
Abstract
Despite being a crucial component of nutrient cycling and soil carbon (C) dynamics in forest ecosystems, there is too little information from past studies to discern whether dissolved organic carbon (DOC) exchanges with soil organic carbon or passes unaltered through soils. In this study, we added 13C-labelled litter-derived DOC into different depth soil columns in a 180-day incubation experiment to determine the fate of DOC in soils, and to monitor the changes in DOC composition when it percolates through the soil. The results showed that δ13C values increased in soil microbes, which indicated that some litter-derived DOC was immobilized by soil microbial communities. Approximately 76% of litter-derived DOC was retained in the soil (60% in topsoil and 16% in midsoil). Meanwhile, 18%, 4%, and 3% of litter-derived DOC were mineralized into CO2 in topsoil, midsoil and subsoil respectively. Only 0.04% of litter-derived DOC leached from the soil column (0–60 cm). These results indicated that DOC was mainly retained on soil, and a small portion was mineralized by microorganisms, with minimal leaching. The composition of water soluble soil organic carbon (WSOC) and leachate DOC (LDOC) were similar between the control and treatment. This indicated that the composition of WSOC and LDOC was more similar to soil C than the added DOC, which supports the previously hypothesized dynamic exchange model. These findings provide new insight by showing that most litter-derived DOC is sequestered in forest soils.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Dissolved organic carbon (DOC) is a key component of the terrestrial carbon (C) cycle. Although a small portion of the total soil organic carbon (SOC) pool, DOC is active in various biotic and abiotic processes in the soil C cycle (De Troyer et al. 2011; Kalbitz et al. 2000; Kindler et al. 2011). At the same time, DOC contributes to C sequestration in soils and microbial and plant nutrition (Scott and Rothstein 2014). Litter-derived DOC is an important source of SOC, but we know little of how it is processed in soil. Thus, it is imperative to understand the contributions of litter-derived DOC to SOC.
When litter-derived DOC infiltrates soil, a series of reactions occur, including retention, mineralization, and leaching into the deeper layers (Moller et al. 2005; Qualls et al. 2002). Incubation experiments have been used to show that added litter-derived DOC is mainly mineralized by microbial communities, retained by soil particles, or leached from soils (Hagedorn et al. 2004; Muller et al. 2009; Qiu et al. 2016). However, the proportions of mineralization, retention and leaching have differed with forest soil type. Previous studies have shown that different soils have different retention capacities relative to texture (Seely et al. 1998), structure (Asano et al. 2006), soil C content (Lilienfein et al. 2004), and mineral composition (Lilienfein et al. 2004; Qualls et al. 2000; Yano et al. 2000). To trace the fate of plant-derived DOC, most researchers directly put soils and plant-derived DOC into glass jars, specimen cups or Erlenmeyer flasks to incubate (Hagedorn et al. 2004; Miao et al. 2017; Qiu et al. 2016; Sanderman et al. 2008b). However, these methods do not reflect effects of different depths of soils and vertical leaching of DOC from soil. The few studies that have used soil column experiments to quantity the fate of litter-derived DOC in soil have shown this method to be effective when tracing the fate of litter-derived DOC (Muller et al. 2009; Scott and Rothstein 2014). However, these soil column experiments did not distinguish contributions of different soil layers, or the experiments underestimated DOC mineralization. Therefore, we refined the approach to quantify how much litter-derived DOC is mineralized, retained or leached in different soil layers in our study site.
The molecular weights and chemical composition of the dissolved organic matter (DOM) change with mineralization, retention or leaching when moving downward through artificial soil columns (Kim et al. 2015). The dynamic exchange model is a common explanation of the changes in the molecular structure of DOM with soil water movement (Kaiser and Kalbitz 2012; Sanderman et al. 2008b; Scott and Rothstein 2014). According to this model, DOM composition changes as a result of exchange reactions between highly sorptive litter-derived compounds and previously sorbed microbial-derived compounds as water percolates through soil. As a result, DOM composition in the subsoil would become more similar to the SOM composition as litter-derived DOC moves into deeper soils. However, we do not know the quantity and the proportions of mineralization, retention and leaching during the process. Furthermore, most soil sorption experiments have lasted from several minutes to days, which is too short to assess DOC mineralization. Additionally, all these studies were of northern forest or grassland soils with relatively low precipitation, with little information available on subtropical forest soils with high annual precipitation. Studies that add 13C-labelled litter-derived DOC into soil columns during incubations offer substantial insight on the changes of quantity and chemistry of DOM that percolates downward through the soil profile. This approach also offers a means to quantify the energetic and metabolic potential of DOC for microbes.
When coupled with an isotopic tracer in a column experiment, spectroscopic methods offer additional metric to identify and quantify the chemical composition of DOM. Fluorescence excitation–emission matrices (EEMs) have been widely used as rapid and nondestructive analytical methods for monitoring DOM (Stedmon et al. 2003; Weishaar et al. 2003). Recently, parallel factor analysis (PARAFAC) has been developed to estimate a set of representative fluorescence components from EEMs (Stedmon and Bro 2008). Generally, DOM is separated into amino acids (free or bound in proteins), low molecular weight components, and high molecular weight components. Amino acids and low molecular components can be easily mineralized by microbial communities. High molecular weight components are generally considered to be less biodegradable than amino acids and low molecular weight components (Birdwell and Engel 2010; Fang et al. 2014; Fellman et al. 2010; Harun et al. 2016).
In this study, we applied 13C-labelled litter-derived DOC to different depth soil columns (10, 30 and 60 cm) in a 180-day incubation experiment to quantify the fate of 13C-labelled litter-derived DOC when they percolated through soils. The changes of litter-derived DOM composition were measured by combining EEMs and PARAFAC. We hypothesized that (1) litter-derived DOC was mainly retained on soil and mineralized by microorganisms when passing through soils; and (2) the changes in the composition of the litter-derived DOM would support the dynamic exchange model. This study will help us better understand the dynamics of DOC in subtropical forest soils and provide validation and insight on the dynamic exchange model.
Methods and materials
Soil collection
Soils were collected from a deciduous, humid, mountainous forest dominated by Fagus lucida in the Badagongshan National Nature Reserve, Hunan Province, in the mid-subtropical zone of China (29°46.04′N, 110°5.24′E). The climate is subtropical mountain humid monsoon with an annual average precipitation of 2105 mm and an annual mean temperature of 11.5 °C. The soil was classified as a Hapludalf with a silt loam texture. The soil consists of an A horizon (approximately 10–15 cm thick), a transitional horizon between the mineral A and B horizons (8–15 cm thick), and a B horizon (approximately 60 cm thick).We dug three trenches and used a shovel to collect soil samples at three fixed depths: 0–10 cm (topsoil), 10–30 cm (midsoil), and 30–60 cm (subsoil). All sampling plots were located in well protected national nature reserves, where vegetation and soil were representative for the given forest type. All sampling plots were located 100–400 m apart from each other. About 125 kg soil was collected at each plot. The topsoil, midsoil and subsoil were 35 kg, 50 kg, and 40 kg, respectively. Soil samples were immediately brought to the laboratory and passed through an 8-mm sieve. Roots, rocks and visible residues were removed manually. We manually mixed each sample with a shovel.
Isotopic labeling of saplings and the preparation of litter-derived DOC
We chose five adjacent Fagus lucida saplings that grew well and were less than 1 m in height in the Badagongshan National Nature Reserve. We sealed these saplings inside a 3 m long × 1 m wide × 1 m high polyethylene film chamber. 13CO2 which was produced by using citric acid and Na 132 CO3 (99 atom% 13C, SIGMA, USA) was delivered into the chamber from 11:00 am to 13:00 pm for ten sunny days in July, 2015. To ensure maximum assimilation of 13CO2 by plants, the 13CO2 concentration in the chamber was kept at approximately 500 ppm. Then, all fresh leaves from these saplings were picked manually, oven-dried, and chopped into small pieces (2–3 mm). Lab analysis indicated that the δ13C value of this litter was 230.5‰. We then obtained litter-derived DOC from the 13C-labelled leaves. For the extraction, we first added distilled water to the litter at a weight ratio of 1:15. Then, after 24 h at 20 °C, the solution was filtered through a 0.45-μm membrane, and distilled water was again added. This procedure was repeated three times. All extracts were combined to measure the concentration, the δ13C value and fluorescence excitation–emission matrices of litter-derived DOC. The concentration and the δ13C value of litter-derived DOC were 71.7 mg L−1 and 318.4‰, respectively.
Soil incubation
Sieved soils were poured into 10 cm diameter PVC tubes to form soil columns with thicknesses of 10, 30 and 60 cm (Fig. 1). The amount of soils for each layer was determined based on bulk density. We placed (1) 0.85 kg of topsoil; (2) 0.85 kg of topsoil and 2 kg of midsoil; (3) 0.85 kg of topsoil, 2 kg of midsoil and 3 kg of subsoil into the 10-cm, 30-cm, and 60-cm PVC tubes, respectively. The soils were individually placed in layers of 0–10 cm, 10–30 cm and 30–60 cm. The bottoms of the PVC tubes were covered with a nylon net to avoid soil loss while allowing liquid to pass through. Each soil layer was also separated with this nylon net. There were 36 replicate PVC tubes for each type of soil column.

Sketch of the soil columns for the incubation experiment
All PVC tubes were pre-incubated at 25 °C for two weeks. After pre-incubation, 13C-labelled litter-derived DOC (25.1 mg) was added drop-wise (5 ml min−1) into half of the PVC tubes by intravenous tubing, and the other half of the PVC tubes received distilled water as the control. During incubation, distilled water (450 ml) was added into each PVC tube every ten days. Each application corresponded to 58 mm of rain. This amount was determined according to the annual average precipitation of our study site. A 1-liter beaker was put at the bottom of each PVC tube for 24 h to collect soil leachate when we added distilled water or litter-derived DOC. The soil leachate was filtered through a 0.45-μm membrane and then was used to measure the DOC concentration (LDOC), δ13C value and the fluorescence characteristics.
The rate of soil C mineralization was measured daily for the first week of the incubation, twice a week for the following five weeks, and weekly for the remaining period of the experiment. At each measurement, three replicates were randomly chosen from each treatment to measure soil CO2 efflux rates. PVC tubes were first flushed with CO2-free air (air through a soda-lime column) for 1 h for the 10-cm PVC tubes, 2.5 h for the 30-cm PVC tubes, and 4 h for the 60-cm PVC tubes, respectively. Then the outflow was connected to an infrared gas analyzer (EGM-4, PP Systems, Amesbury, MA, USA) and a flow rate meter, and then CO2 concentration and the flow rate were recorded. Gas samples were collected in bags to measure 13C composition using cavity ringdown spectroscopy (Carbon Isotope Analyzer, 912-0003, LGR, CA, USA). Soil respiration from the midsoil (10–30 cm) was calculated as the difference between the 0–30 cm and 0–10 cm layers, and soil respiration from the subsoil (30–60 cm) was calculated as the difference between the 0–60 cm and 0–30 cm layers.
Lab analysis
A portion of the collected soil was immediately used to determine soil water content, WSOC and microbial biomass C. Another part of soil was air-dried and sieved (< 2 mm) to determine soil pH, texture, SOC, total N, δ13C, total free Fe oxides (Fed, crystalline plus non-crystalline), poorly crystalline Fe and Al oxides (Feo and Alo), and chelated organically complexed Fe and Al (Fepy and Alpy). The rest of the soils were stored in a refrigerator (< 4 °C) for no more than two weeks before lab incubation.
Three replicate PVC tubes of each treatment were destructively sampled at 10, 20, 30, 60, 90 and 180 days. Soil samples were then collected from each depth: 0–10 cm (topsoil), 10–30 cm (midsoil), and 30–60 cm (subsoil). One subsample of fresh soil was oven-dried to determine the SOC, total N, and soil δ13C. Another subsample of fresh soil was used to extract WSOC and microbial biomass C (MBC).
Soil pH was determined in 1:2.5 (weight: volume) soil solutions, and the soil water content was measured by oven-drying for 48 h at 105 °C. Soil texture was determined with a laser particle size analyzer (Mastersizer 3000, Malvern Panalytical, Malvern, UK). SOC, total N, and their δ13C values were then determined with an elemental analyzer (Flash 2000, Thermo Fisher Scientific, Waltham, MA, USA) interfaced with an isotope ratio mass spectrometer (Delta Plus Advantage, Thermo Finnigan, Bremen, Germany). Fe and Al were extracted by three selective dissolutions to determine the content of three types of mineral functional groups: total free Fe oxides (Fed), poorly crystalline Fe and Al oxides (Feo and Alo), and chelated organically complexed Fe and Al (Fepy and Alpy) (Pansu and Gautheyrou 2006). The concentration of Fed was measured using dithionite–citrate–bicarbonate method (DCB extraction). Feo and Alo were determined by extraction with acid ammonium oxalate at pH 3 in the dark (oxalate extraction). Fepy and Alpy were detected by sodium pyrophosphate extraction. After each of these extractions, the solutions were centrifuged, and the supernatant liquid was filtered through a 0.45 mm filterable membrane. Mineral concentration were then quantified by plasma atomic emission spectrometry (Optima 8000DV, PerkinElmer, Waltham, MA, USA). The crystalline and poorly crystalline metal oxides are characterized by high reactive specific surface area and have a high chemical capacity to establish covalent links with organic matter (Eusterhues et al. 2005; Leinemann et al. 2018). Therefore, higher values of Fed, Feo and Alo indicate stronger SOC stability through organic-mineral associations. The proportion of chelated organo-mineral complexes is given by pyrophosphate extractable Fe and Al (Mpy), and the SOC/Mpy is used to determine whether there is enough stock of Fe and Al ions in the soil for chelation (Masiello et al. 2004).
WSOC was extracted in a paste of 1:15 (weight: volume) of fresh soil and deionized water. The mixed paste was shaken for 5 h at 250 rpm at 25 °C. Subsequently, the supernatant liquid was filtered through a 0.45-μm membrane. Soil MBC was determined by a fumigation–extraction method (Vance et al. 1987). DOC and MBC in extracts were measured using a TOC Analyzer (vario TOC, Elementar, Langenselbold, Germany). To measure the δ13C values of soil leachates or extracts, 2 ml of a 0.5 mol L−1 K2SO4 solution was added to 20 ml soil leachates or extracts and then freeze-dried in order to obtain adequate amounts of dried leachates or extracts (Hagedorn et al. 2004). The δ13C values of leachates or extracts and the δ13C value of microbes were measured by an isotope ratio mass spectrometer (Delta-Plus, Thermo Finnigan, Waltham, MA, USA). Fluorescence excitation–emission spectroscopy of soil leachates or extracts were measured by fluorescence spectrophotometer (FP-6500, Jasco, Ashikaga, Japan). Bandwidths were set to 5 nm for both excitation and emission. A series of emission scans (280–550 nm) were collected over excitation wavelengths ranging from 230 to 400 nm (Stedmon et al. 2003).
Calculations and statistical analysis
We used mass balance and a two-component mixing model to determine the proportions (fDOC, fSOC) of DOC-derived and soil-derived CO2.
and
where fDOC and fSOC refer to the proportion of DOC-derived and SOC-derived CO2, respectively; and δDOC and δSOC refer to the isotope abundance (δ13C) of DOC and SOC, respectively (Phillips and Gregg 2003).
To calculate the amount of CO2–C derived from DOC and SOC under incubation, the following equations were used:
In the Eqs. (1) and (2), Ctotal (Ctotal = CDOC + CSOC) is the total amount of CO2–C released during incubation and δ13Ctotal is the corresponding isotopic value. CDOC is the amount of C derived from DOC and δ13CDOC is the isotopic value of DOC. CSOC is the amount of C derived from SOC and δ13CSOC is the isotopic value of SOC.
Repeated measures ANOVAs were used to assess the differences of δ13C values of soil, microbes, WSOC, LDOC, CO2, LDOC loss, cumulative CO2 emission, and concentrations of WSOC and LDOC among different soil layers at different sampling times. When differences were significant, we conducted post hoc comparisons using the least significant difference procedure. The significance level in all analyses was p < 0.05. All statistical analyses were performed using IBM SPSS statistics (v. 21, International Business Machines Corp., Armonk, NY, USA). Figures were drawn with OriginPro 2016 (OriginLab Corporation, Northampton, MA, USA). The parallel factor analysis (PARAFAC) was performed in MATLAB using the DOMfluor toolbox (Stedmon and Bro 2008).
Results
Soil properties at different soil layers
Physical and chemical properties differed by soil layer (Table 1). Soil pH was 4.6–4.9, soil δ13C was about −25‰, and both of the values increased slightly with soil depth. Soil water content was 62–95%, and it decreased with soil depth. Soil contained 20% clay, 60% silt and 20% sand. The proportion of clay in topsoil was higher than that in midsoil and subsoil. SOC, total N, MBC and WSOC content decreased with soil depth. The concentration of Fed was similar among soil layers, but the concentration of Feo + Alo increased with soil depth. The concentration of Fepy, Alpy, and the ratio of SOC/Mpy decreased with soil depth.
Mineralization and soil retention of DOC
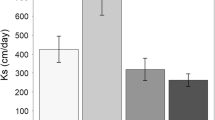
Cumulative CO2 emissions from SOC and DOC were significantly different among soil layers (Fig. 2, SOC: F = 612.7, P < 0.01; DOC: F = 1343.7, P < 0.01). Cumulative CO2 emissions from SOC followed the order of topsoil (4381 ± 146 mg C kg−1 soil) > subsoil (653 ± 297 mg C kg−1 soil) > midsoil (456 ± 286 mg C kg−1 soil), while CO2 emission from DOC followed the order of topsoil (11.55 ± 0.06 mg C kg−1 soil) > midsoil (1.37 ± 0.08 mg C kg−1 soil) > subsoil (0.66 ± 0.34 mg C kg−1 soil) (Table S1 in supplementary material).

Accumulated CO2 emission from SOC (a) and litter-derived DOC (b) during the incubation. Values are mean ± standard deviation (n = 3)
Soil δ13C values were comparable during the incubation, with a slight decrease near the end of the incubation (F = 2.27, P > 0.05, Fig. 3a). The δ13C values of soil microbes showed significant changes during the incubation (F = 12.24, P < 0.01), and the δ13C values at day 30 were significantly higher than at other times (Fig. 3b, Topsoil: F = 6.67, P < 0.01; Midsoil: F = 6.89, P < 0.01; Subsoil: F = 4.94, P < 0.05). The δ13C values increased in the midsoil earlier during the incubation (Fig. 3c, F = 13.44, P < 0.05). The δ13C values of CO2 significantly increased in the first 10 days of the experiment and then decreased (F = 13.28, P < 0.01, Fig. 3e).

The δ13C values of SOC (a), soil microbes (b), WSOC (c), LDOC (d) and CO2 (e) during the incubation. Values are mean ± standard deviation (n = 3)
The δ13C values of LDOC changed during the incubation (F = 755.3, P < 0.01). The topsoil LDOC δ13C values increased sharply and decreased to normal in the first 30 days. Meanwhile, the δ13C values of LDOC for the midsoil and subsoil increased slightly after the first addition of DOC and then gradually declined to initial values in the first 20 days of the incubation (Fig. 3d). The amounts of accumulated LDOC loss from different soil columns were different (F = 1473.1, P < 0.01) and followed the ranking of 0–10 cm (30.03 ± 1.55 mg C) > 0–30 cm (11.97 ± 0.22 mg C) > 0–60 cm (9.16 ± 0.42 mg C) (Fig. 4a). Approximately 21.2% of the added DOC was lost as LDOC (5.32 mg C) from the 0–10 cm soil column. But the 0–30 cm and 0–60 cm soil columns had little leaching losses of DOC (0.03 mg C and 0.01 mg C, respectively, Fig. 4b). Most leaching of DOC occurred immediately after the addition of litter-derived DOC. The loss of LDOC from all three soil columns increased gradually during the incubation.

LDOC (a) and LDOC from DOC (b) during the incubation. Values are mean ± standard deviation (n = 3)
The litter-derived DOC went into three forms of C: CO2, SOC and LDOC. We calculated proportions of DOC that went into the LDOC, CO2, and soil as well as the distribution of DOC at different soil layers. Our results indicated that 21.2, 0.13 and 0.04% of litter-derived DOC was leached from the 0–10 cm, 0–30 cm and 0–60 cm soil columns as LDOC, respectively. CO2 emission accounted for 18.4%, 23.0% and 26.4% of litter-derived DOC in the 0–10 cm, 0–30 cm and 0–60 cm soil columns, respectively. Therefore, 60.4%, 76.8% and 73.6% of litter-derived DOC was retained in the 0–10 cm, 0–30 cm and 0–60 cm soil columns, respectively (Table 2). Therefore, the proportions of litter-derived DOC that went into different soil layers were 60.4% (topsoil) and 16.5% (midsoil), respectively. We did not detect DOC retention in the subsoil. The proportions of litter-derived DOC mineralized into CO2 were 18.4% (topsoil), 4.6% (midsoil), and 3.4% (subsoil), respectively. Therefore, only 0.04% of litter-derived DOC was leached from the 0–60 cm soil column by the end of the experiment (Fig. 5).

The fate of litter-derived DOC at different soil depths. NA: not available, we did not detect DOC retention in the subsoil
Changes in DOC concentration and composition
The concentration of LDOC was highest after the first addition of distilled water or DOC and then it remained relatively steady (Fig. 6a). Only for the topsoil, there was a significant difference between the control and treatment after the first addition (P < 0.01). The WSOC concentration for the topsoil was significantly higher than those for the midsoil and subsoil (F = 791.89, P < 0.01, Fig. 6b). The concentrations in all soil depths increased with the first addition of distilled water or DOC and then decreased gradually.

The concentrations of LDOC (a) and WSOC (b) at different soil depths during the incubation. Values are mean ± standard deviation (n = 3)
The composition of DOC (litter-derived DOC, LDOC, and WSOC) was divided into four different fluorescent components using the PARAFAC model (Table 3; Fig. S1 in supplementary material). Component 1 was aromatic substances with high molecular weight (Excitation Max = 375 nm, Emission Max = 475 nm), component 2 was low molecular weight organic matter (Excitation Max = 325 nm, Emission Max = 400 nm), component 3 was high molecular weight organic matter (Excitation Max = 360 nm, Emission Max = 415 nm), and component 4 was amino acids, free or bound in proteins (Excitation Max = 280 nm, Emission Max = 360 nm). Components 1 and 3 were hydrophobic components, while components 2 and 4 were hydrophilic components. The proportions of hydrophobic components and hydrophilic components in litter-derived DOC were 54% and 46%, respectively.
Compared to litter-derived DOC, WSOC generally had a higher percentage of component 3, lower percentages of component 1 and 4 for all soil columns (Fig. 7a). In general, the relative proportions of all components were comparable between the control and treatment during the incubation. However, the relative proportions of component 1 and 4 in WSOC were quite different among different depths of soil.

Relative proportions of four different fluorescent components of WSOC (a) and LDOC (b) during the incubation. Initial = the relative proportions of four different fluorescent components in litter-derived DOC; C = control; T = treatment, soil columns with added DOC; 0–10 cm, 10–30 cm, 30–60 cm are topsoil, midsoil, and subsoil, respectively. Values are mean ± standard deviation (n = 3). # = a significant difference among different soil layers; * = a significant difference between initial and different layers of soil
The relative proportions of components in LDOC were also different from that of the litter-derived DOC. Compared to litter-derived DOC, LDOC in all the three soil columns had lower percentages of components 1 and 4, and higher percentages of components 3 (Fig. 7b). As soil column depth increased, the relative proportion of component 1 decreased while that of component 3 increased, which illustrated the composition of litter-derived DOC changed after passing through soil columns of different thickness. The relative proportions of all components did not differ between the control and treatment soil columns.
Discussion
The fate of litter-derived DOC
Previous incubation studies of different soils (including calcareous sand, acidic loam, organic soil and mineral soil) indicated that approximately 13–30% of added plant-derived DOC was mineralized into CO2 by the end of the experiment, and the mineralization rate remained unchanged after days 6–30 (Hagedorn et al. 2004; Muller et al. 2009; Miao et al. 2017). In our study, we found that 26.3% of DOC was mineralized into CO2, most of which occurred in the topsoil. Mineralization of litter-derived DOC should be rapid because that DOM contains highly and rapidly degradable, low molecular weight compounds (Van Hees et al. 2005). The larger percentage of mineralization in the top layer was probably caused by greater DOC retention and greater microbial biomass than in midsoil and subsoil. Like other studies, which had different incubation times, soil types and amounts of plant-derived DOC from our study, we also found the overall mineralization proportions to be less than 30%. In topsoil and midsoil where available carbon (such as WSOC, Table 1) is relatively abundant, the mineralized portion of plant-derived DOC is only a small part of the DOC remaining in topsoil and midsoil. However, in the subsoil, most of the added DOC was mineralized by microbes. A recent study also found that the chemical structure of added compounds had a greater effect on the mineralization of SOM than the energy content (Di Lonardo et al. 2017).
The rate of CO2 emission from respired DOC was higher during early incubation (day 1 to 20) than in the later incubation stage. Then, the accumulation rate remained stable during the later incubation stage. This finding showed that some litter-derived DOC was quickly used by microbes in the early incubation; this information may improve biogeochemical models. It is worth mentioning that we found δ13C of litter-derived DOC to be higher than the δ13C of bulk litter, which may be caused by the uneven distribution of 13C among compounds and within plant tissue (Fahey et al. 2011). Thus it is possible that the 13C label was incorporated into leaves in forms that were more readily leached than overall C. Similarly, the DOC with elevated 13C content might be more mineralizable than the overall litter DOC. Consequently, CO2 emission from litter-derived DOC may be overestimated relative to SOC.
Seventy-six percent of litter-derived DOC was retained by soil when it percolated through the soil columns. Of that, 60% was retained by the topsoil, and 16% was retained by the midsoil in the 180-day incubation experiment. Other leaching studies that added various concentrations of DOC into different soils also showed that about 60–73%, 60–70%, and 72–85% of DOC remained in soil (Sanderman and Amundson, 2008a; Muller et al. 2009; Scott and Rothstein, 2014). The high proportion of retention was likely due to physico-chemical processes including adsorption to reactive minerals such as Fe oxides and clay minerals and co-precipitation with Al and Fe, or biological retention (Kaiser and Kalbitz 2012). Most researchers have confirmed that soil adsorption was the main process of DOC retention by soil, and the adsorptive capacities of soils were mainly affected by soil properties, such as texture, structure, soil C content and mineral composition (Kaiser and Zech 1998; Lilienfein et al. 2004). For example, high concentrations of Fe and Al oxides can facilitate sorption (Qualls et al. 2000; Kaiser and Zech 2000a; Kothawala et al. 2009). The soil Feo + Alo contents in our study were higher than many temperate and boreal soils, which might reflect a higher adsorption capacity (Hagedorn et al. 2015; Kothawala et al. 2009).
The added DOC could also be assimilated by microorganisms, with some of that C in living microbial biomass and some of some in nonliving biomass, which then may be adsorbed to mineral surfaces after microbial processing (Guo and Chorover 2003; Kalbitz et al. 2005; Mikutta et al. 2007). In general, even though we could estimate microbial biomass, we could not distinguish between immediate abiotic retention and adsorption of microbial residues after microbial immobilization (Sokol et al. 2019). Thus, we could only trace DOC immobilization as far as assimilation into microbial biomass. Published substrate use efficiencies indicated that per unit mineralized C from lignin and maize residues yields 0.3–1.2 units of microbial biomass at the first days, but this value reduces to less than 0.1 in a few weeks (Bahri et al. 2008; De Troyer et al. 2011; Fanin and Bertrand 2016; Manzoni et al. 2012). When applying those efficiencies to our study, biodegradation only accounted for less than 3% of the added litter-derived DOC. Thus, physico-chemical processes of sorption or precipitation are dominant for DOC retention.
Furthermore, the amount of leaching loss decreased with increasing soil depth. This is probably caused by the different amount of DOC that leached from layer to layer in the soil columns. Less DOC passed through the topsoil to the midsoil and subsoil. Through a 180-day incubation experiment for soil columns of different thickness, we determined where the added DOC went in the different soil layers. Our experiment illustrated that relatively little DOC is mineralized as compared to soil retention. Therefore, retention during percolation through the soil column was the main process, microbial mineralization was a secondary process, and leaching loss from soil was relatively minor.
DOC composition change supports the dynamic exchange model
DOC concentrations in forest soils were regulated by a series of physical, chemical and biological processes that retained, transformed, and released DOC (Kaiser and Kalbitz, 2012; Scott and Rothstein, 2014). Researchers have proposed a dynamic exchange model to explain the movement of DOC (Sanderman et al. 2008b; Kaiser and Kalbitz, 2012). Studies have emphasized that DOC in deeper mineral soil is not only the result of physico-chemical stripping during transport but also the result of microbial processing and subsequent release of organic matter previously sorbed in overlaying horizons. The key point of the dynamic exchange model is that deep soil DOC is more similar to SOC than litter-derived DOC. But this theory has not been adequately tested in situ or with soil column studies. We found that DOC concentration and composition changed as DOM percolated through soil columns. Soil retention and mineralization of DOC led to a decrease of DOC concentration in LDOC, and reduced the relative proportion of the hydrophobic fraction (such as component 1) which indicated the preferential sorption of aromatic compounds. Many field experiments have found similar results with respect to the behavior of the aromatic portion, demonstrating relevance of soil column experiments to field conditions (Dittman et al. 2007; Kaiser et al. 2004; Leinemann et al. 2018).
Our results showed that the composition of WSOC and LDOC at all soil depths were comparable between control and treatment, which indicated that DOC in the subsoil and leachate was similar to WSOC, but not litter-derived DOC. These results indirectly show that DOC exchanged with previously sorbed compounds that were present on Fe oxides or clay minerals rather than adsorbed directly by Fe oxides or clay minerals. This result supported the dynamic exchange model. Other studies have also suggested that DOC at lower depths was derived primarily from the mineral soil itself (De Troyer et al. 2011; Fröberg et al. 2006; Gregorich et al. 2000; John et al. 2003; Karltun et al. 2005). Some studies have shown DOM sorption to be positively related to the concentrations of Fe and Al oxides which have large specific surface areas and provide reactive binding surfaces for OM (Eusterhues et al. 2005). SOC/Mpy can be used to indicate the availability of these sites to bind OM. Complete bonding of OM to oxides is likely when SOC/Mpy = 2–10 (Berhe et al. 2012; Masiello et al. 2004). In our study, the ratio of SOC/Mpy in topsoil was higher than 10, which indicated insufficient Mpy to bind OM. Thus, we attribute high retention of litter-derived DOC in topsoil to the exchanging of litter-derived DOC with older and partly microbial products in soil (Kaiser and Kalbitz 2012).
Our study involved a short-term laboratory incubation to determine the fate of DOC in soil. Therefore, there are limitations associated with our approach. Firstly, we conducted our experiments on reconstructed soil columns, in which soil structure was destroyed and the soil physical properties were quite different from undisturbed natural forest soils (Hagedorn and Bundt 2002; Kaiser and Guggenberger 2005). Secondly, the duration of the experiment was about half a year, and the findings of the experiment are limited to a relatively short term (up to a year). The longer-term pattern and dynamics of soil carbon storage and release could be different. Thirdly, the addition rate of DOC was uniform in the lab experiment, which is quite different from the episodic timing and magnitude of rainfall events under field conditions (Kaiser and Guggenberger 2005). The lab experiment might increase the contact time between DOC and soil, thus increasing the retention of DOC by soil. It is also worth to mention that the water flow in the lab experiment was different from forest soils. Lateral flow is quite common during storm events in field conditions, which might cause DOC to be rapidly transported along hillslopes and bypass contact between DOC and deeper soil, resulting in low retention of DOC by soil. We restricted our study to vertical flow and did not test influences of lateral flow. Nonetheless, our experiment provides some insight on the behavior and fate of litter-derived DOC in soil columns for humid forest ecosystems, and provides a basis for future.
Conclusions
We experimented on soil columns to quantify the fate of litter-derived DOC in different soil layers. During the 180-day incubation, approximately 76% of litter-derived DOC was retained by soil, 26% was mineralized into CO2 by microorganisms and little DOC left (0.04%) the soil columns. Physico-chemical processes of sorption or precipitation are the dominant mechanisms for DOC retention. Proportions of DOC retention and mineralization in topsoil > midsoil > subsoil. Topsoil layers absorbed and retained most DOC, leaving little DOC to leach deeper. The composition of WSOC and LDOC were comparable between the treatment and control, which proved that previously degraded SOM was the major source of WSOC and LDOC, rather than the freshly added litter-derived DOC. These results demonstrate a near-complete DOC exchange process with soil C components when passing through soil columns, in support of the dynamic exchange concept. The findings of how much litter-derived DOC contributes to the soil carbon pool and their dynamics in soil columns shed new light on DOC dynamics in forest ecosystems and provide information that is needed to improve biogeochemical models.
References
Asano Y, Compton JE, Church MR (2006) Hydrologic flowpaths influence inorganic and organic nutrient leaching in a forest soil. Biogeochemistry 81:191–204. https://doi.org/10.1007/s10533-006-9036-4
Bahri H, Rasse DP, Rumpel C, Dignac MF, Bardoux G, Mariotti A (2008) Lignin degradation during a laboratory incubation followed by (13) C isotope analysis. Soil Biol Biochem 40:1916–1922. https://doi.org/10.1016/j.soilbio.2008.04.002
Berhe AA, Harden JW, Torn MS, Kleber M, Burton SD, Harte J (2012) Persistence of soil organic matter in eroding versus depositional landform positions. J Geophys Res-Biogeosci. https://doi.org/10.1029/2011jg001790
Birdwell JE, Engel AS (2010) Characterization of dissolved organic matter in cave and spring waters using UV–Vis absorbance and fluorescence spectroscopy. Org Geochem 41:270–280. https://doi.org/10.1016/j.orggeochem.2009.11.002
Coble PG (1996) Characterization of marine and terrestrial DOM in seawater using excitation emission matrix spectroscopy. Mar Chem 51:325–346. https://doi.org/10.1016/0304-4203(95)00062-3
De Troyer I, Amery F, Van Moorleghem C, Smolders E, Merckx R (2011) Tracing the source and fate of dissolved organic matter in soil after incorporation of a c-13 labelled residue: a batch incubation study. Soil Biol Biochem 43:513–519. https://doi.org/10.1016/j.soilbio.2010.11.016
Di Lonardo DP, De Boer W, Gunnewiek PJAK, Hannula SE, Van der Wal A (2017) Priming of soil organic matter: chemical structure of added compounds is more important than the energy content. Soil Biol Biochem 108:41–54. https://doi.org/10.1016/j.soilbio.2017.01.017
Dittman JA, Driscoll CT, Groffman PM, Fahey TJ (2007) Dynamics of nitrogen and dissolved organic carbon at the Hubbard Brook Experimental Forest. Ecology 88:1153–1166. https://doi.org/10.1890/06-0834
Eusterhues K, Rumpel C, Kogel-Knabner I (2005) Organo-mineral associations in sandy acid forest soils: importance of specific surface area, iron oxides and micropores. Eur J Soil Sci 56:753–763. https://doi.org/10.1111/j.1365-2389.2005.00710.x
Fahey TJ, Yavitt JB, Sherman RE, Groffman PM, Fisk MC, Maerz JC (2011) Transport of carbon and nitrogen between litter and soil organic matter in a Northern Hardwood Forest. Ecosystems 14:326–340. https://doi.org/10.1007/s10021-011-9414-1
Fang H et al (2014) Experimental nitrogen deposition alters the quantity and quality of soil dissolved organic carbon in an alpine meadow on the Qinghai-Tibetan Plateau. Appl Soil Ecol 81:1–11. https://doi.org/10.1016/j.apsoil.2014.04.007
Fanin N, Bertrand I (2016) Aboveground litter quality is a better predictor than belowground microbial communities when estimating carbon mineralization along a land-use gradient. Soil Biol Biochem 94:48–60. https://doi.org/10.1016/j.soilbio.2015.11.007
Fellman JB, Hood E, Spencer RGM (2010) Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: a review. Limnol Oceanogr 55:2452–2462. https://doi.org/10.4319/lo.2010.55.6.2452
Fröberg M, Dan B, Bo B, Bryant C, Mulder J (2006) Concentration and fluxes of dissolved organic carbon (DOC) in Three Norway Spruce stands along a climatic gradient in Sweden. Biogeochemistry 77:1–23. https://doi.org/10.1007/s10533-004-0564-5
Gregorich EG, Liang BC, Drury CF, Mackenzie AF, Mcgill WB (2000) Elucidation of the source and turnover of water soluble and microbial biomass carbon in agricultural soils. Soil Biol Biochem 32:581–587. https://doi.org/10.1016/s0038-0717(99)00146-7
Guo MX, Chorover J (2003) Transport and fractionation of dissolved organic matter in soil columns. Soil Sci 168:108–118. https://doi.org/10.1097/01.ss.0000055306.23789.65
Hagedorn F, Bundt M (2002) The age of preferential flow paths. Geoderma 108:119–132. https://doi.org/10.1016/s0016-7061(02)00129-5
Hagedorn F, Saurer M, Blaser P (2004) A 13C tracer study to identify the origin of dissolved organic carbon in forested mineral soils. Eur J Soil Sci 55:91–100. https://doi.org/10.1046/j.1365-2389.2003.00578.x
Hagedorn F, Bruderhofer N, Ferrari A, Niklaus PA (2015) Tracking litter-derived dissolved organic matter along a soil chronosequence using C-14 imaging: biodegradation, physico-chemical retention or preferential flow? Soil Biol Biochem 88:333–343. https://doi.org/10.1016/j.soilbio.2015.06.014
Harun S, Baker A, Bradley C, Pinay G (2016) Spatial and seasonal variations in the composition of dissolved organic matter in a tropical catchment: the Lower Kinabatangan River, Sabah, Malaysia. Environ Sci 18:137–150. https://doi.org/10.1039/c5em00462d
John B, Ludwig B, Flessa H (2003) Carbon dynamics determined by natural 13C abundance in microcosm experiments with soils from long-term maize and rye monocultures. Soil Biol Biochem 35:1193–1202. https://doi.org/10.1016/s0038-0717(03)00180-9
Kaiser K, Guggenberger G (2005) Storm flow flushing in a structured soil changes the composition of dissolved organic matter leached into the subsoil. Geoderma 127:177–187. https://doi.org/10.1016/j.geoderma.2004.12.009
Kaiser K, Kalbitz K (2012) Cycling downwards-dissolved organic matter in soils. Soil Biol Biochem 52:29–32. https://doi.org/10.1016/j.soilbio.2012.04.002
Kaiser K, Zech W (1998) Rates of dissolved organic matter release and sorption in forest soils. Soil Sci 163:714–725. https://doi.org/10.1097/00010694-199809000-00005
Kaiser K, Zech W (2000) Dissolved organic matter sorption by mineral constituents of subsoil clay fractions. J Plant Nutr Soil Sci 163:531–535. https://doi.org/10.1002/15222624(200010)163:5<531::AID-JPLN531>3.0.CO;2-N
Kaiser K, Guggenberger G, Haumaier L (2004) Changes in dissolved lignin-derived phenols, neutral sugars, uronic acids, and amino sugars with depth in forested Haplic Arenosols and Rendzic Leptosols. Biogeochemistry 70:135–151. https://doi.org/10.1023/b:biog.0000049340.77963.18
Kalbitz K, Solinger S, Park JH, Michalzik B, Matzner E (2000) Controls on the dynamics of dissolved organic matter in soils: a review. Soil Sci 165:277–304. https://doi.org/10.1097/00010694-200004000-00001
Kalbitz K, Schwesig D, Rethemeyer J, Matzner E (2005) Stabilization of dissolved organic matter by sorption to the mineral. Soil Biol Biochem 37:1319–1331. https://doi.org/10.1016/j.soilbio.2004.11.028
Karltun E, Harrison AF, Alriksson A, Bryant C, Garnettd MH, Olssona MT (2005) Old organic carbon in soil solution DOC after afforestation-evidence from 14C analysis. Geoderma 127:188–195. https://doi.org/10.1016/j.geoderma.2004.12.008
Kim EA, Nguyen HV, Oh HS, Hur J, Choi JH (2015) Influence of soil conditions on dissolved organic matter leached from forest and wetland soils: a controlled growth chamber study. Environ Sci Pollut R 23:5203–5213. https://doi.org/10.1007/s11356-015-5740-8
Kindler R et al (2011) Dissolved carbon leaching from soil is a crucial component of the net ecosystem carbon balance. Glob Chang Biol 17:1167–1185. https://doi.org/10.1111/j.1365-2486.2010.02282.x
Kothawala DN, Moore TR, Hendershot WH (2009) Soil properties controlling the adsorption of dissolved organic carbon to mineral soils. Soil Sci Soc Am J 73:1831–1842. https://doi.org/10.2136/sssaj2008.0254
Leinemann T et al (2018) Multiple exchange processes on mineral surfaces control the transport of dissolved organic matter through soil profiles. Soil Biol Biochem 118:79–90. https://doi.org/10.1016/j.soilbio.2017.12.006
Lilienfein J, Qualls RG, Uselman SM, Bridgham SD (2004) Adsorption of dissolved organic and inorganic phosphorus in soils of a weathering chronosequence. Soil Sci Soc Am J 68:620–628. https://doi.org/10.2136/sssaj2004.6200
Manzoni S, Taylor P, Richter A, Porporato A, Agren GI (2012) Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol 196:79–91. https://doi.org/10.1111/j.1469-8137.2012.04225.x
Masiello C, Chadwick O, Southon J, Torn M, Harden J (2004) Weathering controls on mechanisms of carbon storage in grassland soils. Glob Biogeochem Cycle 18:GB4023. https://doi.org/10.1029/2004GB002219
Miao S, Ye R, Qiao Y, Zhu-Barker X, Doane TA, Horwath WR (2017) The solubility of carbon inputs affects the priming of soil organic matter. Plant Soil 410:129–138. https://doi.org/10.1007/s11104-016-2991-1
Mikutta R, Mikutta C, Kalbitz K, Scheel T, Kaiser K, Jahn R (2007) Biodegradation of forest floor organic matter bound to minerals via different binding mechanisms. Geochim Cosmochim Acta 71:2569–2590. https://doi.org/10.1016/j.gca.2007.03.002
Moller A, Kaiser K, Guggenberger G (2005) Dissolved organic carbon and nitrogen in precipitation, throughfall, soil solution, and stream water of the tropical highlands in northern thailand. J Plant Nutr Soil Sci 168:649–659. https://doi.org/10.1002/jpln.200521804
Muller M, Alewell C, Hagedorn F (2009) Effective retention of litter-derived dissolved organic carbon in organic layers. Soil Biol Biochem 41:1066–1074. https://doi.org/10.1016/j.soilbio.2009.02.007
Pansu M, Gautheyrou J (2006) Mineralogical separation by selective dissolution. Handbook of soil analysis: mineralogical, organic and inorganic methods. Springer, New York, pp 167–219
Phillips DL, Gregg JW (2003) Source partitioning using stable isotopes: coping with too many sources. Oecologia 136:261–269. https://doi.org/10.1007/s00442-003-1218-3
Qiu Q, Wu L, Ouyang Z, Li B, Xu Y (2016) Different effects of plant-derived dissolved organic matter (DOM) and urea on the priming of soil organic carbon. Environ Sci-Proc Imp 18:330–341. https://doi.org/10.1039/C5EM00446B
Qualls RG, Haines BL, Swank WT, Tyler SW (2000) Soluble organic and inorganic nutrient fluxes in clearcut and mature deciduous forests. Soil Sci Soc Am J 64:1068–1077. https://doi.org/10.2136/sssaj2000.6431068x
Qualls RG, Haines BL, Swank WT, Tyler SW (2002) Retention of soluble organic nutrients by a forested ecosystem. Biogeochemistry 61:135–171. https://doi.org/10.1023/a:1020239112586
Sanderman J, Amundson R (2008) A comparative study of dissolved organic carbon transport and stabilization in California forest and grassland soils. Biogeochemistry 89:309–327. https://doi.org/10.1007/s10533-008-9221-8
Sanderman J, Baldock JA, Amundson R (2008) Dissolved organic carbon chemistry and dynamics in contrasting forest and grassland soils. Biogeochemistry 89:181–198. https://doi.org/10.2307/40343596
Scott EE, Rothstein DE (2014) The dynamic exchange of dissolved organic matter percolating through six diverse soils. Soil Biol Biochem 69:83–92. https://doi.org/10.1016/j.soilbio.2013.10.052
Seely B, Lajtha K, Salvucci GD (1998) Transformation and retention of nitrogen in a coastal forest ecosystem. Biogeochemistry 42:325–343. https://doi.org/10.2307/1469390
Sokol NW, Sanderman J, Bradford MA (2019) Pathways of mineral-associated soil organic matter formation: integrating the role of plant carbon source, chemistry, and point of entry. Glob Chang Biol 25:12–24. https://doi.org/10.1111/gcb.14482
Stedmon CA, Bro R (2008) Characterizing dissolved organic matter fluorescence with parallel factor analysis: a tutorial. Limnol Oceanogr 6:572–579. https://doi.org/10.4319/lom.2008.6.572
Stedmon CA, Markager S, Bro R (2003) Tracing dissolved organic matter in aquatic environments using a new approach to fluorescence spectroscopy. Mar Chem 82:239–254. https://doi.org/10.1016/S0304-4203(03)00072-0
Van Hees PAW, Jones DL, Finlay R, Godbold DL, Lundstomd US (2005) The carbon we do not see—the impact of low molecular weight compounds on carbon dynamics and respiration in forest soils: a review. Soil Biol Biochem 37:1–13. https://doi.org/10.1016/j.soilbio.2004.06.010
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707. https://doi.org/10.1016/0038-0717(87)90052-6
Weishaar JL, Aiken GR, Bergamaschi BA, Fram MS, Fujii R, Mopper K (2003) Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ Sci Technol 37:4702–4708. https://doi.org/10.1021/es030360x
Yano Y, McDowell WH, Aber JD (2000) Biodegradable dissolved organic carbon in forest soil solution and effects of chronic nitrogen deposition. Soil Biol Biochem 32:1743–1751. https://doi.org/10.1016/s0038-0717(00)00092-4
Acknowledgements
The present study was supported by the National Natural Sciences Foundation of China (31400463, 31470526). We thank Dr. Stephen Sebestyen and two anonymous reviewers for their detailed constructive comments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing financial interests.
Additional information
Responsible Editor: Stephen D. Sebestyen.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Wang, M., Tian, Q., Liao, C. et al. The fate of litter-derived dissolved organic carbon in forest soils: results from an incubation experiment. Biogeochemistry 144, 133–147 (2019). https://doi.org/10.1007/s10533-019-00576-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-019-00576-3