
Abstract
Modern intensive farming caused pronounced changes to the European arable flora. Many species adapted to less intensive traditional farming declined severely, as did the potential of unsown arable vegetation to support higher trophic levels. To reverse these trends, various agri-environment measures were introduced. One such measure is to manage cereal headlands as conservation headlands, involving strict restrictions on pesticide and fertiliser use. An additional modification to management which could reduce crop competition and thus deliver benefits to arable plants is cereal sowing at reduced rates. However, little is known about its benefits to rare and declining arable plants, or to species of value to higher trophic levels, and whether it can be implemented without concomitant increase in undesirable weeds. We set up identical two-factorial experiments in winter wheat and spring barley, combining a nitrogen fertiliser versus no fertiliser treatment with cereal sowing at economic rates versus sowing at rates reduced by 75 %, with added sowing of a mixture of rare arable species. Both experiments also included an uncropped but cultivated control equivalent to another agri-environment measure. Our results show that reduced cereal sowing in conservation headlands can benefit rare and declining species, as well as arable plant diversity, without necessarily resulting in a concomitant increase in undesirable weeds. While such benefits tended to be larger in uncropped cultivated controls, conservation headlands have the advantage of not requiring land being taken out of production. Moreover, as shown in this study, their benefits to arable plants can be maximised by reduced sowing.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
From the late 1940s onwards, intensive methods of arable farming were rapidly adopted both in the UK (Robinson and Sutherland 2002) and in continental Europe (Stoate et al. 2001). Continued mechanisation, along with a substantial increase in the use of herbicides and fertilisers, facilitated a shift away from traditional mixed arable and livestock farming practices. These changes included a move towards simplified crop rotations which no longer included fallow periods, a shift from spring-sown to autumn-sown cereals (Stoate et al. 2001; Robinson and Sutherland 2002), and a shift from ploughing to non-inversion tillage (Chancellor et al. 1984; Cannell 1985; Morris et al. 2010). These in-field changes were accompanied by a reduction in non-productive landscape features and a trend of increasing farm size (Stoate et al. 2001; Robinson and Sutherland 2002).
In their entirety, these developments had profound effects on the non-crop arable flora. Many non-crop species characteristic of traditional arable management declined dramatically, both in the UK (Sutcliffe and Kay 2000; Potts et al. 2010) and across Europe (Hilbig and Bachthaler 1992a; Richner et al. 2015), and trait-based analyses (Storkey et al. 2010; Pinke and Gunton 2014) have provided insights in the underlying mechanisms of decline. At the same time, other species benefited from arable intensification, e.g. through their abilities to evolve herbicide resistance, to efficiently exploit high levels of nutrient availability, and to fit in with simplified cultivation and cropping regimes (Froud-Williams et al. 1983; Hilbig and Bachthaler 1992b; Hald 1999; Sutcliffe and Kay 2000), many of them becoming weeds. The net result of arable intensification was a steep decline in overall abundance and species diversity of non-crop plants, both in the UK (Sutcliffe and Kay 2000; Potts et al. 2010) and across Europe (Richner et al. 2015; Albrecht et al. in press). This is not only relevant in terms of the arable flora per se, with many traditional non-crop arable species now being threatened (Albrecht et al. in press), but also because of the important role that non-crop arable plants play as a food resource for higher trophic levels, including pollinating insects, other farmland invertebrates, and farmland birds (Wilson et al. 1999; Marshall et al. 2003; Franke et al. 2009; Bretagnolle and Gaba 2015). This role has also been demonstrated experimentally in studies showing that rigorous weed control adversely affects such higher trophic levels (Hawes et al. 2003).
The potential benefits of restrictions on agrochemical inputs into cropped field margins to arable plant diversity in general and to rare and threatened species in particular have been recognized since experiments investigating the benefits of reducing pesticide inputs to field headlands were done in the late 1970s and early 1980s, e.g. in Germany (Schumacher 1980) and in the UK (Boatman and Wilson 1988; Sotherton 1990). Similar experiments were subsequently carried out in Sweden (Chiverton 1994; Fischer and Milberg 1997) and in the Netherlands (de Snoo 1995; Kleijn and van der Voort 1997).
In England, in response to the findings of these early studies, conservation headlands were first made available as agri-environment scheme (AES) options as part of the Arable Stewardship Pilot Scheme launched in two regions in 1998 (MAFF 1998). In 2002, these options were extended nationwide as part of the Countryside Stewardship Scheme (Defra 2002). In both schemes, two kinds of cereal headland option were offered, both with similar restrictions on herbicide use, and one with an additional ban on fertiliser application. Assessments of both schemes indicated that non-fertilised conservation headlands tended to be characterised by higher non-crop plant cover and species richness than their fertilised counterparts (Critchley et al. 2004; Walker et al. 2007). However, compared to uncropped cultivated margins, another arable AES option designed to promote arable plants, conservation headlands, even when unfertilised, tended to deliver relatively small benefits for arable plant diversity and rare arable species, when compared against conventionally-managed cereal margins with no restrictions on agrochemical or fertiliser inputs (Critchley et al. 2004; Walker et al. 2007).
One further potential modification to the management of cereal headlands that might positively affect the size of benefit to arable biodiversity offered by conservation headlands is reduction in cereal sowing density, which is currently not promoted by the AES in Britain. Growth of uncompetitive rare arable species in cereal stands is positively related to light penetration levels, and these levels continuously decline while cereals grow, and form an increasingly dense canopy. Accordingly, the primary reason why rare arable weed species perform better in unfertilised cereal stands than in fertilised ones appears to be that a decline to critical threshold light penetration levels below which growth of rare arable species is restricted may occur more rapidly in fertilised stands, resulting in a reduced temporal window for growth of these species (Kleijn and van der Voort 1997). By potentially extending this temporal window, reduced cereal sowing densities may promote both rare as well as more common arable species. On the other hand, sowing of cereal at reduced densities means that more resources become available to individual cereal plants, and we do not know to what extent increased tillering (Kirby 1967; Champion et al. 1998), particularly at high levels of nutrient availability (Aspinall 1961), may counteract the effects of reduced sowing, and thus limit any expected benefit to rare arable species. Moreover, if such benefits can be achieved, it is important that they can be delivered without simultaneously boosting populations of agronomically undesirable species, i.e. weeds (Jones and Smith 2007). This is even more important given farmers’ concerns regarding potential infestations by such undesirable weeds (Still and Byfield 2007), which may in fact be at least partly responsible for the low uptake of UK AES options targeted at boosting rare arable plants (Clothier 2013).
Sowing cereals at reduced density has also recently been advocated in the context of reintroducing rare species by means of sowing (Epperlein et al. 2014). In many instances, such active reintroduction may be required, as natural recolonization may be highly unlikely, due to the fact that after extended periods of intensive management, rare species often are also no longer present in the local soil seed bank after having been lost from the vegetation, and due to many rare species lacking adaptations for dispersal (Albrecht et al. in press). A recent field study by Lang et al. (2016) showed that rare arable species can be reintroduced into cereal crops, provided the crop is managed sympathetically with respect to the needs of these species. Results from such studies also indicate that, as expected, yield losses due to reintroduction of uncompetitive rare species tend to be negligible (Epperlein et al. 2014; Lang et al. 2016).
High rates of establishment in the first year after reintroduction may be crucial for long-term persistence, and at the same time would help bring down the cost of sowing rare species, which can be significant, given that few such species are as yet commercially available (Albrecht et al. in press). Nonetheless, few studies have so far investigated whether using reduced cereal sowing rates could boost rare species establishment (but see Albrecht et al. 2014).
Furthermore, previous studies investigating the effects of reduced cereal sowing density have tended to ignore the potential effects of such a practice on species beneficial to arable faunal biodiversity and on agronomically undesirable species. In particular the latter aspect is important with respect to farmers’ acceptance of reduced sowing rates in cereal headlands to deliver benefits for biodiversity.
In this study, we use experimental manipulation in conventionally-managed cereal fields to address the following questions:
-
(1)
How do sowing rate of major winter and spring cereal crops and application of nitrogen fertiliser affect species richness and overall abundance of desirable and undesirable arable plant species in arable headlands managed for conservation?
-
(2)
What are the effects on establishment of sown rare arable species?
-
(3)
In terms of impacts on the arable flora, how do experimental cereal headlands compare with field margins managed as uncropped cultivated land?
Materials and methods
Experimental design
Our study was carried out at Roundwood Estate, in the Hampshire Downs, England (51°12′N, 1°17′W), in a typical arable landscape characterised by large arable fields with scattered woodland blocks and low hedgerows, with free-draining, thin chalky loams being the predominant soil type (Natural England 2014). A survey of arable plants carried out in 2009 confirmed a rich arable flora containing a large number of rare and declining arable species (Wilson 2010), making the estate a site of international importance for its arable flora according to Plantlife’s Important Arable Plant Area (IAPA) system (Byfield and Wilson 2005). Preliminary inspection of fields revealed low abundances of undesirable weed species, indicating high suitability for AES measures aimed at boosting rare and declining arable species.
To determine the effects of cereal sowing density and of N fertilisation on arable plant species in cereal headlands, separate randomised block experiments were set up in different fields for winter wheat and for spring barley. Accordingly, in the following, these experiments will be referred to as the winter wheat experiment and the spring barley experiment. Both experiments were set up along headlands 12 m wide, and each experiment consisted of four replicate blocks with five treatments. Of these treatments, four corresponded to a 2 × 2 factorial design, with the two factors being cereal sowing density (sown at a standard density falling within the range of recommended densities for achieving optimum yield vs sown at 25 % of this standard density) and N fertilisation (liquid N fertiliser applied at rates typically used for these two respective crops on this type of soil vs no N fertiliser). As shown by crop trials, compared to sowing at a standard density, sowing at 25 % of that density can be carried out with minimal yield loss, provided timely sowing is ensured (Kirby 1967; Spink et al. 2000). The fifth treatment was an uncropped control with no cereal sowing and no fertilisation conforming to the cultivated, uncropped field margin management prescription of the English agri-environment scheme. The length of experimental plots was 10 m for treatments not receiving N fertiliser, and, due to operational requirements for fertiliser spreading, 20 m for treatments receiving N fertiliser.
In the winter wheat experiment, on 27 September 2013, seeds of winter wheat var. Horatio were drilled to a depth of 2.5 cm, using rates of 320 seeds m−2 and of 80 seeds m−2, respectively, on standard-density and quarter-density plots. In the spring barley experiment, on 14 March 2014, spring barley var. Concerto was drilled to a depth of 2.5 cm, using rates of 350 seeds m−2 and of 88 seeds m−2, respectively, on standard-density and quarter-density plots.
Pre-drilling, all treatment plots were cultivated using a Knight triple press, and phosphorus and potassium fertiliser was applied at fixed rates of 50 kg P ha−1 and 60 kg K ha−1. On the same day as the cereals were drilled, seed mixtures containing five rare arable annual species—Kickxia spuria, Lithospermum arvense, Papaver argemone, Scandix pecten-veneris, and Silene noctiflora (nomenclature follows Stace 2010)—were sown into the central 5 m × 5 m area of each plot. Sowing rates varied among species, ranging between 30 seeds m−2 for the largest-seeded species, S. pecten-veneris, and 285 seeds m−2 for the smallest-seeded species, P. argemone, with the remaining species sown at 150 seeds m−2. A similar rationale, based on the assumption of a positive correlation between seed size and establishment probability, has also been applied by e.g. Lang et al. (2016). All of these species have been reported to occur at the estate, albeit generally only locally and at low densities (Wilson 2010). Liquid fertiliser was applied to the growing cereal crop to treatment plots designated to receive N fertiliser, at rates of 240 kg ha−1 N and 48 kg ha−1 S to winter wheat, and of 130 kg ha−1 N and 22 kg ha−1 S to spring barley respectively, in line with regular practice at the estate when growing these crops.
Data collection
A single count of all sown rare arable species, combined with a vegetation survey of all arable plants, was carried out between 28 July and 30 July 2014, just before the harvest of winter wheat and spring barley. Recording was carried out in the central 5 m × 5 m area of each treatment plot in five 0.5 m × 0.5 m quadrats that were placed in a regular pattern, one in the centre of the plot, and the other four in the centres of each of four 2.5 m × 2.5 m quadrants.
In each quadrat, the following parameters were recorded: (1) numbers of individuals of each sown species; (2) total number of cereal tillers; (3) vegetation height, using the drop disc method (Stewart et al. 2001); and (4) vegetation composition, by estimating percent visual cover of all species rooting in the quadrat and of bare ground.
Species classification
The herbaceous non-crop species encountered during vegetation recording were classified into four mutually exclusive groups: (1) common species that are of potential value to the fauna of arable habitats and that are not considered undesirable by farmers; (2) common species considered undesirable, irrespective of their potential value to the fauna; (3) specifically arable species that are rare and/or declining, irrespective of their potential value to the fauna; and (4) common species thought to be of only limited benefit to the fauna and that are not considered undesirable (=‘neutral’ species). Regarding the group of undesirable species, we mostly followed the list of common pernicious weeds by Storkey and Westbury (2007), but added two competitive species encountered in our experiments, Dactylis glomerata and Urtica dioica. Some of the species considered undesirable—in particular Cirsium arvense, Cirsium vulgare, Rumex crispus, Senecio jacobaea, and Urtica dioica—can deliver considerable potential benefit to farmland birds and invertebrates. Nonetheless, for two reasons, a classification into mutually exclusive groups appeared preferable over assigning these species to multiple groups. Firstly, as the aforementioned species are tall-growing and do not flower in the first year after establishment, any benefits specifically to pollinating insects (via nectar and pollen) and to farmland birds (mostly via seeds) are highly unlikely to materialize within a single year after cereal sowing, even in the case of overwintering stubble. Secondly, as this study is concerned specifically with the management of arable headlands in keeping with AES options whose uptake by the farming community is noticeably affected by farmers’ concerns over infestations by undesirable weeds (Clothier 2013), it appeared expedient to focus specifically on the potential benefits to arable fauna brought about by those species other than undesirable weeds.
Regarding the plant species potentially benefitting faunal biodiversity, we considered three different aspects of potential value: (1) to phytophagous insects; (2) to farmland birds; and (3) to insect pollinators. Value to phytophagous insects was assessed on the basis of numbers of unique species-level interactions in the Database of British Insects and their Foodplants (DBIF; available online at http://www.brc.ac.uk/dbif/ and accessed on 8 April 2016; see Smith and Roy 2008). Value to farmland birds was assessed at the genus level, based on information in the review by Holland et al. (2006). Value to insect pollinators was primarily assessed on the basis of a recently published nectar database (Baude et al. 2016), under additional consideration of a plant species’ ability to provide nutritionally valuable pollen collected by insect pollinators, based on the literature (Carvell et al. 2006; Hanley et al. 2008; Kleijn and Raemakers 2008). Rare and declining species were identified on the basis of their IAPA score according to Byfield and Wilson (2005), including all species that had received a rating on the scale from 1 (=of local concern) to 9 (=critically endangered according to Cheffings and Farrell 2005).
Results of the classification of 61 non-crop herbaceous species encountered in the winter wheat and spring barley experiments are summarised in Table 1. For a more detailed description of classification criteria and species-level ratings see Table S1. While these ratings suggest that none of the species classified as rare or declining are of notable benefit to faunal biodiversity, this may, to some extent, reflect the paucity of evidence available for rarer species, as e.g. numbers of interactions in the DBIF tend to be positively correlated with the commonness of species (Smith and Roy 2008).
Statistical analyses
Prior to analyses, count data was summed up at plot level across the five sampled 0.5 m × 0.5 m quadrats, and cover and vegetation height data was averaged. Average cover values were arcsine-transformed (Crawley 2007). Average vegetation height was Box-Cox-transformed, with optimal coefficients for transformation being estimated using spread-level plots as provided in the ‘car’ package v 2.0-12 (Fox and Weisberg 2011) within R v 2.15.1 (R Foundation for Statistical Computing, Vienna, Austria).
Disregarding the control treatment, both experiments conformed to a two-factorial design of cereal sowing density versus nitrogen fertiliser application. Accordingly, we analysed data from each experiment in two ways, (1) as two-factorial design including only treatments involving cereal sowing, and (2) as one-way design including the uncropped cultivated control treatment. For two-factorial analyses, cereal sowing density and nitrogen fertiliser application, along with their two-way interaction, were specified as fixed factors, and block was included as random effect. For one-way analyses, treatment, consisting of five levels, was specified as the sole fixed factor, and block as a random effect. In one-way analyses, in case of a significant treatment effect, pairwise comparisons were carried out using Dunnett tests with Dunnett-Hsu adjustment to investigate differences between the control treatment and each of the four other treatments.
Depending on the type of data, one of two kinds of statistical model was used. Total cover of all non-crop vegetation, summed cover of undesirable species, summed cover of species beneficial for faunal biodiversity, summed cover of rare and declining species, vegetation height and bare ground cover were analysed with linear mixed models (LMM), using Proc MIXED in SAS 9.3 for Windows (SAS Institute Inc., Cary, NC, USA). In contrast, count parameters—including cereal tiller density, total species richness, richness of species of faunal value, richness of rare and declining species with an IAPA score ≥1, and numbers of established plants of sown rare arable species—were analysed with generalized linear mixed models (GLMM) and Poisson errors, using Proc GLIMMIX in SAS 9.3 for Windows (SAS Institute Inc., Cary, NC, USA). Regarding the numbers of established plants of sown species, analyses were carried out for aggregate numbers of plants across all sown species, and individually for those species for which establishment was sufficiently high for allowing successful convergence of the iterative GLMM modelling approach.
Results
Detailed results of two-factorial analyses of the effects of N fertilisation and cereal sowing density are presented in Table S2, with significant results presented below. Results of one-factorial analyses are indicated by asterisks in Figs. 1, 2, 3, and 4, showing which individual cereal headland treatments differ significantly from uncropped controls.

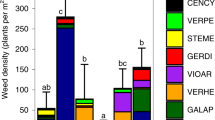
Summed percent cover of a, b all non-crop species, c, d species of faunal value, e, f rare and declining arable species, and g, h undesirable weeds. Left panel winter wheat experiment; right panel spring barley experiment. Back-transformed mean ± SE shown for different combinations of cereal sowing at standard (‘1/1’) versus one-quarter of standard (‘1/4’) densities and N application at typical rates (‘+N’) versus no N (‘−N’), and also for an uncropped cultivated control treatment (‘Control’). Asterisks indicate significant pairwise differences between individual cereal-sown treatments and the uncropped control (Dunnett tests: * 0.01 ≤ P < 0.05; ** 0.001 ≤ P < 0.01 *** P < 0.001)

Species richness per 1.25 m2 of a, b all non-crop species, c, d species of faunal value, and e, f rare and declining arable species. Mean ± SE shown. See also caption and legend of Fig. 1

Establishment of sown rare arable species in terms of numbers of plants per m2 of a, b all sown species pooled together and c, d K. spuria. Mean ± SE shown. See also caption and legend of Fig. 1

Vegetation structural parameters, including a, b percentage bare ground, c, d cereal tiller density, as number of tillers per m2, and e, f vegetation height in metres. Mean ± SE shown. See also caption and legend of Fig. 1
Summed cover and species richness
N fertilisation had a negative effect on summed cover of arable plant species in the spring barley experiment (F1,9 = 5.24; p = 0.048; see trend in Fig. 1b), but not in the winter wheat experiment (Table S2). However, negative effects of fertilisation on arable plant species richness were manifest in both experiments, but were more pronounced in spring barley (F1,9 = 40.59; p < 0.001; see trend in Fig. 2b) than in winter wheat (F1,9 = 11.54; p = 0.008; see trend in Fig. 2a). For spring barley, there was a significant interaction with sowing density (F1,9 = 9.04; p = 0.015), in that the negative effect of fertilisation was more pronounced at the standard rate of sowing than at the reduced rate (see trend in Fig. 2b). In contrast, in winter wheat, sowing density affected species richness independently of fertiliser application, as indicated by a significant main effect (F1,9 = 5.52; p = 0.043), with slightly higher richness at reduced sowing density (see trend in Fig. 2a).
No significant treatment effects were detected regarding summed cover of species of faunal value (Table S2). However, in spring barley, species richness of this group was highly significantly affected by N fertilisation (F1,9 = 14.99; p = 0.004), with a significant interaction (F1,9 = 5.85; p = 0.039) indicating that that this effect was more pronounced at the standard rate of sowing than at the reduced rate (see trend in Fig. 2d). In contrast, in winter wheat, N fertilisation fell short of affecting species richness of this group (F1,9 = 5.10; p = 0.050).
In winter wheat, rare and declining arable species benefited from reduced cereal sowing rates both in terms of summed cover (F1,9 = 5.64; p = 0.042; see trend in Fig. 1e) as well as species richness (F1,9 = 10.21; p = 0.011; Fig. see trend in 2e), whereas in spring barley, such an effect was only observed for rare species richness (F1,9 = 6.89; p = 0.028; see trend in Fig. 2f). N application, on the other hand, had a strong negative effect on rare species in spring barley, both in terms of their summed cover (F1,9 = 11.71; p = 0.008; see trend in Fig. 1f) and species richness (F1,9 = 14.08; p = 0.004; see trend in Fig. 2f), but had no significant effect on either parameter in winter wheat.
Summed cover of undesirable weeds was very low in both experiments, and was not significantly affected by experimental treatments regardless of cereal sown (Table S2; see also Fig. 1g, h).
Establishment of sown rare species
Numbers of established plants pooled across sown species were not affected by the experimental treatments in winter wheat (Table S2), where establishment was generally low. However, in spring barley, where overall establishment was somewhat higher, a significant negative effect of N application was found (F1,9 = 9.14; p = 0.014; see trend in Fig. 3b). For individual species, establishment was generally poor, with the exception of Kickxia spuria, whose plants made up about 2/3 of all recorded individuals (226 out of a total of 343). However, no significant treatment effects were found for this species (Table S2). Papaver argemone was characterised by sporadic establishment, with higher establishment in the spring barley experiment, and the three remaining species, Scandix pecten-veneris, Silene noctiflora and Lithospermum arvense, had very low establishment. For all four species, establishment was too low to allow statistical analysis. For a more detailed breakdown of establishment at species level see Table S3.
Vegetation structure
Effects of experimental treatments on bare ground cover were only observed in the winter wheat experiment, where levels were strongly reduced by N fertilisation (F1,9 = 18.36; p = 0.002; see trend in Fig. 4a). Effects of cereal sowing rate on wheat tiller density were still detectable just before harvest, i.e. lower tiller densities were observed in plots sown at the reduced rate (F1,9 = 20.13; p = 0.002). In contrast, no such differences were detectable in spring barley (F1,9 = 0.09; p = 0.769). N application, on the other hand, had a much more pronounced effect on tiller density in spring barley (F1,9 = 16.15; p = 0.003; see trend in Fig. 4d) than in winter wheat (F1,9 = 5.48; p = 0.044; see trend in Fig. 4c). Vegetation height was strongly increased in both crops by N application (winter wheat: F1,9 = 37.77; p < 0.001; spring barley: F1,9 = 348.60; p < 0.001; see trends in Fig. 4e, f). Reduction of cereal sowing rate was associated with a small but nonetheless significant increase in vegetation height in spring barley (F1,9 = 6.83; p = 0.028; see trend in Fig. 4f).
Comparison of cereal headland treatments with the uncropped cultivated treatment
In the spring barley experiment, summed cover of all arable plant species and of rare and declining species was generally higher in uncropped cultivated control plots than in either of the cereal headland treatments, with significant differences being highlighted in Fig. 1b and f. With the exception of the cereal headland treatment involving cereal sowing at the standard rate in the absence of N application, this was also the case for species of faunal value, as highlighted in Fig. 1d. Generally, many fewer significant pairwise differences between the uncropped cultivated treatment and individual cereal headland treatments were found in the winter wheat experiment (left-hand side of Fig. 1), where, compared to the spring barley experiment (right-hand side of Fig. 1), summed cover generally tended to be higher in cereal-sown plots, particularly in the case of arable species in general and of species of faunal value, as shown in the top two rows of Fig. 1.
Species richness not just of non-crop species in general, but also of species of faunal value and of rare and declining species, was generally not higher on uncropped cultivated plots than on cereal-sown plots, with the exception of spring barley plots sown at the standard rate and receiving N fertiliser, as highlighted for each respective group in Fig. 2b, d, and f.
Similarly, a comparison of sown species establishment pooled across all sown species, between uncropped cultivated treatment and the cereal headland treatments, yielded only a single pairwise difference between a headland treatment and the uncropped treatment, again for spring barley sown at the standard rate and receiving N fertiliser, as highlighted in Fig. 3b.
Structurally, uncropped cultivated control plots differed markedly from cereal headland plots, with vegetation height and, for obvious reasons, cereal tiller density, being markedly lower in the former, as highlighted in Fig. 4c–f. On the other hand, as highlighted in Fig. 4a and b, levels of bare ground just before harvest tended to be roughly similar between uncropped controls and cereal-sown headland treatments.
Discussion
Conservation headlands involving restrictions to agrochemical and fertiliser inputs have the distinctive advantage of providing ecosystem services and supporting rare and declining arable plants without requiring land being taken out of food production (Albrecht et al. in press). This study shows that reduced rates of cereal sowing in such headlands can help boost extant populations of rare and declining arable species, both in terms of total cover as well as in terms of species richness, without necessarily resulting in a pronounced increase in undesirable weeds, if levels of the latter are low to begin with.
These positive effects of a reduction in cereal sowing density on extant rare arable species appeared to be more pronounced in winter wheat. In contrast, fertilisation had pronounced negative effects in spring barley, but not in winter wheat. As evidenced by treatment effects on tiller density, these crop-specific differences in treatment effect on rare species may at least partly have been the result of intrinsic differences between the tested cereals crops in terms of their ability to respond to higher resource availability at the level of individual plants—brought about either by reduced sowing or by added nitrogen—with increased tillering. Such a tillering response was generally more pronounced in the spring barley experiment than in the winter wheat experiment. Unlike the wheat crop, the barley crop compensated perfectly for reduced sowing by increased tillering. Similarly, the tiller density increase in response to N application was more pronounced in barley than in wheat.
On the other hand, while the observed differences between cereals in tillering thus appear to support an explanation of crop-specific responses to treatments, seasonal differences in the timing of cultivation between the two crops may have also have contributed to our results. Many rare arable species show seasonal preferences in terms of emergence (Wilson 1994; Pywell et al. 2010), and the same applies to common species, resulting in marked effects of cultivation season on floristic composition (Hald 1999; Critchley et al. 2006).
Nonetheless, in the case of N fertilisation, similar crop-specific effects on the weed flora to the ones found by us were found by Bischoff and Mahn (2000). In their three-year study on a long-term crop-rotation experiment, peak weed densities were significantly lower on plots receiving N fertiliser than on plots not receiving N in the year when spring barley was planted, whereas in the year when winter wheat was planted, the opposite was the case, indicating that weed densities in spring barley, but not in winter wheat, were suppressed by N application.
Management for rare and declining species
Few insights were possible based on the sowing component of our experiments due to the sporadic establishment of all but one sown rare species, although we found that, pooled across species, in spring barley, establishment was significantly reduced by N application. However, potentially due to this sporadic establishment, we failed to establish any potential effects of reduced cereal sowing density. Recent work by Albrecht et al. (2014) has shown that establishment of rare arable species can indeed be bolstered by sowing cereals at reduced rate, although they tested other cereals in their study, i.e. rye and spelt. The results of previous studies suggest that reduced cereal sowing can also boost size (Svensson and Wigren 1982; Kleijn and van der Voort 1997; Albrecht et al. 2014) and per capita seed production (Peters and Gerowitt 2014) of rare arable plant individuals. In the present study, additional insights regarding the effects of cereal sowing rate on rare and declining species were obtained from analyses of summed cover and species richness of the group of rare and declining arable species found in the experiment, including both unsown and sown species. These analyses demonstrated that reduced cereal sowing rate increased both summed cover and species richness of rare and declining arable species in winter wheat, but only species richness in spring barley.
Regarding the effects of N application, in line with the observed reduction in establishment of sown rare species, analyses both of summed cover and species richness of rare and declining species in spring barley also indicated a strong negative effect of N fertilisation. However, no such effects were detected in the winter wheat experiment.
Taken together, these findings appear to suggest that N application may affect rare and declining species more strongly in spring barley than in winter wheat. However, it is important to keep in mind that these were two separate experiments carried out in different fields. As indicated by the uncropped control treatments in both experiments, which provide an indication of potential maximum values of summed cover and species richness of rare and declining species as well as of sown species establishment, higher maxima for all three variables occurred in spring barley than in winter wheat. This suggests that it may have been more difficult to detect significant N application effects on rare and declining species in winter wheat than in spring barley. Moreover, negative effects on N application on rare species establishment in winter wheat have been demonstrated by previous studies (e.g. Wilson 1999), and our results should thus not be interpreted as a challenge to the perceived wisdom of N application negatively affecting populations of rare arable species in cereal crops.
Management for plant and faunal diversity
Our study failed to produce evidence for reduced rates of cereal sowing to result in increased cover of common arable species of faunal value that are expected to support higher trophic groups. However, reduced sowing of winter wheat resulted in slightly more species-rich arable vegetation, which could mean resource provision for a wider range of fauna (Meek et al. 2002; Asteraki et al. 2004).
In contrast, there was strong evidence for beneficial effects of not applying N fertiliser on non-crop plant diversity, both in terms of overall species richness, as well as in terms of richness of species of faunal value. Both in the winter wheat experiment and in the spring barley experiment, and in agreement with findings from a comparative study of arable options in a previous English AES (Walker et al. 2007), non-crop species richness was much higher in cereal stands not receiving N fertiliser than in those with N application. The same clearly applied to species of faunal value in spring barley, and a similar effect in winter wheat bordered on significance. Again, such positive treatment effects on plant species richness may be indicative of an increased ability of the plant cover to support higher faunal diversity (Meek et al. 2002; Asteraki et al. 2004). Matching these findings for species richness, N application resulted in reduced summed cover of non-crop species in the spring barley experiment, but not in the winter wheat experiment.
In agreement with our findings, Kleijn and van der Voort (1997) demonstrated a clear negative relationship between N application and light penetration beneath the canopy of barley stands, whereas previous studies carried out in wheat crops have shown that N fertilisation can boost both establishment (Bischoff and Mahn 2000) and total biomass of weeds (Rial-Lovera et al. 2016). Together with these findings by other authors, our results suggest that effects of N application on the arable flora may vary between different types of cereal, e.g. being potentially more detrimental in spring barley than in winter wheat.
Summed cover of species of biodiversity value remained unaffected by N application, irrespective of cereal sown. While, due to comparatively low replication, our ability to detect such effects may have been somewhat limited, our findings indicate that restricting N application does not necessarily lead to marked increases in resource provision to arable fauna. In fact, e.g. in wheat crops, certain potentially beneficial species may respond positively to N fertilisation (Rial-Lovera et al. 2016).
Comparison of cereal headlands with uncropped cultivated controls
In spring barley, summed cover of arable species, as well as of species of faunal value and of rare and declining species, tended to be higher in the uncropped cultivated control treatment than in the various cereal headland treatments. However, no such effects were found for overall species richness, for richness species of faunal value, or of rare and declining species, except for comparing uncropped controls with the most intensively-managed type of headland which received N fertiliser and was sown at standard density. In contrast, in winter wheat, there were hardly any differences between control plots and the various types of cereal headland treatments, suggesting that cereal exerted much stronger competitive effects on the non-crop vegetation in the spring barley experiment than in the winter wheat experiment. While these results also fit with the observation of higher compensatory tillering in spring barley, providing an explanation in terms of more intense competition, it is again important to keep in mind that timing of cultivation may also have affected species composition, e.g. due to seasonal preferences for emergence.
Conclusions
In this study, we have demonstrated the potential benefits of reduced cereal sowing densities to rare and declining arable species in conservation headlands, and we have confirmed similar benefits from restrictions in N fertiliser application. However, as suggested by the crop-specific results both in the winter wheat and spring barley experiments, the relative extent of such benefits may vary between different types of cereal, e.g. in relation to attributes such as tillering capacity or crop height (Andrew et al. 2015), or e.g. depending on season of sowing. Thus, building on the findings of this study, further experiments investigating these aspects of managing conservation headlands should focus on establishing under which conditions such management modifications may deliver the greatest benefit to rare and declining arable plants.
References
Albrecht H, Prestele J, Altenfelder S, Wiesinger K, Kollmann J (2014) New approaches to the conservation of rare arable plants in Germany. In: Nordmeyer H, Ulber L (eds) Delivering arable biodiversity. Proceedings 26th German Conference on Weed Biology and Weed Control March 11–13, 2014, Braunschweig. Bundesforschungsinstitut für Kulturpflanzen, Braunschweig, pp 180–189. http://pub.jki.bund.de/index.php/JKA/article/download/2864/3057 Accessed 20 April 2016
Albrecht H, Cambecèdes J, Lang M, Wagner M (in press) Management options for rare arable plants in Europe. Bot Lett 164(4). doi:10.1080/23818107.2016.1237886
Andrew IKS, Storkey J, Sparkes DL (2015) A review of the potential for competitive cereal cultivars as a tool in integrated weed management. Weed Res 55:239–248. doi:10.1111/wre.12137
Aspinall D (1961) The control of tillering in the barley plant. 1. The pattern of tillering and its relation to nutrient supply. Aus J Biol Sci 14:493–505. doi:10.1071/BI9610493
Asteraki EJ, Hart BJ, Ings TC, Manley WJ (2004) Factors influencing the plant and invertebrate diversity of arable field margins. Agr Ecosyst Environ 102:219–231. doi:10.1016/j.agee.2003.07.003
Baude M, Kunin WE, Boatman ND, Conyers S, Davies N, Gillespie MAK, Morton D, Smart SM, Memmott J (2016) Historical nectar assessment reveals the fall and rise of floral resources in Britain. Nature 530:85–88. doi:10.1038/nature16532
Bischoff A, Mahn E-G (2000) The effects of nitrogen and diaspore availability on the regeneration of weed communities following extensification. Agr Ecosyst Environ 77:237–246. doi:10.1016/S0167-8809(99)00104-8
Boatman ND, Wilson PJ (1988) Field edge management for game and wildlife conservation. In: The practice of weed control and vegetation management in forestry, amenity and conservation areas. Aspects of applied biology, vol 16. Association of Applied Biologists, Wellesbourne, pp 89–94
Bretagnolle V, Gaba S (2015) Weeds for bees? A review. Agron Sustain Dev 35:891–909. doi:10.1007/s13593-015-0302-5
Byfield AJ, Wilson PJ (2005) Important Arable Plant Areas: identifying priority sites for arable plant conservation in the United Kingdom. Plantlife International, Salisbury. http://www.plantlife.org.uk/uploads/documents/Important_Arable_Plant_Areas.pdf. Accessed 20 Apr 2016
Cannell RQ (1985) Reduced tillage in north-west Europe—a review. Soil Till Res 5:129–177. doi:10.1016/0167-1987(85)90028-5
Carvell C, Westrich P, Meek WR, Pywell RF, Nowakowski M (2006) Assessing the value of annual and perennial forage mixtures for bumblebees by direct observation and pollen analysis. Apidologie 37:326–340. doi:10.1051/apido:2006002
Champion GT, Froud-Williams RJ, Holland JM (1998) Interactions between wheat (Triticum aestivum L.) cultivar, row spacing and density and the effect on weed suppression and crop yield. Ann Appl Biol 133:443–453. doi:10.1111/j.1744-7348.1998.tb05842.x
Chancellor RJ, Fryer JD, Cussans GW (1984) The effects of agricultural practices on weeds in arable land. In: Jenkins D (ed) Agriculture and the environment—proceedings of ITE symposium no. 13 held at Monks Wood Experimental Station on 28–29 February and 1 March 1984. Institute of Terrestrial Ecology, Cambridge, pp 89–94
Cheffings CM, Farrell L (2005) The vascular plant Red Data List for Great Britain. Species Status No. 7. Joint Nature Conservation Committee, Peterborough
Chiverton PA (1994) Large-scale field trials with conservation headlands in Sweden. In: Boatman N (ed) Field margins: integrating agriculture and conservation. BCPC Monograph No. 58. British Crop Production Council, Farnham, pp 89–94
Clothier L (2013) Campaign for the farmed environment: entry level stewardship option uptake. Defra Agricultural Change and Environment Observatory Research Report No. 32. Department for Environment, Food and Rural Affairs, London. https://www.gov.uk/government/uploads/system/uploads/attachment_data/file/183937/defra-stats-foodfarm-environ-obs-research-setaside-farmenviroment-ELSinCFEjan13-130214.pdf. Accessed 20 April 2016
Crawley MJ (2007) The R book. Wiley, Chichester
Critchley CNR, Allen DS, Fowbert JA, Mole AC, Gundrey AL (2004) Habitat establishment on arable land: assessment of an agri-environment scheme in England, UK. Biol Conserv 119:429–442. doi:10.1016/j.biocon.2004.01.004
Critchley CNR, Fowbert JA, Sherwood AJ (2006) The effects of annual cultivation on plant community composition of uncropped arable field boundary strips. Agr Ecosyst Environ 113:196–205. doi:10.1016/j.agee.2005.05.013
de Snoo GR (1995) Unsprayed field margins: implications for environment, biodiversity and agricultural practice. PhD thesis, University of Leiden
Defra (2002) The Countryside Stewardship Scheme. New arable options from 2002. Department for Environment, Food and Rural Affairs, London
Epperlein LRF, Prestele JW, Albrecht H, Kollmann J (2014) Reintroduction of a rare arable weed: competition effects on weed fitness and crop yield. Agr Ecosyst Environ 188:57–62. doi:10.1016/j.agee.2014.02.011
Fischer A, Milberg P (1997) Effects on the flora of extensified use of field margins. Swed J Agr Res 27:105–111
Fox J, Weisberg S (2011) An R companion to applied regression, 2nd edn. Sage Press, Thousand Oaks
Franke AC, Lotz LAP, van der Burg WJ, van Overbeek L (2009) The role of arable weed seeds for agroecosystem functioning. Weed Res 49:131–141. doi:10.1111/j.1365-3180.2009.00692.x
Froud-Williams RJ, Drennan DSH, Chancellor RJ (1983) Influence of cultivation regime on weed floras of arable cropping systems. J Appl Ecol 20:187–197. doi:10.2307/2403385
Hald AB (1999) The impact of changing the season in which the cereals are sown on the diversity of the weed flora in rotational fields in Denmark. J Appl Ecol 36:24–32. doi:10.1046/j.1365-2664.1999.00364.x
Hanley ME, Franco M, Pichon S, Darvill B, Goulson D (2008) Breeding system, pollinator choice and variation in pollen quality in British herbaceous plants. Funct Ecol 22:592–598. doi:10.1111/j.1365-2435.2008.01415.x
Hawes C, Haughton AJ, Osborne JL, Roy DB, Clark SJ, Perry JN, Rothery P, Bohan DA, Brooks DR, Champion GT, Dewar AM, Heard MS, Woiwod IP, Daniels RE, Young MW, Parish AM, Scott RJ, Firbank LG, Squire GR (2003) Responses of plants and invertebrate trophic groups to contrasting herbicide regimes in the Farm Scale Evaluations of genetically modified herbicide–tolerant crops. Philos T R Soc B 358:1899–1913. doi:10.1098/rstb.2003.1406
Hilbig W, Bachthaler G (1992a) Wirtschaftsbedingte Veränderungen der Segetalvegetation in Deutschland im Zeitraum von 1950–1990. I: Entwicklung der Aufnahmeverfahren, Verschwinden der Saatunkräuter, Rückgang von Kalkzeigem, Säurezeigern, Feuchtezeigern, Zwiebel- und Knollengeophyten, Abnahme der Artenzahlen. Angew Bot 66:192–200
Hilbig W, Bachthaler G (1992b) Wirtschaftsbedingte Veränderungen der Segetalvegetation in Deutschland im Zeitraum von 1950–1990. II: Zunahme herbizidverträglicher Arten, nitrophiler Arten, von Ungräsern, vermehrtes Auftreten von Rhizom- und Wurzelunkräutern, Auftreten und Ausbreitung von Neophyten, Förderung gefährdeter Ackerwildkrautarten, Integrierter Pflanzenbau. Angew Bot 66:201–209
Holland JM, Hutchison MAS, Smith B, Aebischer NJ (2006) A review of invertebrates and seed-bearing plants as food for farmland birds in Europe. Ann Appl Biol 148:49–71. doi:10.1111/j.1744-7348.2006.00039.x
Jones NE, Smith BM (2007) Effects of selective herbicide treatment, row width and spring cultivation on weed and arthropod communities in winter wheat. In: Boatman N, Bradbury R, Critchley N, Holland J, Marshall J, Ogilvy S (eds) Delivering arable biodiversity. Aspects of applied biology, vol 81. Association of Applied Biologists, Wellesbourne, pp 39–46
Kirby EJM (1967) The effect of plant density upon the growth and yield of barley. J Agr Sci 68:317–324. doi:10.1017/S0021859600012806
Kleijn D, Raemakers I (2008) A retrospective analysis of pollen host plant use by stable and declining bumblebees. Ecology 89:1811–1823. doi:10.1890/07-1275.1
Kleijn D, van der Voort LAC (1997) Conservation headlands for rare arable weeds: the effects of fertilizer application and light penetration on plant growth. Biol Conserv 81:57–67. doi:10.1016/S0006-3207(96)00153-X
Lang M, Prestele J, Fischer C, Kollmann J, Albrecht H (2016) Re-introduction of rare arable plants by seed transfer. What are the optimal sowing rates? Ecol Evol 6:5506–5516. doi:10.1002/ece3.2303
MAFF (1998) The Countryside Stewardship Scheme. Arable Stewardship. Information and how to apply. Ministry of Agriculture, Fisheries and Food, London
Marshall EJP, Brown VK, Boatman ND, Lutman PJW, Squire GR, Ward LK (2003) The role of weeds in supporting biological diversity within crop fields. Weed Res 43:77–89. doi:10.1046/j.1365-3180.2003.00326.x
Meek B, Loxton D, Sparks T, Pywell R, Pickett H, Nowakowski M (2002) The effect of arable field margin composition on invertebrate biodiversity. Biol Conserv 106:259–271. doi:10.1016/S0006-3207(01)00252-X
Morris NL, Miller PCH, Orson JH, Froud-Williams RJ (2010) The adoption of non-inversion tillage systems in the United Kingdom and the agronomic impact on soil, crops and the environment—a review. Soil Till Res 108:1–15. doi:10.1016/j.still.2010.03.004
Natural England (2014) National Character Area Profile: 130. Hampshire Downs. Natural England Publications. http://publications.naturalengland.org.uk/file/4546775574642688. Accessed 20 Apr 2016
Peters K, Gerowitt B (2014) Response of the two rare arable weed species Lithospermum arvense and Scandix pecten-veneris to climate change conditions. Plant Ecol 215:1013–1023. doi:10.1007/s11258-014-0358-3
Pinke G, Gunton RM (2014) Refining rare weed trait syndromes along arable intensification gradients. J Veg Sci 25:978–989. doi:10.1111/jvs.12151
Potts GR, Ewald JA, Aebischer NJ (2010) Long-term changes in the flora of the cereal ecosystem on the Sussex Downs, England, focusing on the years 1968–2005. J Appl Ecol 47:215–226. doi:10.1111/j.1365-2664.2009.01742.x
Pywell RF, Hulmes L, Meek WR, Nowakowski M (2010) Practical management of scarce arable populations. In: Boatman N, Green M, Holland J, Marshall J, Renwick A, Siriwardena G, Smith B, de Snoo G (eds) Agri-environment schemes—what have they achieved and where do we go from here? Aspects of applied biology, vol 100. Association of Applied Biologists, Wellesbourne, pp 175–182
Rial-Lovera K, Davies WP, Cannon ND, Conway JS (2016) Weed development in spring wheat after contrasting soil tillage and nitrogen treatment. Ann Appl Biol 169:236–247. doi:10.1111/aab.12294
Richner N, Holderegger R, Linder HP, Walter T (2015) Reviewing change in the arable flora of Europe: a meta-analysis. Weed Res 55:1–13. doi:10.1111/wre.12123
Robinson RA, Sutherland WJ (2002) Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 39:157–176. doi:10.1046/j.1365-2664.2002.00695.x
Schumacher W (1980) Schutz und Erhaltung gefährdeter Ackerwildkräuter durch Integration von landwirtschaftlicher Nutzung und Naturschutz. Natur und Landschaft 55:447–453
Smith RM, Roy DB (2008) Revealing the foundations of biodiversity: the Database of British Insects and their Foodplants. Brit Wildl 20:17–25
Sotherton NW (1990) The environmental benefits of conservation headlands in cereal fields. Pestic Outlook 1:14–18
Spink JH, Semere T, Sparkes DL, Whaley JM, Foulkes MJ, Clare RW, Scott RK (2000) Effect of sowing date on the optimum plant density of winter wheat. Ann Appl Biol 137:179–188. doi:10.1111/j.1744-7348.2000.tb00049.x
Stace C (2010) New flora of the British Isles, 3rd edn. Cambridge University Press, Cambridge
Stewart KEJ, Bourn NAD, Thomas JA (2001) An evaluation of three quick methods commonly used to assess sward height in ecology. J Appl Ecol 38:1148–1154. doi:10.1046/j.1365-2664.2001.00658.x
Still K, Byfield A (2007) New priorities for arable plant conservation. Plantlife International, Salisbury. http://www.plantlife.org.uk/uploads/documents/New_Priorities_for_Arable_Plant_Conservation.pdf. Accessed 20 Apr 2016
Stoate C, Boatman ND, Borralho RJ, Rio Carvalho C, de Snoo GR, Eden P (2001) Ecological impacts of arable intensification in Europe. J Environ Manag 63:337–365. doi:10.1006/jema.2001.0473
Storkey J, Westbury DB (2007) Managing arable weeds for biodiversity. Pest Manag Sci 63:517–523. doi:10.1002/ps.1375
Storkey J, Moss SR, Cussans JW (2010) Using assembly theory to explain changes in a weed flora in response to agricultural intensification. Weed Sci 58:39–46. doi:10.1614/WS-09-096.1
Sutcliffe OL, Kay QON (2000) Changes in the arable flora of central southern England since the 1960s. Biol Conserv 93:1–8. doi:10.1016/S0006-3207(99)00119-6
Svensson R, Wigren M (1982) Några ogräsarters tillbakagång belyst genom konkurrens-, gödslings- och herbicidförsök. Sven Bot Tidskr 76:241–258
Walker KJ, Critchley CNR, Sherwood AJ, Large R, Nuttall P, Hulmes S, Rose R, Mountford JO (2007) The conservation of arable plants on cereal field margins: an assessment of new agri-environment scheme options in England, UK. Biol Conserv 136:260–270. doi:10.1016/j.biocon.2006.11.026
Wilson P (1994) Managing field margins for the conservation of the arable flora. In: Boatman N (ed) Field margins: integrating agriculture and conservation. BCPC Monograph No. 58. British Crop Production Council, Farnham, pp 253–258
Wilson PJ (1999) The effect of nitrogen on populations of rare arable plants in Britain. In: Boatman ND (ed) Field margins and buffer zones: ecology, management and policy. Aspects of applied biology, vol. 54. Association of Applied Biologists, Wellesbourne, pp 93–100
Wilson PJ (2010) The Roundwood Estate—rare arable plant survey. Unpublished report
Wilson JD, Morris AJ, Arroyo BE, Clark SC, Bradbury RB (1999) A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in northern Europe in relation to agricultural change. Agr Ecosyst Environ 75:13–30. doi:10.1016/S0167-8809(99)00064-X
Acknowledgments
This study was part of a project funded by Defra (BD5204). The field experiments were set up in cooperation with Charles Church Partnership and with Jon Harley, the farm manager of Roundwood Estate. Valuable advice was provided by Mike Green from Natural England. We would like to thank Marek Nowakowski (Wildlife Farming Company) for advice and help while setting up the experimental treatments, and Pete Nuttall for help with field recording. Two anonymous referees made valuable comments and suggestions during the review process.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Daniel Sanchez Mata.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Wagner, M., Bullock, J.M., Hulmes, L. et al. Cereal density and N-fertiliser effects on the flora and biodiversity value of arable headlands. Biodivers Conserv 26, 85–102 (2017). https://doi.org/10.1007/s10531-016-1225-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-016-1225-4