
Abstract
Elusive behaviour and financial constraints hamper the long-term monitoring of mammal population, particularly in intensively cultivated and urbanised landscapes, where most survey methodologies cannot be applied effectively. Provided that there is a direct relationship between each species’ density and frequency of road casualties, roadkill counts may represent a cost-effective alternative method to collect abundance data over long periods. We quantified the numbers of casualties of mammal species from 2001 to 2016 along two routes (65 km) crossing the heavily altered central River Po plain (N Italy). Each route was surveyed by car 10 times per month, covering 123,987 km and recording 15,589 road-kills from 15 species (15.3 roadkills/100 km/year). Most widespread mammals previously reported for the study area were recorded. Variation in each species’ roadkill numbers throughout the study period was consistent with available information on their distribution and abundance and the consistency of the patterns outlined on the two roads supported the hypothesis that the frequency of roadkills was related to each species’ density. Seasonal fluctuations in roadkill records could be related to either their reproductive cycles or dispersal patterns. For meso- and large species, the relationship between the occurrence of casualties and a set of 13 habitat variables was assessed by Logistic Regression Analysis. Based on our results, we believe that roadkill counts should be implemented to outline species’ population trends wherever high road density fragments wildlife habitats, and may represent a powerful citizen science-based method to collect large amounts of data over long periods.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Understanding changes in wildlife population trends is a major requisite for setting effective conservation and management actions. Only by monitoring population changes over a long period can important trends be identified and population models and hypotheses tested (Hanski et al. 1991, 2001; Korpimaki and Krebs 1996; Krebs et al. 2001; Battersby and Greenwood 2005; Brady and Slade 2004; Meltofte 2006; Schmidt et al. 2008).
Nonetheless, as the number of surveys is frequently limited by financial or material constraints or both (Lyra-Jorge et al. 2008), few studies have assessed changes in the abundances of mammal populations over periods longer than 3–5 years (Bartel et al. 2008). Mammal behaviour adds a further difficulty: most mammals are elusive and nocturnal, and consequently it can be extremely difficult to count actual numbers over large areas (Gese 2001).
Survey methods for large and medium-sized mammals include the use of sightings, tracks, and other signs, trapping and, more recently, camera-trapping and genetic census. All methods have advantages and disadvantages and reveal different levels of precision and cost–benefit ratios (Gaidet-Drapier et al. 2006; Swann and Perkins 2013; Carvalho et al. 2016). In addition, the probability of detection varies widely among species, habitats and time, and rare species may go undetected even by multi-year studies (Tobler et al. 2008).
Since the early 1900s (Krebs et al. 2001), several alternative methods have been proposed to avoid the obstacle posed by expensive standard methodologies, such as the analysis of game bag records, damage caused and encounter rates, including the count of roadkills (Baker et al. 2004; Gaidet-Drapier et al. 2006; Barthelmess and Brooks 2010; George et al. 2011).
Millions of wild animals are killed on roads each year, 40–50% of which are mammals (e.g. Huijser and Bergers 2000; Lodé 2000; Underhill and Angold 2000; Hell et al. 2005), road casualties currently being the main direct human cause of mortality for many species (Forman and Alexander 1998; Hauer et al. 2002). Roadkills can provide useful information about many aspects of mammal ecology and biology (see Brockie et al. 2009; González-Gallina et al. 2016) and may represent an effective data source for assessing relative abundances and tracking changes in population size for several mammal species (Donaldson and Bennett 2004; González-Gallina et al. 2016; George et al. 2011), provided that there is a direct relationship between their number and species’ density in the surrounding landscape (Baker et al. 2004).
Species differ in their leaning to be run over by vehicles (Taylor and Goldingay 2004; Brockie et al. 2009; Grilo et al. 2009) and the accuracy of road mortality estimates has been reported to be potentially affected by several factors, e.g. body size (Ford and Fahrig 2007), diet (Cook and Blumstein 2013), seasonal movements and dispersion (Neumann et al. 2012), the rate of removal of carcasses by scavengers (Slater 2002), road traffic and survey methods (e.g. car speed and the time interval between successive surveys; Collinson et al. 2014; Santos et al. 2015). Whether each of these potential sources of detection bias may hinder the assessments of absolute abundances and make unreliable any comparison between the estimated abundance of either different species or investigations, the mean number of each species’ casualties assessed over long periods of time may be expected to reflect changes in population abundance and thus provide useful information about population trends. A correlation between density and roadkill counts has been reported, as an example, for red fox Vulpes vulpes (Baker et al. 2004) and European hedgehog Erinaceus europaeus (Roos et al. 2012) in the UK, Eastern barred bandicoot Perameles gunnii in Tasmania (Mallick et al. 1998), raccoon Procyon lotor in Illinois (Gehrt et al. 2002) and several species of carnivores in Portugal (Grilo et al. 2009).
Although some ongoing citizen science-based projects promise the collection of large amounts of data (Vercayie and Herremans 2015), till now roadkill monitoring projects have been conducted at small time-scales, hampering the assessment of long-term trends.
We conducted a 16 year-long monitoring of roadkills in the central Po-Venetian plain (N Italy), an intensively cultivated and densely populated area where widespread human presence and negligible percent cover of semi-natural habitats make most used survey techniques, such as track plots, camera-trapping and DNA-enhanced surveys for scats, difficult to be performed at large scales in terms of either space or time. Moreover, none of these would be a cost-effective method for collecting data about the whole mammal community, including a range of species differing in size, distribution and behaviour. Accordingly, previous investigations have been mostly carried out in protected areas (e.g. Balestrieri et al. 2016a), where protection enhances habitat diversity and species richness, or focused on problematic wildlife, e.g. invasive coypu Myocastor coypus (Balestrieri et al. 2016b) or game species, e.g. European hare Lepus europaeus (Vidus-Rosin et al. 2010, 2012).
By quantifying the monthly and yearly numbers of casualties of each species from January 2001 to December 2016, we aimed to (i) assess long-term population trends of mammal species occurring in agricultural landscapes, (ii) verify the role played by known biological periods in increasing the probability of each species to be struck by vehicles, and (iii) the relationship between the occurrence of casualties and a set of spatial factors which may influence the likelihood of animal-vehicle collisions.
Study area
The Po-Venetian plain is the largest Italian plain (ca. 46,000 km2). The pedogenetic and micro-morphological characteristics of the soils of the lower plain, crossed by the River Po (652 km), support high levels of agricultural productivity and are intensively managed for cattle husbandry and the production of rice, maize, wheat, sugar beet, fruit and horticultural products (Ajassa et al. 1997). As a consequence, 81% of the plain shows low to medium-low levels of landscape diversity (Pileri and Sartori 2004), most residual wood patches (mean size: 4.5 ha) being close to pre-alpine hills or along the main tributaries of the River Po (Lassini et al. 2007). In its central part (Lombardy region), 77% of the plain is farming land, while only 3.65% is covered by residual woodland. We focused on this heavily altered sector of the Po plain, included in the provinces of Pavia (2158 km2) and Lodi (782 km2). In both provinces, road density (Lodi: 0.69 km/km2; Pavia: 0.73 km/km2) was higher than the average for Northern Italy (0.5 km/km2; ACI 2011). In the portion crossed by the monitored roads (ca. 880 km2), in 2009 cultivated land and human settlements covered, respectively, 98% and 1.2% ca. of the total area, while woods and shrubs accounted for less than 0.1%; throughout the study period the percent cover of maize increased (+ 10.8%) and human settlements slightly expanded at the cost of cultivated land (Gherardi et al. 2009).
In agricultural lands, the loss of natural vegetation and fragmentation of uncultivated features have been related to a general decline in biological diversity (Wilcox and Murphy 1985; Donald et al. 2001; Benton et al. 2003). At the start of the monitoring period, 21 mammal species (bats excluded) had been reported for the study area (Table 1; Prigioni et al. 2001).
Methods
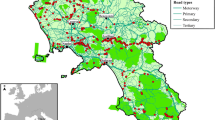
Surveys were carried out by one driver on two provincial, two-lane routes (one for traffic in each direction with no median barriers): SS235 (Route North, RN), connecting Pavia to Lodi (31 km), and SS234 (Route South, RS), connecting Pavia to Casalpusterlengo (34 km; Fig. 1). Between January 2001 and December 2016 we conducted 3834 road surveys (1711 on Route North and 2123 on Route South; Fig. 1). On average (± SD), each route was surveyed 10 ± 4.5 times per month (RN: 8.9 ± 3.9; RS: 11.0 ± 4.7) for the whole duration of the study, covering a total of 123987 km (RN: 58,174 km; RS: 65,813 km). Assuming that most casualties occur during the night, surveys were carried out in the morning, ca. 1–2 h after dawn. Data about car traffic on the two monitored roads were available for the period 2004–2007, when the mean number of vehicles per day kept constant (Fig. 2).

Study area (N Italy, Lombardy region), showing the two monitored routes (RN Route North, connecting Pavia to Lodi; RS Route South, connecting Pavia to Casalpusterlengo)

Mean number of vehicles passing each day on the two monitored routes (Route North, RN, and Route South, RS) between Spring 2004 and Winter 2007
On both routes, posted speed limits ranged between 50 and 70 km/h. Keeping in mind the recommendations by Clevenger et al. (2003), we drove at a speed 10–20 km/h below the posted limits. Consistently, a maximum speed of 50 km/h has been recently reported to be suitable for obtaining time-effective, reliable data (Collinson et al. 2014).
For each casualty, we recorded the species, date and location, using a 1:10000 map of the area. Small mammals were collected to identify the species with accuracy. Records of bats, domestic animals and non-identifiable individuals were discarded.
To account for variation in survey effort between routes and among either years or months, data were expressed as number of animals killed per 100 km surveyed. For the most frequently roadkilled species, the relationship between the number of roadkilled animals/100 km (Y) and year (X) was tested by regression analysis to find the model that best fitted each species’ population trend over time. Spearman’s correlation test was used to assess the consistency between the trends recorded on each route on a yearly basis. Similarity between routes was assessed by Bray–Curtis’ similarity index (Bray and Curtis 1957): INS = 2NNS/(NN + NS), where: NN = total number of individuals at Route North, NS = total number of individuals at Route South, and NNS = sum of the minimum number of each of the species shared between routes. The index ranges between 0 and 1 (identity).
For each meso- and large mammal species recorded, the relationship between the occurrence of casualties and a set of 13 habitat variables was assessed by backward stepwise Logistic Regression Analysis (LRA), testing for the statistical significance of each coefficient in the model by Wald’s test. We classed both RN and RS into 1000 m segments (n= 65) and measured the following variables: (1) number of passageways under roads (UndPass); (2) road lightening (RLight; yes/no); (3) number of watercourses crossed by the route (Nwc); (4) number of canals crossed by the route (Nc); (5) number of crossroads (CrossR); (6) presence of hedgerows or shrub patches close to road edges (HRows); (7) number of field margins crossed on both road sides (Nfs); 8–13% cover (%C) of six main habitats (meadows mw, rice-fields rf, poplar plantations pp, maize fields mf, human settlements hs, woods wo) into a 500 m large belt on each road side.
The first two variables aimed to assess the effects of potential mitigation measures, whilst variables 3–6 tried to assess the influence of linear features and vegetation cover adjacent to road verges (Hrows) on the probability of road-crossing. Nfs was intended as a measure of habitat heterogeneity. Land cover features were derived by the regional land-use map of 2009 (https://www.dati.lombardia.it/Territorio/Uso-Suolo-Dusaf-2009/y6xb-wpka). Maize was the most widespread crop other than rice; other crops, namely wheat, barley, soya-bean, accounted for less than 10% of the whole area and were included in the variable mf.
To avoid collinearity, the correlation between the predictors was measured by Spearman’s coefficient. Variables to be entered in the model were then selected so as to omit those clearly representing redundant information; when two variables were correlated (P < 0.01), the one to be rejected was chosen according to the strength of its correlation with the dependent variable (i.e. Nwc and %Chs; Hosmer and Lemeshow 1989).
Results
We recorded 15,589 mammalian road-kills from 15 species (57% of the mammals reported in the Atlas of Lombardy mammals plus three previously unrecorded species; Prigioni et al. 2001) and 5 orders (Insectivora, Rodentia, Lagomorpha, Carnivora and Artiodactyla) (Table 1). Coypu (67.6% of casualties) and Eastern cottontail Sylvilagus floridanus (28.2%) were the most frequently roadkilled species, followed by European badger Meles meles and common rat Rattus norvegicus (1% each) (Table 1). European hare and common rabbit Oryctolagus cuniculus accounted for 0.1–0.2% of total records. Small mammals included wood mouse Apodemus sylvaticus (0.2%), bank vole Clethrionomys glareolus (0.1%), Savi’s vole Microtus savii (0.1%) among rodents, and European mole Talpa europaea (0.1%) and European hedgehog (0.6%) among insectivores (Table 1). Despite slight differences in numbers due to sampling effort (Table 1), results on the two routes were quite consistent (INS = 0.98). Wild boar Sus scrofa and Savi’s vole were more common on RS, while roe deer Capreolus capreolus were recorded more frequently on RN. Stone marten Martes foina was found only on RS. On average (± SE) we found 15.3 ± 1.5 roadkills/100 km/year (0.56 ± 0.025, excluding non-native cottontails and coypu).
Throughout the study period, the number of roadkills increased steadily for badgers and Eastern cottontails (Fig. 3b, f; Table 2). Wild boars and roe deer were recorded for the first time in 2006 and 2007, respectively, after which their number tended to increase (Fig. 3c, d; Table 2). Between 2001 and 2005 the number of European hedgehog casualties declined sharply, from 0.3 to 0.06 ind./100 km per year (Fig. 3a; Table 2). Hares, rabbits, foxes (Fig. 3e; Table 2) and coypu (Fig. 4) showed a less clear trend, with large fluctuations in the number of roadkilled individuals. The pattern of both fox and coypu variation in abundance could be compared to the number of individuals shot each year during pest-control operations (Province of Lodi, unpubl. data). Numbers followed an opposite trend for foxes (Fig. 5), while coypu’s variation in roadkill numbers was mirrored, with a one-year time-lag, by the rise and fall of the number of individuals shot (Fig. 4). No rabbit was recorded in 2004 and 2009.

Population trend of most frequently roadkilled species as assessed by regression curves (a European hedgehog, b European badger, c roe deer, d wild boar, e: red fox, f: Eastern cottontail)

Comparison between the trend of coypu population assessed by roadkill counts and the number of coypu killed each year for pest-control

Comparison between the trend of red fox population assessed by roadkill counts and the number of foxes killed each year for pest-control
Variation in yearly roadkill numbers followed the same pattern on both monitored routes for most species, including hedgehogs (ρ = 0.61, P = 0.011), badgers (ρ = 0.87, P < 0.001), wild boars (ρ = 0.60, P = 0.015), roe deer (ρ = 0.68, P = 0.004), Eastern cottontails (ρ = 0.93, P < 0.001) and coypu (ρ = 0.94, P < 0.001; df = 16 for all tests).
The number of casualties of roadkilled species varied throughout the year. For hedgehogs, cottontails and coypu, the highest mortality occurred in mid-summer, with coypu showing a second peak in December (Figs. 6, 7). Both badgers and foxes suffered the highest mortality in July–August, with a second peak in April-May and February, respectively (Fig. 6). Spring was the critical period for roe deer, whilst the number of roadkilled wild boars peaked in winter (Fig. 6). Monthly patterns on the two routes were correlated for badgers (ρ = 0.82, P = 0.001), hedgehogs (ρ = 0.87, P < 0.001) and roe deer (ρ = 0.68, P = 0.015).

Monthly variation in the number of roadkilled ind./100 km of route for five mammals widespread in the study area

Monthly variation in the number of roadkilled coypu- and Eastern cottontail ind./100 km of route in the study area
Variables affecting the probability of road casualties were identified for four species (Table 3). The occurrence of roadkilled badgers (recorded on 71% of road stretches) and roe deer (25%) increased with the percent cover of meadows, whilst hedgehog (71%) occurrence was positively related to rice fields. Badger roadkills also increased with the number of crossroads. Fox (54%) mortality was inversely related to the number of underpasses and the cover of maize fields. The probability of foxes being hit was also lower on lit road stretches. In contrast, the occurrence of vegetation (hedgerows and shrubs) next to road edges increased the risk of casualties. Wild boar were recorded on 22% of road stretches, whilst Eastern cottontail, coypu and common rat casualties occurred on all stretches (100%).
Discussion
Counts of roadkills proved an effective method for monitoring population trends of mammalian species for the minimum time-length required to assess trends via regression with a sufficient level of statistical power (15.9 ys; White 2017). Most of the medium- and large-sized mammals reported for the study area were recorded and the consistency of both numbers and patterns on the two roads and seasonal patterns with life-history phenologies supported the hypothesis that the frequency of roadkills was related to each species’ density. Three mammals—Eastern cottontail, wild boar and roe deer—did not occur in the study area at the start of roadkill monitoring. Up to the beginning of the century the Eastern cottontail, a lagomorph native to the American continent introduced to Piedmont region in the mid ‘60 s (Spagnesi 2002), had been recorded only in the western part of Pavia province and on the banks of the River Po. In the last two decades it has rapidly spread and currently occurs in the whole western and central part of the Po plain, where it can reach high densities (Piedmont: 110 ind./km2; Bertolino et al. 2011. Lombardy: 65.6 ind./km2; Vidus-Rosin et al. 2008). In the last 50 years, both the wild boar and roe deer have shown a rapid increase in numbers across Europe (Saez-Royuela and Telleria 1986; Vernesi et al. 2002). In Italy, after having almost gone extinct in the first half of the last century, since the 1960 s both species have started to recover because of countryside abandonment and reintroductions (Perco and Calò 1994; Sacchi 2001). Consistently with roadkill data, until 2006 none of the two species had been reported to occur in the study area (Carnevali et al. 2009). Until 2012 wild boar casualties were recorded only on Route South, suggesting the species’ spreading from the Apennines, whilst the opposite occurred for roe deer, the recovery of which has been driven by the expansion of Alpine residual nuclei (Perco 1987). The number of roadkilled roe deer increased with the percent cover of meadows, which, in agricultural areas, are selected because offering high quality food with respect to cereal fields (Lande et al. 2014).
A positive population trend was recorded also for the European badger. During the monitoring period, on an isolated hill included in the study area badger density was assessed to be higher than the average for continental Europe (0.9–1.4 adult individuals/km2; Balestrieri et al. 2016a).
Road-traffic mortality is considered a major threat to badger population viabilities in many European countries (Griffiths and Thomas 1997), and have been reported as a possible cause of badger decline in the Netherlands (van der Zee et al. 1992). Accordingly, in the study area badgers paid to road traffic one of the highest toll. Their probability of being struck by vehicles increased with the percent cover of meadows, which may represent attractive-to-badgers patches rich in earthworms, the main source of protein for badgers in lowlands of northern Italy (Canova and Rosa 1994; Balestrieri et al. 2004). Roads crossing the two major routes may act as further linear barriers driving badger movements and increasing the risk of casualties. In contrast, risk for foxes increased with the availability of hedgerows and shrubs, suggesting that they select patches and corridors offering cover for moving across their ranges. Avoidance of maize fields was consistent with previous research in the western part of the River Po plain (Balestrieri 1993). Our results suggest that foxes respond better than other mammals to available mitigation measures (underpasses) and road lights may intimidate animals from approaching the road.
Despite depending on hunting effort, which may vary from year to year, the number of both foxes and coypu shot for “pest”-control was related, although in different ways, to the number of roadkills, supporting the hypothesis that road casualties reflect population abundance. Coypu trends confirmed the lack of effectiveness of current management actions for reducing population size (Balestrieri et al. 2016b).
Like most semi-aquatic mammals introduced in Europe, the coypu is a successful colonizer of freshwater ecosystems and, as indicated by the large number of coypu killed each year on the two monitored roads, is currently widespread in agricultural landscapes of northern Italy. In Lombardy population size has been assessed to be ca. 700,000 adult individuals, Lodi being the province with the highest coypu density (Balestrieri et al. 2016b).
Although small mammals tend to avoid roads (McGregor et al. 2008) and short removal times affect their detectability (Slater 2002), roadkill monitoring allowed to record the most widespread rodents. While the wood mouse is the dominant species in cultivated lands, bank voles occur only in residual wood patches (Balestrieri et al. 2015); Savi’s voles prefer open habitats and are considered to be widespread in the Po plain (Canova 1992). Small Soricidae went undetected, while the hedgehog showed a negative trend, with a sharp decline in the number of roadkills between 2001 and 2005. Consistently, between 1995 and 2016 the relative frequency of occurrence of insectivores in the diet of barn owls dropped from 38.2 to 14.6%, suggesting that agricultural changes (increase in maize cover) and the use of pesticides may have affected the abundance of these predators of ground-dwelling invertebrates (Fiandra 2016). In the last decades, a decline in hedgehog populations has been recorded also in the UK (Poulton and Reeve 2010; Roos et al. 2012), where it has been related to both habitat loss and increase in badger numbers (Pettett et al. 2018). Most rice fields occurring in Pavia province, the relationship between rice cover and casualties may suggest that hedgehogs are managing better in the western part of the study area.
As for small mammals, the recorded numbers of hares and rabbits was probably affected by the removal of carcasses by either predators or drivers themselves. Nonetheless, rabbit trend showed two negative peaks consistent with as many epidemic events (myxomatosis in 2004 and hemorrhagic enteritis in 2009), which sharply lowered population density.
With respect to available presence data, the lack of weasel (Mustela nivalis) records was unexpected. Although carcasses can be rapidly removed by corvids, anecdotic reports about a sharp drop in sightings may suggest that this species is actually declining. Consistently, in cultivated lands the availability of their main prey, small rodents, has been reported to be negligible (Balestrieri et al. 2015).
All roadkills were widespread and small to medium-sized mammals (except for wild boars and roe deer), giving rise to the question whether rare (such as polecat Mustela putorius) or larger species may go undetected. In the study area the wild boar is the largest mammal, but elsewhere the road mortality of species as large as wolf Canis lupus (Mech et al. 1988), moose Alces alces and black bear Ursus americanus (Fudge et al. 2007) has been well quantified and related to each species’ density; in Lombardy region, in the last two decades roadkill records have been a major evidence of the persistence of a small reintroduced otter Lutra lutra population (Prigioni et al. 2009; authors’ unpubl. data), suggesting that roadkill monitoring may be effective also for low-density species.
For the most recorded species, seasonal fluctuations in roadkill records could be related to either their reproductive cycles or dispersal patterns. Roe deer casualties peaked during their dispersal phase (Perco 2003), whilst the collision probability for wild boars was the highest during the mating period and particularly in January, when matings are most frequent (Apollonio 2003). As reported by previous studies (Hell et al. 2005; Haigh 2012), for most species (i.e.: red fox, badger, coypu, cottontail and hedgehog), roadkills peaked in summer, when the density of juveniles was the highest. Coypu, badger and fox casualties followed a bimodal pattern, both carnivores showing also a mating season-related peak. In winter, coypu tendency to assemble in available meadows for foraging (pers. obs.) may increase the risk of collisions.
Agricultural intensification is considered a major cause of the decline of many wildlife populations (Pain and Pienkowski 1997; Krebs et al. 1999; Donald et al. 2001), matrix quality affecting both the time devoted by species to the search for resources and their movements (Arberg et al. 1995; Wagner and Fortin 2005). In the heavily altered landscape of the central River Po plain, the number of casualties/100 km was apparently higher than those reported by previous studies throughout Europe (e.g. Ireland: 1.2–1.54 ind/100 km, Smiddy 2002; Haigh 2012; Slovakia: 6.3 ind/100 km Hell et al. 2005; Portugal: 11.6 ind/100 km, Carvalho and Mira 2012; France: 14.5 vertebrates/100 km, Lodé 2000; but see Grilo et al. 2009: 47 carnivores/100 km in Sothern Portugal). Nonetheless, two invasive non-native species—the Eastern cottontail and coypu-, accounted for more than 90% of records, suggesting a high level of environmental degradation and depletion of the native mammal communities. While agricultural intensification, together with increasing urbanisation and introductions, is likely to affect the populations of most specialised and demanding species (Kleijn et al. 1998, 2001), it has been recently demonstrated that the suitability of crop types can differ among species (Gastón et al. 2016), supporting the general perception that some mammals may benefit from agricultural lands (Gehring and Swihart 2003; Laurance 1994). In the River Po plain, the progressive increase in maize production for fodder, which has prolonged its availability until the end of October, is likely to have played a major role, together with the ongoing reduction in the number of huntsmen, in the spread of wild ungulates and badgers. Consistently, the increase in UK badger populations occurred since the 1980s has been related to changes in patterns of agriculture and hunting pressure (Battersby and Tracking Mammals Partnership 2005; Judge et al. 2017).
Animal–vehicle collisions are the main cause of death for many mammalian predators—including “near threatened” (according to IUCN’s criteria) Eurasian otter Lutra lutra (Hauer et al. 2002) and “endangered” Iberian lynx Linx pardinus (Ferreras et al. 1992)-, and also pose human safety at risk, with important economic and social consequences (Williams and Wells 2005; Vignon and Barbarreau 2008; Morelle et al. 2013). Despite these critical issues, we demonstrated that roadkill counts can be considered an efficient method for the long-term monitoring of mammal communities in human landscapes where standard methodologies would prove ineffective or costly. Moreover, roadkill data can be collected by trained volunteers, potentially allowing analyses at very large spatial scales in a short time and with negligible costs. Monitoring projects based on citizen science represent a powerful tool for wildlife management and recently some volunteer-based monitoring projects have been successfully started up (Roos et al. 2012; van der Ree et al. 2015; Vercayie and Herremans 2015). Despite many challenges that need to be evaluated, such as the integration of collected data in the scientific process and variability in species’ detectability across time and space (Schmeller et al. 2009; Bonardi et al. 2011), we suggest that citizen science roadkill counts should be implemented to outline species’ population trends in agricultural landscapes and wherever high road density fragments wildlife habitats.
References
A.C.I. Automobile Club d’Italia (2011) Dotazione di infrastrutture stradali sul territorio italiano. Elaborazioni a cura di Automobile Club d’Italia-Area Professionale Statistica, Rome
Ajassa R, Beretta E, Biagini E, Biancotti A, Bonansea E, Boni P, Brancucci G, Carton A, Cerutti AV, Ferrari R, Giardino M, Laureti L, Maraga F, Marchetti G, Masino A, Motta L, Motta M, Ottone C, Pellegrini L, Rossetti R, Viola E (1997) Mountains, hills and plains in north-western Italy. Suppl Geogr Fis Dinam Quat 2:49–78
Apollonio M (2003) Sus scrofa. In: Boitani L, Lovari S, Vigna Taglianti A (eds) Fauna d’Italia: mammalia III, carnivora-artiodactyla. Calderini, Bologna, pp 261–271
Arberg J, Jansson G, Swenson JE, Angelstam P (1995) The effect of matrix on the occurrence of hazel grouse in isolated habitat fragments. Oecologia 103:265–269
Baker P, Harris S, Robertson CPJ, Saunders G, White PCL (2004) Is it possible to monitor mammal population changes from counts of road traffic casualties? An analysis using Bristol’s red foxes, Vulpes vulpes as an example. Mammal Rev 34:115–130
Balestrieri A (1993) Aree vitali e preferenze ecologiche della volpe (Vulpes vulpes) in due zone delle province di Pavia ed Alessandria. Dissertation, Università degli Studi di Milano, Milan
Balestrieri A, Remonti L, Prigioni C (2004) The diet of the Eurasian badger (Meles meles) in an agricultural river habitat (NW Italy). Hystrix 16:3–12
Balestrieri A, Remonti L, Morotti L, Saino N, Prigioni C, Guidali F (2015) Multilevel habitat preferences of Apodemus sylvaticus and Myodes glareolus in an intensively cultivated agricultural landscape. Ethol Ecol Evol. https://doi.org/10.1080/03949370.2015.1077893
Balestrieri A, Cardarelli E, Pandini M, Remonti L, Saino N, Prigioni C (2016a) Spatial organisation of European badger Meles meles as assessed by camera-trapping. Eur J Wildl Res 62:219–226
Balestrieri A, Zenato M, Fontana E, Vezza P, Remonti L, Caronni FE, Saino N, Prigioni C (2016b) An indirect method for assessing the relative abundance of introduced pest Myocastor coypus (Rodentia) in agricultural landscapes. J Zool 298:37–45
Bartel RA, Knowlton FF, Stoddart LC (2008) Long-term patterns in mammalian abundance in northern portions of the Great Basin. J Mammal 89:1170–1183
Barthelmess EL, Brooks MS (2010) The influence of body-size and diet on road-kill trends in mammals. Biodivers Conserv 19:1611–1629
Battersby J (ed) & Tracking Mammals Partnership (2005) UK mammals: species status and population trends. First report by the tracking mammals partnership. JNCC/Tracking Mammals Partnership, Peterborough
Battersby JE, Greenwood JJD (2005) Monitoring terrestrial mammals in the UK: past, present and future, using lessons from the bird world. Mammal Rev 34:3–29
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Bertolino S, Perrone A, Gola L, Viterbi R (2011) Population density and habitat use of the introduced Eastern Cottontail (Sylvilagus floridanus) compared to the native European hare (Lepus europaeus). Zool Stud 50:315–326
Bonardi A, Manenti R, Corbetta A, Ferri V, Fiacchini D, Giovine G, Macchi S, Romanazzi E, Soccini C, Bottoni L, Padoa-Schioppa E, Ficetola GF (2011) Usefulness of volunteer data to measure the large scale decline of “common” toad populations. Biol Conserv 144:2328–2334
Brady MJ, Slade NA (2004) Long-term dynamics of a grassland rodent community. J Mammal 85:522–561
Bray JR, Curtis JT (1957) An ordination of upland forest communities of southern Wisconsin. Ecol Monogr 27:325–349
Brockie RE, Sadleir RMFS, Linklater WL (2009) Long-term wildlife road-kill counts in New Zealand. N Z J Zool 36:123–134
Canova L (1992) Distribution and habitat preference of small mammals in a biotope of the North Italian plain. Boll Zool 59:417–421
Canova L, Rosa P (1994) Badger Meles meles and fox Vulpes vulpes food in agricultural land in the Western Po plain. Hystrix 5:73–78
Carnevali L, Pedrotti L, Riga F, Toso S (2009) Banca Dati Ungulati: status, distribuzione, consistenza, gestione e prelievo venatorio delle popolazioni di Ungulati in Italia. Rapporto 2001–2005. Biol Cons Fauna 117:1–168
Carvalho F, Mira A (2012) Comparing annual vertebrate road kills over two time periods, 9 years apart: a case study in Mediterranean farmland. Eur J Wildl Res 57:157–174
Carvalho WD, Rosalino LM, Adania CH, Esbérard CEL (2016) Mammal inventories in Seasonal Neotropical Forests: traditional approaches still compensate drawbacks of modern technologies. Iheringia Sér Zool 106:e2016005
Clevenger AP, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109:15–26
Collinson WJ, Parker DM, Bernard RTF, Reilly BK, Davies-Mostert HT (2014) Wildlife road traffic accidents: a standardized protocol for counting flattened fauna. Ecol Evol 4:3060–3071
Cook TC, Blumstein DT (2013) The omnivore’s dilemma: diet explains variation in vulnerability to vehicle collision mortality. Biol Conserv 167:310–315
Donald PF, Green RE, Heath MF (2001) Agricultural intensification and the collapse of Europe’s farmland bird population. Proc R Soc Lond B 268:25–29
Donaldson A, Bennett A (2004) Ecological effects of roads: Implications for the internal fragmentation of Australian parks and reserves. Parks Victoria Technical Series No. 12, Parks Victoria, Melbourne
Ferreras P, Aldama JJ, Beltrán JF, Delibes M (1992) Rates and causes of mortality in a fragmented population of Iberian lynx Felis pardina Temminck, 1824. Biol Conserv 61:197–202
Fiandra M (2016) Variazioni della composizione della comunità dei micromammiferi del parco Adda Sud nel periodo 1994–2016. Dissertation, University of Pavia
Ford AT, Fahrig L (2007) Diet and body size of North American road mortalities. Transp Res D 12:498–505
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol S 29:207–231
Fudge D, Freedman B, Crowell M, Nette T, Power V (2007) Road-kill of mammals in Nova Scotia. Can Field-Nat 121:265–273
Gaidet-Drapier N, Fritz H, Bourgarel M, Renaud P-C, Poilecot P, Chardonnet P, Coid C, Poulet D, Le Bel S (2006) Cost and efficiency of large mammal census techniques: comparison of methods.for a participatory approach in a communal area, Zimbabwe. Biodivers Conserv 15:735–754
Gastón A, Blazquez-Cabrera S, Garrote G, Mateo-Sanchez MC, Beier P, Simon MA, Saura S (2016) Response to agriculture by a woodland species depends on cover type and behavioural state: insights from resident and dispersing Iberian lynx. J Appl Ecol 53:814–824
Gehring TM, Swihart RK (2003) Body size, niche breath, and ecologically scaled responses to habitat fragmentation: mammalian predators in an agricultural landscape. Biol Conserv 109:283–295
Gehrt SD, Hubert GF, Ellis JA (2002) Long term population trends of raccoons in Illinois. Wildl Soc Bull 30:457–463
George L, Macpherson JL, Balmforth Z, Bright PW (2011) Using the dead to monitor the living: can road kill counts detect trends in mammal abundance? Appl Ecol Environ Res 9:27–41
Gese EM (2001) Monitoring of terrestrial carnivore populations. In: Gittleman JL, Funk SM, Macdonald D, Wayne RK (eds) Carnivore conservation. Cambridge University Press, Cambridge, pp 327–396
Gherardi M, Lorito S, Vianello G, Vittori Antisari L (2009) Qualitative and quantitative evaluation of soil depletion due to urbanisation in the areas near the Po River. EQA: Environ Qual 2:29–38
González-Gallina A, Benítez-Badillo G, Hidalgo-Mihart MG, Equihua M, Rojas-Soto OR (2016) Roadkills as a complementary information source for biological surveys using rodents as a model. J Mammal 97:145–154
Griffiths HI, Thomas DH (1997) The conservation and management of the European badger (Meles meles). Nature and environment No 90, Council of Europe publishing, Strasbourg Cedex
Grilo C, Bissonette JA, Santos-Reis M (2009) Spatial-temporal patterns in Mediterranean carnivore road casualties: consequences for mitigation. Biol Conserv 142:301–313
Haigh A (2012) Annual patterns of mammalian mortality on Irish roads. Hystrix, Ital J Mammal 23(2):58–66
Hanski I, Hansson L, Henttonen H (1991) Specialist predators, generalist predators, and the microtine cycle rodent cycle. J Anim Ecol 60:353–367
Hanski I, Henttonen H, Korpimaki E, Oskanen L, Turchin P (2001) Small rodent dynamics and predation. Ecology 76:1505–1520
Hauer S, Ansorge H, Zinke O (2002) Mortality patterns of otters (Lutra lutra) from eastern Germany. J Zool 256:361–368
Hell P, Plavý R, Slameka J, Gašparík J (2005) Losses of mammals (Mammalia) and birds (Aves) on roads in the Slovak part of the Danube Basin. Eur J Wildl Res 51:35–40
Hosmer DW, Lemeshow S (1989) Applied logistic regression. Wiley, New York
Huijser MP, Bergers PJM (2000) The effect of roads and traffic on hedgehog (Erinaceus europaeus) populations. Biol Conserv 95:111–116
Judge J, Wilson GJ, Macarthur R, McDonald RA, Delahay RJ (2017) Abundance of badgers (Meles meles) in England and Wales. Sci Rep 7:276
Kleijn D, Joenje W, Le Coeur D, Marshall EJP (1998) Similarities in vegetation development of newly established herbaceous strips along contrasting European field boundaries. Agr Ecosyst Environ 68:13–26
Kleijn D, Berendse F, Smit R, Gilissen N (2001) Agri-environment schemes do not effectively protect biodiversity in Dutch agricultural landscapes. Nature 413:723–725
Korpimaki E, Krebs CJ (1996) Predation and population cycles of small mammals. Bioscience 10:754–764
Krebs JR, Wilson JD, Bradbury RB, Siriwardena GM (1999) The second silent spring? Nature 400:611–612
Krebs CJ, Boonstra R, Boutin S, Sinclair ARE (2001) What drives the 10-year cycle of snowshoe hares? Bioscience 51:25–35
Lande US, Loe LE, Skjaerli OJ, Meisingset EL, Mysterud A (2014) The effect of agricultural land use practice on habitat selection of roe deer. Eur J Wildl Res 60:69–76
Lassini P, Monzani F, Pileri P (2007) A green vision for the renewal of the Lombardy landscape. In: Pedroli B, et al. (ed) Europe’s living landscapes. Essays on exploring our identity in the countryside. Zeist (NL): Landscape Europe/KNNV, pp 83–100
Laurance WF (1994) Rainforest fragmentation and the structure of small mammal communities in tropical Queensland. Biol Conserv 69:23–32
Lodé T (2000) Effect of a motorway on mortality of wildlife populations. Ambio 29:165–168
Lyra-Jorge MC, Ciocheti G, Pivello VR, Meirelles ST (2008) Comparing methods for sampling large- and medium-sized mammals: camera traps and track plots. Eur J Wildl Res 54:739–744
Mallick SA, Hocking GJ, Driessen MM (1998) Road-kills of the eastern barred bandicoot (Perameles gunni) in Tasmania: an index of abundance. Wildl Res 25:139–145
McGregor RL, Bender DJ, Fahrig L (2008) Do small mammals avoid roads because of the traffic? J Appl Ecol 45:117–123
Mech LD, Fritts SH, Radde GL, Paul WJ (1988) Wolf distribution and road density in Minnesota. Wildl Soc Bull 16:85–87
Meltofte H (2006) Wader populations at Zackenberg, high-arctic Northeast Greenland, 1996–2005. Dan Ornitol Foren Tidsskr 100:16–28
Morelle K, Lehaire F, Lejeune P (2013) Spatio-temporal patterns of wildlife-vehicle collisions in a region with a high-density road network. Nat Conserv 5:53–73
Neumann W, Ericsson G, Dettki H, Bunnefeld N, Keuler NS, Helmers DP, Radeloff VC (2012) Difference in spatiotemporal patterns of wildlife road-crossingsand wildlife-vehicle collisions. Biol Conserv 145:70–78
Pain DJ, Pienkowski MW (1997) Farming and Birds in Europe. Academic Press, London
Perco F (1987) Ungulati. Carlo Lorenzini, Udine
Perco F (2003) Capreolus capreolus. In: Boitani L, Lovari S, Vigna Taglianti A (eds) Fauna d’Italia: mammalia III, carnivora-artiodactyla. Calderini, Bologna, pp 305–326
Perco F, Calò CM (1994) The status of Roe Deer in Italy. In: Wotschikowsky U (ed) II European Roe Deer meeting, 27–30 October 1994, Brixen, pp 43–53
Pettett CE, Johnson PJ, Moorhouse TP, Macdonald DW (2018) National predictors of hedgehog Erinaceus europaeus distribution and decline in Britain. Mammal Rev 48:1–6
Pileri P, Sartori F (2004) Monitoring biodiversity at a wide land scale to support sustainable planning and policy: the proposal of a key indicator based on vegetation cover data deriving from maps. In: Marchetti M (ed) Monitoring and indicators of forest biodiversity in Europe—From ideas to operationality. Saarijarvi (FL): European Forest Institute Proc. 51, pp 455–463
Poulton SMC, Reeve NJ (2010) A pilot study of a method to monitor hedgehogs (Erinaceus europaeus). Mammal Notes, Autumn 2010
Prigioni C, Cantini M, Zilio A (2001) Atlante dei Maammiferi della Lombardia. Regione Lombardia e Università degli Studi di Pavia
Prigioni C, Smiroldo G, Remonti L, Balestrieri A (2009) Distribution and diet of reintroduced otters (Lutra lutra) on the River Ticino (NW Italy). Hystrix Ital J Mammal 20:45–53
Roos S, Johnston A, Noble D (2012) UK Hedgehog datasets and their potential for long-term monitoring. Research Report No. 598, pp 1–63
Sacchi O (2001) Sus scrofa. In: Prigioni C, Cantini M, Zilio A (eds) Atlante dei Mammiferi della Lombardia. Regione Lombardia e Università degli Studi di Pavia, pp 257–261
Saez-Royuela C, Telleria JL (1986) The increased population of the wild boar (Sus scrofa L.) in Europe. Mammal Rev 16:97–101
Santos SM, Marques JT, Lourenço A, Medinas D, Barbosa AM, Beja P, Mira A (2015) Sampling effects on the identification of roadkill hotspots: Implications for survey design. J Environ Manage 162:87e95
Schmeller DS, Henry PY, Julliard R, Gruber B, Clobert J, Dziock F, Lengyel S, Nowicki P, Deri E, Budrys E, Kull T, Tali K, Bauch B, Settele J, Van Swaay C, Kobler A, Babij V, Papastergiadou E, Henle K (2009) Advantages of volunteer-based biodiversity monitoring in Europe. Conserv Biol 23:307–316
Schmidt NM, Berg TB, Forchhammer MC, Hendrichsen DK, Kyhn LA, Meltofte H, Høye TT (2008) Vertebrate predator-prey interactions in a seasonal environment. Adv Ecol Res 40:345–370
Slater EM (2002) An assessment of wildlife road casualties: the potential discrepancy between numbers counted and numbers killed. Web Ecol 3:33–42
Smiddy P (2002) Bird and mammal mortality on roads in counties Cork and Waterford, Ireland. Bull Irish Biogeogr Soc 26:29–38
Spagnesi M (2002) Silvilago Sylvilagus floridanus. In: Spagnesi M, De Marinis AM (eds) Mammiferi d’Italia, Quaderni di Conservazione della Natura, No. 14, Ministero Ambiente–Istituto Nazionale Fauna Selvatica, Bologna, pp 156–157
Swann DE, Perkins N (2013) Inventory of Terrestrial Mammals in the Rincon Mountains Using Camera Traps. In: Gottfried GJ; Folliott PF, Gebow BS, Eskew LG, Collins LC (eds) Merging science and management in a rapidly changing world: Biodiversity and management of the Madrean Archipelago III, 2012 May 1–5, Tucson, AZ. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, USA
Taylor BD, Goldingay RL (2004) A survey of wildlife killed on three major roads in northeast New South Wales. Wildl Res 31:83–91
Tobler MW, Carrilo-Percastegui SE, Pitman RL, Mares R, Powell G (2008) An evaluation of camera traps for inventorying large-and-medium sized terrestrial rainforest mammals. Anim Conserv 11:169–178
Underhill JE, Angold PG (2000) Effects of roads on wildlife in an intensively modified landscape. Environ Rev 8:21–39
van der Ree R, Smith DJ, Grilo C (2015) Handbook of road ecology. Wiley, Chichester
van der Zee FF, Wiertz J, Terbraak CJF, Vanapeldoorn RC (1992) Landscape change as a possible cause of the badger Meles meles L. decline in the Netherlands. Biol Conserv 61:17–22
Vercayie D, Herremans M (2015) Citizen science and smartphones take roadkill monitoring to the next level. In: Seiler A, Helldin J-O (eds) Proceedings of IENE 2014 International Conference on Ecology and Transportation, Malmö, Sweden. Nature Conservation11:29–40
Vernesi C, Pecchioli E, Caramelli D, Tiedemann R, Randi E, Bertorelle G (2002) The genetic structure of natural and reintroduced roe deer (Capreolus capreolus) populations in the alps and Central Italy, with reference to the mitochondrial DNA phylogeography of Europe. Mol Ecol 11:1285–1297
Vidus-Rosin A, Gilio N, Meriggi A (2008) Introduced lagomorphs as a threat to “native” lagomorphs: The case of the Eastern cottontail (Sylvilagus floridanus) in northern Italy. In: Alves PC, Ferrand N, Hacklander K (eds) Lagomorph biology, evolution, ecology and conservation. Springer, Eidenberg, pp 153–165
Vidus-Rosin A, Montagna A, Meriggi A, Serrano S (2010) Density and habitat requirements of sympatric hares and cottontails in northern Italy. Hystrix, Ital J Mammal. https://doi.org/10.4404/hystrix-20.2-4440
Vidus-Rosin A, Lizier L, Meriggi A, Serrano S (2012) Habitat selection and segregation by two sympatric lagomorphs: the case of European hares (Lepus europaeus) and Eastern cottontails (Sylvilagus floridanus) in northern Italy. Acta Theriol. https://doi.org/10.1007/s13364-012-0084-y
Vignon V, Barbarreau H (2008) Collisions entre véhicules et ongulés sauvages: quel cout économique? Faune Sauvag 279:31–35
Wagner H, Fortin MJ (2005) Spatial analysis of landscapes: concepts and statistics. Ecology 86:1975–1987
White ER (2017) Minimum time required to detect population trends: the need for long-term monitoring programs. PeerJ Prepr. https://doi.org/10.7287/peerj.preprints.3168v3
Wilcox BA, Murphy DD (1985) Conservation strategy: effects of fragmentation on extinction. Am Nat 125:879–887
Williams AF, Wells JK (2005) Characteristics of vehicle-animal crashes in which vehicle occupants are killed. Traffic Inj Prev 6:56–59
Acknowledgements
We are grateful to the Province of Lodi for the necessary authorizations and logistical help during the research period. Special thanks to E. Maioli, A. Vanelli (Regione Lombardia-UTR Città Metropolitana di Milano, Via Haussmann, 7-26900 Lodi) and C. Consonni and his staff (Regione Lombardia-Direzione Generale Presidenza, P.zza di Lombardia, 1-20124 Milano) for sharing with us their data for data about, respectively, the Control Plan of both coypu and foxes in the Province of Lodi and ungulate road kills in Lombardy. P. Cristiani (Provincia di Pavia, Via Taramelli—27100 Pavia) provided data on daily vehicle traffic on the two monitored routes.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have neither conflict of interest nor funding to declare.
Additional information
Communicated by Dirk Sven Schmeller.
Rights and permissions
About this article

Cite this article
Canova, L., Balestrieri, A. Long-term monitoring by roadkill counts of mammal populations living in intensively cultivated landscapes. Biodivers Conserv 28, 97–113 (2019). https://doi.org/10.1007/s10531-018-1638-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-018-1638-3