
Abstract
Purpose
There is suggestive but limited evidence for a relationship between meat intake and breast cancer (BC) risk. Few studies included Hispanic women. We investigated the association between meats and fish intake and BC risk among Hispanic and NHW women.
Methods
The study included NHW (1,982 cases and 2,218 controls) and the US Hispanics (1,777 cases and 2,218 controls) from two population-based case–control studies. Analyses considered menopausal status and percent Native American ancestry. We estimated pooled ORs combining harmonized data from both studies, and study- and race-/ethnicity-specific ORs that were combined using fixed or random effects models, depending on heterogeneity levels.
Results
When comparing highest versus lowest tertile of intake, among NHW we observed an association between tuna intake and BC risk (pooled OR 1.25; 95 % CI 1.05–1.50; trend p = 0.006). Among Hispanics, we observed an association between BC risk and processed meat intake (pooled OR 1.42; 95 % CI 1.18–1.71; trend p < 0.001), and between white meat (OR 0.80; 95 % CI 0.67–0.95; trend p = 0.01) and BC risk, driven by poultry. All these findings were supported by meta-analysis using fixed or random effect models and were restricted to estrogen receptor-positive tumors. Processed meats and poultry were not associated with BC risk among NHW women; red meat and fish were not associated with BC risk in either race/ethnic groups.
Conclusions
Our results suggest the presence of ethnic differences in associations between meat and BC risk that may contribute to BC disparities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Breast cancer (BC) incidence rates vary by race/ethnicity in the USA. Non-Hispanic white (NHW) women have the highest age adjusted rates (128.0 per 100,000), whereas Hispanic women have among the lowest rates (93.2 per 100,000) [1]. In spite of the lower incidence rates, Hispanic women are more likely to be diagnosed at advanced disease stages and with estrogen receptor (ER)-negative tumors [2, 3]. Racial/ethnic differences in the distribution of risk factors such as reproductive history, alcohol consumption, and menopausal hormone therapy use [2, 4, 5] may partially explain the disparity in incidence, but do not account for all of the observed variability [5]. While migrant studies found a rise in incidence rates of BC upon immigration to the USA from countries with traditionally low BC incidence rates, such as Latin America and Asia [4, 6], consideration of known risk factors do not fully explain the observed rate differences between the US and foreign-born Hispanic women [4]. Differences in the frequency of predisposing genetic variants may also play a role. Hispanics are a genetically admixed population made up of European, Native American (NA), and African ancestry components. Higher European ancestry is associated with increased BC risk in both the US Hispanic and Mexican women [7, 8], and BC susceptibility loci were identified among Latinas via admixture mapping, and more recently, through genome-wide association analyses [9, 10]. Altogether, the current evidence suggests the presence of unmeasured or poorly characterized BC risk factors might be particularly relevant for Latina women, a growing population.
Diet, particularly meat intake, has not been considered in investigations of BC among Latina women. The World Cancer Research Fund and the American Institute for Cancer Research recommend limiting red and processed meat intake based on conclusive links between meat intake and colorectal cancer [11]. Epidemiological evidence for positive associations between intakes of meat, poultry, and fish and BC risk is less conclusive, but suggestive [11, 12]. Possible mechanisms include oxidative damage from bioavailable heme–iron [13], exposure to exogenous growth-promoting hormones used in animal food production [14], and intake of mutagenic xenobiotic compounds such as heterocyclic amines (HCAs), polycyclic aromatic hydrocarbons (PAHs), and N-nitroso compounds (NOCs) [15, 16]. Meta-analyses of large prospective studies yielded weakly positive associations that failed to reach statistical significance [17, 18]. In contrast, another meta-analysis including cohort and case–control studies performed on pre-menopausal women reported positive summary associations between meat intake and BC risk [19], although there is substantial heterogeneity across studies regarding the choice of model covariates and control selection. In addition, genetic variants may modify the association with meat intake. To date, several studies have investigated variants in mutagen metabolism, with two reporting significant interactions with meat intake [20, 21].
In this study, we investigated the association between meat, poultry, and fish intake and BC risk among NHW and the US Hispanic women. Our goals were to understand the role of meat/fish intake in BC risk and its potential impact on the observed BC incidence rate disparity.
Methods
Study population
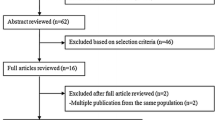
The Breast Cancer Health Disparities Study (BCHD) [22] is a consortium of three case–control studies (two from the USA and one from Mexico). In this analysis, we included data from the two population-based US case–control studies: the 4-Corners Breast Cancer Study (4-CBCS) and the San Francisco Bay Area Breast Cancer Study (SFBCS). Protocols were approved by the Institutional Review Board for Human Subjects at each institution, and all participants signed written informed consents prior to study enrollment.
4-Corners Breast Cancer Study (4-CBCS)
This study consists of NHW, Hispanic, or NA women aged 25–79 years who resided in non-reservation areas within the states of Arizona, Colorado, New Mexico, or Utah at the time of diagnosis (cases) or selection into the study (controls) [23]. State tumor registries were used to identify cases and confirm eligibility criteria, which included a histologically confirmed diagnosis of in situ or invasive breast cancer between October 1999 and May 2004. Information on tumor ER/PR status was also obtained from registry data as indicated in pathology reports. Controls were selected within target populations from sources ranging from commercial mailing lists to driver’s license lists, and frequency matched on ethnicity and 5-year age distribution of cases. Participation rates were 63 and 36 % for Hispanic cases and controls, respectively, and 71 and 47 % for NHW cases and controls, respectively. Trained interviewers administered a structured computerized questionnaires in English or Spanish to collect participant information up to the reference year (the year prior to diagnosis for cases or selection for controls). Dietary intake was assessed using a computerized version of a validated dietary history questionnaire (CARDIA) which captures more than 300 food items [24] and was modified to accommodate foods commonly eaten in the Southwestern US [25]. Weight and height were measured at the time of interview. Of those interviewed, blood for DNA extraction was collected from 76.6 % of cases and 82.4 % of controls.
San Francisco Bay Area Breast Cancer Study (SFBCS)
Participants in SFBCS were NHW, Hispanic, and African American women ages 35–79 years newly diagnosed with a first primary histologically confirmed invasive breast cancer between April 1995 and April 2002 for Hispanic women and between April 1995 and April 1999 for NHW and African American women who resided in the San Francisco Bay Area at the time of diagnosis (counties of San Francisco, San Mateo, Santa Clara, Alameda, and Contra Costa) [4, 26]. Cases were ascertained via the Greater Bay Area Cancer Registry and screened by telephone for self-reported race/ethnicity and study eligibility (89 % participation among those contacted). Information on tumor ER/PR status was obtained from registry data as indicated in pathology reports. All eligible Hispanic and African American women and a 10 % random sample of eligible NHW women were invited to participate in an in-person interview. Controls residing in the San Francisco Bay Area were selected via random digit dialing using the Waksberg method, and frequency matched to cases by race/ethnicity and 5 year age group. They were also screened by telephone for self-reported race/ethnicity and study eligibility (92 % participation among those contacted). Among those eligible for the in-person interview, participation rates were 89 and 88 % for Hispanic cases and controls, respectively, and 86 and 83 % for NHW cases and controls, respectively. Trained bilingual interviewers administered a structured questionnaire to collect participant information up to the reference year (the calendar year prior to diagnosis for cases or selection for controls). Height and weight were measured in person. Dietary intake during the reference year was assessed using a modified version of the Block’s Health History and Habits Questionnaire which captured 85 food items [27]. A biospecimen component was added to the investigation in 1997, and among those eligible, 93 % of cases and 92 % of controls contributed a blood or mouthwash sample.
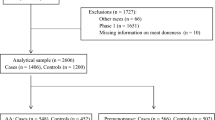
We excluded 158 individuals with missing or extreme caloric intake, defined as daily intake of <600 or >6000 kcal, 301 individuals with in situ BC diagnosis or for whom BC was not the first primary cancer diagnosis, and 128 individuals who self-identified as American Indian/Native American. Prior to further exclusions, there were 8242 study participants: 2064 cases and 2392 controls from 4-CBCS, and 1695 cases and 2091 controls from SFBCS. DNA for genotyping was available for 5544 participants (~67.3 %); thus, analyses adjusted for or stratified by admixture information were performed on this smaller subset. Lastly, participants with missing covariate or exposure data were dropped from the final fitted models. The final main effects models included 7,470 participants, whereas final models utilizing genetic data included 5,079 participants.
Data harmonization and exposure variables
Adjustment variables
Data were harmonized across the two studies. Adjustment variables included body mass index (BMI), calculated as self-reported weight (kg) divided by height (m) squared. Race/ethnicity was self-reported. Age corresponds to age during the reference year. Education was defined as the highest educational level attained (less than high school, high school/GED, post-high school). Reports of history of first-degree relative with breast cancer were dichotomized (yes/no). Parity was defined as the number of live and stillborn births (0, 1–2, 3–4, 5+ births), while age at first birth was defined as age at first live or still birth. These were combined into a single reproductive history variable (nulliparous, <20, 20–24, 25–29, ≥30 years). Lifetime physical activity was scored from 1 (low) to 4 (high), based on study-specific cut-points for hours per week of vigorous activity during the reference year, and at ages 15, 30, and 50. Women were classified as pre-menopausal or post-menopausal based on responses to menstrual history questions. All women who reported having periods during the referent year were classified as pre-menopausal. Women taking menopausal hormone therapy and still having periods were classified as post-menopausal if their age was above the 95th percentile of age distribution among women reporting natural menopause (no periods for ≥12 months) within their corresponding race/ethnicity groups and study center. This age cutoff varied by study: 58 years for NHWs and 56 for Hispanics in 4-CBCS, and 55 for NHWs and 56 for Hispanics in the SFBCS. Alcohol intake (gm/day) was calculated based on lifetime consumption in 4-CBCS and consumption during the reference year in SFBCS, and was categorized as none, <5, 5 to <10, and ≥10. Lastly, dietary variables included in the adjusted models were daily caloric intake (kcal/day) and nutrient density adjusted daily intake of fiber and total fat (g/kcal/day).
Meat/fish consumption
In order to harmonize meat/fish intake variables across studies, we combined each studies’ questionnaire meat items into six categories: red meat, processed meat, white meat (combined poultry and fish intake), poultry, fish, and tuna. Both questionnaires included similar meat items for each category. Red meat includes items such as beef steaks, burgers, roasts, veal, ribs, pork (chops, steaks, roasts, ribs, fresh hams), lamb, and any dishes that included fresh meat as an ingredient. Processed meats include hotdogs, sausages, bacon, luncheon meats, processed ham, and any dishes that included these items. Chicken and turkey make up the poultry category, while fish includes any seafood items, such as white and dark fishes, and shellfish. White meat is a combination of all poultry and fish intake. While the fish variable includes tuna intake, we chose to analyze tuna separately since questionnaires contained questions specific for tuna and dishes containing tuna. As caloric intake could confound associations between meat and BC, the combined meat/fish variables were adjusted for energy intake using the nutrient density method [28] and are expressed as grams per 1,000 kcal of energy intake per day. Lastly, using the nutrient density adjusted variables we categorized intake levels by calculating study- and race-/ethnicity-specific tertiles based on distributions of energy-adjusted meat/fish intake among controls, then combining corresponding tertile levels across studies and race–ethnic groups. For tuna intake, a substantial number of controls (>10 %) reported zero intake during the referent period. These participants were grouped into the lowest tertile group, while the rest were split into the second and third tertile groups based on median levels of intake among controls.
Ancestry informative markers
Genotyping (Goldengate Chemistry, San Diego, CA) was performed as part of a larger effort to investigate the association between variants in genes related to inflammation, hormones, and energetic factors and BC risk in the BCHD Study [22]. We obtained genotype information for 104 Ancestry Informative Markers (AIMs), which were used to categorize women based on percent level of NA ancestry, also previously described [22]. Briefly, using the program STRUCTURE 2.0, individual ancestry for each study participant was estimated assuming two founding populations (European and NA). A three founding population model was assessed, but did not fit the population structure with the same level of repeatability and correlation among runs. Ancestry is expressed as percent Native American.
Statistical analysis
Distributions of covariates by race/ethnicity were summarized using frequencies (proportions) and means (standard deviations), and group differences were assessed using Chi-square and Student’s t tests, respectively. Measures of association between overall meat/fish intake in tertile categories and BC risk were calculated using unconditional logistic regression models. Covariates included age, study center, menopausal status (when not stratified), family history of BC, education, alcohol consumption, parity/age at first birth, physical activity, BMI during reference year, daily caloric intake, intake of fiber, and total fat. Oral contraceptive (never/ever) was accounted for in analyses among pre-menopausal women, and hormone replacement therapy use (never/ever) was accounted for in post-menopausal analyses and analyses of all women combined, but neither significantly change estimates (<10 % change), and thus were omitted from the final results. Trend tests were performed by modeling the indicator variables continuously; median levels of intake are not meaningful given the manner in which the variables were harmonized. All analyses were stratified by race/ethnicity and menopausal status. Among Hispanic women, analyses were stratified by admixture tertile categories which were calculated based on the distribution of NA ancestry among Hispanic controls. Tests of heterogeneity of odds ratios by menopausal status, race/ethnicity, and admixture categories were computed using likelihood ratio tests of models including and excluding interaction terms between these variables and meat/fish intake. Sub-analyses included mutual adjustment for other meat, poultry, and fish intake variables. Lastly, multinomial logistic regression was used to model the risk of BC by tumor estrogen receptor status, with controls serving as the reference group.
As mentioned previously, 4-CBCS and SFBCS employed different instruments to measure dietary intake, leading to study differences in daily intake values of meat and fish. To address this, we calculated study- and race-/ethnicity-specific tertiles. To further address any potential heterogeneity across studies, we performed additional analyses to combine study-specific odds ratios for each racial/ethnic group. We obtained these combined ORs by using random [29] or fixed effects models depending on the level of study heterogeneity as determined by Cochran’s Q [29] and I 2 [30] statistics. Specifically, among NHW women the Cochran Q was <0.05 or I 2 > 50 %; thus, random effects models were employed exclusively. There was less study heterogeneity among Hispanic women, with Cochran Q > 0.05 and I 2 < 50 %, with the exception of poultry intake. Therefore, combined odds ratios for all variables, except poultry, were obtained using fixed effect models, whereas for poultry intake random effects models were used. We also obtained combined ORs stratified by estrogen receptor status. We first calculated study- and race-specific results for each breast cancer subtype (ER+ and ER−) compared to controls using unconditional logistic regression and then combined these results using random/fixed effects models, as above.
All hypothesis tests were two-sided. Analyses were performed using the statistical software STATA SE 12.0 (STATA Corporation, College Station, TX).
Results
Characteristics of study participants stratified by race/ethnicity are given in Table 1. Overall, NHW cases and controls were older than Hispanic cases and controls and were more likely to have a first-degree family history of breast cancer. In addition, NHW cases and controls consumed more alcohol, were more often nulliparous and older at first birth, were more likely to use oral contraceptives and undergo hormone replacement therapy, and attained higher levels of education than Hispanic cases and controls, whereas the latter had higher BMI and daily caloric intake levels. Compared to controls, both NHW and Hispanic cases were more likely to have a first-degree family history of BC, consume more alcohol, and be nulliparous. Among Hispanics, higher education levels and older age at first birth were associated with BC risk, while higher BMI was inversely associated with BC risk. In general, Hispanic women consumed less meats and fish than NHW women, with the exception of processed meat, for which consumption was lower among Hispanic women in SFBCS and higher in 4-CBCS.
Meat/fish variables and BC risk among NHW and Hispanic women
We investigated the association between six meat/fish variables and BC risk among NHW women (Table 2) and among Hispanic women (Table 3), stratifying by menopausal status, pooling data from both case–control studies. Among NHW women, tuna intake was the only meat/fish variable associated with BC risk (T3 vs. T1 OR 1.25; 95 % CI 1.05–1.50), with comparable OR estimates for pre- and post-menopausal women (Table 2). We observed a similar association between high intake of tuna and BC risk among Hispanic women (overall T3 vs. T1 OR 1.21; 95 % CI 1.02–1.44) (Table 3), with a significant association among post-menopausal women only (T3 vs. T1 OR 1.29, 95 % CI 1.04–1.61); however, there was no evidence of statistically significant heterogeneity by menopausal status. In contrast, among Hispanics, high intake of processed meats was associated with increased BC risk (T3 vs. T1 OR 1.42; 95 % CI 1.18–1.71), with similar results for pre- and post-menopausal women. The observed difference in ORs associated with processed meats intake between NHW and Hispanic women was statistically significant (2df heterogeneity p value 0.03; data not shown). Among Hispanic women, we also observed an inverse association between high poultry intake and BC risk (T3 vs. T1 OR 0.80; 95 % CI 0.67–0.95) (Table 3). The inverse association was limited to pre-menopausal women, but there was no evidence of statistically significant heterogeneity by menopausal status. Furthermore, there was no evidence of effect modification of association between poultry and BC risk by race. Similarly, we observed an inverse association between high white meat intake and BC risk (T3 vs. T1 OR 0.80; 95 % CI 0.67–0.95), with no significant heterogeneity by menopausal status (Table 3). Mutual adjustment for other meat/fish intake variables did not drastically change estimates.
Given the wide variation in NA ancestry among Hispanic women, we investigated whether the associations with meat/fish variables differed by tertiles of NA ancestry (Supplemental Table 1). We observed that the positive association between diets high in processed meats and BC risk was found only in Hispanic women with intermediate (T3 vs. T1 OR 1.62; 95 % 1.10–2.40) and high (T3 vs. T1 OR 1.88; 95 % 1.19–2.95) NA ancestry, but not among women within the low category (T3 vs. T1 OR 0.97; 95 % 0.66–1.45), suggesting a possible modifying effect of NA ancestry; however, tests for heterogeneity were not statistically significant. Similarly, we observed that the inverse association with poultry intake was restricted to women with intermediate NA ancestry, but again we found no evidence of significant heterogeneity. Furthermore, interaction tests were not significant when modeling admixture continuously. In unstratified models that included admixture as a covariate, associations between processed meat (T3 vs. T1 OR 1.45; 95 % CI 1.15–1.82; data not shown) and poultry intake (T3 vs. T1 OR 0.79; 95 % CI 0.64–0.98; data not shown) and BC risk remained statistically significant, whereas associations between tuna intake and BC risk did not.
Meat/fish variables and BC risk according to ER status
When considering BC subtypes defined by ER status, among NHW women (Table 4) we observed that the positive association with tuna intake was limited to ER+ BC cases (T3 vs. T1 OR 1.46; 95 % CI 1.18–1.81). No significant associations were observed for women with ER− BC (heterogeneity p = 0.003). Among Hispanic women (Table 5), associations with processed meat intake were only statistically significant among ER+ BC (T3 vs. T1 OR 1.45; 95 % CI 1.16–1.81, heterogeneity p = 0.004) as were associations with poultry (T3 vs. T1 OR 0.77; 95 % CI 0.63–0.94, heterogeneity p = 0.04) and white meat intake (T3 vs. T1 OR 0.74; 95 % CI 0.60–0.91, heterogeneity p = 0.02). Further stratification by menopausal status was not performed due to small numbers.
Meta-analysis results
To address any possible residual heterogeneity across the two case–control studies that was not accounted for by variable harmonization, adjustment for study, and use of study- and race-/ethnicity-specific exposure cut-points, we also combined study- and race-/ethnicity-specific ORs via random/fixed effects models, for NHW and Hispanic women separately. Results varied by study to a greater extent among NHW, where all significant associations seemed restricted to the SFBCS study. Upon pooling results via random effects models, tuna intake was still positively associated with BC risk (T3 vs. T1 combined OR 1.31; 95 % CI 0.90–1.91; p trend = 0.014) (Table 6).
Less heterogeneity across the two case–control studies was observed among Hispanics. Processed meat intake was positively associated in both 4-CBCS and SFBS studies. Likewise, white meat intake was consistently associated with decreased BC risk in both studies. Consequently, in combined analyses both processed meats (T3 vs. T1 combined OR 1.38; 95 % CI 1.14–1.67) and white meats (T3 vs. T1 combined OR 0.78; 95 % CI 0.65–0.93) were positively and inversely associated with BC risk, respectively (Table 6). Tuna and poultry intake were no longer statistically significantly associated with BC risk in meta-analysis, although results for poultry are still suggestive of an inverse association (Table 6).
Meta-analyses stratified by ER status yielded similar findings as those obtained with pooled analyses. Tuna associations among NHW women were limited to ER+ cases, whereas processed meat and white meat associations with BC risk maintained statistical significance only among Hispanic ER+ cases (Supplemental Table 2). Similarly to results from pooled analyses, poultry and tuna were not associated with either ER+ nor ER− among Hispanic women.
Discussion
In this pooled case–control analysis, we found evidence that diets high in tuna intake may increase the risk of BC risk among both NHW and Hispanic women, whereas positive associations with diets high in processed meats and inverse associations with diets high in poultry and white meat were found among Hispanic women only. The associations among Hispanic women did not seem to be modified by NA ancestry. Our findings were similar when combining estimates via random/fixed models: Tuna intake was positively associated with BC risk among NHW women only, whereas processed and white meat intake was associated with increased and decreased risk of BC, respectively, among Hispanic women. To our knowledge, this is one of the first investigations of meat/fish intake and BC risk in a large population of the US Hispanics.
Previous investigations of fish intake and BC risk have produced inconclusive results. A meta-analysis that included 11 prospective studies concluded that fish intake was not associated with BC risk [31]. Furthermore, a 2013 prospective study conducted among the US black women also failed to find an association [32], as did a more recent prospective study conducted in Japan, where fish makes up a relatively higher proportion of daily dietary consumption [33]. In contrast, a prospective study from Denmark reported that overall intake of fish was associated with higher incidence rates of BC independently of fish fat content or preparation methods [34]. Several case–control studies have investigated fish intake and BC risk with inconclusive results, some reporting no evidence of association [20, 35–37], evidence of inverse associations for fatty fish among both pre- and post-menopausal women in Korea [38] and post-menopausal women in the USA [39], or evidence of a positive association [40].
In this study, although overall fish consumption was not associated with BC risk, we found a positive association with tuna intake. None of the previously mentioned studies reported findings for specific fish species, such as tuna. The health benefits of fish intake are often attributed to the consumption of omega 3 polyunsaturated fatty acids (n-3 PUFA), which may act in several pathways that inhibit tumor progression [41]. In the previously mentioned meta-analysis, Zheng et al. reported a protective effect for marine n-3 PUFA in a dose-responsive manner [31]. Nevertheless, the benefits of PUFA intake may be outweighed by exposures to chemical contaminants such as persistent organic pollutants and metals, potentially found in fish, contingent on species, portion size, and frequency of consumption [42]. Certain metals such as mercury and cadmium may activate estrogen receptors in the absence of estradiol, and there is epidemiological support for an association between exposure to these metals and an increased risk of BC [43]. In terms of PUFA content, tuna ranks relatively low compared to other commonly consumed species [44], but may be responsible for a greater share of exposures to chemical pollutants [42]. In the USA, tuna is frequently consumed in canned form, and different types of canned tuna contain varying amounts of mercury contamination [45]. In our study, the 4-CBCS FFQ captured tuna intake information in terms of various canned tuna and tuna salad, while the SFBCS FFQ asked about overall tuna intake, such as fresh, canned, or as part of a dish. Thus, we could not investigate associations with specific tuna products separately. Therefore, this issue deserves further investigation. We cannot discard the possibility that the association between tuna intake and BC risk may be driven by chance, given the number of comparisons we made, or by residual confounding by factors unmeasured in our study.
Several studies investigated processed meat intake and BC risk by grouping food items such as hotdogs, bacon, sausages, and luncheon meats, with equivocal results. Among ten prospective studies, four studies reported statistically significant positive associations with processed meats [20, 46–48], one a non-statistically significant positive association [49], and five studies reported no associations [32, 50–53]. A pooled analysis of eight additional cohort studies reported no association with processed meats [18]. In addition, among seven population-based case–control studies that considered processed meats separately from unprocessed red meat, five reported a positive association with processed meats [36, 54–57], and two reported non-statistically significant positive associations [39, 58]. Recently, a meta-analysis of all available prospective studies reported a positive association with processed meats and breast cancer risk [59]; however, we note that this study included overlapping studies; therefore, the conclusions may not be fully representative of the available data. Among three hospital-based case–control studies, two reported non-statistically significant positive associations [60, 61], and one reported no associations with processed meats [62]. In our study, we found that processed meat intake was associated with elevated BC risk among Hispanic women only, with no evidence of heterogeneity by menopausal status. We do not have a clear explanation for the racial/ethnic difference in our results. Although we attempted to adjust for most well-known putative confounders, the presence of additional unmeasured or unknown confounding factors is a possibility, particularly factors that may be unique to Hispanic women. Another possibility is the presence of a threshold effect for processed meats. In our study, Hispanic women consumed significantly higher mean levels of processed meats than NHW women (14.2 vs. 12.4 g/day among controls) during the reference year. However, once processed meat intake is nutrient density adjusted (gm/1,000 kcal/day), the differences in consumption per 1,000 kcal are not as large. Hispanic cases and controls in the SFBCS study had lower levels of nutrient density adjusted processed meat consumption compared to NHW women, while Hispanic cases and controls in the 4-CBCS had higher levels compared to NHW women.
Given that breast development occurs during adolescence, early life exposures might be more influential in determining future BC risk than exposures during midlife [63]; a study done within the Nurses’ Health Study II cohort reported a positive association between early adulthood total red meat consumption and BC risk [64]. In a related study using the same cohort, an association was reported between BC risk in pre-menopausal women and total red meat consumption and total processed meats during high school [65]. Acculturation after migration to the USA may contribute to a net increase in intake of unhealthy food sources among Mexican women [66]. Thus, it is plausible that Hispanic adolescent exposures to higher levels of processed meats may explain our results. Another possibility is that the differences in the observed associations for processed meats may be due to ethnic differences in genetic susceptibility to exposure to meat-related carcinogens. Two studies, among Danish and Chinese populations, found evidence of interaction between red and smoked meat intake, respectively, and the carcinogen metabolism enzymes NAT1 and NAT2 [20, 21]. In our analyses, adjustment and stratification by NA ancestry categories did not change results dramatically, suggesting that associations with processed meats, or other risk factors captured by this exposure, are similar across Hispanic women, regardless of possible differences at the genetic level. Further research is necessary to clarify the nature of the association between processed meats and BC risk among Hispanics.
It is also unclear why poultry and white meat were protective only among Hispanic women. Although the poultry association did not retain statistical significance when combining results via meta-analysis for all women combined, there was still evidence of a significant association in stratified analyses by menopausal status. Evidence for poultry and white meat intake is inconclusive, with many investigations of poultry yielding null results [20, 37, 61, 67], with a few finding positive associations with BC risk [40, 58]. Like processed meats, there could be differences in the timing of exposure (e.g., earlier in adolescence vs. late) [65]. We cannot discard residual confounding, as high intake of white meats may indicate overall healthier eating patterns [68]. White meat and processed meat intake were negatively correlated, but the correlation coefficients were very small, and did not differ by race/ethnicity.
To our knowledge, this is the first study to examine the association between meat/fish intake and BC risk among Hispanic women in comparison with NHWs. Our study has many strengths, such as the inclusion a large number of Hispanic women in addition to NHWs, with a large proportion of women contributing data on meat intake and ER and PR status. In addition, we were able to consider global genetic ancestry in an effort to control for the known genetic heterogeneity among Hispanics. Limitations include the harmonized food intake data from two different FFQs, which could introduce artificial variability in consumption levels. We attempted to address this by creating tertiles based on study-specific cutoff levels, and by adjusting meat/fish intake levels by energy intake. We also conducted meta-analysis of study- and race-/ethnicity-specific odds ratios in order to corroborate findings using these harmonized categorical variables and account further for possible inter-study heterogeneity. Another limitation of our study was the inability to harmonize cooking methods information, which would have allowed estimation of mutagen consumption and also closer investigation of the tuna intake associations. We also recognize that participation rates were lower in 4-CBCS compared to SFBCS, adding the possibility of selection bias and biased exposure reports. All our analyses adjusted for study center, so much of the variability introduced by these factors may have been attenuated.
In summary, we report that diets high in tuna fish may increase risk of BC among NHW and possibly also among Hispanic women. Moreover, we observed that diets high in processed meats may increase risk of BC risk among Hispanic women, albeit not comparable evidence was observed among NHWs. Further research is needed to understand the possible reason for the ethnic differences in associations with processed meat intake and the role of processed meats in BC formation among Latinas.
References
Howlader N, Noone A, Krapcho M et al (2014) SEER Cancer Statistics Review, 1975–2011, National Cancer Institute. Bethesda, MD, http://seer.cancer.gov/csr/1975_2011/, based on November 2013 SEER data submission, posted to the SEER web site, April 2014
Sweeney C, Baumgartner KB, Byers T et al (2008) Reproductive history in relation to breast cancer risk among Hispanic and non-Hispanic white women. Cancer Causes Control 19:391–401. doi:10.1007/s10552-007-9098-1
Siegel R, Naishadham D, Jemal A (2012) Cancer statistics for Hispanics/Latinos, 2012. CA Cancer J Clin 62:283–298. doi:10.3322/caac.21153
John EM, Phipps AI, Davis A, Koo J (2005) Migration history, acculturation, and breast cancer risk in Hispanic women. Cancer Epidemiol Biomark Prev 14:2905–2913. doi:10.1158/1055-9965.EPI-05-0483
Chlebowski RT, Chen Z, Anderson GL et al (2005) Ethnicity and breast cancer: factors influencing differences in incidence and outcome. J Natl Cancer Inst 97:439–448. doi:10.1093/jnci/dji064
Ziegler RG, Hoover RN, Pike MC et al (1993) Migration patterns and breast cancer risk in Asian-American women. JNCI J Natl Cancer Inst 85:1819–1827. doi:10.1093/jnci/85.22.1819
Fejerman L, John EM, Huntsman S et al (2008) Genetic ancestry and risk of breast cancer among U.S. Latinas. Cancer Res 68:9723–9728. doi:10.1158/0008-5472.CAN-08-2039
Fejerman L, Romieu I, John EM et al (2010) European ancestry is positively associated with breast cancer risk in Mexican women. Cancer Epidemiol Biomark Prev 19:1074–1082. doi:10.1158/1055-9965.EPI-09-1193
Fejerman L, Chen GK, Eng C et al (2012) Admixture mapping identifies a locus on 6q25 associated with breast cancer risk in US Latinas. Hum Mol Genet 21:1907–1917. doi:10.1093/hmg/ddr617
Fejerman L, Ahmadiyeh N, Hu D et al (2014) Genome-wide association study of breast cancer in Latinas identifies novel protective variants on 6q25. Nat Commun 5:5260. doi:10.1038/ncomms6260
World Cancer Research Fund/American Institute for Cancer Research (2010) Continuous update project report. Food, nutrition, physical activity, and the prevention of breast cancer
Romieu I (2011) Diet and breast cancer. Salud Publica Mex 53:430–439
Huang X (2003) Iron overload and its association with cancer risk in humans: evidence for iron as a carcinogenic metal. Mutat Res 533:153–171
Andersson AM, Skakkebaek NE (1999) Exposure to exogenous estrogens in food: possible impact on human development and health. Eur J Endocrinol 140:477–485. doi:10.1530/eje.0.1400477
Felton JS, Knize MG, Wu RW et al (2007) Mutagenic potency of food-derived heterocyclic amines. Mutat Res 616:90–94. doi:10.1016/j.mrfmmm.2006.11.010
Rothman N, Poirier MC, Baser ME et al (1990) Formation of polycyclic aromatic hydrocarbon-DNA adducts in peripheral white blood cells during consumption of charcoal-broiled beef. Carcinogenesis 11:1241–1243. doi:10.1093/carcin/11.7.1241
Alexander DD, Morimoto LM, Mink PJ, Cushing CA (2010) A review and meta-analysis of red and processed meat consumption and breast cancer. Nutr Res Rev 23:349–365. doi:10.1017/S0954422410000235
Missmer SA, Smith-Warner SA, Spiegelman D et al (2002) Meat and dairy food consumption and breast cancer: a pooled analysis of cohort studies. Int J Epidemiol 31:78–85. doi:10.1093/ije/31.1.78
Taylor VH, Misra M, Mukherjee SD (2009) Is red meat intake a risk factor for breast cancer among premenopausal women? Breast Cancer Res Treat 117:1–8. doi:10.1007/s10549-009-0441-y
Egeberg R, Olsen A, Autrup H et al (2008) Meat consumption, N-acetyl transferase 1 and 2 polymorphism and risk of breast cancer in Danish postmenopausal women. Eur J Cancer Prev 17:39–47. doi:10.1097/CEJ.0b013e32809b4cdd
Lee H, Wang Q, Yang F et al (2012) SULT1A1 Arg213His polymorphism, smoked meat, and breast cancer risk: a case–control study and meta-analysis. DNA Cell Biol 31:688–699. doi:10.1089/dna.2011.1403
Slattery ML, John EM, Torres-Mejia G et al (2012) Genetic variation in genes involved in hormones, inflammation and energetic factors and breast cancer risk in an admixed population. Carcinogenesis 33:1512–1521. doi:10.1093/carcin/bgs163
Slattery ML, Sweeney C, Edwards S et al (2007) Body size, weight change, fat distribution and breast cancer risk in Hispanic and non-Hispanic white women. Breast Cancer Res Treat 102:85–101. doi:10.1007/s10549-006-9292-y
McDonald A, Van Horn L, Slattery M et al (1991) The CARDIA dietary history: development, implementation, and evaluation. J Am Diet Assoc 91:1104–1112
Slattery ML, Caan BJ, Duncan D et al (1994) A computerized diet history questionnaire for epidemiologic studies. J Am Diet Assoc 94:761–766
John EM, Horn-Ross PL, Koo J (2003) Lifetime physical activity and breast cancer risk in a multiethnic population: the San Francisco Bay area breast cancer study. Cancer Epidemiol Biomark Prev 12:1143–1152
Horn-Ross PL, John EM, Lee M et al (2001) Phytoestrogen consumption and breast cancer risk in a multiethnic population: the Bay Area Breast Cancer Study. Am J Epidemiol 154:434–441
Willett WC, Howe GR, Kushi LH (1997) Adjustment for total energy intake in epidemiologic studies. Am J Clin Nutr 65:1220S–1228S (discussion 1229S–1231S)
DerSimonian R, Laird N (1986) Meta-analysis in clinical trials. Control Clin Trials 7:177–188
Higgins JPT, Thompson SG (2002) Quantifying heterogeneity in a meta-analysis. Stat Med 21:1539–1558. doi:10.1002/sim.1186
Zheng J, Hu X, Zhao Y et al (2013) Intake of fish and marine n-3 polyunsaturated fatty acids and risk of breast cancer: meta-analysis of data from 21 independent prospective cohort studies. BMJ 346:f3706. doi:10.1136/bmj.f3706
Genkinger JM, Makambi KH, Palmer JR et al (2013) Consumption of dairy and meat in relation to breast cancer risk in the Black Women’s Health Study. Cancer Causes Control 24:675–684. doi:10.1007/s10552-013-0146-8
Kiyabu GY, Inoue M, Saito E et al (2015) Fish, n-3 polyunsaturated fatty acids and n-6 polyunsaturated fatty acids intake and breast cancer risk: the Japan Public Health Center-based prospective study. Int J Cancer. doi:10.1002/ijc.29672
Stripp C, Overvad K, Christensen J et al (2003) Fish intake is positively associated with breast cancer incidence rate. J Nutr 133:3664–3669
Bessaoud F, Daurès J-P, Gerber M (2008) Dietary factors and breast cancer risk: a case control study among a population in Southern France. Nutr Cancer 60:177–187. doi:10.1080/01635580701649651
Hermann S, Linseisen J, Chang-Claude J (2002) Nutrition and breast cancer risk by age 50: a population-based case–control study in Germany. Nutr Cancer 44:23–34. doi:10.1207/S15327914NC441_4
Hu J, La Vecchia C, DesMeules M et al (2008) Meat and fish consumption and cancer in Canada. Nutr Cancer 60:313–324. doi:10.1080/01635580701759724
Kim J, Lim S-Y, Shin A et al (2009) Fatty fish and fish omega-3 fatty acid intakes decrease the breast cancer risk: a case–control study. BMC Cancer 9:216. doi:10.1186/1471-2407-9-216
Ambrosone CB, Freudenheim JL, Sinha R et al (1998) Breast cancer risk, meat consumption and N-acetyltransferase (NAT2) genetic polymorphisms. Int J Cancer 75:825–830
Bao P-P, Shu X-O, Zheng Y et al (2012) Fruit, vegetable, and animal food intake and breast cancer risk by hormone receptor status. Nutr Cancer 64:806–819. doi:10.1080/01635581.2012.707277
Larsson SC, Kumlin M, Ingelman-Sundberg M, Wolk A (2004) Dietary long-chain n-3 fatty acids for the prevention of cancer: a review of potential mechanisms. Am J Clin Nutr 79:935–945
Domingo JL, Bocio A, Falcó G, Llobet JM (2007) Benefits and risks of fish consumption part I. A quantitative analysis of the intake of omega-3 fatty acids and chemical contaminants. Toxicology 230:219–226. doi:10.1016/j.tox.2006.11.054
Byrne C, Divekar SD, Storchan GB et al (2013) Metals and breast cancer. J Mammary Gland Biol Neoplasia 18:63–73. doi:10.1007/s10911-013-9273-9
Sidhu KS (2003) Health benefits and potential risks related to consumption of fish or fish oil. Regul Toxicol Pharmacol 38:336–344. doi:10.1016/j.yrtph.2003.07.002
Burger J, Gochfeld M (2004) Mercury in canned tuna: white versus light and temporal variation. Environ Res 96:239–249. doi:10.1016/j.envres.2003.12.001
Taylor EF, Burley VJ, Greenwood DC, Cade JE (2007) Meat consumption and risk of breast cancer in the UK Women’s Cohort Study. Br J Cancer 96:1139–1146. doi:10.1038/sj.bjc.6603689
Pala V, Krogh V, Berrino F et al (2009) Meat, eggs, dairy products, and risk of breast cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort. Am J Clin Nutr 90:602–612. doi:10.3945/ajcn.2008.27173
Pouchieu C, Deschasaux M, Hercberg S et al (2014) Prospective association between red and processed meat intakes and breast cancer risk: modulation by an antioxidant supplementation in the SU.VI.MAX randomized controlled trial. Int J Epidemiol 43:1583–1592. doi:10.1093/ije/dyu134
Fung TT, Hu FB, Holmes MD et al (2005) Dietary patterns and the risk of postmenopausal breast cancer. Int J Cancer 116:116–121. doi:10.1002/ijc.20999
van der Hel OL, Peeters PHM, Hein DW et al (2004) GSTM1 null genotype, red meat consumption and breast cancer risk (The Netherlands). Cancer Causes Control 15:295–303. doi:10.1023/B:CACO.0000024255.16305.f4
Wu K, Sinha R, Holmes MD et al (2010) Meat mutagens and breast cancer in postmenopausal women–a cohort analysis. Cancer Epidemiol Biomark Prev 19:1301–1310. doi:10.1158/1055-9965.EPI-10-0002
Kabat GC, Cross AJ, Park Y et al (2009) Meat intake and meat preparation in relation to risk of postmenopausal breast cancer in the NIH-AARP diet and health study. Int J Cancer 124:2430–2435. doi:10.1002/ijc.24203
Ferrucci LM, Cross AJ, Graubard BI et al (2009) Intake of meat, meat mutagens, and iron and the risk of breast cancer in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial. Br J Cancer 101:178–184. doi:10.1038/sj.bjc.6605118
Goodman MT, Nomura AM, Wilkens LR, Hankin J (1992) The association of diet, obesity, and breast cancer in Hawaii. Cancer Epidemiol Biomark Prev 1:269–275
Steck SE, Gaudet MM, Eng SM et al (2007) Cooked meat and risk of breast cancer—lifetime versus recent dietary intake. Epidemiology 18:373–382. doi:10.1097/01.ede.0000259968.11151.06
Fu Z, Deming SL, Fair AM et al (2011) Well-done meat intake and meat-derived mutagen exposures in relation to breast cancer risk: the Nashville Breast Health Study. Breast Cancer Res Treat 129:919–928. doi:10.1007/s10549-011-1538-7
Mourouti N, Kontogianni MD, Papavagelis C et al (2015) Meat consumption and breast cancer: a case–control study in women. Meat Sci 100:195–201. doi:10.1016/j.meatsci.2014.10.019
Chandran U, Zirpoli G, Ciupak G et al (2013) Racial disparities in red meat and poultry intake and breast cancer risk. Cancer Causes Control 24:2217–2229. doi:10.1007/s10552-013-0299-5
Guo J, Wei W, Zhan L (2015) Red and processed meat intake and risk of breast cancer: a meta-analysis of prospective studies. Breast Cancer Res Treat 151:191–198. doi:10.1007/s10549-015-3380-9
Richardson S, Gerber M, Cenée S (1991) The role of fat, animal protein and some vitamin consumption in breast cancer: a case control study in southern France. Int J Cancer 48:1–9
Zhang C-X, Ho SC, Chen Y-M et al (2009) Meat and egg consumption and risk of breast cancer among Chinese women. Cancer Causes Control 20:1845–1853. doi:10.1007/s10552-009-9377-0
Franceschi S, Favero A, La Vecchia C et al (1995) Influence of food groups and food diversity on breast cancer risk in Italy. Int J Cancer 63:785–789
Engelman RW, Day NK, Good RA (1994) Calorie intake during mammary development influences cancer risk: lasting inhibition of C3H/HeOu mammary tumorigenesis by peripubertal calorie restriction. Cancer Res 54:5724–5730
Farvid MS, Cho E, Chen WY et al (2014) Dietary protein sources in early adulthood and breast cancer incidence: prospective cohort study. BMJ 348:g3437. doi:10.1136/bmj.g3437
Farvid MS, Cho E, Chen WY et al (2015) Adolescent meat intake and breast cancer risk. Int J Cancer 136:1909–1920. doi:10.1002/ijc.29218
Batis C, Hernandez-Barrera L, Barquera S et al (2011) Food acculturation drives dietary differences among Mexicans, Mexican Americans, and Non-Hispanic Whites. J Nutr 141:1898–1906. doi:10.3945/jn.111.141473
Dai Q, Shu X-O, Jin F et al (2002) Consumption of animal foods, cooking methods, and risk of breast cancer. Cancer Epidemiol Biomark Prev 11:801–808
Daniel CR, Cross AJ, Graubard BI et al (2011) Prospective investigation of poultry and fish intake in relation to cancer risk. Cancer Prev Res 4:1903–1911. doi:10.1158/1940-6207.CAPR-11-0241
Acknowledgments
The Breast Cancer Health Disparities Study was funded by Grant CA14002 from the National Cancer Institute to Dr. Slattery. The San Francisco Bay Area Breast Cancer Study was supported by Grants CA63446 and CA77305 from the National Cancer Institute, Grant DAMD17-96-1-6071 from the US Department of Defense, and Grant 7PB-0068 from the California Breast Cancer Research Program. The collection of cancer incidence data used in this study was supported by the California Department of Public Health as part of the statewide cancer reporting program mandated by California Health and Safety Code Section 103885; the National Cancer Institute’s Surveillance, Epidemiology and End Results Program under contract HHSN261201000036C awarded to the Cancer Prevention Institute of California; and the Centers for Disease Control and Prevention’s National Program of Cancer Registries, under agreement #1U58 DP000807-01 awarded to the Public Health Institute. The 4-Corners Breast Cancer Study was funded by Grants CA078682, CA078762, CA078552, and CA078802 from the National Cancer Institute. The research also was supported by the Utah Cancer Registry, which is funded by contract N01-PC-67000 from the National Cancer Institute, with additional support from the State of Utah Department of Health, the New Mexico Tumor Registry, and the Arizona and Colorado cancer registries, funded by the Centers for Disease Control and Prevention National Program of Cancer Registries and additional state support. The contents of this manuscript are solely the responsibility of the authors and do not necessarily represent the official view of the National Cancer Institute or endorsement by the State of California Department of Public Health, the National Cancer Institute, and the Centers for Disease Control and Prevention or their Contractors and Subcontractors. The Mexico Breast Cancer Study was funded by Consejo Nacional de Ciencia y Tecnología (CONACyT) (SALUD-2002-C01-7462). Mariana C. Stern received support from Grant RSF-09-020-01-CNE from the American Cancer Society, from award number 5P30 ES07048 from the National Institute of Environmental Health Sciences and award number P30CA014089 from the National Cancer Institute. Andre E. Kim received support from Grant 5T32 ES013678 from the National Institute of Environmental Health Sciences.
Author contributions
We would also like to acknowledge the contributions of the following individuals to the study: Sandra Edwards for data harmonization oversight; Jennifer Herrick for data management and data harmonization; Erica Wolff and Michael Hoffman for laboratory support; Jocelyn Koo for data management for the San Francisco Bay Area Breast Cancer Study; Dr. Tim Byers for his contribution to the 4-Corners Breast Cancer Study; and Dr. Josh Galanter for assistance in selection of AIMs markers.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Martha L. Slattery and Mariana C. Stern: Co-senior authors.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Kim, A.E., Lundgreen, A., Wolff, R.K. et al. Red meat, poultry, and fish intake and breast cancer risk among Hispanic and Non-Hispanic white women: The Breast Cancer Health Disparities Study. Cancer Causes Control 27, 527–543 (2016). https://doi.org/10.1007/s10552-016-0727-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10552-016-0727-4