
Abstract
In marine ecosystems, cleaning is a mutualistic relationship in which so-called cleaners remove ectoparasites, diseased tissue, or mucus from the body of their clients, and thus help to maintain a healthy reef community. In spite of its importance in many marine habitats, this interaction remains poorly understood, particularly at oceanic islands. Here, we present the first comprehensive study of cleaning interactions in a reef fish assemblage at Rocas, the only atoll in the South Atlantic. We recorded 318 cleaning events, in which six fish species, including two endemic ones, and two shrimp species acted as cleaners. The clients serviced by these cleaners were 21 bony fish species, one shark and one sea turtle. The cleaner wrasse Thalassoma noronhanum and the cleaner goby Elacatinus phthirophagus were the cleaners with the greatest number of events and species richness of clients. Additionally, 82% of clients in the cleaning events were non-piscivores, and the abundance of both cleaners and clients positively influenced the number of cleaning events (R2 = 0.4; p < 0.001). Our results indicate that Rocas atoll has a high species richness of cleaner species despite its small size and highlight the importance of studies of cleaning symbiosis, even in isolated places with low species richness, for a better comprehension of this association in reefs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cleaning symbiosis is considered as a key interaction within animal populations, since in these interactions cleaners remove ectoparasites and diseased tissue from their clients, and thus help to maintain healthy communities (Losey 1972; Losey et al. 1999). In the marine environment, these interactions usually take place at stationary reef locations, known as cleaning stations; normally massive coral colonies, sponges and large rocks (Slobodkin and Fishelson 1974). At least 208 fish species and 51 shrimp species have been recorded acting as cleaners (Vaughan et al. 2016), which are classified in two groups (facultative and dedicated) according to the life stage in which cleaning behavior is displayed and the degree of dependence on these interactions (Côté 2000; Grutter 2005; Vaughan et al. 2016). Facultative cleaners only clean at juvenile stages or sporadically as a complementary food resource, whereas dedicated cleaners are highly specialized species that clean during their entire lifetime (Grutter 2005; Vaughan et al. 2016).
Mutualistic relationships between cleaners and clients are influenced by multiple factors. First, species abundance determines the rate of the encounters between cleaners and clients (Floeter et al. 2007). Second, parasite load influences the frequency clients search for cleaning services (Grutter 2002). Third, cleaner preferences for some clients are influenced by greater parasite loads of the latter (Darcy et al. 1974). Fourth, a series of morphological traits and specific behaviors, such as contrasting and vivid colors, degree of jaw protrusion, and fast jaw movements for prey capture (Losey 1987; Losey et al. 1999; Baliga and Mehta 2016). Fifth, tendency of clients to school at cleaning stations that foster cleaning interactions (Cheney et al. 2009; Quimbayo et al. 2016). Finally, biogeographic and evolutionary processes that probably limited the presence of particular cleaners species in some regions (Quimbayo et al. 2012).
Given these factors, questions about how cleaning interactions occur and what ecological factors affect these interactions remain open to debate. In this sense, the identification of cleaners and clients, as well as quantification of matches between them, are the first step to understand the drivers of cleaning interactions over the broad spectrum of natural conditions and evolutionary histories encompassed by distinct biogeographic regions (Floeter et al. 2007). Despite the known geographical variation on cleaning interactions features, most studies have been conducted mainly in coastal reefs of Indo-Pacific and Caribbean (e.g., Johnson and Ruben 1988; Poulin and Grutter 1996; reviews in Côté 2000 and Vaughan et al. 2016). Only recently these associations have been investigated in other regions, such as the southwestern Atlantic (SWA) and the eastern Atlantic (e.g. Sazima et al. 1998, 2000; Francini-Filho and Sazima 2007; Quimbayo et al. 2012; Morais et al. 2016). Still, studies on cleaning symbiosis in the SWA are mostly restricted to case studies focused on one or two cleaner species (e.g., Sazima et al. 1999; Francini-Filho and Sazima 2007; Krajewski 2007; Coni et al. 2010). Only one comprehensive study has assessed the network structure of an assemblage of cleaners and clients in this region (Sazima et al. 2010b). Additionally, about a half of the studies on cleaning interactions in the SWA were done in coastal reefs and reefs in the continental shelf (Sazima et al. 1998, 1999, 2000; Sazima and Sazima 2001). Consequently, oceanic islands in the SWA remain poorly understood, as the studies on this subject focused on the oceanic Fernando de Noronha Archipelago (e.g., Sazima and Moura 2000; Sazima et al. 2004, 2005, 2010a; Francini-Filho and Sazima 2007). A comprehensive assessment of cleaning interactions in oceanic islands is key for a better understating of reef fish ecology in the SWA, given that the these ecosystems harbor about 70% of all cleaner species reported for the Brazilian Province (Côté 2000; Floeter et al. 2008; Vaughan et al. 2016).
Here, we provide the first comprehensive study of cleaning interactions at the Rocas atoll, an oceanic island in the SWA. More specifically, this study aims to answer: (1) What species do take part in cleaning interactions at the Rocas atoll? (2) How do cleaning interactions vary among cleaners (i.e. number of cleaning events, number of serviced clients, cleaners servicing or not at cleaning stations)? (3) What are the species and trophic group of clients? (4) How morphologic traits of clients influence cleaning interactions? (5) Are cleaning interactions influenced by the abundance of cleaners and clients? Collectively, the answers to these questions will provide a glimpse of this complex interaction at an isolated reef site, and identify some resemblances and dissimilarities of cleaning interactions among oceanic islands and between oceanic islands and the continental areas in the SWA. Additionally, this study provides a template for future studies on cleaning interactions and, therefore, will allow for comparisons between Rocas and other locations in which cleaners-clients assemblages have been comprehensively studied (see Floeter et al. 2007 for a macroecological perspective).
Materials and methods
Study site
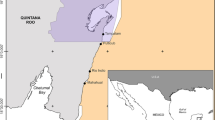
Cleaning interactions were recorded at Rocas atoll (03°50′S, 33°49′W) located 230 km off the northeastern coast of Brazil. This is the only atoll in the SWA, and is one of the smallest in the world (Longo et al. 2015), with a total area of 5.5 km2 (Fig. 1). Rocas is mainly composed of a coralline algal-vermetid gastropod association and is part of a seamount chain known as the Fracture Zone of Fernando de Noronha (Kikuchi and Leão 1997). At this atoll, two sandy cays lie on the western side of the reef flat and several pools of various sizes and depths are found within the atoll. Some of these pools remain connected with the open ocean (open pools) during low tide, whereas others do not (closed pools). The tidal regime that affects the atoll is semi-diurnal and mesotidal, with maximum amplitudes of 3.8 m (Gherardi and Bosence 2001). During our study, water visibility varied between 2 and 10 m and the temperature ranged from 28 to 31 °C.

The Rocas atoll, about 250 km off the Brazilian coast (southwestern Atlantic), and the studied sites within it, including the species recorded cleaning in each site. Sandy cays are depicted in yellow, open pools in light blue and closed pools in dark blue. Symbols indicate the species of cleaners at each study site
Observational sessions were conducted during low tide in four closed and three open pools with perimeters between 0.26–2.64 km and depth ranging 1–5 m. We did a total of 184 direct observations, 87 photographic records and 47 remote video records of cleaning interactions. These observations were distributed over sessions of two hours daily along 22 days, totaling 44 h of observations in May 2016. We considered that a cleaning interaction began when we detected a physical contact between a cleaner and a client, and ended when the cleaner or the client withdrew (Johnson and Ruben 1988; Sazima et al. 2000). During each event, we identified the species and visually estimated the body size (total length, TL) of the client and the cleaner. The abundance of species was estimated with underwater visual censuses (UVCs; N = 90) along 40 m2 (20 × 2 m) transects (Krajewski and Floeter 2011; Pinheiro et al. 2011; Longo et al. 2015).
Additionally, we classified each client fish species according to its diet (herbivore-detritivore, mobile invertebrate feeder, omnivore, planktivore and piscivore), level in the water column (benthic, bentho-pelagic and pelagic), and social organization (solitary or gregarious) as defined by Mouillot et al. (2014). Shark and the sea turtles were also classified according to diet, level in the water column, and social organization, following Rincon-Diaz et al. (2011), Bell (2013) and Froese and Pauly (2016).
Data analyses
Quantitative networks of ecological interactions were used to assess the number of cleaning events and species richness of clients. For the network, we used the number of cleaning interactions between each cleaner species and its client species. We ran these analyses with the function “plotweb” within the package “bipartite” (Dormann et al. 2008). To examine how functional traits of clients affected the probability of them to be cleaned, we used a generalized linear model (GLM) with binomial distribution. Number of cleaning events per client species relative to total abundance of each client in the UVCs was used as response variable. The functional traits examined in this analysis were the same referred above: dietary type, level in water column and social organization. To evaluate the relationship between the proportion of cleaning events and the relative abundance of clients and cleaners, we used a linear regression with 95% confidential intervals over model predictions. Species falling above or below this interval were considered, respectively, as interacting disproportionally more and less than expected given their abundance. All the analyses were performed in R software version 3.2.4 (R Core Team 2016).
Results
We recorded 318 cleaning interactions, in which six fish species from four families and two shrimp species from two families acted as cleaners (Fig. 2). Fishes cleaned in 94% of the events (Table S1). The endemic wrasse Thalassoma noronhanum (Labridae) cleaned in 75.5% of the events and had the richest assemblage of clients (18 species), followed by the endemic goby Elacatinus phthirophagus (Gobiidae, 12.6%), the angelfish Pomacanthus paru (Pomacanthidae, 2.3%), the damselfish Stegastes rocasensis (Pomacentridae, 2.3%), the surgeonfish Acanthurus chirurgus (Acanthuridae, <1%), and the damselfish Abufdefduf saxatilis (Pomacentridae, <1%; Figs. 2 and 3).

The eight cleaner species found at the Rocas atoll: a the endemic wrasse Thalassoma noronhanum, b the endemic goby Elacatinus phthirophagus, c the angelfish Pomacanthus paru, d the damselfish Stegastes rocasensis, e the surgeonfish Acanthurus chirurgus, f the damselfish Abufdefduf saxatilis, and the shrimps g Lysmata grabhami and h Stenopus hispidus. From these, only the goby E. phthirophagus is a dedicated cleaner, whereas the remainder species are facultative cleaners

Network of cleaning interactions at the Rocas atoll. Circles are proportional to relative abundance of fish species (abundance of shrimps was not quantified). Lines indicate cleaner–client interaction; line width is proportional to the number of cleaning events (Table S1)
Apart from cleaning by E. phthirophagus and T. noronhanum, the remaining cleaning services were mostly performed by juvenile fishes (Fig. 4 A-C, F). Cleaning by shrimps were rarer than by fishes, as Lysmata grabhami (Hippolytidae) and Stenopus hispidus (Stenopodidae) cleaned, respectively, in 3.7% and 2.3% of the events (Figs. 2- 3). Thalassoma noronhanum and E. phthirophagus cleaned on stations based on Montastraea cavernosa corals or anthozoan colonies (Fig. 4), whereas the shrimps L. grabhami and S. hispidus cleaned exclusively in rock crevices and serviced mainly clients with nocturnal habits, such as the squirrelfish Holocentrus adscensionis, the soldierfish Myripristis jacobus, and the cardinalfish Apogon americanus (Table S1; Fig. 4).

Some examples of cleaning interactions at the Rocas atoll; a a juvenile Thalassoma noronhanum approaches a posing Acanthurus chirurgus at mid water, b an adult T. noronhanum approaches a group of the parrotfish Sparisoma axillare waiting to be serviced at a cleaning station, c an adult Elacatinus phthirophagus cleans the back of the damselfish Chromis multilineata at a cleaning station, d Lysmata grabhami cleans the gills of Holocentrus adscensionis, e a juvenile Pomacanthus paru cleans a posing Abufdefduf saxatilis, f a juvenile Stegastes rocasensis inspects the carapace of a resting hawksbill turtle Eretmochelys imbricata
We recorded 23 client species from 15 families (21 ray-finned fishes, a shark and a sea turtle), from which 15 are diurnal (65.2%), two nocturnal (8.7%), and six active at day and night (Table S1). The clients more frequently recorded in the cleaning events were the surgeonfish Acanthurus chirurgus (41%) and the squirrelfish Holocentrus adscensionis (22%). In contrast, frequency of other client species was low, with each of them representing 1–7% of cleaning events (Fig. 3). The only shark recorded in this study, Ginglymostoma cirratum, was cleaned by T. noronhanum, E. phthirophagus and S. rocasensis, whereas the hawksbill turtle, Eretmochelys imbricata, was cleaned by A. saxatilis and A. chirurgus (Fig. 4 A-C, F), the latter species observed cleaning once or twice.
The majority of the clients, considering the total number of events recorded, were herbivore-detritivores (30.4%), followed by mobile invertebrate feeders (26%), piscivores (17.4%), planktivores and omnivores (13% each). Client species preferentially cleaned were those in groups, at higher levels in the water column and in two trophic groups: herbivore-detritivore and omnivore (GLM: p < 0.05; Table 1).
The proportion of cleaning events of a given species was positively related to its abundance in the reef (R2 = 0.4; p < 0.001; Fig. 5). However, some of the most representative cleaners, especially E. phthirophagus and T. noronhanum, cleaned disproportionately more than expected from their abundances (Fig. 5). In contrast, A. saxatilis and S. rocasensis cleaned disproportionately less than expected by their abundances. Among the clients, the parrotfishes Sparisoma amplum and Sparisoma frondosum were cleaned more and the damselfish Stegastes rocasensis less than expected by their relative abundances on the reef (Fig. 3). The client assemblages serviced by the wrasse T. noronhanum and the goby E. phthirophagus showed some differences. For instance, the wrasse attended mainly non-piscivorous clients that arrived in large groups and remained in the water column, whereas clients of the goby included more piscivores that remained solitary and were associated with the bottom.

Linear regression model between the relative abundance of fish cleaners and clients and proportion of cleaning events at Rocas atoll. Blue points, and associated fish figures, represent cleaner fish individuals, and black squares represent client individuals. Cleaners: Thalassoma noronhanum (Tha nor), Elacatinus phthirophagus (Ela pht), Stegastes rocasensis (Ste roc), and Abufdefduf saxatilis (Abu sax). Line represents the linear model and shaded area represents the confidence intervals of 95% generated based on standard errors of model’s predictions
Discussion
Our study revealed a particularly rich assemblage of cleaners at Rocas, despite the small area and general low species richness of the atoll (Floeter et al. 2008). Within the southwestern Atlantic, the species richness of cleaners at Rocas may be considered high and comparable to the richness of cleaners at the much larger Fernando de Noronha Archipelago (e.g., Francini-Filho and Sazima 2007; Sazima et al. 2010a; Sazima et al. 2013). A likely explanation for the cleaner richness at the atoll is its biogeographic connection to Fernando de Noronha Archipelago through a seamount chain, which would result in several shared marine species between these locations (Floeter et al. 2008; Hachich et al. 2015).
We also found some novelties among the cleaning interactions at Rocas atoll. For instance, Thalassoma noronhanum acted as a cleaner and maintained cleaning stations as terminal phase adult (Fig. 4B), a previously unrecorded behavior for this species, even when considering a previous study at Rocas (Campos et al. 2007; Francini-Filho and Sazima 2007). This behavior is likely related to the versatile foraging of this wrasse (Sazima et al. 2005), as well as the maintenance of some ontogenetic traits at the adult stage, such as the flexibility across the vertebral column, body depth, high protrusion and fast jaw movements for prey capture (Baliga and Mehta 2015, 2016). Another singularity found was the aggregation of several parrotfish individuals at cleaning stations of the Noronha wrasse (Fig. 4B). Although clients’ aggregation at cleaning stations is habitual in the presence of active cleaners and some clients even wait at temporarily vacant stations (Sazima et al. 1999), the behavior and postures adopted by the parrotfishes at one of the cleaning stations was unique. We also recorded the damselfish S. rocasensis cleaning fishes including the nurse shark Ginglymostoma cirratum, a behavior previously unreported, as this fish was known to clean sea turtles only (Sazima et al. 2010a).
The dominance of the wrasse Thalassoma noronhanum and the goby Elacatinus phthirophagus we found in this study was previously recorded for the Fernando de Noronha Archipelago (Francini-Filho and Sazima 2007). However, at the latter site T. noronhanum keeps cleaning stations at midwater only (Francini-Filho and Sazima 2007), whereas at Rocas this species kept substrate-based stations. The differences among the client assemblage serviced by the wrasse and the goby agree with Francini-Filho et al. (2000), who found that Thalassoma species avoid interactions with potentially dangerous species, whereas cleaner gobies have no differences in the trophic group of clients at their cleaning stations (Côté and Soares 2011). Our results about preferentially cleaned clients differ from several studies at some other West Atlantic localities, where planktivores are among the most serviced clients (Johnson and Ruben 1988; Wicksten 1995, 1998; Arnal et al. 2000; Sazima et al. 2000). The reasons for this difference may be due to the relatively low abundance of planktivores, besides topography and water flow conditions within the atoll, among other possible variables.
At Rocas atoll, the angelfish Pomacanthus paru rarely acted as a cleaner and, when doing so, serviced only a few clients. In contrast, this species is an active cleaner with a high species richness of clients at other localities in the western Atlantic, such as Florida in the USA (Brockmann and Hailman 1976), and the Abrolhos Archipelago in Brazil (Sazima et al. 1999). However, busy cleaning activity and a varied assemblage of clients of this angelfish were recorded at Fernando de Noronha Archipelago (Sazima et al. 2013), but only at a few sites where the wrasse T. noronhanum was uncommon. This supports the idea of high cleaning efficiency of T. noronhanum where this species is abundant (Francini-Filho and Sazima 2007; present paper).
In our study, the hawksbill turtle was cleaned mostly by juvenile fishes, a trend recorded for sea turtles at other locations including Fernando de Noronha Archipelago (Sazima et al. 2010a). Cleaners of turtles are usually herbivorous, mobile invertebrate feeders and omnivorous reef fishes that likely supplement their diets with small invertebrates and/or algal epibionts that dwell on the hard and soft parts of sea turtles (Ferreira et al. 2004; Sazima et al. 2010a). However, at the continental Abrolhos Archipelago, the cleaner goby Elacatinus figaro was recorded nipping at the carapace of a hawksbill turtle (Carneiro-Proietti et al. 2012), which would be unexpected given the specialized cleaning habits of this goby (Sazima et al. 2000; Sazima et al. 2010b). Clearly, the relationship between cleaner fishes and sea turtles still deserves a close attention of marine biologists (Sazima et al. 2010a).
The low frequency of cleaning events by shrimps in the present study is unexpected, given that shrimp species are important cleaners at other locations in the western Atlantic, such as St. Croix (Johnson and Ruben 1988) and Bay Islands (Titus et al. 2015), both in the Caribbean. The low activity of cleaner shrimps at Rocas may be related to the high efficiency and abundance of fish cleaners (especially T. noronhanum), or by the fact that shrimps clean mostly in rock crevices (Sazima et al. 2004) or at night (Bonaldo et al. 2015). In fact, all of our sampling were conducted during the day and, therefore, would not adequately represent nocturnal cleaning activity.
We believe that three main factors may explain the positive relationship between the abundance of species and the number of cleaning interactions in our study. First, cleaners could interact more frequently with abundant clients than with rare ones, resulting in a positive relationship between client abundance and number of cleaning interactions (Floeter et al. 2007). Second, more abundant cleaners would attracted more clients, which would seek cleaning stations attended by a larger number of cleaners since clients could be serviced by several cleaners at the same time (Slobodkin and Fishelson 1974; Grutter 1997). Third, gregarious clients could interact with cleaners more often than solitary clients, both because they would more easily find patchily distributed cleaning stations and because they might have greater parasite loads (Pitcher and Parrish 1993; Sasal 2003).
In conclusion, our study is the first comprehensive characterization of cleaning symbiosis at an oceanic island in the SWA and other areas as well. We recorded some novelties at Rocas atoll and several differences and similarities between our results and those of studies at other SWA oceanic islands and the mainland coast. Moreover, our results reveal that the knowledge of cleaning interactions in the SWA is still in its infancy. Further studies at other poorly known oceanic islands, such as Trindade Island and St. Peter and St. Paul’s. Archipelago, are needed to properly assess the nature of cleaning interactions at isolated sites. The same applies for coastal sites in Brazil, where these associations still remain largely underexplored. Our study provides a template for future studies on cleaning interactions and will allow comparisons between Rocas and other locations where the cleaners-clients assemblages have been comprehensively studied, which, in turn, would allow all-inclusive analyses such as those in Floeter et al. (2007) or network analyses in Sazima et al. (2010b).
References
Arnal C, Côté IM, Sasal P, Morand S (2000) Cleaner-client interactions on a Caribbean reef: influences of correlates of parasitism. Behav Ecol Sociobiol 47:353–358. doi:10.1007/s002650050676
Baliga VB, Mehta RS (2016) Ontogenetic allometry in shape and flexibility underlies life history patterns of labrid cleaning behavior. Integr Comp Biol. doi: 10.1093/icb/icw028
Baliga VB, Mehta RS (2015) Linking cranial morphology to prey capture kinematics in three cleaner wrasses: Labroides dimidiatus, Larabicus quadrilineatus, and Thalassoma lutescens. J Morphol 276:1377–1391. doi:10.1002/jmor.20425
Bell I (2013) Algivory in hawksbill turtles: Eretmochelys imbricata food selection within a foraging area on the northern great barrier reef. Mar Ecol 34:43–55. doi:10.1111/j.1439-0485.2012.00522.x
Bonaldo RM, Grutter AS, Sazima I, Krajewski JP (2015) 24/7 service: nocturnal cleaning in a tropical indo-Pacific reef. Mar Biodivers. doi:10.1007/s12526-014-0289-1
Brockmann HJ, Hailman JP (1976) Fish cleaning symbiosis: notes on juvenile angelfishes (Pomacanthus, Chaetodontidae) and comparisons with other species. Z Tierpsychol 42:129–138
Campos CEC, Silva MB, Targino SG et al (2007) Simbiose de limpeza de Thalassoma noronhanum (Boulenger, 1890) (Labridae) na Reserva Biológica do Atol das Rocas, Rio Grande do Norte, Brasil. Rev Etol 8:63–70
Carneiro-Proietti M, Reisser J, Secchi ER (2012) Foraging by immature hawksbill sea turtles at Brazilian islands. Mar Turt Newsl 135:4–8
Cheney KL, Grutter AS, Blomberg SP, Marshall NJ (2009) Blue and yellow signal cleaning behavior in coral reef fishes. Curr Biol 19:1283–1287. doi:10.1016/j.cub.2009.06.028
Coni EOC, Nunes JACC, Ferreira CM et al (2010) The Spanish hogfish Bodianus rufus (Labridae) acting as cleaner of nocturnal fish in the north-east of Brazil. Mar Biodivers Rec 3:1–2. doi:10.1017/S1755267210000187
Côté IM (2000) Evolution and ecology of cleaning symbiosis in the sea. Oceanogr Mar Biol 38:311–355
Côté IM, Soares MC (2011) Gobies as cleaners. In: Van Tassell JL, Kovacíc M, Kapoor BG (eds) Patzner RA. Biol. Gobies. CRC Press, Boca Raton, pp 525–551
Darcy GH, Maisel E, Ogden JC (1974) Cleaning preferences of the gobies Gobiosoma evelynae and G. prochilos and the juvenile wrasse Thalassoma bifasciatum. Copeia 1974:375–379
Dormann CF, Fruend J, Bluethgen N, Gruber B (2008) Introducing the bipartite package: Analysing ecological networks. R news 8(2):8–11
Ferreira CEL, Floeter SR, Gasparini JL et al (2004) Trophic structure patterns of Brazilian reef fishes: a latitudinal comparison. J Biogeogr 31:1093–1106. doi:10.1111/j.1365-2699.2004.01044.x
Floeter SR, Rocha LA, Robertson DR et al (2008) Atlantic reef fish biogeography and evolution. J Biogeogr 35:22–47. doi:10.1111/j.1365-2699.2007.01790.x
Floeter SR, Vázquez DP, Grutter AS (2007) The macroecology of marine cleaning mutualisms. J Anim Ecol 76:105–111. doi:10.1111/j.1365-2656.2006.01178
Francini-Filho RB, Sazima I (2007) A comparative study of cleaning activity of two reef fishes at Fernando de Noronha Archipelago, tropical West Atlantic. Environ Biol Fish 83:213–220. doi:10.1007/s10641-007-9322-6
Francini-Filho R, Moura RL, Sazima I (2000) Cleaning by the wrasse Thalassoma noronhanum, with two records of predation by its grouper client Cephalopholis fulva. J Fish Biol 56 (4):802–809. doi:10.1006/jfbi.1999.1197
Froese R, Pauly D (2016) FishBase. World wide web electronic publication. Available: www.fishbase.org. Accessed 2016 Jun 13
Gherardi DFM, Bosence DWJ (2001) Composition and community structure of the coralline algal reefs from Atol das Rocas, South Atlantic, Brazil. Coral Reefs 19:205–219. doi:10.1007/s003380000100
Grutter AS (2002) Cleaning symbioses from the parasites’ perspective. Parasitology 124:s61–s81. doi:10.1017/S0031182002001488
Grutter AS (1997) Effect of the removal of cleaner fish on the abundance and species composition of reef fish. J Fish Biol 50:1303–1308. doi:10.1007/s004420050217
Grutter AS (2005) Cleaning mutualism in the sea. In: Rohde K (ed) mar. Parasitol. CSIRO Publishing, Collingwood, pp 264–278
Hachich NF, Bonsall MB, Arraut EM et al (2015) Island biogeography: patterns of marine shallow-water organisms in the Atlantic Ocean. J Biogeogr 45:1871–1882. doi:10.1111/jbi.12560
Johnson WS, Ruben P (1988) Cleaning behavior of Bodianus rufus, Thalassoma bifasciatum, Gobiosoma evelynae, and Periclimenes pedersoni along a depth gradient at Salt River submarine canyon, St. Croix. Environ Biol Fish 23:225–232. doi:10.1007/BF00004913
Kikuchi RKP, Leão ZMAN (1997) Rocas (southwestern equatorial Atlantic, Brazil): an atoll built primarily by coralline algae. Proc 8 Th Int coral reef Sym 1:731–736
Krajewski JP, Floeter SR (2011) Reef fish community structure of the Fernando de Noronha Archipelago (equatorial western Atlantic): the influence of exposure and benthic composition. Environ Biol Fish 92:25–40. doi:10.1007/s10641-011-9813-3
Krajewski JP (2007) Cleaning by the occasional cleaner Diplodus argenteus (Perciformes: Sparidae) in south Brazil: why so few client species? J Mar Biol Assoc UK 87:1013. doi:10.1017/S0025315407055981
Longo GO, Morais RA, Martins CDL et al (2015) Between-habitat variation of benthic cover, reef fish assemblage and feeding pressure on the benthos at the only atoll in south Atlantic: Rocas atoll, NE Brazil. PLoS One 10:e0127176. doi:10.1371/journal.pone.0127176
Losey GS (1972) The ecological importance of cleaning symbiosis. Copeia 4:960–970
Losey GS (1987) Cleaning symbiosis. Symbiosis 4:229–258
Losey GS, Grutter AS, Rosenquist G et al (1999) Cleaning symbiosis: a review. In: Oliveira RF, Goncalves EJ (eds) Almada VC. Behav. Conserv. littoral fishes. Lisboa, Portugal, pp 379–395
Morais RA, Brown J, Ferreira CEL et al (2016) Mob rulers and part-time cleaners: two reef fish associations at the isolated Ascension Island. J mar Biol Assoc UK:1–13. doi:10.1017/S0025315416001041
Mouillot D, Villeger S, Parravicini V, Kulbicki M, Arias-Gonzalez JE, Bender M, Chabanet P, Floeter SR, Friedlander A, Vigliola L, Bellwood DR (2014) Functional over-redundancy and high functional vulnerability in global fish faunas on tropical reefs. Proc Natl Acad Sci 111(38):13757–13762. doi:10.1073/pnas.1317625111
Pinheiro HT, Ferreira CEL, Joyeux JC et al (2011) Reef fish structure and distribution in a south-western Atlantic Ocean tropical island. J Fish Biol 79:1984–2006. doi:10.1111/j.1095-8649.2011.03138.x
Pitcher TJ, Parrish JK (1993) Functions of schoaling behaviour in teleosts. In: Pitcher TJ (ed) Behav. Teleost Fishes. Belknap Press, London, pp 363–439
Poulin R, Grutter AS (1996) Cleaning symbioses: proximate and adaptive explanations. BioScience 46:512–517. doi:10.2307/1312929
Quimbayo JP, Dias MS, Schlickmann OC, Mendes TC (2016) Fish cleaning interactions on a remote island from the tropical eastern Pacific. Mar Biodivers. doi:10.1007/s12526-016-0493-2
Quimbayo JP, Floeter SR, Noguchi R et al (2012) Cleaning mutualism in Santa Luzia (Cape Verde Archipelago) and São Tomé Islands, tropical eastern Atlantic. Mar Biodivers Rec 5:e118. doi:10.1017/S175526721200108X
Core Team R (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Rincon-Diaz MP, Diez CE, Van Dam RP, Sabat AM (2011) Foraging selectivity of the hawksbill sea turtle (Eretmochelys imbricata) in the Culebra Archipelago, Puerto Rico. J Herpetol 277–282. doi:10.1670/10-120.1
Sasal P (2003) Experimental test of the influence of the size of shoals and density of fish on parasite infections. Coral Reefs 22:241–246. doi:10.1007/s00338-003-0313-6
Sazima C, Bonaldo RM, Krajewski JP, Sazima I (2005) The Noronha wrasse: a “jack-of-all-trades” follower. J Ichthyol Aquat Biol 9:97–108
Sazima C, Grossman A, Sazima I (2010a) Turtle cleaners: reef fishes foraging on epibionts of sea turtles in the tropical southwestern Atlantic, with a summary of this association type. Neotrop Ichthyol 8:187–192. doi:10.1590/S1679-62252010005000003
Sazima C, Guimarães PRJ, Reis SF, Sazima I (2010b) What makes a species central in a cleaning mutualism network? Oikos 119:1319–1325. doi:10.1111/j.1600-0706.2009.18222.x
Sazima I, Moura RL (2000) Shark (Carcharhinus perezi), Cleaned by the Goby (Elacatinus randalli), at Fernando de Noronha Archipelago, Western South Atlantic. Copeia 2000(1):297–299
Sazima C, Sazima I (2001) Plankton-feeding aggregation and occasional cleaning by adult butterflyfish, Chaetodon striatus (Chaetodontidae), in southwestern Atlantic. Cybium 25:145–151
Sazima I, Grossman A, Sazima C (2004) Hawksbill turtles visit moustached barbers: cleaning symbiosis between Eretmochelys imbricata and the shrimp Stenopus hispidus. Biota Neotrop 4:2–7. doi:10.1590/S1676-06032004000100011
Sazima I, Krajewski JP, Bonaldo RM, Sazima C (2013) A vida dos peixes em Fernando de Noronha. Campinas-Brazil
Sazima I, Moura RL, Gasparini JL (1998) The wrasse Halichoeres cyanocephalus (Labridae) as a specialized cleaner fish. Bull Mar Sci 63:605–610
Sazima I, Moura RL, Sazima C (1999) Cleaning activity of juvenile angelfish, Pomacanthus paru, on the reefs of the Abrolhos Archipelago, western South Atlantic. Environ Biol Fish 56:399–407. doi:10.1023/A:1007531925845
Sazima I, Sazima C, Francini-Filho RB, Moura RL (2000) Daily cleaning activity and diversity of clients of the barber goby, Elacatinus figaro, on rocky reefs in southeastern Brazil. Environ Biol Fish 59:69–77. doi:10.1023/A:1007655819374
Slobodkin LB, Fishelson L (1974) The effect of the cleaner-fish Labroides dimidiatus on the point diversity of fishes on the reef front at Eilat. Am Nat 108:369–376
Titus BM, Daly M, Exton DA (2015) Temporal patterns of Pederson shrimp (Ancylomenes pedersoni Chace 1958) cleaning interactions on Caribbean coral reefs. Mar Biol 162:1651–1664. doi:10.1007/s00227-015-2703-z
Vaughan DB, Grutter AS, Costello MJ, Hutson KS (2016) Cleaner fishes and shrimp diversity and a re-evaluation of cleaning symbioses. Fish Fish. 00: 1–19. doi:10.1111/faf.12198
Wicksten MK (1995) Associations of fishes and their cleaners on Coral reefs of Bonaire, Netherlands Antilles. Copeia 1995(2):477. doi:10.2307/1446914
Wicksten MK (1998) Behaviour of cleaners and their client fishes at Bonaire, Netherlands Antilles. J Nat His 32(1):13–30. doi:10.1080/00222939800770021
Acknowledgements
This study was carried out under the “Programa de Monitoramento de Longa Duração das Comunidades Recifais de Ilhas Oceânicas – PELD” (CNPq 403740/2012-6). We thank Jarian Dantas, ICMBio and SOS Mata Atlântica for logistical support. JPQ, NLT, FL and RAM thank CAPES and CNPq for financial support. IS thanks Cristina Sazima and all his former students of cleaning interactions for wholehearted companionship in fieldwork, and Marlies Sazima for loving support over the years.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 34 kb)
Rights and permissions
About this article

Cite this article
Quimbayo, J.P., Nunes, L.T., Ozekoski, R. et al. Cleaning interactions at the only atoll in the South Atlantic. Environ Biol Fish 100, 865–875 (2017). https://doi.org/10.1007/s10641-017-0612-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-017-0612-3