, Vila-Gispert A, Bhagat Y (2013) Morphological differences between native and non-native pumpkinseed in traits associated with locomotion. Environ Biol Fish 96:507–518
Yoder JB, Clancey E, Des Roches S, Eastman JM, Gentry L, Godsoe W, Hagey TJ, Jochimsen D, Oswald BP, Robertson J, Sarver BAJ, Schenk JJ, Spear SF, Harmon LJ (2010) Ecological opportunity and the origin of adaptive radiations. J Evol Ecol 23:1581–1596
Acknowledgements
Funding was provided by a PhD Grant from the University of Girona to JN, and Scientific Research Grants (CGL2006-01755/BOS, CGL2010-15958/BOS) from the Spanish Ministry of Science and Technology to AVG, and by the Ontario Ministry of Training, Colleges and Universities in the form of an Ontario Graduate Scholarship to SY. Financial support was also provided by the Generalitat de Catalunya (ref. 2014 SGR 484). The authors would like to thank Lluis Benejam, Quim Carol, Jordi René-Mor and Yakuta Bhagat for their field and laboratory assistance.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed.
Rights and permissions
About this article

Cite this article
Vila-Gispert, A., Yavno, S., Naspleda, J. et al. Patterns of morphological variation among native and non-native pumpkinseed (Lepomis gibbosus) populations: shared and unique aspects of diversification. Environ Biol Fish 100, 969–980 (2017). https://doi.org/10.1007/s10641-017-0626-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-017-0626-x