
Abstract
Global initiatives to reforest degraded areas have intensified in recent years, arising from advances in agricultural frontiers that mainly alter natural landscapes and suppress vegetation. However, little is known about the influence of agricultural landscapes on the re-establishment of groups that perform key ecosystem services in restoration areas, such as pollination and pest control. Using trap nest methodology, we aimed to evaluate how aspects of the landscape influence the abundance, richness, diversity, and composition of trap-nesting bees and wasps as well as their natural enemies in restored areas located within highly managed landscapes. Samplings were conducted monthly from August 2018 to August 2019 in nine reforested areas of Seasonal Semideciduous forest older than 15 years. We found evidence of a negative influence of monoculture area (i.e., surrounding soybean/corn agriculture) on the abundance of wasps and their natural enemies, indicating a preference for more heterogeneous landscapes, possibly related to the greater availability of prey and lesser edge effect in this type of landscape. Bee diversity indicates a negative relationship with increasing distance from the edge of reforestation to the edges of the nearest forest fragment. Our results also suggest that some bee and wasp species are more affected by habitat loss than other species in general, which was indicated by a change in community composition. We conclude that the nesting of solitary bees and wasps in reforested areas was influenced by characteristics of the surrounding landscape, which should be considered in restoration projects to maximize the effectiveness of ecological services such as pollination and predation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Insect biodiversity is declining and severely threatened worldwide (Sánchez-Bayo and Wyckhuys 2019; Brühl and Zaller 2019; Cardoso et al. 2020). Some of the main causes for this condition are the loss and simplification of natural habitats, mainly related to intensive land use on farms with high-impact local management (Hipólito et al. 2018), as well as to increase in the use of pesticides and fertilizers (Brühl and Zaller 2019). Factors on different spatial scales can intensify the erosion of local diversity; changes in the horizontal and vertical structure of the natural landscape (Flores et al. 2018; Fornoff et al. 2021), increased land use intensity (Flores et al. 2018), heterogeneity of the agricultural matrix (Coutinho et al. 2020), and reduced habitat diversity have been identified as a set of factors that lead to the decline in the diversity and abundance of global insects, including invertebrates such as bees and wasps, as well as other organisms (Hendrickx et al. 2007; Sánchez-Bayo and Wyckhuys 2019; Zattara and Aizen 2021).The advancement of these agricultural landscapes has caused almost irreversible changes to the spatial structure of the highly diversified and structured original landscape (Krebs et al. 1999), resulting in a global decline in pollinator diversity and richness (Baldock 2020).
In this context, the study and practice of ecological restoration as an activity to promote biodiversity has gained strength in recent decades, representing an important instrument to halt and mitigate the deterioration of ecosystem health (Borges et al. 2021). Due to the urgency and need for higher investment in ecological restoration projects, the United Nations (UN) has instituted the period 2021 to 2031 as the decade of ecosystem restoration (Waltham et al. 2020).
The greatest challenge of ecological restoration is to restore not only the productivity of an ecosystem but also the biodiversity and ecological interactions, which are essential for the functioning and self-regeneration of habitats degraded or totally devastated by human actions (Lamb and Gilmour 2003; Calmon et al. 2011). Despite efforts to achieve this goal, many restoration projects fail even when the physical environment has been restored (Jones and Davidson 2016). A relevant cause is the lack of structural complexity of the landscape and the specific characteristics required by the biota of each location, limiting and restricting key groups and their numerous ecosystem services (Peipoch et al. 2015; Tonietto and Larkin 2018) that are important for the maintenance of restored environments.
Pollination and predation are listed as the main health-sustaining services of natural and man-made terrestrial ecosystems among these ecosystem services (Kumar 2010). Bees are recognized as the main pollinators in wild and cultivated ecosystems (Tscharntke et al. 1998; Zattara and Aizen 2021), with more than 20,000 species cataloged (Ascher and Pickering 2021). Particularly in restored areas, the presence of pollinators such as bees is essential over time for the success of areas undergoing restoration in the initial stages, contributing to the persistence of plant communities (Montalvo et al. 1997; Holzschuh et al. 2009; Dixon 2009; Cariveau et al. 2020). Wasps play an important role in regulating the population of herbivorous insects in the ecosystem as they are predators of other invertebrates such as spiders, caterpillars, and cockroaches (Crivelato et al. 2017). Wasps also act as occasional pollinators (Sumner et al. 2018).
Many studies have recently evaluated the responses of communities of trap-nesting bees and wasps and their natural enemies to ecosystem modifications (Tscharntke et al. 1998; Taki et al. 2008; Rocha-Filho et al. 2020), including landscape simplification (Wilson et al. 2020). Bees and wasps that nest in pre-existing cavities spend most of their lives building and supplying their nests. They depend directly on the structure of the vegetation due to the need for natural cavities in tree hollows and the specific food resources to feed the immature and adult bees (Krombein 1967). This direct relationship with plant resources and the physical structure of the environment, together with the ease of standardized sampling through the trap nest technique, gives the group of cavity-nesting bees and wasps a character of bioindicators that can be used in the evaluation of areas undergoing restoration (Araújo et al. 2018a, b, 2020, 2021; Fornoff et al. 2021).
The surrounding landscape can have a great influence on the structure of pollinator communities (Zirbel et al. 2019; Sexton et al. 2021). The structural complexity of the habitat is a driver of species richness and diversity in natural environments (Loyola and Martins 2008, 2011). Changes in the horizontal (types of land cover) and vertical (stratification of vegetation) structure are often caused by the increase in high-impact agricultural borders (Kennedy et al. 2013; Rocha-Filho et al. 2017; Hipólito et al. 2018), leading to simplifications in the habitat structure (Steiner et al. 2003). This simplification of the habitat reduces diversity and alters the composition of the community of several taxonomic groups (Wilson et al. 2020), including cavity-nesting bees and wasps (Ebeling et al. 2012; Flores et al. 2018, 2019; Araújo et al. 2021).
Factors such as the size and shape of the restored areas (Araújo et al. 2018a), size of the vegetation cover area, and connectivity and diversity of the surrounding agricultural and urban matrix can positively influence the community of cavity-nesting bees and wasps (Holzschuh et al. 2010; Hadley and Betts 2012; Cariveau et al. 2020; Fornoff et al. 2021; Sexton et al. 2021). Restored areas positioned in matrices of intensive agricultural lands can serve as islands or corridors of habitat and sustain high levels of biodiversity (Araújo et al. 2018a, 2020, 2021). Despite the evident influence of fragmentation and effects of the landscape on the community of these bees and wasps and the importance of a diverse agricultural landscape for the conservation of natural environments (Hendrickx et al. 2007; Flores et al. 2018, 2019; Rocha-Filho et al. 2020), studies evaluating the effects of the configuration of these landscapes on the abundance, richness, and composition of this community in restored areas are relatively recent and scarce (Krewenka et al. 2011; Tonietto and Larkin 2018; Campbell et al. 2019; Araújo et al. 2018a, b, 2020, 2021; Fornoff et al. 2021; Sexton et al. 2021).
Standardized monitoring of the restoration of bee and wasp fauna in areas under restoration helps to understand how this community is influenced by the surrounding landscape. This knowledge is essential to mitigate and reverse the decline of these insects, contributing to the establishment of guidelines for the implementation of environmental conservation projects and the implementation and evaluation of areas under restoration (Chapin et al. 2000).
The current study aimed to evaluate whether patterns of the cavity-nesting bee and wasp community and their natural enemies sampled in reforestation areas are affected by aspects of the structure of landscapes as the size of areas under restoration, size of the vegetation cover area, and size of the surrounding agricultural (i.e., monoculture) area. We proposed the following hypotheses: there would be greater abundance, richness, and diversity of bees and wasps that nest in traps and their natural enemies in (i) larger reforested areas, (ii) with larger areas of forest fragments nearby, (iii) with decreases in the distance from the edge of reforestation to the edges of the nearest forest fragment, and (iv) with decreases in soybean/corn monoculture areas. We expected variation in the composition of bees and wasps with the presence of specialized taxa in less simplified landscapes, that is, with greater reforestation, a greater proportion of native vegetation, smaller areas of soybean/corn monoculture nearby, and more connected (shorter distances).
Materials and methods
Study areas
Samplings were carried out in nine reforestation areas (REFs) located on the banks of the Capivara Reservoir on the Paranapanema River (22° 47′ 45′ S, 51° 00′ 12′′ W), near the mouth of the Tibagi River basin, in the state of Paraná, southern Brazil (Table S1; Fig. 1).

Location of the studied reforestation areas (REFs) delimited by a circle with a radius of 2000 m. *The habitat area is represented by forest fragments + reforestation and others, outside the sampled buffers
Aiming to better evaluate the conditions of areas undergoing restoration, the present study selected reforested areas with similar characteristics in terms of age (between 15 and 18 years, implemented in the same event) (Table S1), methodology for planting seedlings and seeds, geography, climatic conditions, altitude (334–338 m), and relief (shallow) (Mendonça 2000).
The reservoir was formed in the 1970s with the construction of a hydroelectric plant between the municipalities of Porecatu (state of Paraná) and Taciba (state of São Paulo). The study region was previously covered by seasonal semideciduous forests (Scervino and Torezan 2015). All reforestations were implemented on the margins of the Capivara Reservoir, starting in 2002. The manual planting of seedling “cocktails” was adopted with 3 × 2 m spacing, using 40 to 50 plant species native to the Tibagi River basin with a high proportion of early pioneer (ex. Aegiphilla sellowiana, Guazuma ulmifolia, Trema micrantha, and Schinus terebinthifolius) and secondary species (ex. Cordia tichotoma, Gallesia integrifolia, Peltophorum dubium), without fertilization and with manual and mechanized maintenance for the rapid control of grasses (Cavalheiro et al. 2002).
The studied reforestation areas are in little diversified landscapes with a predominant matrix of rotating soybean (Glycine. max (L.) Merril) and corn (Zea mays L.) monoculture (Scervino and Torezan 2015). The landscape has been drastically altered since 1920 due to intensive agricultural and urban activities. Today only 2% of the original vegetation remains, represented by a mosaic of small fragments and remnants of seasonal semideciduous forests in different degrees of conservation, primarily in secondary successional stages and suffering from logging and poaching (Scervino and Torezan 2015).
According to the Köppen classification, the region’s climate is Cfa, humid subtropical (Mesothermal), a marginal tropical zone with a hot summer and a mean temperature during the hottest month of around 22 °C. In winter, the area has a dry climate with temperatures below 18 °C in the coldest month, and rains are distributed throughout the year with a mean rainfall index between 1400 and 1600 mm/year (Mendonça 2000).
Samplings of bees, wasps, and their natural enemies
For the sampling of cavity-nesting bees and wasps and their natural enemies, we used two types of trap nests (Krombein 1967; Taki et al. 2008): (a) bamboo canes closed at one end by the node, with varying lengths and diameters, ranging from 8.0 to 30.0 cm and 0.5 to 3 cm, respectively, and grouped in bundles of 50 canes; (b) black cardboard tubes closed at one end, 8 cm long and 0.8 and 1.0 cm in diameter, inserted into a wooden block, 20 × 20 × 5 cm, with a capacity for 82 cardboard tubes per block (41 of each diameter). The two types of trap nests were made available at the sampling points on individual wooden platforms with an approximate height of 1.5 m and covered with plastic protection (transparent PET tile). Two wooden blocks (164 tubes) and two bamboo bundles (100 bamboo canes) were placed on each platform, exposed to opposite sides, totaling 264 trap nests per platform. This sampling method ensures the capture of reproductively active and resident species, excluding those that are just transiting through the reforestation areas (Morato and Martins 2006).
A 500 m transect was implemented in each reforestation area (REF), with five sampling points, 100 m equidistant from each other, and making a total of five sets of traps per REF. At each sampling point, a platform was installed with the set of traps, using the same number of trap nests, with equivalent dimensions, totaling 264 trap nests, 1320 cavities per reforestation area, and 11880 cavities considering the set of nine REFs. All platforms were installed from 10 to 20 m from the edge of the REFs, but they suffer from the evident edge effect due to the corridor shape of all reforestations.
Trap nests were sampled monthly from August 2018 to August 2019, totaling 13 samples in each area. An otoscope was used for the inspection of traps in the field. The occupied trap nests were removed, identified, capped at one end with a test tube and taken to the laboratory. Each trap nest removed was immediately replaced by another of equivalent size. In the laboratory, the nests were monitored daily to record emerging individuals. A few months after collection, the nests were opened, photographed, and inspected for non-emerged individuals, adults or in larval form, usually dead. The nests were classified according to their characteristics such as architecture, building materials, number of cells, and available food. The individuals found alive were killed in ethyl acetate or in a freezer, mounted on pins, labeled, dried in an oven, and stored in entomological drawers. The specimens were identified to the lowest possible taxonomic level, based on dichotomous keys for identification and reference material. The identification of nests with only non-emerged individuals was performed by comparing the characteristics with similar nests and with reference material. Mixed nests, where two or more species nested at the same time, were separated in the data analysis. All the material collected in this study is deposited in the Collection of the Laboratory of Animal Genetics and Ecology of the State University of Londrina. Subsequently, part of this material will be deposited at the Zoology Museum of the State University of Londrina.
Landscape variables
Images extracted from a LANDSAT 8, OLI sensor (Orbit/point: 222/076) with a resolution of 30 m and passing date of 10/22/2018 were used to obtain the landscape metrics. Initially, we defined a circular buffer 2000 m in radius, starting from the central point of the sampling transect (sampling point 3) of each focal reforestation (Fig. 1). This buffer pattern was chosen according to the flight radius and foraging of medium to large-sized bee species, the most commonly found in this study (Greenleaf et al. 2007; Zurbuchen 2010; Pokorny et al. 2015). In the buffers, we classified the images in the following themes based on supervised classification using the Maximum Likelihood Classification algorithm: water, soybean/corn monoculture matrix, reforestation, forest fragment, and urban areas. After classification, we described and estimated the following landscape variables through the polygon areas: (a) Reforestation area (ha), (b) Forest fragment area (ha), and (c) Soybean/corn monoculture area (ha). Within the buffers analyzed, areas of exposed soil were classified as a soybean/corn monoculture matrix area. Flood areas in the process of restoration at the edges of the reforestation areas were classified as reforestation areas. They showed a reflected electromagnetic radiation shine similar to reforestation (Moskalenko 2016) and can be considered areas of extreme importance for the foraging of bees and wasps due to the presence of ruderal plants (Coutinho et al. 2020). All forest fragments found in the analyzed buffers were characterized as Seasonal Semideciduous Forest in the secondary successional stage, with areas of clearings and tangles of vines. All images generated from the supervised classification showed high overall accuracy and Kuiper indices (> 0.75).
To quantify the degree of isolation of the focal reforestations, we used the distances (m) between the focal reforestations and the nearest forest fragment, estimated from the edge closest to the focal reforestations to the edge of the nearest forest fragment (Bender et al. 2003).
All metrics and classifications were calculated using the software QGIS v. 3.14 (QGis Development Team 2020).
Data analysis
To determine the abundance of cavity-nesting bees and wasps, the number of established nests was counted, whether these nests emerged or not. To determine the abundance of natural enemies, the number of parasitized nests was counted, whether the individuals emerged or not. This abundance pattern represents the nesting rate, that is, the number of nests found, and parasitism (= number of nests attacked by natural enemies) of each species.
To compare the richness of bees, wasps, and enemy species of each REF and to evaluate the sampling efficiency, we used integrated rarefaction/extrapolation curves based on the “Hill numbers” approach with asymptotic analysis, q = 0 (Real richness) and a confidence interval of 0.95, calculated using the iNEXT package in the R 3.2.3 program (Chao 2014; R Core Team 2020) and plotted using the ggplot2 package (R Core Team 2020).
The diversity of bees, wasps, and natural enemies of each REF was calculated using the Shannon-Wiener index (H′), considering the abundance of nests collected and using the Diversity function of the Vegan package in the program R 3.2.3 (R Core Team 2020).
Generalized linear models (GLMs) were adjusted to evaluate the influence of landscape characteristics on the abundance, richness, and diversity of bees, wasps, and natural enemies, which constitute the response variables. For this, each response variable was related to the explanatory variables of the landscape, consisting of reforestation area, forest fragment area, soybean/corn monoculture area, and distance. These landscape descriptors were selected since they did not present a significant correlation between them. It is important to note that the water areas were classified but not included in the analysis, as it was negatively correlated with the explanatory variable soybean/corn monoculture matrix. To evaluate the abundance of bees, wasps, and natural enemies, generalized linear models with binomial error distribution were used due to the super-dispersion of the data. For the richness of bees, wasps, and natural enemies, generalized linear models with Poisson error distribution were used for counting data. For the diversity of bees, wasps, and natural enemies, generalized linear models with Gaussian error distribution were used. For all response variables, candidate models were classified using the corrected Akaike information criterion for small samples (AICc) (Akaike 1974). For each response variable, 16 competing models were used to explain the patterns, including a null model representing the lack of effect. Only additive models were used in these analyses. Subsequently, the corrected Akaike Information criterion for small samples was estimated, the ΔAICc (difference between the AICc of each model concerning the best model) and the Akaike weight (wAIC) (the probability of a given model being the best among a set of competing models) (Johnson and Omland 2004). Models with ΔAICc < 2.0 and wAIC > 0.1 were considered equally plausible to explain the patterns observed (Burnham and Anderson 2003). The models selected by the Akaike information criterion (AIC) were performed with the MuMIn package (Bartón 2016) and the model diagnostics were performed with the RT4Bio package (Reis Jr et al. 2015) in the R 3.2.3 program (R Core Team 2020).
Distance-Based Redundancy Analysis - dbRDA (Legendre and Anderson 1999) using the Bray–Curtis index was carried out to investigate the influence of landscape characteristics on the composition of bees, wasps, and natural enemies. For this analysis, five sampling units per REF were considered, referring to the five sampling points of each area. The composition data were previously standardized to minimize the effects of sampling units with a very high abundance. The abundance of nests of each species was divided by the total abundance of nests in each sampling unit. This procedure was performed separately for each studied group. The distance matrices of the composition of bees, wasps, and natural enemies were used as the response variables and the predictor matrix comprised the landscape characteristics (reforestation area, forest fragment area, soybean/corn monoculture area, and distance between reforestation area and forest area). For each group, “ANOVA-like” permutation tests (Legendre and Legendre 1998) were performed to assess the statistical significance of the entire model, of the two first axes, and the association between species composition and the variables selected. Finally, the dbRDA results were plotted to visualize the relationship between the species and the landscape characteristics. The dbRDA was performed using the vegan package (Oksanen et al. 2018) in R 3.2.3 (R Core Team 2020).
Results
A total of 1000 nests were identified in the nine reforestation areas evaluated during the study. Of these, 101 were bee nests and 898 were wasp nests (Table S2). The sampled bees belong to 10 morpho-species from two families, Apidae and Megachilidae, while the wasps belong to 16 morpho-species from four families: Crabronidae, Pompilidae, Sphecidae, and Vespidae (Table S2). The abundances of wasp species that hunt spiders (58.66%) and of species that hunt lepidopteran larvae (41.38%) were close.
Eighteen morpho-species of natural enemies were recorded in 218 parasitized nests, from which 11 were classified as parasitoids (Bombyliidae, Chrysididae, Eulophidae, Ichneumonidae, Leucospidae, Ripiphoridae and Sarcophagidae) and seven as kleptoparasites (Apidae, Chrysididae, Megachilidae). The parasitoids attacked 11 host species, and Auplopus sp. 1 was the wasp species with the most parasitoids (7 morpho-species) (Table S3).
The highest values of bee diversity were found in REF 8 and REF 4, areas with large and nearby forest fragments (Tables S1 and S2). The lowest bee diversity value was found in REF 5 (Table S2). The diversity of wasps and natural enemies was relatively high in all sampled areas (Table S2). The greatest diversity of wasps found was in REF 4 and the lowest in REF 8 (Table S2). The greatest diversity of natural enemies was also found in REF 4 and the lowest diversity of natural enemies was found in REF 9 and REF 7 (Table S3).
The rarefaction curves of bees did not present stabilization (Fig. 2). The areas with a greater abundance of nests (REF7) and greater richness of bees (REF2, REF4, and REF8) indicated stabilization in the rarefaction curve, around 50 nests collected and 8 to 15 expected species. The REF5 reforestation area presented the lowest species richness with only the occurrence of one recorded bee species. The extrapolation curves of bees in less favored reforestation areas (REF3, REF5, and REF6), that is, small reforestation areas with few forest fragment areas nearby and large areas of surrounding monoculture, indicated stabilization with relatively low richness, around 25 sampled nests and 1 to 4 expected species (Fig. 2).

Rarefaction curves based on the number of bee, wasp, and natural enemy species, by number of nests, sampled from August 2018 to August 2019 in nine reforestation areas of seasonal semideciduous forest, located in the state of Paraná, southern Brazil
Wasp rarefaction curves indicated sampling efficiency in most reforestation areas and a pattern in the number of species, around 100 nests collected and 8 to 15 expected species (Fig. 2). The largest number of wasp nests was found in the second largest reforestation area (REF8), indicating stabilization of the curve and confirming the sampling efficiency for the group of wasps that nest in the areas (Fig. 2).
The natural enemies indicated stabilization of the rarefaction curve in only a few areas (REF1, REF7, REF8, and REF9). The areas with the greatest abundance of nests attacked by natural enemies indicated stability in the curves from 25 nests collected and from 5 to 12 expected natural enemy species (Fig. 2). Reforestation areas with a smaller area (REF1) and those with an absence of a nearby forest fragment (REF5) indicated an expected richness of parasite species greater than that of wasps and bees (Fig. 2).
We observed the replacement of the sampled species of bees, wasps, and natural enemies along the gradients of the reforestation areas (Fig. 3a), forest fragment areas (Fig. 3b), and soybean/corn monoculture areas (Fig. 3c). Bees, wasps, and natural enemies presented species that only occurred in more integral regions, that is, with greater areas of reforestation (Tetrapedia diversipes, Podium sp. 1, Caenochrysis crotonis and Ichneumonidae sp. 2) (Fig. 3a), larger areas of forest fragments (Euglossa townsendi, Eufriesea aff. auriceps, Megachile guaranitica, Minixi sp. 1, Chrysis intricata, Caenochrysis crotonis, Ichneumonidae sp. 2, Priochilus sp. 1) (Fig. 3b), and smaller areas of soybean/corn monoculture (Priochilus sp 1, Zethus sp. 2, Euglossa townsendi, Eufriesea aff. auriceps, Ichneumonidae sp. 2, Caenochrysis crotonis and Tetrapedia diversipes) (Fig. 3c). Regarding the monoculture area gradient, a group of species with a much higher relative abundance was observed in regions with smaller monoculture areas (Fig. 3c). All orchid bee species occurred in reforestation areas with nearby forest fragments and with less monoculture areas (REF1, REF2, REF4, and REF8).

Relative abundance of nests of species of bees (green), wasps (red), and their natural enemies (blue), collected from August 2018 to August 2019 in nine reforestation areas in northern Paraná, according to the: a size of the reforestation area (ha); b size of the forest fragments (ha); c size of the soybean/corn monoculture area (ha), evaluated within the 2000 m radius
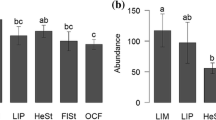
The models that best explained the abundance of wasps and natural enemies included only the monoculture matrix area as an explanatory variable through a negative relationship. This indicates that the abundance of wasps and natural enemies decreased with the increase in the monoculture matrix area (Tables 1 and S4; Fig. 4). The abundance of bees did not vary according to any of the explanatory variables used (Table 1 and S4). While the diversity of wasps and natural enemies did not change due to explanatory variables (Tables 1 and S4), the diversity of bees was explained by the model that indicates distance as an explanatory variable, through a negative relationship. This indicates that bee diversity decreases with increasing distance between the reforestation area and the nearest forest area. A second plausible model includes only the size of the forest area, indicating that bee diversity increases directly with this variable (Tables 1 and S4). No relationship was found between the different variables and any of the three analyzed groups regarding species richness (Tables 1 and S4). For bees, the first and second axes of dbRDA explained 51.0% (pseudoF = 3.21, P = 0.15) and 34.9% (pseudoF = 2.21, P = 0.31) of dissimilarity in species composition, respectively. The permutation test for the entire model revealed a marginally significant association between species composition and the explanatory variables of landscape characteristics (pseudoF = 1.59, P = 0.06). Reforestation area had a marginally significant effect on the species composition (pseudoF = 2.03, P = 0.06). The other landscape characteristics had no significant effects: forest fragment area (pseudoF = 0.95, P = 0.44), soybean/corn monoculture area (pseudoF = 1.69, P = 0.11), and distance between reforestation area and forest area (pseudoF = 1.66, P = 1.44). They explained together 19.9% of the total variation in the data set. Carloticola paraguayensis and Euglossa pleosticta were the species most associated with larger reforestation areas (Fig. 5).

Relationship between response variables of the studied groups (bees, wasps, and natural enemies) and explanatory variables of the landscape: A influence of the monoculture area on the abundance of wasps; B influence of the monoculture area on the abundance of natural enemies; C influence of distance on the Shannon (H′) diversity of bees

dbRDA of species composition of the bees (A), wasps (B) and natural enemis (C) collected from August 2018 to August 2019 in nine reforestation areas in northern Paraná whith biplot of explanatory variables of the landscape
For wasps, the first and second axes of dbRDA explained 73.9% (pseudoF = 9.61, P < 0.001) and 15.8% (pseudoF = 2.06, P = 0.23) of dissimilarity in species composition, respectively. The permutation test for the entire model revealed a significant association between species composition and the explanatory variables of landscape characteristics (pseudoF = 3.25, P < 0.001). Reforestation area (pseudoF = 7.77, P < 0.001) and forest area (pseudoF = 2.95, P < 0.005) had a significant effect on the species composition (pseudoF = 2.03, P = 0.06). Soybean/corn monoculture area (pseudoF = 1.19, P = 0.28) and distance between reforestation area and forest area (pseudoF = 1.10, P = 0.34) had no significant effects. They explained together 29.0% of the total variation in the data set. Auplopus sp. 1 was the species most associated with larger reforestation areas, and Priochilus captivum and Pachodynerus guadulpensis were the species most associated with smaller reforestation areas. Pachodinerus grandis was the species most associated with larger forest areas (Fig. 5).
For natural enemies, the first and second axes of dbRDA explained 48.3% (pseudoF = 2.69, P = 0.20) and 24.3% (pseudoF = 1.35, P = 0.72) of dissimilarity in species composition, respectively. The permutation test for the entire model revealed a marginally significant association between species composition and the explanatory variables of landscape characteristics (pseudoF = 1.39, P = 0.08). Reforestation area had a significant effect on the species composition (pseudoF = 2.41, P = 0.02). The other landscape characteristics had no significant effects: forest fragment area (pseudoF = 0.59, P = 0.80), soybean/corn monoculture area (pseudoF = 1.34, P = 0.20), and distance between reforestation area and forest area (pseudoF = 1.20, P = 0.25). They explained together 14.5% of the total variation in the data set. Ichneumonidae sp. 1 and Macrosiagon sp. 1 were the species most associated with larger reforestation areas, and Melittobia sp. 1 and Chrysis intrincata were the species most associated with smaller reforestation areas. (Fig. 5).
Discussion
Our results revealed that aspects of the landscape differently affected the populations of cavity-nesting bees and wasps in reforestation areas, as well as affecting the community of natural enemies of these insects. We detected a negative effect of the amount of soybean/corn monoculture on the abundance of wasp nests and on the abundance of nests attacked by natural enemies sampled in the reforestation areas evaluated. Contrary to our expectations, the richness of bee, wasp, and natural enemy species did not present any variation due to the metrics of the landscape evaluated. Bee diversity was the only variable affected by a landscape factor, being negatively correlated with the increasing distance from the nearest forest fragment and positively correlated with the size of nearby forest fragments. We found an influence of the size of the reforestation areas regarding species composition, altering the composition of bees, wasps, and natural enemies. The wasp composition also varied depending on the area size of the nearby forest fragments. The results obtained only partially confirm the hypotheses raised, indicating that bees, wasps, and their natural enemies sampled in reforestation areas respond differently to variations in the landscape.
The lower abundance of wasps observed in areas with a larger area of monoculture is possibly related to the reduced diversification of the studied agricultural landscapes. These intensely simplified ecosystems are hostile to the majority of species, being favorable only to a few species that adapt well to the dominant condition of low environmental heterogeneity (Hendrickx et al. 2007; Torné-Noguera et al. 2020; Rocha-Filho et al. 2017; Araújo et al. 2018a, 2021). Generally, plants grown in the agricultural matrix do not offer the food resources used by adult wasps (nectar) or only offer it at certain times of the year, through intense flowering and for a short period (Silveira 2004; Hipólito et al. 2018). This resource limitation restricts foraging in areas adjacent to cultivated fields, such as areas under restoration, which in turn suffer from the recurrent impacts of fragmentation caused by agricultural matrices (Rocha-Filho et al. 2017). These matrices of conventional agriculture also interfere negatively with the supply of solitary wasp nests due to the extinction or drastic reduction in species of forage plants and old woody plants that provide preexisting cavities (Holzschuh et al. 2009, 2010; Flores et al. 2019; Araújo et al. 2021). A study found a positive correlation between wasp abundance and crop diversification in family farming areas due to the increase in the variety of agricultural crops introduced in the landscape (Coutinho et al. 2020). It is important to remember that in this study the explanatory variable soybean/corn monoculture matrix was negatively correlated with the water area, which also tends to result in a simplification of habitats for wasps and bees (Flores et al. 2018). In this context, it is important that future studies consider and discuss the effects of the water area on these communities.
Other factors have also been identified as having an impact on the abundance in wasp populations. Land use has been recognized as one of these factors, since recordings of some rarer wasp species have occurred with greater abundance in more “natural” environments, such as fallow land, reforestation of native species, and forest interiors (Matos et al. 2013; Rocha-Filho et al. 2017; Araújo et al. 2021). The authors reported a significant contribution from diversified and organic crops to the maintenance of wasp species by providing resources for feeding and nesting for their populations (Matos et al. 2013). The increase in the cultivated area with the monoculture of grains tends to hinder the colonization of reforestation by species from fragments of native vegetation nearby, which are unable to move through the highly homogeneous matrix. For example, wasps of the subfamily Eumeninae avoid crossing conventionally managed cereal fields, needing strips of fallow fields or organic cultivation fields to move between the native vegetation in the landscape (Hendrickx et al. 2007; Holzschuh et al. 2009, 2010). In young regrowth forests in subtropical South-East China, the abundance of cavity-nesting wasps increased with increasing canopy cover and the subsequent microclimatic alteration brought about by this change (Fornoff et al. 2021).
The effects of the proportion of monoculture area on the community of natural enemies are related to the effect of this landscape variable on the host (wasps). More than 90% of the collected sampled natural enemies were found in wasp nests (Spearman’s Correlation statistic: rs = 0.903; p-value < 0.0005). When monoculture affects the abundance of host wasps, it directly affects the abundance of their natural enemies through ecological interaction between related trophic niches (Veddeler et al. 2005). Recent studies corroborate this statement since there was no direct correlation between the abundance and richness of the parasites and any of the landscape structure variables, but a direct relationship with the hosts was suggested (Holzschuh et al. 2010; Araújo et al. 2018b; Staab et al. 2018). Fornoff et al. (2021) found a direct relationship between the abundance of the “herbivore hunting wasps” community with the abundance of their natural enemies.
The lack of influence of the monoculture area on the abundance of bee nests collected in the reforestation areas was not expected. Thus, the abundance of bees could be influenced on a finer scale by the landscape related to structural complexity factors of the vegetation of the evaluated reforestations, such as plant species richness, height, the density of tree species, canopy cover, and basal area of the trees (a proxy for tree biomass) around sampling points (Steffan-Dewenter et al. 2002; Loyola and Martins 2008, 2011; Araújo et al. 2018a; Coutinho et al. 2020; Fornoff et al. 2021). The surrounding vegetation did not influence the abundance of bees in recovering fallow areas within intensive farming matrices in northern Florida (Campbell et al. 2017). The authors suggested that the availability of preexisting cavities is the only factor that influences the abundance of bees in areas of intensive agriculture. This results from factors often absent from the landscape, such as pre-existing cavities, floral sources, among others.
The fact that the richness of bees, wasps, and natural enemies was not related to any of the landscape variables evaluated could again be explained by the spatial scale adopted in this study. Coutinho et al. (2020) found a positive correlation between the richness of bees and wasps and the size of the surrounding agricultural area considering wide spatial scales (above 2000 m buffer). However, this relationship was observed only in areas with an extremely diversified family farming matrix or surrounded by more structurally diverse environments, such as areas of native vegetation. In homogeneous landscapes, factors related to the structural complexity of local vegetation such as the richness of plant species and the height and density of trees may be more important in determining the richness of bees and wasps than general aspects of the landscape such as the size of the agricultural area (Tscharntke et al. 1998; Araújo et al. 2018a, 2021).
In agreement with the findings of the current study, the diversity of bees is often positively related to the diversity and vegetation cover of a landscape (Staab et al. 2018; Flores et al. 2019; Araújo et al. 2021; Fornoff et al. 2021). Large areas of continuous forest or more connected areas showed higher indices of bee diversity than small and poorly preserved forest fragments (Morato and Campos 2000). This results from the great heterogeneity of these environments, providing a higher number of resources both for nesting (greater abundance of old and decomposing trees, providing a greater number of nesting sites) and for foraging and provisioning of nests (greater abundance and diversity of resources plants), favoring the increase in bee diversity indices (Morato and Martins 2006; Fabian et al. 2014; Sexton et al. 2021). According to hypotheses derived from the theories of islands and metapopulations, larger forest fragments inserted in agricultural matrixes of great impact may be serving as a “source”, providing greater abundance and species richness for the studied reforestation areas (“sink” areas). These areas would contribute to the increase in the diversity of bees in patches of adjacent favorable vegetation (Bush 2000), as with areas undergoing restoration. The distance from the “source” can also influence the colonization of new areas (Bush 2000), which corroborates what was found in our study where the diversity of bees was greater in restored areas closer to forest fragments.
The fact that the composition of wasps sampled in reforestation areas varied according to the size of the nearby forest fragment areas is also supported by the theories of island and metacommunities. Larger forest areas would provide greater abundance and species richness, influencing the composition of the communities found in adjacent restoration areas (Steckel et al. 2014; Staab et al. 2018). Decreasing the size of forest fragments can also increase interspecific competition (Hadley and Betts 2012), change pollinator/plant network patterns (Hagen et al. 2012; Hadley and Betts 2012), increase local extinctions, and decrease the quality and quantity of resources offered to wasps (Hadley and Betts 2012). These alterations in environmental dynamics resulting from the fragmentation of native habitat favor some species adapted to edges and degraded environments and restrict other more specialized species (Stangler et al. 2015; Oliveira and Gonçalves 2017; Rocha-Filho et al. 2017). More specialized species have a preference for larger and more preserved fragments (Morato and Campos 2000; Araújo et al. 2018a, 2021) and thus are only found in larger reforestation areas with larger adjacent forested areas. With this evidence, we emphasize the importance of preserving forest fragments and conservation units to maintain the diversity of wasp and bee communities. Recent studies on a gradient of restoration in the Amazon have shown greater diversity of bees and wasps nesting in “mixed reforestations” (with native species planting) and in areas of secondary forest, indicating that the abundance, richness, and diversity of bees and wasps is greater in “intermediate” environments. This suggests that the variation in bee abundance and species richness along the gradient may reflect variation in food resource availability and variations in habitat structure (Araújo et al. 2021).
Wasps present a great capacity to colonize disturbed environments since they find greater prey abundance and a large quantity of ruderal plants that serve as a source of food. This seems to be the most likely factor that explains the absence of an influence of the landscape on the diversity of this group. This result was similar in all the reforestation areas studied (Morato and Campos 2000), indicating a greater capacity of wasps to colonize areas undergoing restoration in a shorter period of time (Araújo 2020; 2021). Concerning the diversity of natural enemies, their greater richness in some areas could be more related to the greater abundance of wasp and bee nests than to environmental factors due to their direct relationship with the hosts (Torné-Noguera et al. 2020; Fornoff et al. 2021).
The size of the area under recovery can directly affect the attempt to restore degraded environments. The size acts as a limiting factor by restricting the richness and abundance of species of colonizing plants, which alters the availability of niches and the heterogeneity of the habitat and hinds the process of ecological succession (Suganuma et al. 2014). The edge effect is also a relevant factor related to the size of the areas under restoration. Due to the irregular shape of the majority of the studied reforestation areas and the proximity to the agricultural matrices, the edge effect acts more intensely in smaller and narrower reforestation areas (Hendrickx et al. 2007; Araújo et al. 2018a; Zirbel et al. 2019). Smaller reforestation areas may be more vulnerable to disturbances from the adjacent agricultural matrices such as the drift of pesticide “clouds”, fire, flooding, wind, solar radiation, and invasive species (Kennedy et al. 2013; Rocha-Filho et al. 2017; Araújo et al. 2018a, 2021; Fornoff et al. 2021). These variations related to the size and irregular shape can affect the formation of the canopy and understory of reforestation areas. They reflect in the structure of the community of plant species that supply resources such as pollen, resins, and pre-existing cavities, favoring the appearance of areas of clearings with a predominance of invasive plant species that do not provide resources for bees and wasps (Montalvo et al. 1997; Morato and Campos 2000; Rocha-Filho et al. 2020). These alterations may favor some species more efficient in colonizing disturbed environments (Morato and Campos 2000; Araújo et al. 2018a) and limit the restoration of species that require specialized resources (Hendrickx et al. 2007; Flores et al. 2018, 2019; Staab et al. 2018). Variation in the composition of wasps and bees that nest in pre-existing cavities has already been observed depending on the size and shape of reforested riparian areas (Araújo et al. 2018a). Smaller and narrower reforestation areas lacked specialized species due to the low plant heterogeneity and the greater intensity of the edge effect in these environments (Araújo et al. 2018a).
We also detected an evident association of bee species such as Euglossa pleosticta, E. townsendi, Eufriesea auriceps, Megachile guaranitica, and Carloticola paraguayensis showing a preference for natural habitats, i.e., landscapes with greater areas of reforestation and forest. Some of these species can be considered indicators of well-structured environments since they are often found in well-preserved forest fragments (Mello and Gaglianone 2019; Sobreiro et al. 2019). In comparison, in smaller reforestation areas we detected a greater association with species of bees with generalist habits, that is, species of broader trophic niches, cosmopolitan, found in all biomes (Aguiar and Zanella 2005). For example, the species Centris analis and C. tarsata are known for their excellent nesting capacity in environments with extensive land and urban use, often found in areas of open vegetation, forest edges, and related to diverse crops of fruit trees (Aguiar and Zanella 2005; Matos et al. 2013). Another generalist species also associated with small reforestation areas established in the vicinity of large monoculture areas was Epanthidium tigrinum. This species presents wide geographical distribution, little demand for nesting sites, polyletic behavior, absence of seasonality patterns, and absence of natural enemies, resulting in a high capacity to colonize environments that suffer constant disturbances (Gomes et al. 2020). These generalist species may also be benefiting from the high abundance of ruderal herbs belonging to the Asteraceae and Leguminosae families, usually associated with these areas of intensive land use and edges of reforestation areas (Matos et al. 2013). Recent studies indicate that the size and shape of riparian areas in the process of restoration influence the composition of cavity-nesting bees and wasps (Araújo et al. 2018a).
Wasps of the genera Pachodynerus, Trypoxylum, Monobia, and Priochilus were sampled in all the studied reforestation areas, although with high abundance in small reforestation areas and with few forest areas nearby. These wasp genera are known for their wide geographical distributions and high abundances in diverse environments such as ecological corridors (Holzschuh et al. 2009), open vegetation areas (Tscharntke et al. 1998), and agro-systems (Matos et al. 2013; Coutinho et al. 2020). They are considered generalist wasps with a high capacity to colonize disturbed environments (Buschini and Buss 2010). This high adaptability to disturbed environments may be related to the greater availability and variety of the prey of these species (larvae of Lepidoptera and spiders) usually found in these locations (Holzschuh et al. 2010; Krewenka et al. 2011). Their medium/large body size allows them to fly long distances and consequently colonize more distant patches in the landscape, as long as they are connected by fallow strips or organic farming (Holzschuh et al. 2009). The species Auplopus sp. 1 was associated with larger fragments and closer to forest areas. Species of this genus usually have a relatively small size and depend on specific food resources to supply their nests (spiders of specific species), being considered rare or exclusive to the neotropical region (Flores et al. 2018, 2019; Rocha-Filho et al. 2020).
We can conclude that reforestation areas such as those evaluated in the present study can be considered refuges or amenable to colonization for several species of bees, wasps, and their natural enemies, as long as these areas are in favorable landscapes (Araújo 2018a, 2021; Fornoff et al. 2021). Reforestation areas can be considered as corridor areas that connect habitat patches, contributing to the heterogeneity of the landscape. They provide essential resources for the survival of these organisms, functioning as an important tool in the conservation of local biodiversity (Staab et al. 2018). Despite the similarity in the characteristics of establishment and management of the reforested areas evaluated, aspects of the surrounding landscapes influenced the nesting of bees and wasps and, consequently, their natural enemies. Areas under restoration can be considered important refuges for forest species and can assist in the mobility of fauna, increasing the permeability of the landscape. Thus, these are important areas for the conservation of regional species, which provide essential services for the maintenance of ecosystems, including those that are restored. Restoration plans should analyze the effects of human activities on the restored landscapes and develop actions that seek to maximize the quality and heterogeneity of the surrounding matrices to guarantee ecological pollination and predation services, mitigate the decline in biodiversity, and assist in the restoration of degraded areas.
References
Aguiar CML, Zanella FCV (2005) The bee community (Hymenoptera: Apoidea: Apiformis) of an area in the border of the caatinga domain (Itatim, BA—Brazil). Neotrop Entomol 34:15–24
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Automat Contr 19:716–723. https://doi.org/10.1109/TAC.1974.1100705
Araújo GJ, Monteiro GF, Messias MCTB, Antonini Y (2018a) Restore it, and they will come: trap-nesting bee and wasp communities (Hymenoptera: Aculeata) are recovered by restoration of riparian forests. J Insect Conserv 22:245–256. https://doi.org/10.1007/s10841-018-0058-8
Araújo GJ, Fagundes R, Antonini Y (2018b) Trap-nesting Hymenoptera and their network with parasites in recovered riparian forests Brazil. Neotrop Entomol 47:26–36. https://doi.org/10.1007/s13744-017-0504-4
Araújo GJ, Stork-Tonon D, Izzo TJ (2020) Temporal stability of cavity-nesting bee and wasp communities in different types of reforestation in southeastern Amazonia. Restor Ecol 28:1528–1540. https://doi.org/10.1111/rec.13250
Ascher JS, Pickering J (2021) Discover life bee species guide and world checklist (Hymenoptera: Apoidea: Anthophila). http://www.discoverlife.org/mp/20q?guide=Apoidea_species. Accessed 27 Aug 2021
Baldock K (2020) Opportunites and threats for pollinator conservation in global towns and cities. Curr Opin Insect Sci 38:63–71. https://doi.org/10.1016/j.cois.2020.01.006
Bartón K (2016) Mumin: multi-model inference. R package version 1.10.6. https://cran.R-project.Org/package=mumin. Accessed 20 Oct 2021
Bender DJ, Tischendorf L, Fahrig L (2003) Using patch isolation metrics to predict animal movement in binary landscapes. Landsc Ecol 18:17–39. https://doi.org/10.1023/A:1022937226820
Borges FLG, Oliveira MR, Almeida TC, Majer JD, Garcia LC (2021) Terrestrial invertebrates as bioindicators in restoration ecology: a global bibliometric survey. Ecol Indic 125:107458. https://doi.org/10.1016/j.ecolind.2021.107458
Brühl CA, Zaller JG (2019) Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front Environ Sci 7:177. https://doi.org/10.3389/fenvs.2019.00177
Burnham KP, Anderson DR (2003) Model selection and multimodel inference. Technometrics 45:181–181
Buschini M, Buss C (2010) Biologic aspects of different species of Pachodynerus (Hymenoptera; Vespidae; Eumeninae). Brazilian J Biol 70:623–629. https://doi.org/10.1590/S1519-69842010000300020
Bush MB (2000) Ecology of a changing planet. Prentice-Hall Inc, Upper Saddle River
Calmon M, Brancalion PHS, Paese A et al (2011) Emerging threats and opportunities for large-scale ecological restoration in the Atlantic forest of Brazil. Restor Ecol 19:154–158. https://doi.org/10.1111/j.1526-100X.2011.00772.x
Campbell JW, Smithers C, Irvin A et al (2017) Trap nesting wasps and bees in agriculture: a comparison of sown wildflower and fallow plots in Florida. Insects 8:107. https://doi.org/10.3390/insects8040107
Campbell AJ, Gigante Carvalheiro L, Gastauer M et al (2019) Pollinator restoration in brazilian ecosystems relies on a small but phylogenetically-diverse set of plant families. Sci Rep 9:1–10. https://doi.org/10.1038/s41598-019-53829-4
Cardoso P, Bartonb PS, Birkhoferc K, Chichorroa F et al (2020) Scientists’ warning to humanity on insect extinctions. Biol Conserv 242:108–426. https://doi.org/10.1016/j.biocon.2020.108426
Cariveau DP, Bruninga-Socolar B, Pardee GL (2020) A review of the challenges and opportunities for restoring animal-mediated pollination of native plants. Emerg Top Life Sci 4:99–109. https://doi.org/10.1042/ETLS20190073
Cavalheiro AL, Torezan JMD, Fadeli L (2002) Recuperação de áreas degradadas: procurando por diversidade e funcionamento dos ecossistemas. In: Medri ME, Bianchini E, Shibatta OA, Pienta JA (eds) A bacia do rio Tibagi, 1st edn. Londrina, Paraná, Brasil, pp 213–224
Chao A, Gotelli NJ, Hsieh TC et al (2014) Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84:45–67. https://doi.org/10.1890/13-0133.1
Chapin FS, Zavaleta ES, Eviner VT et al (2000) Consequences of changing biodiversity. Nature 405:234–242. https://doi.org/10.1038/35012241
Coutinho JGE, Angel-Coca C, Boscolo D, Viana BF (2020) Heterogeneous agroecosystems support high diversity and abundance of trap-nesting bees and wasps among tropical crops. Biotropica 52:991–1004. https://doi.org/10.1111/btp.12809
Crivelato A, Lenis P, Dolacio T et al (2017) Notes on the nesting biology and foraging trips of the solitary wasp Pachodynerus guadulpensis (Saussure, 1853) (Vespidae: Eumeninae). Behav Ecol Conserv Syposium 2017:539–550
de Araújo GJ, Izzo TJ, Storck-Tonon D et al (2021) Re-establishment of cavity-nesting bee and wasp communities along a reforestation gradient in southern Amazonia. Oecologia 196:275–288. https://doi.org/10.1007/s00442-021-04920-z
Dixon KW (2009) Pollination and restoration. Science 325:571–573. DOI: https://doi.org/10.1126/science.1176295
Ebeling A, Klein AM, Weisser WW, Tscharntke T (2012) Multitrophic effects of experimental changes in plant diversity on cavity-nesting bees, wasps, and their parasitoids. Oecologia 169:453–465. https://doi.org/10.1007/s00442-011-2205-8
Fabian Y, Sandau N, Bruggisser OT et al (2014) Plant diversity in a nutshell: testing for small-scale effects on trap nesting wild bees and wasps. Ecosphere 5(2):8. https://doi.org/10.1890/ES13-00375.1
Flores LMA, Zanette LRS, Araújo FS (2018) Effects of habitat simplification on assemblages of cavity nesting bees and wasps in a semiarid neotropical conservation area. Biodivers Conserv 27:311–328
Flores LMA, Zanette LRS, Boscolo D, Araújo FS (2019) Landscape structure effects on bee and wasp assemblages in a semiarid buffer zone. Landsc Online 76:1–17. https://doi.org/10.3097/LO.201976
Fornoff F, Staab M, Zhu CD, Klein AM (2021) Multi-trophic communities re-establish with canopy cover and microclimate in a subtropical forest biodiversity experiment. Oecologia 196:289–301. https://doi.org/10.1007/s00442-021-04921-y
Gomes AMS, Silva CI, Cavalcante AM et al (2020) Bionomy and nesting behavior of the bee Epanthidium tigrinum (Schrottky, 1905) (Hymenoptera: Megachilidae) in trap-Nests. Sociobiology 67:247–255. https://doi.org/10.13102/sociobiology.v67i2.4759
Greenleaf S, Williams N, Winfree R et al (2007) Bee foraging ranges and their relationship to body size. Oecologia 153:589–596. https://doi.org/10.13102/sociobiology.v67i2.4759
Hadley AS, Betts MG (2012) The effects of landscape fragmentation on pollination dynamics: absence of evidence not evidence of absence. Biol Rev 87:526–544. https://doi.org/10.1111/j.1469-185X.2011.00205.x
Hagen M, Kissling WD, Rasmussen C et al (2012) Biodiversity, species interactions and ecological networks in a fragmented world. Adv Ecol Res 46:89–210. https://doi.org/10.1016/B978-0-12-396992-7.00002-2
Hendrickx F, Maelfait JP, Van Wingerden W et al (2007) How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes. J Appl Ecol 44:340–351. https://doi.org/10.1111/j.1365-2664.2006.01270.x
Hipólito J, Boscolo D, Viana BF (2018) Landscape and crop management strategies to conserve pollination services and increase yields in tropical coffee farms. Agric Ecosyst Environ 256:218–225. https://doi.org/10.1016/j.agee.2017.09.038
Holzschuh A, Steffan-Dewenter I, Tscharntke T (2009) Grass strip corridors in agricultural landscapes enhance nest-site colonization by solitary wasps. Ecol Appl 19:123–132. https://doi.org/10.1890/08-0384.1
Holzschuh A, Steffan-Dewenter I, Tscharntke T (2010) How do landscape composition and configuration, organic farming and fallow strips affect the diversity of bees, wasps and their parasitoids? J Anim Ecol 79:491–500. https://doi.org/10.1111/j.1365-2656.2009.01642.x
Johnson JB, Omland KS (2004) Model selection in ecology and evolution. Trends Ecol Evol 19:101–108. https://doi.org/10.1016/j.tree.2003.10.013
Jones ME, Davidson N (2016) Applying an animal-centric approach to improve ecological restoration. Restor Ecol 24:836–842. https://doi.org/10.1111/rec.12447
Kennedy CM, Lonsdorf E, Neel MC et al (2013) A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol Lett 16:584–599. https://doi.org/10.1111/ele.12082
Krebs J, Wilson J, Bradbury R et al (1999) The second silent spring? Nature 400:611–612. https://doi.org/10.1038/23127
Krewenka KM, Holzschuh A, Tscharntke T, Dormann CF (2011) Landscape elements as potential barriers and corridors for bees, wasps and parasitoids. Biol Conserv 144:1816–1825. https://doi.org/10.1016/j.biocon.2011.03.014
Krombein KV (1967) Trap nesting wasps and bees. Life histories, nests and associates, Washington, D.C
Kumar P (2010) Valuation and ecosystem services: an assessment of conceptual underpinnings. In: Koizumi T, Okabe K, Thompson I (eds) The role of forest biodiversity in the sustainable use of ecosystem goods and services in Agro-forestry Sisheries and Forestry. Proceeding of International Symposium for the Convention on Biological Divesity, Tokyo, pp 29–35
Lamb D, Gilmour D (2003) Rehabilitation and restoration of degraded forests. IUCN, Switzerland
Legendre P, Anderson MJ (1999) Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol Monogr 69:1–24. https://doi.org/10.1890/0012-9615(1999)069
Legendre P, Legendre L (1998) Numerical ecology. Elsevier, Amsterdam
Loyola RD, Martins RP (2008) Habitat structure components are effective predictors of trap-nesting Hymenoptera diversity. Basic Appl Ecol 9:735–742. https://doi.org/10.1016/j.baae.2007.06.016
Loyola RD, Martins RP (2011) Small-scale area effect on species richness and nesting occupancy of cavity-nesting bees and wasps. Rev Bras Entomol 55:69–74. https://doi.org/10.1590/S0085-56262011000100011
Matos MCB, Sousa-Souto L, Almeida RS, Teodoro AV (2013) Contrasting patterns of species richness and composition of solitary wasps and bees (Insecta: Hymenoptera) according to land-use. Biotropica 45:73–79. https://doi.org/10.1111/j.1744-7429.2012.00886.x
Mello BNS, Gaglianone MC (2019) Nesting biology of sympatric species of Megachilidae bees in a conservation area in brazilian Atlantic Forest. Sociobiology 66:52–60. https://doi.org/10.13102/sociobiology.v66i1.2881
Mendonça FA (2000) A tipologia climática: gênese, características e tendências. In: Stipp NMF (ed) Macrozoneamento ambiental da bacia hidrográfica do rio tibagi (PR). Londrina, Paraná, pp 21–62
Montalvo AM, Williams SL, Rice KJ et al (1997) Restoration biology: a population biology perspective. Restor Ecol 5:277–290. https://doi.org/10.1046/j.1526-100X.1997.00542.x
Morato EF, Campos LAO (2000) Efeitos da fragmentação florestal sobre vespas e abelhas solitárias em uma área da Amazônia Central. Rev Bras Zool 17:429–444. https://doi.org/10.1590/S0101-81752000000200014
Morato EF, Martins RP (2006) An overview of proximate factors affecting the nesting behavior of solitary wasps and bees (Hymenoptera: Aculeata) in preexisting cavities in wood. Neotrop Entomol 35:285–298. https://doi.org/10.1590/S1519-566X2006000300001
Moskalenko A (2016) Bee forage mapping based on multispectral images Landsat 8. Zemleustrìj kadastr ì monìtorìng zemelʹ 0:32–35
Oksanen J, Blanchet FG, Kindt R (2018) Vegan: community ecology package. R oachage version 2.5–5. http://cc.oulu.fi/~jarioksa/
Oliveira P, Gonçalves R (2017) Trap-nesting bee and wasps (Hymenoptera, Aculeata) in a Semidecidual Seasonal Forest fragment, southern Brazil. Pap Avulsos Zool 13:149–156. https://doi.org/10.11606/0031-1049.2017.57.13
Peipoch M, Brauns M, Hauer FR et al (2015) Ecological simplification: human influences on Riverscape Complexity. Bioscience 65:1057–1065. https://doi.org/10.1093/biosci/biv120
Pokorny T, Loose D, Dyker G et al (2015) Dispersal ability of male orchid bees and direct evidence for long-range flights. Apidologie 46:224–237. https://doi.org/10.1007/s13592-014-0317-y
Qgis Development Team (2021) Qgis geographic information system, Qgis Software, Qgis association. https://www.qgis.org
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://ww.R-project.org/
Reis R Jr, Oliveira ML, Borges GRA(2015) Rt4Bio:R tools for biologists (RT4Bio). R package version 1.0
Rocha-Filho LC, Rabelo LS, Augusto SC, Garófalo CA (2017) Cavity-nesting bees and wasps (Hymenoptera: Aculeata) in a semi-deciduous Atlantic forest fragment immersed in a matrix of agricultural land. J Insect Conserv 21:727–736. https://doi.org/10.1007/s10841-017-0016-x
Rocha-Filho LC, Montagnana PC, Boscolo D, Garófalo CA (2020) Green patches among a grey patchwork: the importance of preserving natural habitats to harbour cavity-nesting bees and wasps (Hy menoptera) and their natural enemies in urban areas. Biodivers Conserv 29:2487–2514. https://doi.org/10.1007/s10531-020-01985-9
Sánchez-Bayo F, Wyckhuys KAG (2019) Worldwide decline of the entomofauna: a review of its drivers. Biol Conserv 232:8–27. https://doi.org/10.1016/j.biocon.2019.01.020
Scervino RP, Torezan JMD (2015) Factors affecting the genesis of vegetation patches in anthropogenic pastures in the Atlantic forest domain in Brazil. Plant Ecol Divers 8:475–482. https://doi.org/10.1080/17550874.2015.1044582
Sexton AN, Benton S, Browning AC, Emery SM (2021) Reproductive patterns of solitary cavity-nesting bees responsive to both local and landscape factors. Urban Ecosyst 24(6):1271–1280. https://doi.org/10.1007/s11252-021-01116-4
Silveira FA (2004) Monitoring pollinating wild bees. In: Ruggiero M, Buchamnn S, Adams L (eds) The north american pollinator initiative. Fortaleza, Ceará, Brazil, pp 73–76
Sobreiro AI, Peres LLS, Boff S et al (2019) Continuous micro-environments associated orchid bees benefit from an Atlantic Forest remnant, Paraná State, Brazil. Sociobiology 66:293–305. https://doi.org/10.13102/sociobiology.v66i2.3443
Staab M, Pufal G, Tscharntke T, Klein AM (2018) Trap nests for bees and wasps to analyse trophic interactions in changing environments—A systematic overview and user guide. Methods Ecol Evol 9:2226–2239. https://doi.org/10.1111/2041-210X.13070
Stangler ES, Hanson PE, Steffan-Dewnter I (2015) Interactive effects of habitat fragmentation an microcimate on trap-nesting Hymenoptera and their trophic interactions in small secondary rainforest remnants. Biodivers Conserv 24:563–577. https://doi.org/10.1007/s10531-014-0836-x
Steckel J, Westphal C, Peters MK et al (2014) Landscape composition and configuration differently affect trap-nesting bees, wasps and their antagonists. Biol Conserv 172:56–64. https://doi.org/10.1016/j.biocon.2014.02.015
Steffan-Dewenter I, Münzenberg U, Bürger C et al (2002) Scale-dependent effects of landscape context on three pollinator guilds. Ecology 83:1421–1432. https://doi.org/10.1890/0012-9658(2002)083[1421:SDEOLC]2.0.CO;2
Steiner NC, K Köhler W (2003) Effects of landscape patterns on species richness–A modelling approach. Agric Ecosyst Environ 98:353–361. https://doi.org/10.1016/S0167-8809(03)00095-1
Suganuma MS, Assis GB, Durigan G (2014) Changes in plant species composition and functional traits along the successional trajectory of a restored patch of Atlantic forest. Community Ecol 15:27–36. https://doi.org/10.1556/ComEc.15.2014.1.3
Sumner S, Law G, Cini A (2018) Why we love bees and hate wasps. Ecol Entomol 43:836–845. https://doi.org/10.1111/een.12676
Taki H, Kevan PG, Viana BF et al (2008) Artificial covering on trap nests improves the colonization of trap-nesting wasps. J Appl Entomol 132:225–229. https://doi.org/10.1111/j.1439-0418.2007.01237.x
Tonietto RK, Larkin DJ (2018) Habitat restoration benefits wild bees: a meta-analysis. J Appl Ecol 55:582–590. https://doi.org/10.1111/1365-2664.13012
Torné-Noguera A, Arnan X, Rodrigo A, Bosch J (2020) Spatial variability of hosts, parasitoids and their interactions across a homogeneous landscape. Ecol Evol 10:3696–3705. https://doi.org/10.1002/ece3.6158
Tscharntke T, Gathmann A, Steffan-Dewenter I (1998) Bioindication using trap-nesting bees and wasps and their natural enemies: community structure and interactions. J Appl Ecol 35:708–719. https://doi.org/10.1046/j.1365-2664.1998.355343.x
Veddeler D, Schulze CH, Steffan-Dewenter I et al (2005) The contribution of tropical secondary forest fragments to the conservation of fruit-feeding butterflies: Effects of isolation and age. Biodivers Conserv 14:3577–3592. https://doi.org/10.1007/s10531-004-0829-2
Waltham NJ, Elliott M, Lee SY et al (2020) UN Decade on Ecosystem Restoration 2021–2030 what chance for success in restoring coastal ecosystems? Front Mar Sci 7:5. https://doi.org/10.3389
Wilson RS, Leonhard SD, Burwell CJ, Fuller C, Smith TJ et al (2020) Landscape simplification modifies trap-nesting bee and wasp communities in the subtropics. Insects 11:853. https://doi.org/10.3390/insects11120853
Zattara EE, Aizen MA (2021) Worldwide occurrence records suggest a global decline in bee species richness. One Earth 4:114–123. https://doi.org/10.1016/j.oneear.2020.12.005
Zirbel CR, Grman E, Bassett T, Brudvig LA (2019) Landscape context explains ecosystem multifunctionality in restored grasslands better than plant diversity. Ecology 100(4):e0263. https://doi.org/10.1002/ecy.2634
Zurbuchen A, Landert L, Klaiber J et al (2010) Maximum foraging ranges in solitary bees: only few individuals have the capability to cover long foraging distances. Biol Conserv 143:669–676. https://doi.org/10.1016/j.biocon.2009.12.003
Acknowledgements
We are grateful to: Instituto Chico Mendes (MMA-Brazil), Instituto Ambiental do Paraná (IAP) for the collecting permits; the owners for authorizing the study on their properties. We are greatly indebted to the following taxonomists: Carlos J. E. Lamas, Cecília Waichert, Daercio (A) A. Lucena, David (B) Muniz, Gabriel. (A) R. Melo, Léo C. Rocha–Filho, and Rogério (B) Lopes for the identification of part of the material collected in this study. This study was financed by CNPq–PELD (441540/2016–3), Fundação Araucária and Coordination for the Improvement of Higher Education Personnel – Brazil (CAPES) – Finance Code 001. ALG thanks CAPES and CNPq / MCTI / CONFAP-FAPs / PELD (No. 21/2020) for the scholarship. SMM thanks PIBIC–CNPq. SHS receives a fellowship from CNPq (305343 / 2018–1). We also thank two anonymous referees for providing many insightful comments.
Funding
CNPQ-PELD, Fundação Araucária and CAPES.
Author information
Authors and Affiliations
Contributions
ALG and SHS conceived the initial idea; ALG, SMM, and NU conducted the fieldwork; ALG and SMM conducted lab analysis on biological material; ALG and PNM carried out the data analyses; and ALG led the writing with assistance from all the authors.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest or competing interests.
Additional information
Communicated by Eckehard G. Brockerhoff.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gobatto, A.L., Miranda, P.N., Uemura, N. et al. Agricultural landscape influences on the solitary bees and wasps that nest in ecological restoration sites. Biodivers Conserv 32, 523–544 (2023). https://doi.org/10.1007/s10531-022-02510-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-022-02510-w