
Abstract
During almost a century of permanence in the Mediterranean, the warm water species Teredo bartschi has adapted to progressively colder climates up to overwintering at water temperatures only a few degrees above zero. A fine-grained analysis of discoveries, synonyms, museum collections and grey literature establishes that this species entered the Mediterranean since at least 1935. Coming from tropical waters through the Suez Canal, the species has undergone to a long period of acclimatization in the Levantine Basin of the Mediterranean and then pushed north at the beginning of this century until it has invaded the Lagoon of Venice. The invasion routes are reconstructed and presented. The lagoon of Venice is a microtidal bar-built estuary located in the northernmost part of the Mediterranean and represents the highest latitude reached by the species on a global scale. Here for over ten years, T. bartschi has now become invasive forming stable and abundant populations. This paper presents some biometrics of hard parts such as pallets and shells as well as the description of siphons, useful for the identification and characterization of the species. The shape of the pallets of the Venetian population differs from the Aqaba’s (Giordania) and Mersin’s (Turkey) populations. Phenotypic variation are probably due to environmental effects on morphology.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Shipworms are xylotrophic bivalves (Teredinidae Rafinesque 1815), strongly adapted to live inside and feed on wood. The wood is finely minced by the rocking, rasping movement of the valves (Miller 1924a, b) and its digestion is supported by the actions of bacterial endosymbionts (Turner 1966; Distel 2003; Betcher et al. 2012). Other widespread wood-boring bivalves belong to the families Pholadidae Lamarck 1809 and Xylophagidae Purchon 1941.
Wood is naturally present in the marine environment in many forms from living mangroves to floating and drifting logs, and wood that has sunk to the seafloor. Brought offshore by rivers and currents, driftwood may travel by sea for months or years covering thousands of miles (Edmonson 1962; Carlton et al. 2017; Treneman et al. 2018). Human presence overseas was suggested to Columbus by sailors who retrieved carved wood objects far from the coast (Phillips and Phillips 1993). Hence, floating wood is used as a vehicle for the spreading of species like shipworms (Thiel and Gutow 2005). Waterlogged wood finally sinks and reaches the seabed supplying new habitats for seafloor organisms both in coastal shallows and oceanic depths, where Xylophagaidae dominate (Laurent et al. 2009; Bienhold et al. 2013; Romano et al. 2014; Kintisch 2016). Maritime structures encompass: pilings and piers as permanent structures, timber logs and manufactured wood debris as drifting material, wrecks and archaeological sites as sunken waterlogged wood. During the early geographical explorations, the wooden hulls of ships provided ideal habitat for woodboring molluscs, and shipworms used them to travel from port to port and colonize the world (Columbus 1492–1493; Pigafetta 1524; Sellius 1733; Vallisneri 1733). Again, visiting the estuaries and lagoons of the Mosquito coast and Veragua during his fourth voyage to the West Indies (1502–1504), Columbus experienced a massive shipworm attack that forced him to abandon two of his caravels, the Gallega and the Vizcaìna (de Navarrete 1807). Wood vessels and maritime routes were perfect vectors for the diffusion of shipworms connecting ports, often located in estuaries (Carlton 1989).
The spread of shipworms continued even after the end of the era of galleons colonizing new seas all-over the world (Wolff 2005; Borges et al. 2014a; Shipway et al. 2014; Appelqvist et al. 2015a; Velásquez and López 2015) and consequently the geographic origin of many shipworm species is questionable.
The preferred way of dissemination of these sea travellers are undoubtedly the commercial routes (Hill and Kofoid 1927); so it is no wonder to witness their constant dissemination in the market globalization era that promotes the spread of a plethora of non-indigenous species (NIS) (Levine and D'Antonio 2003; Meyerson and Mooney 2007; Seebens et al. 2013; Turbelin et al. 2017). The opening of the Suez Canal (November 17th, 1869), connecting the Eastern Mediterranean with the Red Sea and the Indo-Pacific, gave a further impulse to the colonization of NIS (Por 1978; Por 1990; Leppäkoskii et al. 2002; Galil 2008; Rilov and Galil 2009; Katsanevakis et al. 2014; Galil et al. 2017; 2018).
The Mediterranean and the Red Sea have been connected by a canal in many historical periods: Persian (ca 500 BCE), Ptolemaic (ca 250 BCE), Roman (ca 100 CE) and Islamic (ca 650 CE). The waterway was made possible by a canal, called "Canal of the Pharaohs" or "Trajan's Canal", which connected the Pelusiac Branch of the Delta Nile across the Wadi Tumilat with the Bitter Lakes in turn connected with the Red Sea (Redmount 1995; Cooper 2009; Aubert 2015). There is no documentation as to whether the ancient Mediterranean—Red Sea connection allowed an exchange of species between the two seas, a possibility made difficult by the great diversity of the waters crossed. Mathematical models have shown that the Mediterranean is more readily colonized than the Red Sea by organisms crossing the Suez Canal as free-drifting propagules due to the direction and velocity of water currents (Agur and Sasfriel 1981) and here we could also include driftwood. Ballast water and hull fouling could also contribute significantly to the species exchange between the seas (Shefer et al. 2004). This waterway is considered the main responsible for half of the marine alien species introduction in the Mediterranean. In fact, most of the alien species have been found in the Eastern Mediterranean basin, where the Suez Canal flows, among them warm-water species account for 88.4% (Zenetos et al. 2012). Zenetos et al. (2012) listed 986 alien species in Eastern Mediterranean basin, 215 are molluscs, 120 of which are considered established. The shipworms Teredo bartschi Clapp 1923 and Teredothyra dominicensis Bartsch 1921 were recently added to the list (Borges et al. 2014b; Shipway et al. 2014).
The Lagoon of Venice is the main Italian hotspot of species introduction counting 71 NIS, out of which 55 are considered established. The list included 29 macroalgae, 13 molluscs (not including T. bartschi, 13 crustaceans, four annelids, four cnidarians, four tunicates, three bryozoans, and one pycnogonid (Marchini et al. 2015)).
Throughout history, shipworms have had a very strong impact on vessels and maritime infrastructures with huge economic losses (Rayes et al. 2015). The concern for the resurgence of severe wood attacks, reported in recent decades from many parts of the world (e.g. Al-Hamdani et al. 2011; Paalvast and Van der Velde 2011a; Shipway et al. 2014; Appelqvist et al. 2015b) was felt also in the Lagoon of Venice prompting scientific public institutions to resume the studies of Roch (Roch 1957a, b; Roch and Santhakumaran 1967). Our surveys led to the identification of Teredo bartschi, a non-indigenous shipworm never found before now in the Lagoon of Venice (Roch 1957a, b; Roch and Santhakumaran 1967) and which was foretold to expand in the Mediterranean (Munari 1974).
In this paper, we described the main characteristics of the Venetian population, bringing a review and a historical analysis of the expansion route of this species.
Materials and methods
Environmental context
The Lagoon of Venice is a bar-built estuary (Kjerfve 1994) with a surface area of about 550 km2, roughly 80% of which covered by water and about 10% by salt marshes. The mean depth is about 1.2 m, with only 5% of the lagoon deeper than 5 m (Molinaroli et al. 2009). The basin is connected to the Adriatic Sea through three inlets (Lido, Malamocco, Chioggia) which allow tidal flushing twice a day. Tides are the largest in the Mediterranean, nevertheless the lagoon can be classified as microtidal with extensive nanotidal areas (Tagliapietra and Volpi Ghiradini 2006). The mean tidal range is 61 cm (calculated on 1986–2004), with an estimated maximum of 121 cm. Most of the lagoon waters are classified as (mixo)euhaline/(mixo)polyhaline, according to the Venice System (Anonymous 1959).
The north-eastern coast of the Adriatic Sea, in the middle of which lays the Venice Lagoon, is characterized by a climate colder than the rest of the Adriatic and the Mediterranean. This is due to the presence of the Alps, the contribution of cold waters from the rivers of Alpine origin, the exposure to the north-east Bora winds and the distance from the circulation of the warm Mediterranean currents. Known as the "Venetian biogeographic gap", this climatic gap results in a reduction of thermophilic species and the presence of glacial relics such as the brown alga Fucus virsoides J Agardh, endemic of the North Adriatic Sea (Gulf of Venice), the only Mediterranean species of Fucaceae, a widespread family in Atlantic temperate waters (Marcello 1962; Tagliapietra et al. 2011).
The average salinity is about 30 PSU, presenting steep salinity gradients close to the bay-head river mouths. Water temperature follows strictly the air temperature, from over 30 °C during summer to a few degrees above zero in winter, with rare freezing events. On the whole, the lagoon can be considered as mesotrophic (Sfriso et al. 2019), even if eutrophic conditions could be reached around the City of Venice.
Currently, 50,000/100,000 wooden poles of different size are used for marking the large navigation canals, for mooring large and small boats, for the protection of saltmarshes, for fisheries, together with boats and maritime structures, including cultural heritage traditional structures (i.e. wooden water-doors (Tagliapietra et al. 2019)). The anthropogenic supply of wood acts as a replacement, or supplements to the natural river supply, generating high concentrations of the appropriate substrate over short distances, unintentionally restoring an environment suitable for the life of marine woodborers.
Collection sites and materials
The first specimens identified as Teredo bartschi were extracted on October 30th 2013, from Spruce panels (Picea abies (L) Karst) (3000 × 70 × 20 mm) collected at the ancient harbour of the Arsenal of Venice (45°26′13”N, 012°21′14"E). These panels have been hung on a floating pontoon, at a depth of one meter from May to October 2013.
After the detection, other wood samples stored in our repertory (represented in Fig. 1 and summarized in Table 1) revealed the presence of T. bartschi.

Sampling site map with the detail of Venice lagoon in Mediterranean
From 2014 till now, we have regularly collected T. bartschi at the Experimental Site (45°24′23.35”N, 12°19′27.15”E) and at the Arsenal.
We also investigated the cooling outlet of the Thermal Power plant of Venice at Fusina (45°25′26.10”N, 12°14′50.65"E) on the Naviglio (a branch of the Brenta river estuary) placing Spruce collectors and Scots pine standard panels and checking them periodically from January to October 2017. Samples were collected also from Oak dolphins at the Malamocco-Marghera Canal at the Naviglio bifurcation (45°25′4.71”N, 12°15′34.34”E, during summer 2016).
Hydroclimagraphs (Hedgpeth 1957) have been produced to describe temperature and salinity regimes (Fig. 2) of the sampling sites. Venice Water Authority kindly provided raw data.

Hedgepth’s hydroclimograms of the five multimetric field stations where Teredo bartschi was collected. Analemmatic lines join the average monthly temperature and salinity, variability bars show the standard deviations
Sample treatment
All materials were processed at the ISMAR Venice laboratories. Wood samples were dissected with the help of chisels, carving knives and scalpels. Considering the high shipworm density, not all the individuals were picked up. The early samples were preserved in denatured 70% ethylic alcohol, whereas from 2013, onwards specimens were stored in 95% ethylic alcohol to be suitable for further genetic analysis.
Shipworms were identified using the keys of Turner (Turner 1966; Turner and Johnson 1971) and compared with the papers of Cragg et al. (2009) and Borges et al. (2014b). Turner (1966) reported Teredo bartschi having a wide variability in palettes morphology that was taken into consideration during identification (Table 2).
Specimens were measured under the dissecting microscope or photographed and measured later using Free and Open source software such as ImageJ (Schneider et al. 2012) and Inkscape (Harrington et al. 2004–2005). Digital radiography was performed using an Ecoray Orange 1040 HF apparatus set at voltage: 50 kV, current intensity: 0.5 mA, exposure time 7 ms, with a distance between the X-ray generator and the flat panel detector of 70 cm. All images contained in this text were generated with Gimp (Kimball et al. 1997–2014) and Inkscape.
Biometrics were carried out both on integer shipworms and on pallets enucleated from the fragment of shipworms in which the shell was missing. A pallet is composed by a stalk and a flattened calcareous blade covered distally by a thin periostracum cap. The principal dimensions of these anatomical parts, namely the length of the blade (BL), length of the calcareous part of the blade (CBL), the maximum width of the blade (BW) and the length of the stalk (SL), were measured and their relationships and proportions derived (Fig. 3). The total length of the palette (PL = pallet length = BL + SL) is obtained adding the length of the blade (BL) to the length of the stalk (SL). CBL measurement can be useful when treating samples with worn pallets, in which the fragile periostracal cap is lost, as often happens in archaeological remains or in the analysis of woods with dead animals. A single blade for each individual was considered (n = 96 obtained from 17 samples of wood, taken in 8 sites in different years).

Pallet morphology. BW—blade width, BL—blade length, SL—stalk length, CBL—length of the calcareous part of the blade
We performed a simple morphometric analysis of the pallets, quantifying the relationships between the anatomical structural parts of the palette. Subsequently, we have performed similar measurements on the palettes images found in the scientific literature (Table 2). We extracted information of a large number of individuals from the graphs reported by Cragg et al. (2009) and collected at the Gulf of Aqaba (29°25′N 34° 55′ E), then we added information obtained from the pictures in Borges et al. (2014b) collected at Mersin (Turkey, 36°33′ N, 34°15′ E). Afterwards, we compared our data with the description of the new species given by Clapp (1923, Museum of Comparative Zoology, Tampa FL MCZ45301) and with the measures of the palette of Teredo aegytptia Roch 1935 (Port Said, Egypt) obtained from the original description (Roch and Moll 1935). To these, we added measures taken from the drawings of Turner (1966) of a set of species synonymized with T. bartschi, i.e., Teredo batilliformis Clapp 1923 (Bermuda), Teredo balatro Iredale 1932 (Australia), Teredo shawi Iredale 1932 (Australia), Teredo hiloensis Edmondson 1942 (Hawaii), Teredo grobbai Moll 1937 (Iraq). The shell diameter (SD) related to the enucleated palette was obtained by applying the allometric relationship between BW and SD calculated for the whole animals. The body size (BS or the body length from the mouth to the tip of the syphons), the maximum diameter of the shells (SD) and the width of the pallet blade (BW) were measured on integer animals (n = 74, preserved in ethylic alcohol, extracted from 10 wood samples taken in 6 sites, in different years). Gill chambers were inspected to determine the spawning mode (synchronous or sequential brooders) looking for different larval stages. The number of pediveligers was counted under a stereomicroscope in a subset of 20 shipworms, by opening the branchial chambers or by transparency when these were few and easily identifiable in small individuals. The maximum SD of fully developed pediveligers was measured on 10 random larvae for each adult.
Entry holes at the wood surface were counted in a sub-sample area (5 × 5 cm) of Pinus sylvestris panels in which the dominance of T. bartschi was ascertained.
A number of regression analyses were performed between couples of biometrics from different datasets (including dataset from literature), and linear, logarithmic and power models were estimated, and statistical significance calculated. In one case, to check if regression lines from different datasets are different from each other, an ANCOVA (type II test) was performed. Statistical analyses were performed using the R software environment for statistical computing, v. 3.6.1 (R Development Core Team 2019), available as free software under the GNU General Public License.
Results
After the first shipworm identified as Teredo bartschi in October 2013, we carried out a retrospective inspection of wood samples in our possession. All the materials, we examined, revealed the presence of T. bartschi often in very large numbers. The oldest woods, we were able to re-examine, dated back to April 2007 from a mooring pole of the Venice Sailing Marina along with samples of September 2007, berthing dolphins, from a public transport pontoon located along the Grand Canal. Actually, already in 2007, there had been reports of sudden crashes of large poles. The examined poles showed a massive infestation especially near the bottom. The samples in April also showed signs of dormancy.
Distribution and Abundance
Teredo bartschi was found both in the intertidal and in the subtidal down to about two meters below the mean sea level. Deeper waters have not yet been investigated for shipworms in the lagoon of Venice. In 2013, the first individuals identified as T. bartschi were small (6.5 mm long), carrying 37 pediveligers. Just few palettes were extracted from the 2007 softwood samples in association with Lyrodus pedicellatus (complex) (Quatrefages 1849) which was the dominant species. In 2009, T. bartschi was found in Scots pine panels associated with L. pedicellatus and Teredo navalis Linnaeus 1758 in variable percentage from 0 to 100%: T. bartschi (avg. 39.6% St.Dev. 30.7), L. pedicellatus (avg. 28.4% St.Dev. 25.8), T. navalis (avg. 32.0% St.Dev. 33.3) (20 wood samples, 128 individuals, 5 sites). It was also present in Scots Pine panels from 2010 ranging from 25 to 46.2%. The species composition found in Larch (Larix decidua Mill 1768) panels exposed at the Dogi’s Palace during summer-fall 2011 was dominated by T. bartschi (avg. 57.8% St.Dev. 21.4) followed by L. pedicellatus (avg. 40.0% St.Dev. 23.3) and just few T. navalis (avg. 2.2% St.Dev. 1.9) (3 wood samples, 90 individuals, 30 individuals randomly collected per sample). Samples collected at the Arsenal after being exposed during the summer 2016, hosted T. bartschi avg 53.7% St.Dev. 11.6, L. pedicellatus avg 39.6% St.Dev. 13.9, T. navalis avg 6.7% St.Dev. 11.5 (3 wood samples, 28 individuals). Standard Scot Pine test panels were placed at the discharge outlet of the cooling system of the thermoelectric power plant at Fusina from January 2017. They were not colonised during the winter, but the settlement starting from March led to the complete failure in October. Teredo bartschi was dominant 97.8% while L. pedicellatus was present with only 2.2% (3 wood samples, 90 individuals, 30 individuals randomly collected per sample). Table 3 reportes a synopsis of the species percentage encountered during the sampling.
In Fig. 2, the hydroclimagraphs shows the monthly salinity and temperature averages at the SAMANET permanent monitoring stations where T. bartschi was found (averaged over the decade 2009–2018, the standard deviation is shown in brackets). The minima and maxima values of temperature and salinity recorded in the considered decade are −1.87/34.25 °C and 0.01/38.62 PSU. The 5th-95th percentile range is 4.22/28.22 °C and 10.44/34.70 PSU. Located along the estuarine gradient, the station recorded the lowest water temperatures (Dese Estuary) and the highest salinity was recorded at the power plat water discharge output (Fusina) (Table 1).
During the exposure of the test panels at the Dese Estuary, (August–September 2009), very few shipworms were found and all of them were T. bartschi. During the winter, in the proximity of the power plant, the water was warm and the temperature more constant all around the year, while salinity was close to that of the middle reaches of the lagoon (Arsenal of Venice). On January 11th 2017, at noon, water temperature was 18.1 °C at the cooling outlet (sal. 31.5), whereas it was much lower in the rest of the lagoon e.g. 4.1 °C at the Arsenal (sal. 32.6 PSU). During the warmer months, the temperature differences decreased e.g. on April 13th at noon at the cooling outlet temperature was 24.6 °C (sal. 31.7 PSU) versus 17.9 °C (sal. 31.8 PSU) at the Arsenal.
Teredo bartschi proved to be also more resistant to hypo/anoxia than the other shipworm species present in the lagoon. In July 2013, there was an extensive anoxia around the City of Venice with massive die-off of fishes lasting for few days. The wood samples collected after the anoxia have shown the almost exclusive survival of T. bartschi.
In 2013, the wooden board collected in Taranto revealed the presence of a large number of T. bartschi, a sign of a viable population.
Main features
The typical pattern of the attack of Teredo bartschi is the result of a massive, almost synchronous settlement of larvae leading to a very high density of small-size adults. After the attack, little wood matrix remained among the calcareous tubes that were often twisted and coiled (Fig. 4). The general appearance of the superficial attack was similar to that reported by Clapp (1923 Plate 3/4) when describing the species. The entrance holes measure about 350 μm, and often present a calcareous sept between the palettes (Fig. 5). In highly infested softwood samples, after only one summer of exposure, we counted hole densities ranging from 36,000 (at Arsenal, 2017) to 400,000 (at Fusina, 2017) per square meter. High density led to the growth of small animals that build chaotic tunnels (Fig. 6). High mortality was common during winter.

Wood samples hosting Teredo bartschi, on the left: appearance of a massive settlement, on the right: detail of the entrance hole with the calcareous set

Detail of the multi-layer tube of a frictioned Teredo bartschi. The sample was collected from the eroded intertidal portion of an Oak pile

X-ray of Pinus sylvestris panels (standard EN275) deployed on January 2017 at the cooling outlet of the Thermoelectric Power plant at Fusina. a April 13th 2017, b September 15th 2017
Reproduction and settlement
Teredo bartschi was able to produce from less than ten to few hundred fully developed larvae. Large individuals with body lengths of 20 mm up to 80 mm carried hundreds (400–500) of pediveligers, but also small individuals long less than one centimetre (5–7 mm) were found to carry up to 30 larvae. It was often possible to observe their movement inside the mother, or even see them swimming when removed from the gill chamber. The smallest individual bearing larvae was found on December 12th 2013 at ISMAR docks. It was 3 mm long and carried in its branchial chamber nine actively moving pediveligers (Fig. 7). The water temperature was 4 °C.

Teredo bartschi with bearing synchronous larvae
Pediveliger inside the adults measured 275–325 μm in diameter, a little less than the size of the entrance hole. The larvae were always at the same developmental stage, confirming that T. bartschi is a synchronous larviparous brooder. Individuals carrying well-developed pediveligers were found throughout the year, and individuals with and without larvae were often found together.
The settlement of the larvae began in late spring (May–June, at water temperature over 20 °C) but it could be quite variable. For example, in 2019 until July, there was no X-ray appreciable settlement on panels deployed in February, even though the water temperature had risen above 20 °C in early June. Then, at the end of August a massive colonization was recorded (100% coverage sensu EN275 (1992), Fig. 6). Panels placed at the cooling outlet of the power station were not colonized during the winter (temperature over 18 °C) and the settlement started in March (temperature over 20 °C). However, in this last case, the few individuals visible on the X-ray were not extracted and identified.
Morphology
The body size ranged from 1.19 mm to 62.03 mm with Shell Diameter between 0.73 and 4.92 mm. Pallets closely matched the description given by Turner (Turner 1966; Turner and Johnson 1971), Cragg et al. (2009) and Borges et al. (2014b). The dimensional relationships of the pallets allowed comparison with the images reported in scientific literature to validate the taxonomic identification. Furthermore, the identification of the ratios between the width of the blade, the shell and the length of the body could be used to infer the dimensions of the entire animal even in the absence of the soft parts of the body, such as, for example, in archaeological finds. The fresh periostracal cap had U-shaped distal margin, extending into characteristic lateral "horns" (looking like the Greek letter “psi”). Distal margins were moderately wide open, especially in little specimens. The colour of the cap was amber to light-brown, getting darker as the animal grows. The calcareous portion of the blade, not reaching the periostracal margin, was visible through the cap. It was shaped into a rounded form often resembling the Mithridates Crater (Figs. 3, 8a).

Pallets of the most common shipworms from the lagoon of Venice. a Teredo bartschi, b Lyrodus pedicellatus, c Teredo navalis
Pallets
Regression models have been calculated between main biometrics (Table 4, Figs. 9, 10).

Linear relationships between blade width (BW) and the other anatomical parts of the pallet (SL—stalk length, CBL—length of the calcareous part of the blade, BL—blade length, PL—pallet length = SL + BL)

Allometric relationships between body size (BS, on the secondary axis, in red) blade width (BW) and shell diameter (SD)
All of them showed high statistical significance.
The SD/BW ratio was quite stable (Mean = 3.1 SD = 0.26).
The BW was the better-preserved character as to be used as a term of comparison with the other biometric data. The relationship between SD and BW was analysed by fitting linear, logarithmic and allometric models. The linear relationships also presented high R2 value (BW = 0.297 SD + 0.0644, R2 = 0.964), suggesting that the general shape of the palette does not change much during the somatic growth in the investigated dimensional range.
The allometric relationship between SD and BW proved to be the most suitable but also the logarithmic regression presented fitted well the data with a high R2 value as well (BW = 0.597 ln SD + 0.333, R2 = 0.946), allowing the comparison with the model produced by Cragg et al. (2009) (BW = 0.59 ln SD + 0.62, R2 = 0.85). The linear relationships also presented high R2 value (BW = 0.297 SD + 0.0644, R2 = 0.964), suggesting that the general shape of the pallets does not change much during the somatic growth, in the investigated dimensional range.
The dimensional range of pallets and shell diameters of T. bartschi from Aqaba (Jordan) and Mersin (Turkey) was generally comparable to that of T. bartschi from Venice (Italy).
Aqaba pallets are proportionally wider than the Venice pallets that, in turn, were larger than Mersin’s (Aqaba: PL = 2.16 BW – 0.025, R2 = 0.906, df = 58, p-value < 2.2 e−16; Venice PL = 2.72 BW – 0.219, R2 = 0.835, df = 95, p-value < 2.2 e−16; Mersin PL = 2.96 BW + 0.128, R2 = 0.693, df = 11, p-value = 0.000414). Regression lines fitted on Mersin, Aqaba and Venice datasets (Aqaba: PL = 2.16 BW – 0.025, R2 = 0.906, df = 58, p-value < 2.2 e−16; Venice PL = 2.72 BW – 0.219, R2 = 0.835, df = 95, p-value < 2.2 e−16; Mersin PL = 2.96 BW + 0.128, R2 = 0.693, df = 11, p-value = 0.000414) were compared to each other by ANCOVA, which showed statistical difference in terms of both intercept (p-value < 2.2 e−16) and slope (p-value = 0.006057). The larger the pallets, the more noticeable the differences.
The holotype of T. bartschi reported by Clapp (1923) had a more elongated blade than the Venetian T. bartschi. One of the three palettes of T. aegyptia reported by Roch and Moll (1935) fitted the series of Aqaba whereas the other two palettes of T. aegyptia fell perfectly within the dimensional range of the Lagoon of Venice. The other species synonymized by Turner (1966) were similar to the Venice specimens with the exception of T. batilliformis, which was close to Aqaba’s. T. shawi sided the most elongated individuals of Venice and its calcareous portion of the blade resembled those of many T. bartschi found in the Lagoon of Venice (those resembling the "Mithridates Crater"). On specimen of T. aegyptia fell in the common area between Aqaba and Venice, the second in the common area between Venice and Mersin whereas the third had very elongated palettes approaching the Clapp’s typus. T. aegyptia was reported by Roch and Moll (1935) from Port Said, Egypt (Fig. 11).

Tubes
The size and thickness of the tube depended on the size of the animal. In case of massive attack and small organisms, the tube was 2–3 mm wide on average, lined with a thin limestone layer 40–60 μm thick. The tunnels were produced during the warm season but then they were sealed in the terminal part, near the valves, by a calcareous dome-shaped diaphragm, indicating dormancy. The dome was often made up of few calcareous layers which are easy to be separated from each other. We observed the diaphragm was produced also by other species such as Lyrodus pedicellatus. The multi-layer tube-tip, showed in Fig. 5, was taken from the intertidal bioeroded belt of the oak dolphin post. The pallets were thick and looked as they were worn out possibly due to rubbing against an unknown object.
Siphons
The structure of the siphons and papillae could be a valid foothold in the identification of species morphologically similar. In small individuals, both siphons were transparent, while, in larger individuals, they had a particular colour pattern although variable in intensity.
The papillae are found inside the siphons and are placed in radial symmetry. Their arrangement seemed to be characteristic and it can help in the identification of animals still living in the wood. Indeed, Roch (1940) had already proposed an identification key for Mediterranean species using this character.
The structure of the adult shipworm's siphons is described below. The inhalant (incurrent) siphon carried a fringe of papillae around its entrance. The observed specimens had six short siphonal papillae. The inhalant siphon is dotted on its entire surface by reddish-brown spots that line up along its length to form dark stripes ending in the space between the papillae, it is, therefore, characterized by a banding of alternating light and dark stripes, in equal number to the papillae. The exhalant (excurrent) siphon was shorter than the inhalant one and had no papillae. It was dorsally bluish white (i.e. not pigmented), whereas, in the ventral portion facing the inhalant siphon, it was reddish-brown spotted. The density and colour intensity of the points forming the lines was variable (Fig. 12).

Siphons of an adult (SD 4 mm—a), of young specimen (SD 2 mm—b) and the cross-section of the inhalant syphon as reported by the dashed red line on image (c)
Discussion and conclusions
In the Lagoon of Venice, Teredo bartschi was found both subtidally and intertidally in microtidal (mixo)euhalyne and nanotidal (mixo)polyhaline environments. Teredo bartschi can be considered established, or naturalized, in the lagoon as it has produced free-living, self-maintaining and self-perpetuating populations over, at least, ten years from 2007 to 2020. Furthermore, these populations were vigorous and ubiquitous, being T. bartschi often the dominant shipworm species.
The oldest evidence of the presence of T. bartschi in the Lagoon of Venice, we were able to recover, dated back to April 2007. Found in April 2007 wood samples together with the presence of well-developed but empty calcareous tubes, the evidence of dormancy (tubes closed by the calcareous dome) supported the hypothesis of a settlement dating back to at least the previous year, 2006. Pallets were still present in some tubes. The observation of the samples collected before 2013 suggested a progressive increase in the proportion of T. bartschi. In 2007, the local navigation company, ACTV SpA, reported sudden crashes of large poles. In the same year, the University of Padua was commissioned to assess the quality and durability of wood samples from mooring poles subjected to shipworms attack in the Lagoon of Venice. The research concluded that the cause of the shorter duration of the wood did not depend on intrinsic characteristics of the material. It was suggested, instead, that modifications of the lagoon habitat (Solidoro et al. 2010) might have influenced the biology of the shipworms making them more abundant (Urso and Crivellaro 2008). The concern, cited in the report, about the reduced durability of wooden poles in the lagoon has appeared in the local press since 2004, and now, after the present study, the cause of the problem can be likely attributed to the arrival of T. bartschi.
In the investigated sites, T. bartschi displayed a distribution similar to Lyrodus pedicellatus. However, it was more tolerant of the low salinity of the estuary. The temperature and salinity ranges, recorded at the collection sites in the lagoon of Venice (Fig. 2), were wider than those reported in scientific literature, showing an unexpected tolerance towards low temperatures for a species considered of tropical origin (Turner 1966; Cragg et al. 2009; Borges et al. 2014b).
Teredo bartschi appeared to be able to reach sexual maturity very early, preserving vital larvae in the coldest months with water temperatures few degrees above the zero. Hoagland (1986) reported two temperatures particularly meaningful for the reproduction of T. bartschi, i.e., 16 °C was the temperature below which the pediveligers were not active, while 20 °C was the optimal temperature for the settlement. In the Lagoon of Venice, the passage between these two temperatures takes place during two to three weeks, generally from the end of April to the beginning of May. Although just after leaving their mother, the pediveligers were already competent, i.e. able to respond to cues that induces the settlement, they could have a planktonic phase of about two weeks to favour their dispersal (Hoagland 1986). Therefore, this dispersal would postpone the first settlement towards the end of May and the beginning of June. Conversely, the passage 20–16 °C takes place during two to three weeks, generally from the end of September to mid-October (see Fig. 2).
We observed T. bartschi populations growing rapidly during summer. In just two months, the shipworm was able to destroy completely a standard panel of P. sylvestris. The density influenced the somatic growth (Cragg et al. 2009) but T. bartschi was fertile even when they were very small. Teredo bartschi is a small shipworm, perhaps the smallest. The smallest sexually mature shipworm with pediveliger in the gill chamber found in Venice was 3 mm long (Fig. 7), the same size of the smallest T. batschi with larvae in the gill chamber found at Oyster Creek (NJ, USA), which measured 4 mm (Hoagland and Crocket 1978). Our observations confirmed that this species is, indeed, a synchronous larviparous brooder, i.e. all the larvae from a single individual mature at the same time and are released together, but not all the individuals of a population ripen together so that during the breeding season there were often multiple settlements. These characteristics produced particularly virulent attack, which in short time led to the destruction of maritime structures, such as mooring poles, dolphins, pontoons and boats. The findings of T. bartschi in the lagoon of Venice have shed new light on the plasticity of this species and on its ability to colonize new habitats.
Teredo bartschi has hitherto been referred to as a “warm water” species, ranging from tropical to warm temperate waters (Turner 1966; Turner and Johnson 1971; Carlton 1995; Cragg et al. 2009; Borges et al. 2014b) but it was also recorded at latitudes over ± 30.0 °C, considered the limit of the subtropical belt (Corlett 2013). In the northern hemisphere, it was found in Portugal (Olhão 37°01′ N, Borges et al. 2014b), in Turkey (Mersin 36° 34′ N, Borges et al. 2014b) and in Japan (at Maizuru 35°28′ N; MCZBASE Malacology 278105). In the southern hemisphere, it reached latitudes over 40° S in Patagonia (Argentina, Puerto Madryn 42°46′ S; MCZBASE Malacology 278062 and 170816), and in Tasmania (Triabunna, Spring Bay, 42°30′ S) under the synonym of Teredo fragilis Tate, 1888 (Turner 1966) (MCZBASE Malacology 357242, 357256). The lagoon of Venice was, nevertheless, the highest latitude at which T. bartschi has ever been found (45°30′ N).
Over the last decade, T. bartschi established in the lagoon of Venice viable populations proving not only to be able to survive overwintering at water temperatures few degrees above zero, but becoming invasive and exceeding in density the more abundant local species such as L. pedicellatus. The Venetian population of T. bartschi showed a remarkable tolerance also for salinity. This species was generally found in full-salinity waters (Turner 1966; Cragg et al. 2009) but has also been reported in brackish estuaries (22 PSU, Leonel et al. 2002). In the lagoon of Venice, it was found both in euhaline waters and estuarine traits with wide salinity fluctuations (Fig. 2), albeit in low density.
The presence of T. bartschi in the Mediterranean was certainly due to the opening of the Suez Canal in 1869. This appeared clear tracking-back the available records through a careful analysis of the scientific literature and museum records. T. bartschi was first collected in the Mediterranean no later than the early-mid 1930s. Borges et al. (2014b) believed their report to be the first record of established T. bartschi in the Mediterranean, but they overlooked earlier 1930s Egyptian records, as well as records in experimental wood panels in Israel of the 1960s. Not mentioned, for example, in Borges et al. (2014b) there was T. aegyptia, which was described from Port Said (Egypt) in the Mediterranean (Roch and Moll 1935) and is a synonym of T. bartschi (Turner 1966). Roch (1940) and others considered another possible synonym of T. aegyptia: Teredo elongata Quatrefages, 1849, described from the Indian Ocean, and thus entered into the consideration of early Mediterranean records under that name. Nevertheless, Turner (1966) and Munari (1978) considered T. elongata an unrecognizable species (nomen dubium), despite Rancurel's (1954) attempt to redescribe it. Tryon (1862) appeared to have reported T. elongata on the East Coast of Africa, where it could have thus been poised to enter the soon-to-be opened (1869) Suez Canal. Tryon's record of the species in "East coast of Africa", however, was based on a report from "côte ouest d'Afrique," as Tryon himself stated, and thus appeared to be a mistranslation; the record was in fact from Grand-Bassam, Ivory Coast, on the West Coast of Africa (Tryon 1862). In turn, this Ivory Coast record was based upon Teredo petitii Recluz, 1849, which Tryon misinterpreted as a synonym of T. elongata (J. T. Carlton personal communication), currently a synonym of Psiloteredo senegalensis (Blainville, 1824) (see Turner 1966).
Quatrefage's material is preserved in alcohol at the Muséum d’Histoire Naturelle de Paris (Rancurel 1954), and it would be of interest to determine if DNA could be extracted and then be sequenced. Until such time as the potential for archival genetic work could be explored, Tiller and Bavay's (1905) report of T. elongata from Port Said, collected between 1903 and 1905, should not be considered in our analysis, although it is not impossible that T. bartschi might have established in the Mediterranean by that time.
The history of T. bartschi in the Mediterranean is thus as follows:
-
Port Said, Egypt, pre-1935. Reported by Roch and Moll, 1935 (as T. aegyptia). Roch and Moll did not provide a collection date, but their paper was presented in May 1935, and thus the collections, which their extensive paper were based upon, should have been received by them prior to 1935. In the same paper, T. aegyptia was also reported from Ismailia, in the middle reaches of the Suez Canal.
-
Tel Aviv, Jaffa, Haifa, Israel, 1961–1964. Teredo bartschi was collected in wood panels in Israel by the W. F. Clapp laboratory; the specimens are on deposit in the Museum of Comparative Zoology—Harvard University (MCZBASE 2019, Cat. Number Malacology 169815, 170520, 170522, 170524, 170527, 170571, 170657, 170671, 170673, 356963, 356964, 356965, 356970, 356971, 356972, 356973, 357176, 357178, 357179, 357180). Borges et al. (2014a, b) acknowledged these Israel records, based on data published in the Global Biodiversity Information System (GBIF; https://www.gbif.org/species/2288844), but did not accept them, being uncertain as to whether the specimens were collected in driftwood or wooden test panels. However, according to the data in GBIF, Clapp's Israel records were from fixed wood panels.
-
In an annotate catalogue, Sabelli et al. (1990) mentioned T. bartschi as a Mediterranean species, without any reference to sites, dates or any other information, so did Coll et al. (2010).
-
No other findings were mentioned until 2002, when T. bartschi was recorded from the Kumkuyu Marina near Mersin (Turkey) by Borges et al. (2014b). The species was considered introduced and established in the site since it was found in two distinct records ten years apart (2002–2003 and 2012–2013).
-
The Venetian specimens of T. bartschi were identified for the first time in 2013 but the species has been recovered also from wood sampled preserved since 2007 [2006].
-
The presence of a vital and abundant population, probably long established, in the Taranto Lagoon in 2013, as intermediate latitude between Venice (Italy) and Mersin (Turkey) supported the idea of a progressive northward expansion (Fig. 13).
Fig. 13 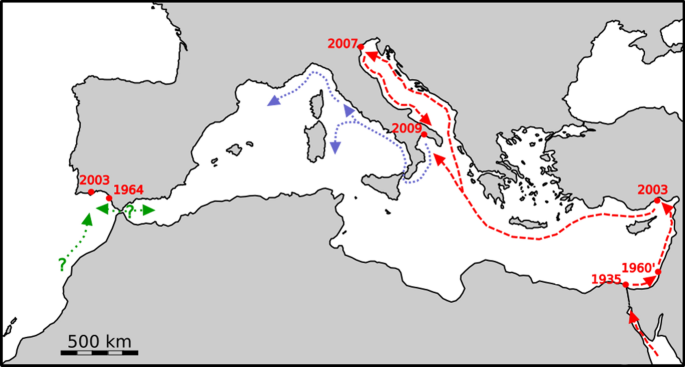
The spreading of Teredo bartschi in the Mediterranean (red arrows). Suez Canal, Egyptian coast (1935, Port Said), Israel (1960′), Turkey (2003), Venice (2007), Taranto (2009). The dates refer to the first records. Purple arrows show possible future routes following the main superficial currents. Green arrows indicate the putative routes to and from the Gulf of Cádiz

At local scale, colonization can be made possible by particular environmental conditions, both natural and anthropogenic, which in turn can promote dissemination on a larger scale. This led us to consider the hypothesis stated by Hoagland and Turner (1980) to explain the range extension in the eastern United States, involving the colonization of a power plant. In 1971, a breeding population of T. bartschi was found on wood structures at the heated effluent of the Nuclear Power Plant at Oyster Creek, NJ USA (Oyster Creek, Forked Creek; 39° 48′ N) and in the adjacent sites (Turner 1973). A few years later, in 1975, T. bartschi was found again at the Millstone Nuclear Power cooling plant (Connecticut 41°18′ N). Water temperatures at Millstone outlet were 12–14 °C above ambient, i.e., 0.6 °C in January–February to 25.5 °C in August–September (United States Nuclear Regulatory Commission 1984) resulting in a range from 12.5 °C to about 40 °C. The salinity ranged from 26 to 32 PSU. In 1981, the population was considered stable. In their hypothesis, the larvae could have been arrived in some way to the power plant, surviving and then breeding with a subsequent natural selection during the shut-down period of the power plants or when a few individuals from the original stock left the outlet (Hoagland and Turner 1980). The cooling water increased local water temperature to a level favourable for T. bartschi to settle and reproduce. The area, surrounding the warm outfall of power plant, thus acted as a stepping-stone for the colonization of a larger area. Teredo bartschi has not been found breeding in natural-temperature waters north of Cape Hatteras (Hoagland 1986).
Hence, we investigated the warm water discharge outlet of the Thermal Power plant of Venice at Fusina on the Naviglio (a branch of the Brenta river estuary). Panels placed at the cooling outlet were not colonized during January and February. Later, the massive colonization of T. bartschi occurred during the summer, leading to the complete failure of the collectors. January temperature of 18 °C suggested that settlement had happened at higher temperatures. Hoagland (1986) reported that larvae are found in the brood pouch throughout the year but that they are unable to move at temperatures below 16 °C. Our observations confirmed the presence of mature and viable larvae in the gill chamber of adults in periods with extremely low temperatures (4 °C). They were seen to be actively moving in the laboratory (about 20 °C), which means that they were ready for free life as soon as the water had reached an acceptable temperature. Bringing larvae in the gills chamber throughout the year implies being ready to release pediveligers as soon as the environmental conditions allow a successful settlement. This strategic trait would give the population the ability to expand by rafting over cold stretches of sea to meet favourable conditions. At Oyster Creek, New Jersey, shipworms stopped their activity when the water dropped below 13 °C (Hoagland and Crocket 1982a, b). In the Lagoon of Venice, this temperature is reached around mid-November.
At present, the range of temperatures and salinity of the Lagoon of Venice probably represents the "cold" extremes of the environmental requirements of T. bartschi in Europe. Here, the shipworms are subjected to very variable combinations of salinity and temperature, making the lagoon a particularly selective environment. A big reproductive effort with large number of offspring offers a source of genetic variation on which the environment can act by selecting strains able to stand to colder environments. Climate change might have contributed to the spread of this species, but a combination of factors is more likely the cause (Paalvast and Van der Velde 2011b; Borges et al. 2014b). Combining a wide tolerance to cold to the extension of the breeding season (Appelqvist and Havenhand 2016), the colonization of high latitude European coasts becomes the next probable future scenario.
Such a selection process could also have had repercussions on the morphology of the populations. The shape of the pallets, described by the relation between length and width, distinguished well the three populations of Aqaba, Mersin and Venice (Fig. 11). The population of Aqaba had proportionally wider palettes, followed by Venice and Mersin. Two pallets of T. aegyptia fell in the Venice range, while the third was decidedly elongated. The three pallets came from two different locations, Port Said on the Mediterranean coasts and Ismailia at the beginning of the Canal. This could explain the different proportions. The populations of Venice and Aqaba were well distinguishable by the relationships between the width of the pallets (BW) and the diameter of the shell, the SD of Aqaba being relatively smaller than Venice (Aqaba SD = 1.83BW1.15; Venice SD = 3.10BW1.03) (Fig. 14a). Both the typus of T. bartschi and the synonymous species overlapped the dimensions of the Venetian contingent. Teredo shawi was located between Venice and Aqaba. For T. aegyptia and for Mersin, it was not possible to obtain the size of the shell. On the contrary, the relations between PL and SD were similar (Fig. 14b). The data of Aqaba were largely overlapping with those of Venice and in line with both the types and Turner's iconography. Teredo batillifoirmis represented the only exception having a larger SD/PL ratio.

a Relationships between blade width (BW) and Shell Diameter (SD) b Relationships between blade width (PL) and Shell Diameter (SD) from Venice (Present work), Aqaba (Cragg et al. 2009), the Teredo bartschi typus (Clapp 1923), and synonymised species from literature (Turner 1966; Roch and Moll 1935) For T. aegyptia and for Mersin, it was not possible to obtain the size of the shell
The general idea is that the Venetian population differed, in the relationships between the skeletal parts, from the population of Aqaba, which had wider palettes and/or smaller shells, while it did not differ much from specimens coming from other sites. Since pallet width defined the diameter of the siphonal opening, Cragg et al. (2009) suggested that this opening tends to be larger in waters poor of plankton, allowing a larger water flux. This hypothesis implies a mixed xylophagous/planktivorous feeding as suggested for other species (Mann and Gallager 1985a, b; Paalvast and van der Velde 2013). This speculation fits with the trophism of the two aquatic environments: the Gulf of Aqaba, oligotrophic surrounded by desert and the lagoon of Venice, a mesotrophic estuary. Nevertheless, if this hypothesis is valid, the shape of the Mersin pallets would suggest waters with a trophism higher than those of the Venice lagoon that is almost eutrophic. In order to give a more decisive contribution to this issue, a comparative analysis on the plankton of these locations would be needed. It is also possible that we are dealing with multiple causes shaping the siphonal openings width including the presence of suspended materials in the water, the wood type or the size of the pediveliger larvae, not known from the other localities. Among the causes, the different genetic heritage of the founders of the colonies cannot be excluded. The mere geographical origin of the specimens obtained from scientific iconography is not sufficient to formulate any hypothesis.
The morphology and arrangement of the siphonal papillae and the chromatic pattern of the siphons may constitute diagnostic characters useful for the identification of the species (Roch 1940). The description of the siphons of the teredinids was absent in the literature, except for the work of Roch (1940), which, in turn, mentioned Sellius (1733) and Meyer and Möbius (1865) as unique precedents. We could not find any other description of the siphons and especially of the papillae, in literature. Clapp (in Turner 1966) stated that the siphons of T. bartschi have a colour similar to those of Bankia gouldi (Bartsch 1908) but added that the colouring of the siphons of B. gouldi is too variable to constitute a valid diagnostic character in the systematic identification.
According to Hoagland, T. bartschi showed a series of biological traits that make it adapted to the mangrove environment, where wood is highly available (Hoagland 1986; Hoagland and Crocket 1982a, b). The most important traits are the reproductive strategy and physiological plasticity of the larvae. Releasing larvae at the pediveliger stage, ready to settle, allows a high survival rate as long as there is a large amount of wood available in close proximity. Other adaptations to the mangrove environment are: a wide larval tolerance towards temperature and salinity; pediveliger with short free-life phase (Lane et al. 1954) (and therefore with spatial dispersion in the mid-range); tendency to settle in the wood when salinity goes down (Hoagland 1986). Teredo bartschi's pediveliger also prefers fresh, non-colonized adult wood (Hoagland 1986), a possible adaptation to settle in living mangrove trees. Such adaptations can be beneficial also in temperate estuaries. In the pristine state, in fact, wood is also abundant in estuaries and lagoons. Although not growing directly in brackish waters, wood is generally abundantly transported by rivers. Nowadays, humid and temperate riparian forests are largely reduced and watercourses are continually cleaned from trunks and branches so that the availability of natural wood is becoming very limited. However, the Lagoon of Venice lagoon is a special case. A large amount of wood of anthropogenic origin is in permanent contact with the water. Tidal currents facilitate the spread of larvae stimulated to settle down by the presence of fresh wood and saline gradients. Typically r-selected, these characteristics favour the dispersion of the species in estuaries spaced from stretches of unfavourable sea. Teredo bartschi would always be ready to colonize quickly the estuary with a fast and massive swarm of larvae as soon as the carrier/driftwood approaches it. Once the larvae are released, they can drop down, be passively transported or even move actively to get closer to the wood or choose a more suitable current. The active vertical movement of T. bartschi larvae can be as fast as 7.7 mm s−1 (Isham and Tiemey 1953; Turner and Johnson 1971; Mann and Wolff 1983). In their search for substrate, pediveligers would respond to chemical cues such as salinity lowering (Hoagland 1986) or molecules released from submerged wood and/or from conspecifics (Toth et al. 2015). After settlement, the next generation grows fast and repeats the cycle until the piece of wood leaves the estuary, for example after a flood. Therefore, estuaries would be used as stepping stones of the dispersion. The arrival in an estuary during a favourable period, followed by a recrudescence of environmental variables, such as, for example, a harsh winter, could represent a phase of selection that contributes to increase the breadth of the ecological niche by selecting more tolerant animals. This plasticity can give also an advantage to T. bartschi in overcoming difficult, hypersaline, oligotrophic and warm stretches of sea (Cragg et al. 2009).
The timing and geographical distribution of the findings support the hypothesis of a northward migration facilitated by the counter clockwise surface current that from Egypt heads towards the eastern shores of the Levantine sub basin touching Israel and Turkey, then crossing the Cretan-Aegean basins to spill in the Northern Ionian Basin. In front of the Otranto Strait, the current splits the left stream feeding the Gulf of Taranto and the right branch penetrating the Adriatic up to the Gulf of Venice then descends along the Italian coasts joining the left branch. Lessepsian species can be “trapped” for long time in the Levantine basin (Pascual et al. 2017), this could justify the lack of reports of T. bartschi (or one of its synonym) in the western Mediterranean basin. Nevertheless, T. bartschi could penetrate the Tyrrhenian Sea from the Gulf of Taranto through the Strait of Messina. Once in the western basin, its shorter residence times, compared to the Levantine basin (Tanhua et al. 2013), could allow a faster dispersal of the species. Driftwood follows the same fate of plastic floating litter. Studies on the transport of marine litter in the sea (Zambianchi et al. 2017), increased after its inclusion in the Marine Strategy Framework Directive (EC 2008), will contribute to the understanding of the spreading of species that use rafting for dispersion.
The doubt remains whether the populations found in the Gulf of Càdiz at Rota (Spain, 1964) and Olhao (Portugal, 2003) are the result of a migration from the Mediterranean or have an Atlantic origin, also considering the lack of records along the African Atlantic coasts. The surface currents entering the Mediterranean from Gibraltar could favour a secondary penetration of T. bartschi in the Western basin from the Atlantic.
This hardy species seems to take advantage from the opportunity offered by new environments. There are hypotheses and evidences that, during their journey, individuals are selected so they are able to overcome cold seas and to explode in lush populations. In the expansion within the Mediterranean, which has lasted about one hundred years, they have slipped into the cul de sac of the Gulf of Venice with its lagoons, perfect selection point and hub. The next possible question could be to investigate whether T. bartschi is now ready to colonize the Northern Europe.
References
Anonymous (1959) The venice system for the classification of marine waters according to salinity. Limnol Oceanogr 3:346–347
Agur Z, Safriel UN (1981) Why is the Mediterranean more readily colonized than the Red Sea, by organisms using the Suez Canal as a passageway? Oecologia 49:359–361. https://doi.org/10.1007/BF00347598
Al-Hamdani ZK, Appelqvist C, Björdal CG, Gregory D, Manders M (2011) Guidelines for predicting of decay by shipworms in the Baltic Sea. p 47
Appelqvist C, Havenhand JN (2016) A phenological shift in the time of recruitment of the shipworm, Teredo navalis L, mirrors marine climate change. Ecol Evol 6:3862–3870. https://doi.org/10.1002/ece3.2126
Appelqvist C, Havenhand JN, Toth GB (2015a) Distribution and abundance of teredinid recruits along the Swedish coast—are shipworms invading the Baltic Sea? J Mar Biol Assoc UK 95:783–790. https://doi.org/10.1017/S0025315414001830
Appelqvist C, Al-Hamdani ZK, Jonsson PR, Havenhand JN (2015b) Climate envelope modeling and dispersal simulations show little risk of range extension of the shipworm, Teredo navalis (L), in the Baltic Sea. PLoS ONE 10(3):e0119217. https://doi.org/10.1371/journal.pone.0119217
Aubert J-J (2015) Trajan’s canal: river navigation from the Nile to the Red Sea. In: De Romanis F, Maiuro M (eds) Across the Ocean: nine essays on Indo-Mediterranean trade. Brill Pubbl, Leiden/Boston, pp 33–42
Betcher MA, Fung JM, Han AW, O’Connor R, Seronay R, Concepcion GP et al (2012) Microbial distribution and abundance in the digestive system of five shipworm species (Bivalvia: Teredinidae). PLoS ONE 7(9):e45309. https://doi.org/10.1371/journal.pone.0045309
Bienhold C, Pop Ristova P, Wenzhöfer F, Dittmar T, Boetius A (2013) How deep-sea wood falls sustain chemosynthetic life. PLoS ONE 8(1):e53590. https://doi.org/10.1371/journal.pone.0053590
Borges LMS, Merckelbach LM, Sampaio I, Cragg SM (2014a) Diversity, environmental requirements, and biogeography of bivalve wood-borers (Teredinidae) in European coastal waters. Front Zool 11:13. https://doi.org/10.1186/1742-9994-11-13
Borges LMS, Sivrikaya H, Cragg S (2014b) First records of the warm water shipworm Teredo bartschi (Bivalvia, Teredinidae) in Mersin, southern Turkey and in Olhão. Portugal Bioinvasions Rec 3(1):25–28. https://doi.org/10.3391/bir.2014.3.1.04
Carlton JT (1989) Man’s role in changing the face of the ocean: biological invasions and implications for conservation of near-shore environments. Conserv Biol 3:265–273. https://doi.org/10.1111/j.1523-1739.1989.tb00086
Carlton JT (1995) Shipping Study: The Role of Shipping in the Introduction of Nonindigenous Aquatic Organisms to the Coastal Waters of the United States (other Than the Great Lakes) and an Analysis of Control Options. U.S. Coast Guard, Office of Engineering, Logistics, and Development, p 275
Carlton JT, Chapman JWC, Geller JB, Miller JA, Carlton DA, McCuller MI, Treneman N, Steves BP, Ruiz GM (2017) Tsunami-driven rafting: transoceanic species dispersal and implications for marine biogeography. Science 357(6358):1402–1406. https://doi.org/10.1126/science.aao1498
Clapp WF (1923) New species of Teredo from Florida. Proc Boston Soc Nat Hist 37:31–38
Coll M, Piroddi C, Steenbeek J, Kaschner K, Ben Rais Lasram F, Aguzzi J et al (2010) The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PLoS ONE 5(8):e11842. https://doi.org/10.1371/journal.pone.0011842
Columbus C (1492–1493) Journal of Christopher Columbus (During his First Voyage, 149293): and Documents Relating the Voyages of John Cabot and Gaspar Corte Real (Cambridge Library Collection - Hakluyt First Series) (Markham C (ed)). Cambridge: Camb U Pr. https://doi.org/10.1017/CBO9780511708411
Cooper JP (2009) Egypt’s Nile-Red Sea Canals: Chronology, Location, Seasonality and Function. In Blue LK, Cooper JP, Thomas R, Whitewright J (Eds.), Connected Hinterlands: Proceedings of Red Sea Project IV, Oxford: Archaeopress, pp 195–210
Corlett RT (2013) Where are the subtropics? Biotropica 45(3):273–275
Cragg SM, Jumel M-C, Al-Horani FA, Hendy IW (2009) The life history characteristics of the wood-boring bivalve Teredo bartschi are suited to the elevated salinity, oligotrophic circulation in the Gulf of Aqaba, Red Sea. J Exp Mar Biol Ecol 375:99–105. https://doi.org/10.1016/j.jembe.2009.05.014
Distel DL (2003) The biology of marine wood boring bivalves and their bacterial endosymbionts. In: Goodell B, Nicholas DD, Schultz TP, editors. Wood Deterioration and Preservation. Washington: Am Chem Soc Pr 253–271. https://doi.org/10.1021/bk-2003-0845.ch014
Edmonson CH (1962) Teredinidae, ocean travellers. Occasional Papers of the Bernice P Bishop Museum 23(3):45–59
EN275 (1992) Test Method for determining the protective effectiveness of a preservative against marine borers. European Committee for Standardization. p 21
European Commission (2008) Directive 2008/56/EC – Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 establishing a framework for Community action in the field of marine environmental policy (Marine Strategy Framework Directive)
Galil BS (2008) Alien species in the Mediterranean Sea—which, when, where, why? In: Davenport J et al. (eds) Chall Mar Ecosyst Hydrobiologia 606:105–116. https://doi.org/10.1007/s10750-008-9342-z
Galil B, Marchini A, Occhipinti-Ambrogi A, Ojaveer H (2017) The enlargement of the Suez Canal—Erythraean introductions and management challenges. Management of Biological Invasions 8:141–152
Galil BS, Marchini A, Occhipinti-Ambrogi A (2018) Mare Nostrum, Mare Quod Invaditur—The History of Bioinvasions in the Mediterranean Sea. In Histories of Bioinvasions in the Mediterranean, Springer, Cham pp 21–49
Geiger R, Pohl W (1953) Revision of Koppen–Geiger Climate Maps of the Earth. Darmstadt, Germany, Justus Perthes
Harrington B, the Inkscape developer team (2004–2005) Inkscape. Version 2, June (1991) Copyright (C) 1989, 1991 Free Software Foundation, Inc. 51 Franklin Street, Fifth Floor, Boston, MA 02110 – 1301, USA. http://www.inkscape.org/
Hedgpeth JW (1957) Classification of marine environments. In Hedgpeth JW Treatise on Marine Ecology and Paleoecology. Geol Soc Am Mem 67(1):17–27
Hill CL, Kofoid CA (eds) (1927) Marine Borers and Their Relation to Marine Construction on on the Pacific coast: Being the Final Report of the San Francisco Bay Marine Piling Committee. San Francisco, CA, p 357
Hoagland KE (1986) Effects of temperature, salinity and substratum on larvae of the shipworm Teredo bartschi Clapp and Teredo navalis Linnaeus (Bivalvia: Teredinidae). American Malacological Bulletin 4:89–99
Hoagland KE, Crocket L (1978) Analysis of populations of boring and fouling organisms he vicinity of the Oyster Creek Nuclear Generating Station. Quarterly Report September 1978-November 1978. U.S. Nuclear Regulatory Commission, NUREG/CR-0812, 66 pp
Hoagland KE, Crocket L (1982a) Ecological Studies of Wood-Boring Bivalves in the Vicinity of the Oyster Creek Nuclear Generating Station Progress Report June-August 1981 - February 1982, U.S. Nuclear Regulatory Commission, NUREG/CR-1939 Vol. 4, p 45
Hoagland KE, Crocket L (1982b) Ecological Studies of Wood-Boring Bivalves in the Vicinity of the Oyster Creek Nuclear Generating Station Progress Report December 1981 - February 1982, U.S. Nuclear Regulatory Commission, NUREG/CR-2727 Vol. 2, p 29
Hoagland KE, Turner RD (1980) Range extensions of teredinids (shipworms) and polychaetes in the vicinity of a temperate-zone nuclear generating station. Mar Biol 58(1):55–64. https://doi.org/10.1007/BF00386880
Isham LB, Tiemey JQ (1953) Some aspects of the larval development and metamorphosis of Teredo (Lyrodus) pedicellata de Quatrefages. Bull Mar Sci Gulf Caribb 2:574–589
Katsanevakis S, Wallentinus I, Zenetos A, Leppäkoski E, Çinar ME, Oztürk B, Grabowski M, Golani D, Cardoso AC (2014) Impacts of marine invasive alien species on ecosystem services and biodiversity: a pan-European review. Aquat Invasions 9:391–423. https://doi.org/10.3391/ai.2014.9.4.01
Kimball S, Mattis P & the GIMP developer team (1997–2014) GIMP 2.8.22. http://www.gimp.org/
Kintisch E (2016) Arctic shipworm discovery alarms archaeologists. Science 351(6276):901. https://doi.org/10.1126/science.351.6276.901
Kjerfve B (1994) Coastal lagoon processes. (ed) Elsevier Oceanography Series 60. Elsevier, Amsterdam
Lane CE, Tierney JQ, Hennacy RE (1954) The respiration of normal larvae of Teredo bartschi Clapp. Biolological Bulletin 106(3):323–327. https://doi.org/10.2307/1538766
Laurent MCZ, Gros O, Brulport JP, Gaill F, Le Bris N (2009) Sunken wood habitat for thiotrophic symbiosis in mangrove swamps. Mar Environ Res 67:83–88. https://doi.org/10.1016/j.marenvres.2008.11.006
Leppäkoskii E, Gollasch S, Olenin S (eds) (2002) Invasive Aquatic Species of Europe: Distribution, Impacts and Management. Kluwer Academic Publishers Dordrecht, The Netherlands, p 583. https://doi.org/10.1007/978-94-015-9956-6 ISBN: 1-4020-0837-6
Leonel RMV, Lopes S, Aversari M (2002) Distribution of wood-boring bivalves in the Mamanguape River estuary, Paraiba, Brazil. J Mar Biol Assoc UK 82:1039–1040
Levine JM, D’Antonio CM (2003) Forecasting biological invasions with increasing international trade. Conserv Biol 17:322–326. https://doi.org/10.1046/j.1523-1739.2003.02038.x
Mann R, Gallager SM (1985a) Physiological and biochemical energetics of larvae of Teredo navalis L. and Bankia gouldi (Bartsch) (Bivalvia: Teredinidae). J Exp Mar Biol Ecol 85(3):211–228
Mann R, Gallager SM (1985b) Growth, morphometry and biochemical composition of the wood boring molluscs Teredo navalis L., Bankia gouldi (Bartsch), and Nototeredo knoxi (Bartsch) (Bivalvia: Teredinidae). J Exp Mar Biol Ecol 85(3):229–251
Mann R, Wolf CC (1983) Swimming behaviour of larvae of the ocean quahog Arctica islandica in response to pressure and temperature. Mar Ecol Prog Ser 13:211–218
Marcello A (1962) Lacuna floristica del Veneziano e sue condizioni bioclimatiche. Memorie di Biogeografia Adriatica 5:53–118
Marchini A, Ferrario J, Sfriso A, Occhipinti-Ambrogi A (2015) Current status and trends of biological invasions in the Lagoon of Venice, a hotspot of marine NIS introductions in the Mediterranean Sea. Biological invasions 17(10):2943–2962
MCZBASE (2019) The Database of the Zoological Collections of the Museum of Comparative Zoology - Harvard University, https://mczbase.mcz.harvard.edu/ accessed on September 25th, 2019
Meyer HA, Möbius K (1865) Fauna der Kieler Bucht, Bd. I: Die Hinterkiemer Oder Opisthobranchia. Verlag Wilhelm Engelmann, Leipzig
Meyerson LA, Mooney HA (2007) Invasive Alien Species in an Era of Globalization. Front Ecol Environ 5(4):199–208. https://doi.org/10.1890/1540-9295(2007)5[199:IASIAE]2.0.CO;2
Miller RC (1924a) The boring habits of the shipworm. Sci Monthly 19(4):434–440
Miller RC (1924b) The boring mechanism of Teredo. Univ Calif Publ Zool 26(4):41–80
Molinaroli E, Guerzoni S, Sarretta A, Masiol M, Pistolato M (2009) Thirty-year changes (1970 to 2000) in bathymetry and sediment texture recorded in the Lagoon of Venice sub-basins, Italy. Mar Geol 258(1–4):115–125
Munari L (1974) Contributo alla conoscenza dei teredinidae nel mediterraneo. Conchiglie I Milano 10(34):71–84
Munari L (1978) Appunti critici sui pholadacea del Mediterraneo (Mollusca, Bivalvia). Soc Ven Sc Nat - Lavori 3:8–18
Navarrete M-F de (1807) Lettera di Cristoforo Colombo Vicerè ed Ammiraglio delle Indie Diretta ai Sovrani di spagna, Nostri Signori Cristianissimi e potentissimi, In: Marmocchi FC (1841) Raccolta di Viaggi dalla scoperta del nuovo continente a Dì Nostri. 2:107–146
Paalvast P, van der Velde G (2011a) Distribution, settlement, and growth of first-year individuals of the shipworm Teredo navalis L (Bivalvia: Teredinidae) in the Port of Rotterdam area, the Netherlands. Int Biodeter Biodegr 65(3):379–388. https://doi.org/10.1016/j.marpolbul.2011.05.009
Paalvast P, van der Velde G (2011b) New threats of an old enemy: the distribution of the shipworm Teredo navalis L. (Bivalvia: Teredinidae) related to climate change in the Port of Rotterdam area, the Netherlands. Mar Pollut Bull 62(8):1822–1829. https://doi.org/10.1016/j.marpolbul.2011.05.009
Paalvast P, van der Velde G (2013) What is the main food source of the shipworm (Teredo navalis)? A stable isotope approach. J Sea Res 80:58–60. https://doi.org/10.1016/j.seares.2013.03.003
Pallary P (1911) Catalogue des Mollusques du littoral méditerranéen de 1’Egypte, vol 7. Memoires de l'Institut Egyptien, p 207
Pascual M, Rives B, Schunter C, Macpherson E (2017) Impact of life history traits on gene flow: a multispecies systematic review across oceanographic barriers in the Mediterranean Sea. PLoS ONE 12(5):e017641. https://doi.org/10.1371/journal.pone.0176419
Phillips WD, Phillips CR (1993) The Worlds of Christopher Columbus. Camb U Pr. p 340
Pigafetta A (1524) The First voyage round the world, by Magellan by Pigafetta Antonio, ca. 1480/91-ca. 1534. Primo viaggio intorno al globo terraqueo. English; Stanley, Henry Edward John Stanley, Baron, pp 1827–1903
Por FD (1978) Lessepsian Migration – the influx of Red Sea Biota into the Mediterranean by way of the Suez Canal Ecol Stud. vol. 23, Springer, p 228
Por FD (1990) Lessepsian migration. An appraisal and new data. Bull Inst Oceanogr Monaco no special 7:1–10
Quatrefages De JLA (1849) Mémoire sur le genre Taret (Teredo Linn). Ann Sci Nat Zool, Sér 3, 11:19–73, pl. 1–2
R Development Core Team (2019) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://nam02.safelinks.protection.outlook.com/?url=http%3A%2F%2Fwww.r-project.org%2F&data=04%7C01%7Ckeppele%40si.edu%7C35c0c329c18c48c2a40308d8c95ab3f0%7C989b5e2a14e44efe93b78cdd5fc5d11c%7C0%7C0%7C637480738706983921%7CUnknown%7CTWFpbGZsb3d8eyJWIjoiMC4wLjAwMDAiLCJQIjoiV2luMzIiLCJBTiI6Ik1haWwiLCJXVCI6Mn0%3D%7C1000&sdata=Lh7k1WdrRsruh8HX6fiNr1eRokBeVk5f%2BKXwZUE4V8c%3D&reserved=0
Rancurel P (1954) Teredo elongata Quatrefages: redescription du type des collections du Muséum d’Histoire Naturelle de Paris. Bulletin de l’IFAN. Série A: Sci Naturelles 16(2):455–465. http://www.documentation.ird.fr/hor/fdi:010008978
Rayes CA, Beattie J, Duggan IC (2015) Boring through history: an environmental history of the extent, impact and management of marine woodborers in a global and local context, 500 BCE to 1930s CE. Environ Hist 21(4):477–512. https://doi.org/10.3197/096734015X14414683716163
Redmount CA (1995) The Wadi Tumilat and the ‘Canal of the Pharaohs’. J Near Eastern Stud 54:127–135. http://www.jstor.com/stable/545471
Rilov G, Galil B (2009) Marine Bioinvasions in the Mediterranean Sea – History, Distribution and Ecology. In Rilov G, Crooks JA (Eds) Biological Invasions in Marine Ecosystems– Berlin, Heidelberg: 549–575. https://doi.org/10.1007/978-3-540-79236-9_31
Roch F (1957a) Zur Ökologie der Terediniden, holzbohrenden Crustaceen und Balaniden in der Lagune von Venedig. (Vorläufige Mitteilung.) Anz math-nat Kl Öst Akad Wiss. pp 223–227
Roch F (1957b) Le Teredinidi della Laguna Veneta e dipendenza dalle condizioni idrografiche locali per quanto riguarda la loro distribuzione geografica. Boll Mus Civ St Nat Venezia 10:87–111
Roch F (1940) Die Terediniden des Mittelmeeres. Thalassia 4(3):147, 8 tavv
Roch F, Moll F (1935) Über einige neue Teredinidenarten. Sitzber Akad Wiss Wien Math-Naturwiss Kl Biol Mineral Erdk Abt 1 144(5–6):263–279. https://www.zobodat.at/pdf/SBAWW_144_0263-0279.pdf
Roch F, Santhakumaran LN (1967) Notes on the Teredinidae from the Lagoon of Venice (Italy). Boll Pesca Piscic Idrobiol Roma 22(1):37–48
Romano C, Voight JR, Pérez-Portela R, Martin D (2014) Morphological and genetic diversity of the wood-boring xylophaga (Mollusca, Bivalvia): new species and records from deep-sea Iberian Canyons. PLoS ONE 9(7):e102887. https://doi.org/10.1371/journal.pone.0102887
Sabelli B, Giannuzzi-Savelli R, Bedulli D (1990) Catalogo annotato dei molluschi marini del Mediterraneo. Bologna. Libreria Naturalistica Bolognese, p 348
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9(7):671–675. https://doi.org/10.1038/nmeth.2089
Seebens H, Gastner M, Blasius B (2013) The risk of marine bioinvasion caused by global shipping. Ecol Lett 16(6):782–790. https://doi.org/10.1111/ele.12111
Sellius G (1733) Historia naturalis teredinis seu xylophagi marini, tubulo-conchoidis speciatim belgici: cum tabulis ad vivum coloratis. Apud Hermannum Besseling: Trajecti ad Rhenum (Utrecht) Netherlands 30(11):1–353, 4 pl
Sfriso A, Buosi A, Mistri M, Munari C, Franzoi P, Sfriso AA (2019) Long-term changes of the trophic status in transitional ecosystems of the northern Adriatic Sea, key parameters and future expectations: the lagoon of Venice as a study case. Nat Conserv 34:193–215. https://doi.org/10.3897/natureconservation.34.30473
Shefer S, Abelson A, Mokady O, Geffen E (2004) Red to Mediterranean Sea bioinvasion: natural drift through the Suez Canal, or anthropogenic transport? Mol Ecol 13(8):2333–2343. https://doi.org/10.1111/j.1365-294X.2004.02227.x
Shipway RJ, Borges LMS, Müller J, Cragg SM (2014) The broadcast spawning Caribbean shipworm, Teredothyra dominicensis (Bivalvia, Teredinidae), has invaded and become established in the eastern Mediterranean Sea. Biol Invasions 16:2037. https://doi.org/10.1007/s10530-014-0646-9
Solidoro C, Bandelj V, Bernardi Aubry F, Camatti E, Ciavatta S, Cossarini G, Facca C, Franzoi P, Libralato S, Melaku Canu D, Pastres R, Pranovi F, Raicevich S, Socal G, Sfriso A, Sigovini M, Tagliapietra D, Torricelli P (2010) Response of Venice Lagoon ecosystem to natural and anthropogenic pressures over the last 50 years (ch 19) In: Kennish MJ, Paerl HW (eds.). Coastal Lagoons. Critical Habitats of Environmental Change, CRC Press: 483–511
Tagliapietra D, Aloui-Bejaoui N, Bellafiore D, De Wit R, Ferrarin C, Gamito S, Lasserre P, Magni P, Mistri M, Pérez-Ruzafa A, Pranovi F, Reizopoulou S, Rilov G, Solidoro C, Tunberg B, Valiela I, Viaroli P (2011) The Ecological Implications of Climate Change on the Lagoon of Venice, Unesco Report n. 2 Scientific Workshop 26–27 May 2011, UNESCO Venice Office, Venice, Italy, p 45 http://www.unesco.org/new/fileadmin/MULTIMEDIA/FIELD/Venice/pdf/report%202 %20final.pdf
Tagliapietra D, Sigovini M, Keppel E, Guarneri I, Palanti S, Veronese N, Abbate A (2019) Bioerosion effects of Sea Level Rise on the Doge’s Palace water doors in Venice (Italy). Facies 65:34. https://doi.org/10.1007/s10347-019-0577-0
Tagliapietra D, Volpi Ghirardini A (2006) Notes on coastal lagoon typology in the light of the EU Water Framework Directive: Italy as a case study. Aquat Conserv 16:457–467. https://onlinelibrary.wiley.com/doi/abs/10.1002/aqc.768
Tanhua T, Hainbucher D, Schroeder K, Cardin V, Álvarez M, Civitarese G (2013) The Mediterranean Sea system: a review and an introduction to the special issue. Ocean Sci 9:789–803. https://doi.org/10.5194/os-9-789-2013
Thiel M, Gutow L (2005) The ecology of rafting in the marine environment. I. The floating substrata. Oceanogr Mar Biol 42:181–264. https://doi.org/10.1201/9780203507810.ch6
Tiller L, Bavay A (1905) Les mollusques testacés du Canal de Suez. B Soc Zool Fr 30:170–181
Toth GB, Larsson AI, Jonsson PR, Appelqvist C (2015) Natural populations of shipworm larvae are attracted to wood by waterborne chemical cues. PloS ONE 10(5):e0124950. https://doi.org/10.1371/journal.pone.0124950
Treneman NC, Carlton JT, Borges LMS, Shipway JR, Raupach MJ, Altermark B (2018) Species diversity and abundance of shipworms (Mollusca: Bivalvia: Teredinidae) in woody marine debris generated by the Great East Japan Earthquake and Tsunami of 2011. Aquat Invasions 13:87–100. https://doi.org/10.3391/ai.2018.13.1.07
Tryon GW (1862) Monograph of the family Teredidae. Proc Acad Nat Sci Philadelphia 14:453–482
Turbelin AJ, Malamud BD, Francis RA (2017) Mapping the global state of invasive alien species: patterns of invasion and policy responses. Global Ecol Biogeogr 26(1):78–92. https://doi.org/10.1111/geb.12517
Turner RD (1966) A survey and illustrated catalogue of the Teredinidae (Mollusca: Bivalvia) The Museum of Comparative Biology Harvard University: Cambridge, MA. p 265
Turner RD (1973) First report on marine borers in Oyster Creek, Waretown, New Jersey. Museum of Comparative Zoology, Harvard University. p 30
Turner RD, Johnson AC (1971) Biology of marine wood boring molluscs. In: Jones EBG, Eltringham SK (eds) Marine borers, fungi and fouling organisms of wood. Organization for Economic Co-operation and Development, Paris, pp 259–301
Urso T, Crivellaro A (2008) Relazione tecnica relativa alle indagini sulla qualità e il degrado del legno di palificazioni per ormeggi nella Laguna di Venezia, Dipartimento Territorio e Sistemi Agro Forestali Università di Padova, Legnaro (PD), Maggio 2008, p 17
United States Nuclear Regulatory Commission (1984) Final environmental statement Related to the operation of Millstone Nuclear Power Station Unit 3, Northeast Nuclear Energy Company, Docket No. 50–423, p 67
Vallisneri A (1733) Osservazioni utilissime intorno alle Brume delle Navi. Op Fis Med Venezia 2:53–57
Velásquez M, López IM (2015) First record of Spathoteredo spatha (Mollusca: Teredinidae) in Venezuela. Rev Mex Biodivers 86(1):262–264. https://doi.org/10.7550/rmb.46308
Wolff WJ (2005) Non-indigenous marine and estuarine species in the Netherlands. Zool Med Leiden 79(1):3–116
Zambianchi E, Trani M, Falco P (2017) Lagrangian transport of marine litter in the Mediterranean Sea. Front Environ Sci 5:5. https://doi.org/10.3389/fenvs.2017.00005
Zenetos Α, Gofas S, Morri C, Rosso A, Violanti D, Raso JE et al (2012) Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Medit Mar Sci 13(2):328–352. https://doi.org/10.12681/mms.327
Acknowledgments
Many thanks are due to the editor James T. Carlton (Williams College, MA USA) for his careful, accurate and punctual revision of historical sources, his highly appreciated scientific suggestions and stylistic notes. We are grateful to the reviewers, Nancy C. Treneman, (Oregon Institute of Marine Biology, USA), and a second anonymous colleague, for their careful reading of our manuscript and their valuable comments and suggestions. Thanks to Dr. Luca Perale for performing radiographs at his Vet Lab in Venice and to Mr Loris Dametto (CNR-ISMAR) for its technical help in fieldwork. We thank the Venice Water Authority (Provveditorato Interregionale per le Opere Pubbliche per il Veneto, Trentino Alto Adige e Friuli Venezia Giulia, former Magistrato alle Acque) which kindly provided the SAMANET hydroclimatic data. Thanks are due to Dr. Patrizio Terlicher, Mr. Ferro and to the staff of the Territorial Office for Biodiversity of the State Forestry Corps, Tarvisio (UD, Italy), for the courtesy with which they provided us with the test panels. We thank Dr. Reuben Shipway (University of Portsmouth, UK) who confirmed our taxonomic identifications by DNAbarcoding. Finally, we thank the Italian network Long-Term-Ecological-Research, of which the Venice lagoon is a macrosite (LTEREUIT016) for useful suggestions in the discussion of data.
Funding
This research has been supported by the Italian Flagship Project RITMARE (Ricerca ITaliana per il MARE- The Italian Research for the Sea), coordinated by the Italian National Research Council and funded by the Italian Ministry of Education, University and Research.
Author information
Authors and Affiliations
Contributions
DT has performed the field and laboratory work, has analysed the data and written the first draft of the manuscript and edited the final version. IG has contributed to field and laboratory work, analysed the data, helped producing the figures, manipulates the images and has edited the manuscript. EK has contributed to field, laboratory work, and has edited the manuscript. MS has contributed to field and laboratory work and helped producing the figures. All authors have been involved in project development and have approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Tagliapietra, D., Guarneri, I., Keppel, E. et al. After a century in the Mediterranean, the warm-water shipworm Teredo bartschi invades the Lagoon of Venice (Italy), overwintering a few degrees above zero. Biol Invasions 23, 1595–1618 (2021). https://doi.org/10.1007/s10530-021-02461-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-021-02461-3